
Human RAD51 Protein Forms Amyloid-like Aggregates In Vitro

 , , , , , , , and
, , , , , , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. RAD51 Protein Forms Aggregates In Vitro That Are Resistant to SDS
2.2. RAD51 Binds to Thioflavin T during Aggregation
2.3. RAD51 Aggregates Bind to Congo Red and Glow under Polarized Light
2.4. RAD51 Aggregates Resistant to 1% SDS Have a Fibrillar Structure Visible under an Electron Microscope
2.5. X-ray Diffraction of RAD51 Fibrils Shows a Typical Cross-β Diffraction Pattern Typical for Amyloids
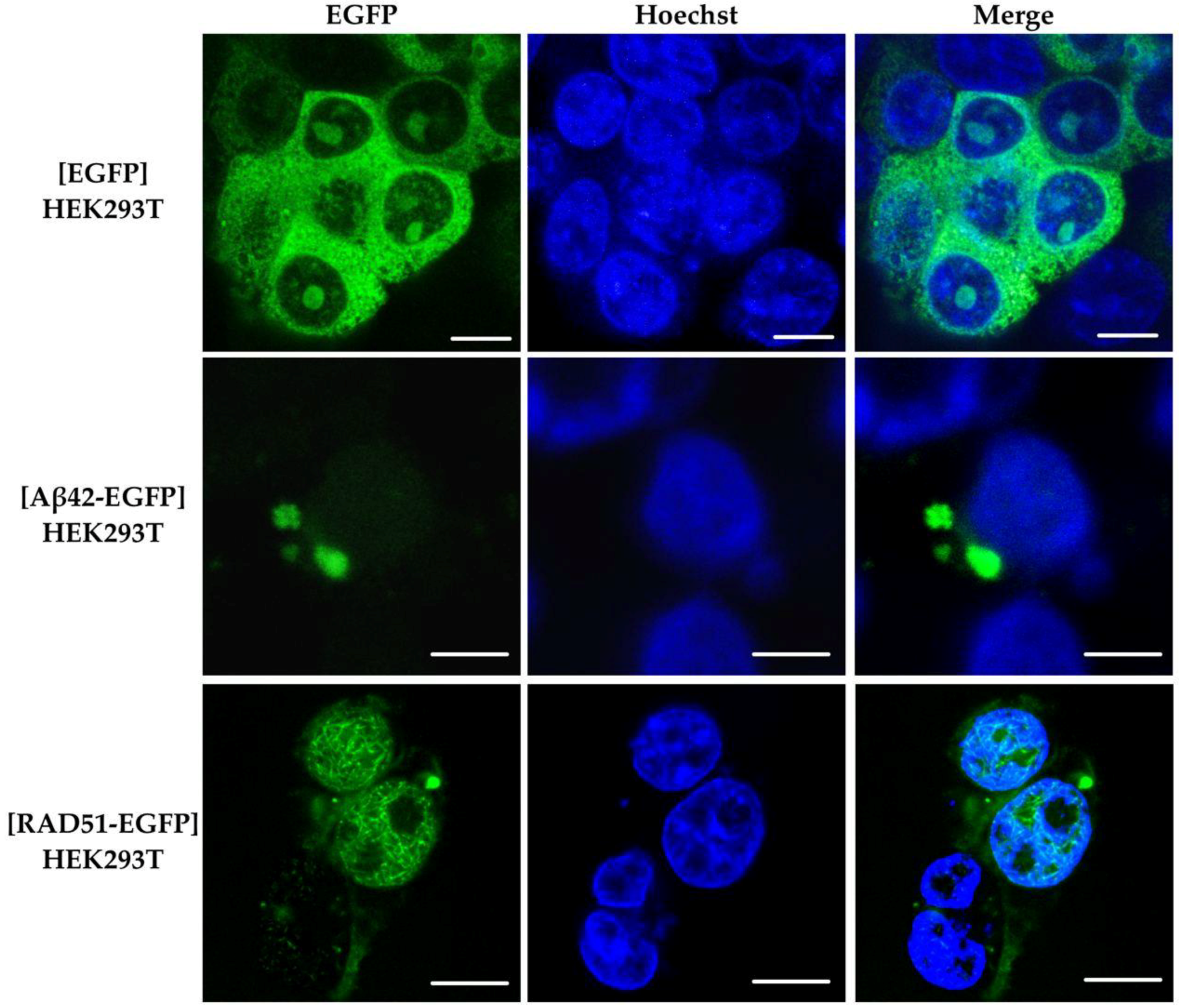
2.6. Aggregation of RAD51 in HEK293T Cells
2.7. Bioinformatic Analysis of RAD51’s Amyloidogenic Potential
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Cultivating Conditions
4.2. Cell Cultures
4.3. Plasmids
4.4. Mammalian Cell Transfection
4.5. Immunofluorescence
4.6. In Vitro Protein Expression and Purification
4.7. In Vitro Protein Aggregation
4.8. RAD51 Aggregation Dynamics
4.9. Microscopy Assays (TEM, Polarized, Confocal)
4.10. RAD51 X-ray Diffraction Analysis
4.11. RAD51 Aggregation Analysis in the C-DAG System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benson, M.D.; Buxbaum, J.N.; Eisenberg, D.S.; Merlini, G.; Saraiva, M.J.M.; Sekijima, Y.; Sipe, J.D.; Westermark, P. Amyloid Nomenclature 2020: Update and Recommendations by the International Society of Amyloidosis (ISA) Nomenclature Committee. Amyloid 2020, 27, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Chatani, E.; Yuzu, K.; Ohhashi, Y.; Goto, Y. Current Understanding of the Structure, Stability and Dynamic Properties of Amyloid Fibrils. Int. J. Mol. Sci. 2021, 22, 4349. [Google Scholar] [CrossRef] [PubMed]
- Maskevich, A.A.; Stsiapura, V.I.; Kuzmitsky, V.A.; Kuznetsova, I.M.; Povarova, O.I.; Uversky, V.N.; Turoverov, K.K. Spectral Properties of Thioflavin T in Solvents with Different Dielectric Properties and in a Fibril-Incorporated Form. J. Proteome Res. 2007, 6, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Frieg, B.; Gremer, L.; Heise, H.; Willbold, D.; Gohlke, H. Binding Modes of Thioflavin T and Congo Red to the Fibril Structure of Amyloid-β(1–42). Chem. Commun. 2020, 56, 7589–7592. [Google Scholar] [CrossRef]
- Bagriantsev, S.N.; Kushnirov, V.V.; Liebman, S.W. Analysis of Amyloid Aggregates Using Agarose Gel Electrophoresis. Methods Enzymol. 2006, 412, 33–48. [Google Scholar] [CrossRef]
- Kushnirov, V.V.; Alexandrov, I.M.; Mitkevich, O.V.; Shkundina, I.S.; Ter-Avanesyan, M.D. Purification and Analysis of Prion and Amyloid Aggregates. Methods 2006, 39, 50–55. [Google Scholar] [CrossRef]
- Howie, A.J. Origins of a Pervasive, Erroneous Idea: The “Green Birefringence” of Congo Red-stained Amyloid. Int. J. Exp. Pathol. 2019, 100, 208–221. [Google Scholar] [CrossRef] [Green Version]
