
Hepatobiliary Thyroid Hormone Deficiency Impacts Bile Acid Hydrophilicity and Aquaporins in Cholestatic C57BL/6J Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
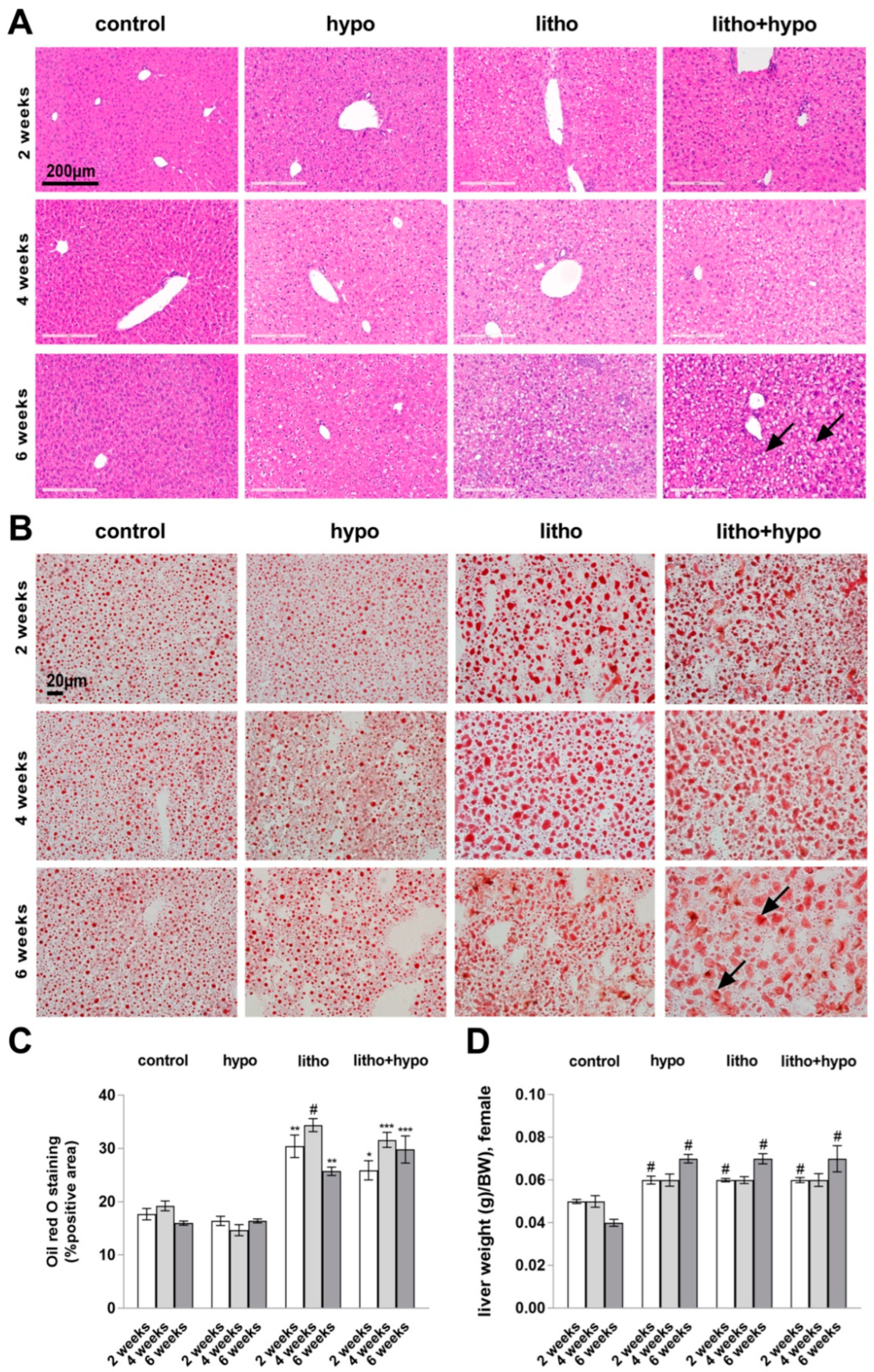
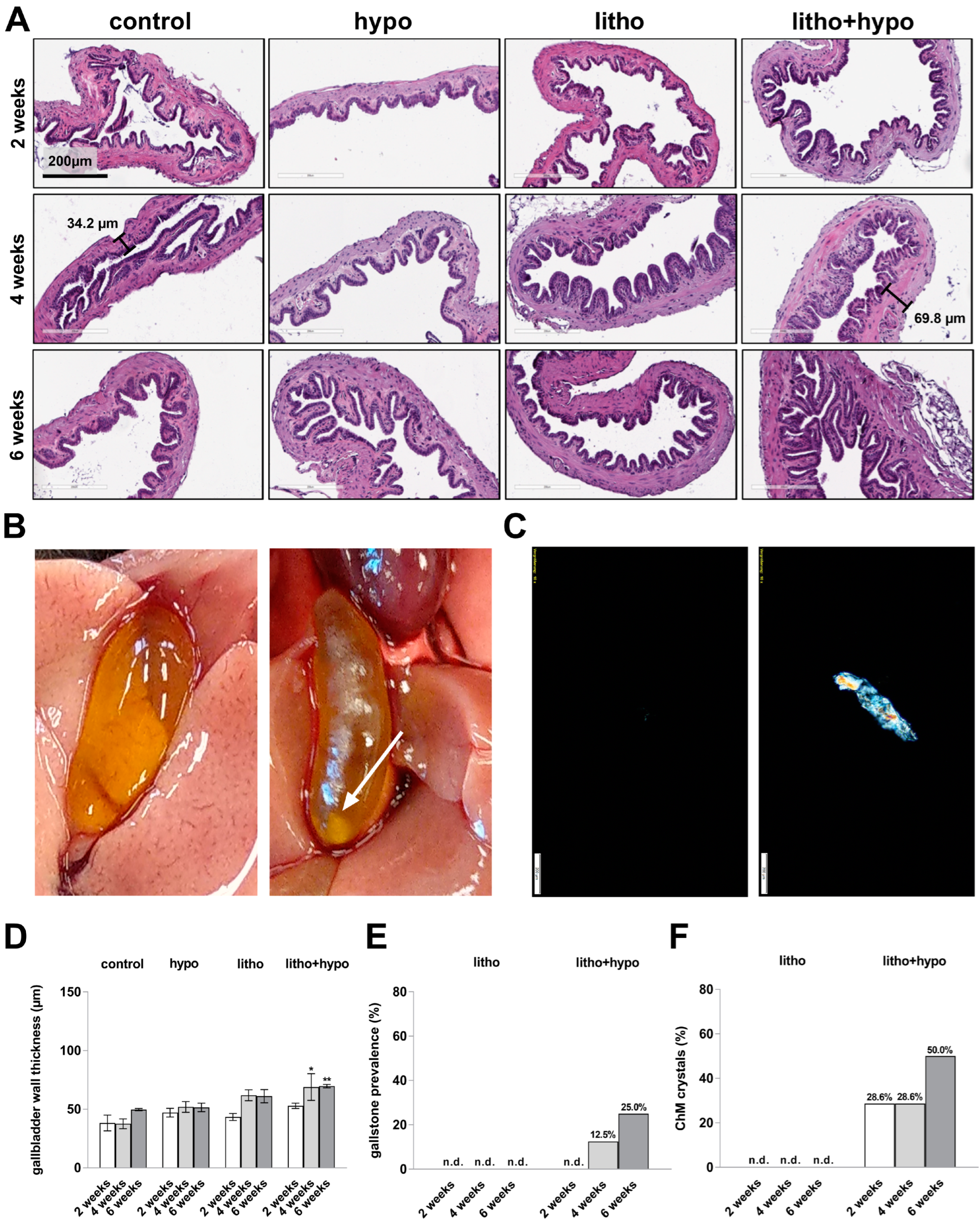
2.1. Hepatobiliary Histology and Function under Thyroid Hormone Deficient and Lithogenic Conditions in Female C57BL/6J Mice
2.2. TH Deficiency Leads to Cholesterol Gallstone Formation in Lithogenic Diet Supplemented Female C57BL/6J Mice
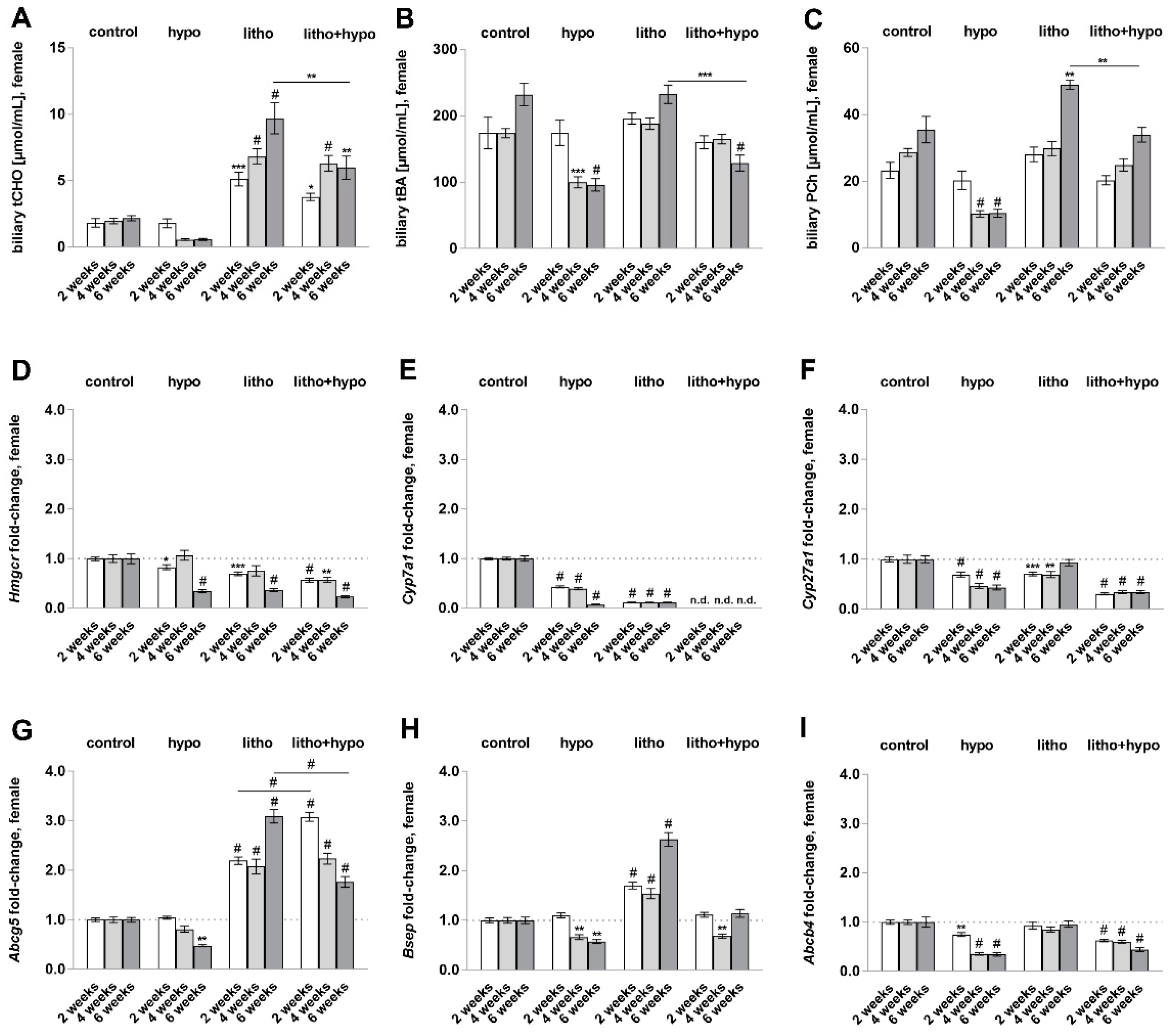
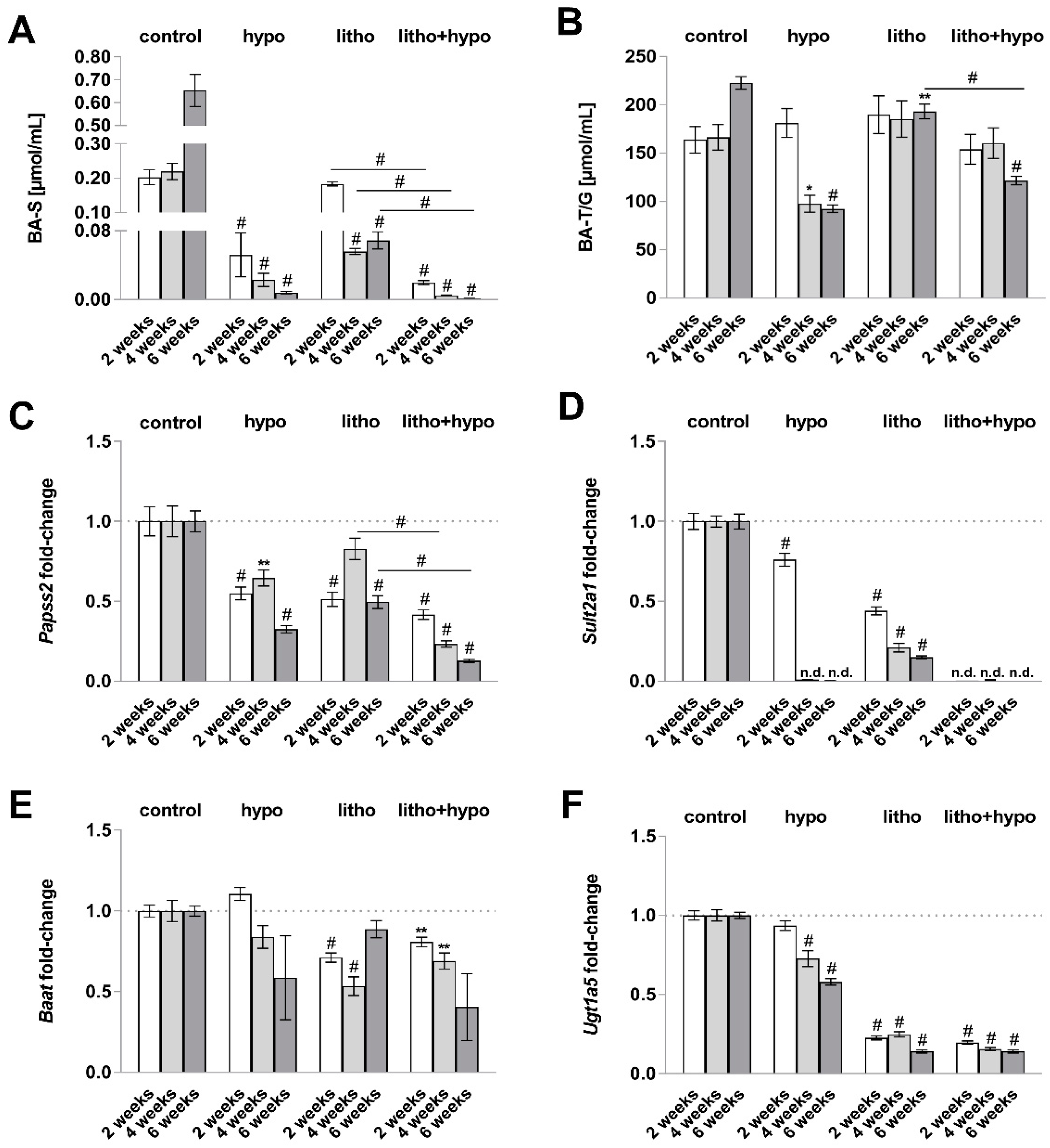
2.3. TH Deficiency Diminishes Bile Acid Hydrophilicity in Lithogenic Diet Supplemented Female C57BL/6J Mice
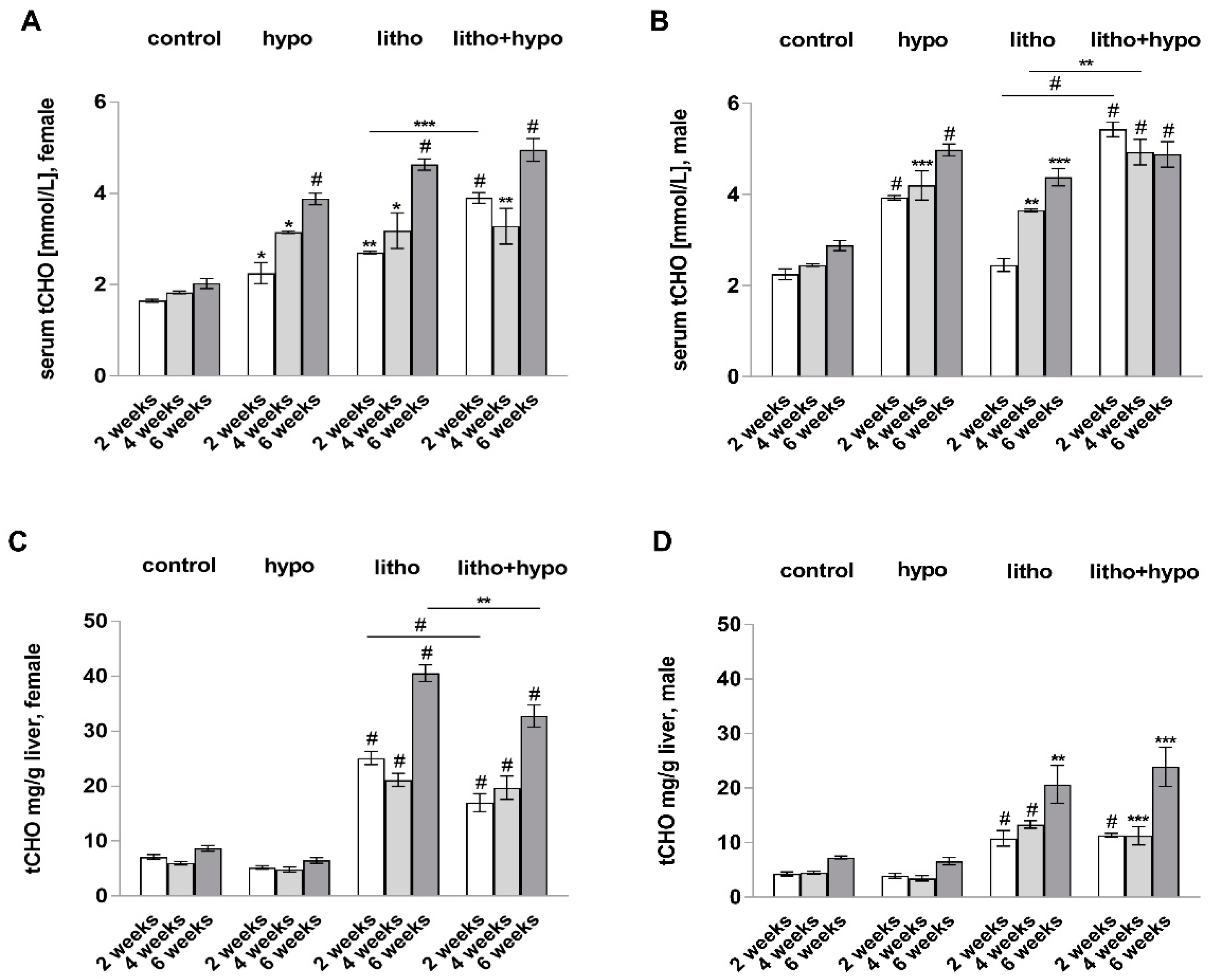
2.4. TH Deficiency Reduces Hepatic Total Cholesterol Content in Lithogenic Diet Supplemented Female but Not Male C57BL/6J Mice
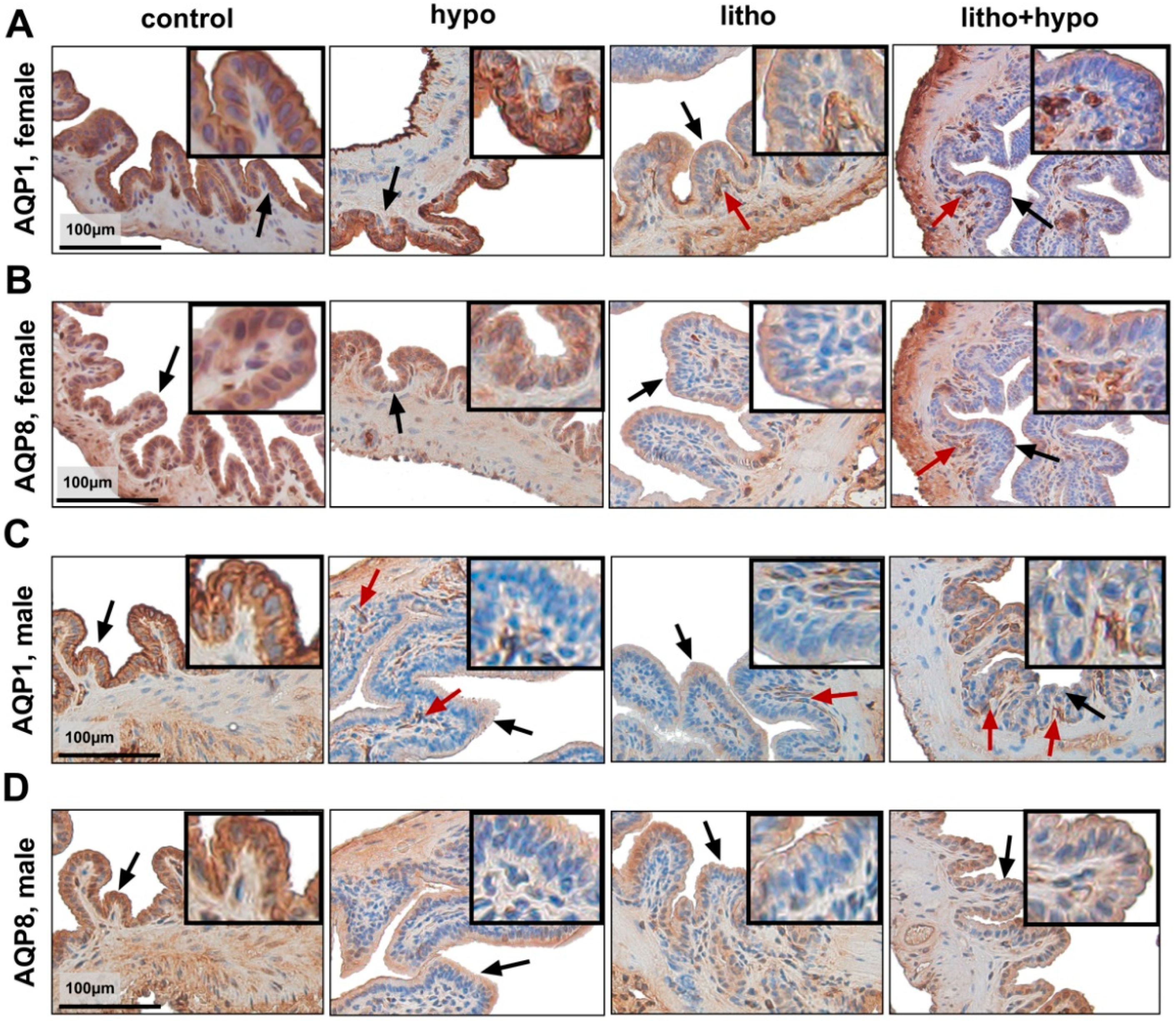