- Yakupova, E.I.; Bobyleva, L.G.; Vikhlyantsev, I.M.; Bobylev, A.G. Congo Red and Amyloids: History and Relationship. Biosci. Rep. 2019, 39, 20181415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, K.L.; Serpell, L.C. X-ray Fibre Diffraction Studies of Amyloid Fibrils. Methods Mol. Biol. 2012, 849, 121–135. [Google Scholar] [CrossRef]
- Vaquer-Alicea, J.; Diamond, M.I. Propagation of Protein Aggregation in Neurodegenerative Diseases. Annu. Rev. Biochem. 2019, 88, 785–810. [Google Scholar] [CrossRef]
- Santos, T.G.; Lopes, M.H.; Martins, V.R. Targeting Prion Protein Interactions in Cancer. Prion 2015, 9, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Tahaney, W.M.; Mazumdar, A.; Savage, M.I.; Brown, P.H. Molecularly Targeted Therapies for P53-Mutant Cancers. Cell. Mol. Life Sci. 2017, 74, 4171–4187. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Zhao, Y.; Xu, Y.; Zheng, M.; Feng, Z.; Hu, W. Mutant P53 in Cancer: Accumulation, Gain-of-Function, and Therapy. J. Mol. Biol. 2017, 429, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Salot, S.; Sengupta, S.; Navalkar, A.; Ghosh, D.; Jacob, R.; Das, S.; Kumar, R.; Jha, N.N.; Sahay, S.; et al. P53 Amyloid Formation Leading to Its Loss of Function: Implications in Cancer Pathogenesis. Cell Death Differ. 2017, 24, 1784–1798. [Google Scholar] [CrossRef] [Green Version]
- Torreira, E.; Moreno-Del Álamo, M.; Fuentes-Perez, M.E.; Fernández, C.; Martín-Benito, J.; Moreno-Herrero, F.; Giraldo, R.; Llorca, O. Amyloidogenesis of Bacterial Prionoid RepA-WH1 Recapitulates Dimer to Monomer Transitions of RepA in DNA Replication Initiation. Structure 2015, 23, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.L.; Che, M.X.; Zhao, J.; Zhou, C.J.; Xie, M.Y.; Li, H.Y.; He, J.H.; Hu, H.Y. Structural Transformation of the Amyloidogenic Core Region of TDP-43 Protein Initiates Its Aggregation and Cytoplasmic Inclusion. J. Biol. Chem. 2013, 288, 19614–19624. [Google Scholar] [CrossRef] [Green Version]
- Smaldone, G.; Caruso, D.; Sandomenico, A.; Iaccarino, E.; Focà, A.; Ruggiero, A.; Ruvo, M.; Vitagliano, L. Members of the GADD45 Protein Family Show Distinct Propensities to Form Toxic Amyloid-like Aggregates in Physiological Conditions. Int. J. Mol. Sci. 2021, 22, 10700. [Google Scholar] [CrossRef]
- Carver, A.; Zhang, X. Rad51 Filament Dynamics and Its Antagonistic Modulators. Semin. Cell Dev. Biol. 2021, 113, 3–13. [Google Scholar] [CrossRef]
- Bonilla, B.; Hengel, S.R.; Grundy, M.K.; Bernstein, K.A. RAD51 Gene Family Structure and Function. Annu. Rev. Genet. 2020, 54, 25–46. [Google Scholar] [CrossRef]
- Liu, L.; Malkova, A. Break-Induced Replication: Unraveling Each Step. Trends Genet. 2022, 38, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Yu, X.; Shinohara, A.; Egelman, E.H. Similarity of the Yeast RAD51 Filament to the Bacterial RecA Filament. Science 1993, 259, 1896–1899. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jacobs, S.A.; West, S.C.; Ogawa, T.; Egelman, E.H. Domain Structure and Dynamics in the Helical Filaments Formed by RecA and Rad51 on DNA. Proc. Natl. Acad. Sci. USA 2001, 98, 8419–8424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haaf, T.; Golub, E.I.; Reddy, G.; Radding, C.M.; Ward, D.C. Nuclear Foci of Mammalian Rad51 Recombination Protein in Somatic Cells after DNA Damage and Its Localization in Synaptonemal Complexes. Proc. Natl. Acad. Sci. USA 1995, 92, 2298–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scully, R.; Chen, J.; Plug, A.; Xiao, Y.; Weaver, D.; Feunteun, J.; Ashley, T.; Livingston, D.M. Association of BRCA1 with Rad51 in Mitotic and Meiotic Cells. Cell 1997, 88, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, L.; Yu, D.S.; Lo, T.; Anand, S.; Lee, M.; Blundell, T.L.; Venkitaraman, A.R. Insights into DNA Recombination from the Structure of a RAD51-BRCA2 Complex. Nature 2002, 420, 287–293. [Google Scholar] [CrossRef]
- Morati, F.; Modesti, M. Insights into the Control of RAD51 Nucleoprotein Filament Dynamics from Single-Molecule Studies. Curr. Opin. Genet. Dev. 2021, 71, 182–187. [Google Scholar] [CrossRef]
- Kryndushkin, D.S.; Alexandrov, I.M.; Ter-Avanesyan, M.D.; Kushnirov, V.V. Yeast [PSI+] Prion Aggregates Are Formed by Small Sup35 Polymers Fragmented by Hsp104. J. Biol. Chem. 2003, 278, 49636–49643. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Lin, T.Y.; Chang, D.; Guo, Z. Thioflavin T as an Amyloid Dye: Fibril Quantification, Optimal Concentration and Effect on Aggregation. R. Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [Green Version]
- Barton, J.; Sebastian Arias, D.; Niyangoda, C.; Borjas, G.; Le, N.; Mohamed, S.; Muschol, M. Kinetic Transition in Amyloid Assembly as a Screening Assay for Oligomer-Selective Dyes. Biomolecules 2019, 9, 539. [Google Scholar] [CrossRef] [Green Version]
- Sulatskaya, A.I.; Rodina, N.P.; Sulatsky, M.I.; Povarova, O.I.; Antifeeva, I.A.; Kuznetsova, I.M.; Turoverov, K.K. Investigation of α-Synuclein Amyloid Fibrils Using the Fluorescent Probe Thioflavin T. Int. J. Mol. Sci. 2018, 19, 2486. [Google Scholar] [CrossRef] [Green Version]
- Sivanathan, V.; Hochschild, A. Generating Extracellular Amyloid Aggregates Using E. Coli Cells. Genes Dev. 2012, 26, 2659–2667. [Google Scholar] [CrossRef] [Green Version]
- Sunde, M.; Blake, C. The Structure of Amyloid Fibrils by Electron Microscopy and X-ray Diffraction. Adv. Protein Chem. 1997, 50, 123–159. [Google Scholar] [CrossRef] [PubMed]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef]