2.5. Hepatobiliary Aquaporin Expression and Localization Sex-Dependently Differ by TH Deficiency in C57BL/6J Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Collection of Serum, Bile Fluid, and Tissue Samples
4.3. Serum Measurements
4.4. Bile Measurements
4.5. Hepatic Total Cholesterol Content
4.6. Histological Analysis
4.7. Immunohistochemistry
4.8. Quantitative Real-Time PCR
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inkinen, J.; Sand, J.; Nordback, I. Association between common bile duct stones and treated hypothyroidism. Hepatogastroenterology 2000, 47, 919–921. [Google Scholar] [PubMed]
- Volzke, H.; Robinson, D.M.; John, U. Association between thyroid function and gallstone disease. World J. Gastroenterol. 2005, 11, 5530–5534. [Google Scholar] [CrossRef] [PubMed]
- Laukkarinen, J.; Sand, J.; Nordback, I. The underlying mechanisms: How hypothyroidism affects the formation of common bile duct stones-a review. HPB Surg. 2012, 2012, 102825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Erpecum, K.J.; Van Berge-Henegouwen, G.P. Gallstones: An intestinal disease? Gut 1999, 44, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Portincasa, P.; Wang, D.Q. Intestinal absorption, hepatic synthesis, and biliary secretion of cholesterol: Where are we for cholesterol gallstone formation? Hepatology 2012, 55, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Ness, G.C.; Pendleton, L.C.; Li, Y.C.; Chiang, J.Y. Effect of thyroid hormone on hepatic cholesterol 7 alpha hydroxylase, LDL receptor, HMG-CoA reductase, farnesyl pyrophosphate synthetase and apolipoprotein A-I mRNA levels in hypophysectomized rats. Biochem. Biophys. Res. Commun. 1990, 172, 1150–1156. [Google Scholar] [CrossRef]
- Hashimoto, K.; Cohen, R.N.; Yamada, M.; Markan, K.R.; Monden, T.; Satoh, T.; Mori, M.; Wondisford, F.E. Cross-talk between thyroid hormone receptor and liver X receptor regulatory pathways is revealed in a thyroid hormone resistance mouse model. J. Biol. Chem. 2006, 281, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonde, Y.; Plösch, T.; Kuipers, F.; Angelin, B.; Rudling, M. Stimulation of murine biliary cholesterol secretion by thyroid hormone is dependent on a functional ABCG5/G8 complex. Hepatology 2012, 56, 1828–1837. [Google Scholar] [CrossRef] [Green Version]
- Gautherot, J.; Claudel, T.; Cuperus, F.; Fuchs, C.D.; Falguieres, T.; Trauner, M. Thyroid hormone receptor beta1 stimulates ABCB4 to increase biliary phosphatidylcholine excretion in mice. J. Lipid Res. 2018, 59, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, D.J.; Wallace, R.B.; Rodabough, R.J.; Greenland, P.; LaCroix, A.Z.; Limacher, M.C.; Larson, J.C. Effect of estrogen therapy on gallbladder disease. JAMA 2005, 293, 330–339. [Google Scholar] [CrossRef]
- Alexander, M.; Portman, O.W. Different susceptibilities to the formation of cholesterol gallstones in mice. Hepatology 1987, 7, 257–265. [Google Scholar] [CrossRef]
- Kube, I.; Tardio, L.B.; Hofmann, U.; Ghallab, A.; Hengstler, J.G.; Führer, D.; Zwanziger, D. Hypothyroidism Increases Cholesterol Gallstone Prevalence in Mice by Elevated Hydrophobicity of Primary Bile Acids. Thyroid 2021, 31, 973–984. [Google Scholar] [CrossRef]
- Moschetta, A.; Bookout, A.L.; Mangelsdorf, D.J. Prevention of cholesterol gallstone disease by FXR agonists in a mouse model. Nat. Med. 2004, 10, 1352–1358. [Google Scholar] [CrossRef]
- Calamita, G.; Ferri, D.; Bazzini, C.; Mazzone, A.; Botta, G.; Liquori, G.E.; Paulmichl, M.; Portincasa, P.; Meyer, G.; Svelto, M. Expression and subcellular localization of the AQP8 and AQP1 water channels in the mouse gall-bladder epithelium. Biol. Cell 