- Fowler, D.M.; Koulov, A.V.; Alory-Jost, C.; Marks, M.S.; Balch, W.E.; Kelly, J.W. Functional Amyloid Formation within Mammalian Tissue. PLoS Biol. 2006, 4, 0100–0107. [Google Scholar] [CrossRef] [PubMed]
- Ramshini, H.; Parrini, C.; Relini, A.; Zampagni, M.; Mannini, B.; Pesce, A.; Saboury, A.A.; Nemat-Gorgani, M.; Chiti, F. Large Proteins Have a Great Tendency to Aggregate but a Low Propensity to Form Amyloid Fibrils. PLoS ONE 2011, 6, e16075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, B.; van Niel, G.; Fowler, D.M.; Hurbain, I.; Luk, K.C.; Stayrook, S.E.; Lemmon, M.A.; Raposo, G.; Shorter, J.; Kelly, J.W.; et al. N-Terminal Domains Elicit Formation of Functional Pmel17 Amyloid Fibrils. J. Biol. Chem. 2009, 284, 35543–35555. [Google Scholar] [CrossRef] [Green Version]
- Stroud, J.C.; Liu, C.; Teng, P.K.; Eisenberg, D. Toxic Fibrillar Oligomers of Amyloid-β Have Cross-β Structure. Proc. Natl. Acad. Sci. USA 2012, 109, 7717–7722. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, L.C.; Melo-Ferreira, B.; Braga, C.A.; Fontes, G.N.; Mattos, L.J.; Lima, L.M.T.R. Stepwise Oligomerization of Murine Amylin and Assembly of Amyloid Fibrils. Biophys. Chem. 2013, 180–181, 135–144. [Google Scholar] [CrossRef]
- Raderschall, E.; Bazarov, A.; Cao, J.; Lurz, R.; Smith, A.; Mann, W.; Ropers, H.H.; Sedivy, J.M.; Golub, E.I.; Fritz, E.; et al. Formation of Higher-Order Nuclear Rad51 Structures Is Functionally Linked to P21 Expression and Protection from DNA Damage-Induced Apoptosis. J. Cell Sci. 2002, 115, 153–164. [Google Scholar] [CrossRef]
- Richardson, C.; Stark, J.M.; Ommundsen, M.; Jasin, M. Rad51 Overexpression Promotes Alternative Double-Strand Break Repair Pathways and Genome Instability. Oncogene 2004, 23, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Walsh, I.; Seno, F.; Tosatto, S.C.E.; Trovato, A. PASTA 2.0: An Improved Server for Protein Aggregation Prediction. Nucleic Acids Res. 2014, 42, W301–W307. [Google Scholar] [CrossRef] [PubMed]
- Garbuzynskiy, S.O.; Lobanov, M.Y.; Galzitskaya, O.V. FoldAmyloid: A Method of Prediction of Amyloidogenic Regions from Protein Sequence. Bioinformatics 2010, 26, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Conchillo-Solé, O.; de Groot, N.S.; Avilés, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: A Server for the Prediction and Evaluation of “Hot Spots” of Aggregation in Polypeptides. BMC Bioinform. 2007, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Maurer-Stroh, S.; Debulpaep, M.; Kuemmerer, N.; De La Paz, M.L.; Martins, I.C.; Reumers, J.; Morris, K.L.; Copland, A.; Serpell, L.; Serrano, L.; et al. Exploring the Sequence Determinants of Amyloid Structure Using Position-Specific Scoring Matrices. Nat. Methods 2010, 7, 237–242. [Google Scholar] [CrossRef]
- Ahmed, A.B.; Znassi, N.; Château, M.T.; Kajava, A.V. A Structure-Based Approach to Predict Predisposition to Amyloidosis. Alzheimer’s Dement. 2015, 11, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Jeyasekharan, A.D.; Liu, Y.; Hattori, H.; Pisupati, V.; Jonsdottir, A.B.; Rajendra, E.; Lee, M.; Sundaramoorthy, E.; Schlachter, S.; Kaminski, C.F.; et al. A Cancer-Associated BRCA2 Mutation Reveals Masked Nuclear Export Signals Controlling Localization. Nat. Struct. Mol. Biol. 2013, 20, 1191–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendra, E.; Venkitaraman, A.R. Two Modules in the BRC Repeats of BRCA2 Mediate Structural and Functional Interactions with the RAD51 Recombinase. Nucleic Acids Res. 2010, 38, 82–96. [Google Scholar] [CrossRef]
- Buisson, R.; Dion-Côté, A.M.; Coulombe, Y.; Launay, H.; Cai, H.; Stasiak, A.Z.; Stasiak, A.; Xia, B.; Masson, J.Y. Cooperation of Breast Cancer Proteins PALB2 and Piccolo BRCA2 in Stimulating Homologous Recombination. Nat. Struct. Mol. Biol. 2010, 17, 1247–1254. [Google Scholar] [CrossRef] [Green Version]
- Buchhop, S.; Gibson, M.K.; Wang, X.W.; Wagner, P.; Stürzbecher, H.W.; Harris, C.C. Interaction of P53 with the Human Rad51 Protein. Nucleic Acids Res. 1997, 25, 3868–3874. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Furlong, R.A.; Narain, Y.; Rankin, J.; Wyttenbach, A.; Rubinsztein, D.C. Alpha-Synuclein Overexpression Promotes Aggregation of Mutant Huntingtin. Biochem. J. 2000, 346, 577. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hu, H.Y. Sequestration of Cellular Interacting Partners by Protein Aggregates: Implication in a Loss-of-Function Pathology. FEBS J. 2016, 283, 3705–3717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demeyer, A.; Benhelli-Mokrani, H.; Chénais, B.; Weigel, P.; Fleury, F. Inhibiting Homologous Recombination by Targeting RAD51 Protein. Biochim. Biophys. Acta-Rev. Cancer 2021, 1876, 188597. [Google Scholar] [CrossRef]
- Orhan, E.; Velazquez, C.; Tabet, I.; Sardet, C.; Theillet, C. Regulation of Rad51 at the Transcriptional and Functional Levels: What Prospects for Cancer Therapy? Cancers 2021, 13, 2930. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.R.; Bernstein, K.A. RAD-Ical New Insights into RAD51 Regulation. Genes 2018, 9, 629. [Google Scholar] [CrossRef] [Green Version]
- Antony, H.; Wiegmans, A.P.; Wei, M.Q.; Chernoff, Y.O.; Khanna, K.K.; Munn, A.L. Potential Roles for Prions and Protein-Only Inheritance in Cancer. Cancer Metastasis Rev. 2012, 31, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Park, S.; Pentek, C.; Liebman, S.W. Tumor Suppressor Protein P53 Expressed in Yeast Can Remain Diffuse, Form a Prion, or Form Unstable Liquid-like Droplets. iScience 2021, 24, 102000. [Google Scholar] [CrossRef]
- Lin, H.Y.; Tan, G.Q.; Liu, Y.; Lin, S.Q. The Prognostic Value of Serum Amyloid A in Solid Tumors: A Meta-Analysis. Cancer Cell Int. 2019, 19, 62. [Google Scholar] [CrossRef]
- Findeisen, P.; Zapatka, M.; Peccerella, T.; Matzk, H.; Neumaier, M.; Schadendorf, D.; Ugurel, S. Serum Amyloid A as a Prognostic Marker in Melanoma Identified by Proteomic Profiling. J. Clin. Oncol. 2009, 27, 2199–2208. [Google Scholar] [CrossRef]
- Biaoxue, R.; Hua, L.; Wenlong, G.; Shuanying, Y. Increased Serum Amyloid A as Potential Diagnostic Marker for Lung Cancer: A Meta-Analysis Based on Nine Studies. BMC Cancer 2016, 16, 836. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Liu, F.; Higuchi, K.; Sawashita, J.; Fu, X.; Zhang, L.; Zhang, L.; Fu, L.; Tong, Z.; Higuchi, K. Serum Amyloid A Expression in the Breast Cancer Tissue Is Associated with Poor Prognosis. Oncotarget 2016, 7, 35843. [Google Scholar] [CrossRef] [PubMed]
- Fourie, C.; Shridas, P.; Davis, T.; de Villiers, W.J.S.; Engelbrecht, A.M. Serum Amyloid A and Inflammasome Activation: A Link to Breast Cancer Progression? Cytokine Growth Factor Rev. 2021, 59, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Beatty, G.L. Serum Amyloid A Proteins and Their Impact on Metastasis and Immune Biology in Cancer. Cancers 2021, 13, 3179. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Iashchishyn, I.A.; Pansieri, J.; Nyström, S.; Klementieva, O.; Kara, J.; Horvath, I.; Moskalenko, R.; Rofougaran, R.; Gouras, G.; et al. S100A9-Driven Amyloid-Neuroinflammatory Cascade in Traumatic Brain Injury as a Precursor State for Alzheimer’s Disease. Sci. Rep. 2018, 8, 12836. [Google Scholar] [CrossRef]
- Lv, Z.; Li, W.; Wei, X. S100A9 Promotes Prostate Cancer Cell Invasion by Activating TLR4/NF-ΚB/Integrin Β1/FAK Signaling. Onco. Targets. Ther. 2020, 13, 6443. [Google Scholar] [CrossRef]
- Markowitz, J.; Carson, W.E. Review of S100A9 Biology and Its Role in Cancer. Biochim. Biophys. Acta 2013, 1835, 100. [Google Scholar] [CrossRef]
- Brat, D.J.; Gearing, M.; Goldthwaite, P.T.; Wainer, B.H.; Burger, P.C. Tau-Associated Neuropathology in Ganglion Cell Tumours Increases with Patient Age but Appears Unrelated to ApoE Genotype. Neuropathol. Appl. Neurobiol. 2001, 27, 197–205. [Google Scholar] [CrossRef]
- Stridsberg, M.; Eriksson, B.; Lundqvist, G.; Skogseid, B.; Wilander, E.; Öberg, K. Islet Amyloid Polypeptide (IAPP) in Patients with Neuroendocrine Tumours. Regul. Pept. 1995, 55, 119–131. [Google Scholar] [CrossRef]
- Bononi, P.L.; Martinez, A.J.; Nelson, R.B.; Amico, J.A. Amyloid Deposits in a Prolactin-Producing Pituitary Adenoma. J. Endocrinol. Investig. 1993, 16, 339–343. [Google Scholar] [CrossRef]
- Murphy, C.L.; Kestler, D.P.; Foster, J.S.; Wang, S.; Macy, S.D.; Kennel, S.J.; Carlson, E.R.; Hudson, J.; Weiss, D.T.; Solomon, A. Odontogenic Ameloblast-Associated Protein Nature of the Amyloid Found in Calcifying Epithelial Odontogenic Tumors and Unerupted Tooth Follicles. Amyloid 2008, 15, 89–95. [Google Scholar] [CrossRef]
- Kestler, D.P.; Foster, J.S.; Macy, S.D.; Murphy, C.L.; Weiss, D.T.; Solomon, A. Expression of Odontogenic Ameloblast-Associated Protein (ODAM) in Dental and Other Epithelial Neoplasms. Mol. Med. 2008, 14, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Chandramowlishwaran, P.; Sun, M.; Casey, K.L.; Romanyuk, A.V.; Grizel, A.V.; Sopova, J.V.; Rubel, A.A.; Nussbaum-Krammer, C.; Vorberg, I.M.; Chernoff, Y.O. Mammalian Amyloidogenic Proteins Promote Prion Nucleation in Yeast. J. Biol. Chem. 2018, 293, 3436–3450. [Google Scholar] [CrossRef] [PubMed]
- Campeau, E.; Ruhl, V.E.; Rodier, F.; Smith, C.L.; Rahmberg, B.L.; Fuss, J.O.; Campisi, J.; Yaswen, P.; Cooper, P.K.; Kaufman, P.D. A Versatile Viral System for Expression and Depletion of Proteins in Mammalian Cells. PLoS ONE 2009, 4, e6529. [Google Scholar] [CrossRef] [PubMed]
- Kachkin, D.V.; Khorolskaya, J.I.; Ivanova, J.S.; Rubel, A.A. An Efficient Method for Isolation of Plasmid DNA for Transfection of Mammalian Cell Cultures. Methods Protoc. 2020, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Sopova, J.V.; Koshel, E.I.; Belashova, T.A.; Zadorsky, S.P.; Sergeeva, A.V.; Siniukova, V.A.; Shenfeld, A.A.; Velizhanina, M.E.; Volkov, K.V.; Nizhnikov, A.A.; et al. RNA-Binding Protein FXR1 Is Presented in Rat Brain in Amyloid Form. Sci. Rep. 2019, 9, 18983. [Google Scholar] [CrossRef] [PubMed]
- Sivanathan, V.; Hochschild, A. A Bacterial Export System for Generating Extracellular Amyloid Aggregates. Nat. Protoc. 2013, 8, 1381. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachkin, D.V.; Volkov, K.V.; Sopova, J.V.; Bobylev, A.G.; Fedotov, S.A.; Inge-Vechtomov, S.G.; Galzitskaya, O.V.; Chernoff, Y.O.; Rubel, A.A.; Aksenova, A.Y. Human RAD51 Protein Forms Amyloid-like Aggregates In Vitro. Int. J. Mol. Sci. 2022, 23, 11657. https://doi.org/10.3390/ijms231911657
Kachkin DV, Volkov KV, Sopova JV, Bobylev AG, Fedotov SA, Inge-Vechtomov SG, Galzitskaya OV, Chernoff YO, Rubel AA, Aksenova AY. Human RAD51 Protein Forms Amyloid-like Aggregates In Vitro. International Journal of Molecular Sciences. 2022; 23(19):11657. https://doi.org/10.3390/ijms231911657
Chicago/Turabian StyleKachkin, Daniel V., Kirill V. Volkov, Julia V. Sopova, Alexander G. Bobylev, Sergei A. Fedotov, Sergei G. Inge-Vechtomov, Oxana V. Galzitskaya, Yury O. Chernoff, Aleksandr A. Rubel, and Anna Y. Aksenova. 2022. "Human RAD51 Protein Forms Amyloid-like Aggregates In Vitro" International Journal of Molecular Sciences 23, no. 19: 11657. https://doi.org/10.3390/ijms231911657