2005, 97, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Carle, A.; Laurberg, P.; Pedersen, I.B.; Knudsen, N.; Perrild, H.; Ovesen, L.; Rasmussen, L.B.; Jorgensen, T. Epidemiology of subtypes of hypothyroidism in Denmark. Eur. J. Endocrinol. 2006, 154, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Araujo, R.L.; Andrade, B.M.; Padron, A.S.; Gaidhu, M.P.; Perry, R.L.S.; Carvalho, D.P.; Ceddia, R.B. High-fat diet increases thyrotropin and oxygen consumption without altering circulating 3,5,3′-triiodothyronine (T3) and thyroxine in rats: The role of iodothyronine deiodinases, reverse T3 production, and whole-body fat oxidation. Endocrinoloy 2010, 151, 3460–3469. [Google Scholar] [CrossRef]
- Kahle, M.; Horsch, M.; Fridrich, B.; Seelig, A.; Schultheiß, J.; Leonhardt, J.; Irmler, M.; Beckers, J.; Rathkolb, B.; Wolf, E.; et al. Phenotypic comparison of common mouse strains developing high-fat diet-induced hepatosteatosis. Mol. Metab. 2013, 2, 435–446. [Google Scholar] [CrossRef]
- Xia, S.-F.; Duan, X.-M.; Hao, L.-Y.; Li, L.-T.; Cheng, X.-R.; Xie, Z.-X.; Qiao, Y.; Li, L.-R.; Tang, X.; Shi, Y.-H.; et al. Role of thyroid hormone homeostasis in obesity-prone and obesity-resistant mice fed a high-fat diet. Metabolism 2015, 64, 566–579. [Google Scholar] [CrossRef]
- Lehmann, G.L.; Larocca, M.C.; Soria, L.R.; Marinelli, R.A. Aquaporins: Their role in cholestatic liver disease. World J. Gastroenterol. 2008, 14, 7059–7067. [Google Scholar] [CrossRef]
- van Erpecum, K.J.; Wang, D.Q.H.; Moschetta, A.; Ferri, D.; Svelto, M.; Portincasa, P.; Hendrickx, J.J.; Schipper, M.; Calamita, G. Gallbladder histopathology during murine gallstone formation: Relation to motility and concentrating function. J. Lipid Res. 2006, 47, 32–41. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Ma, T.; Verkman, A.S. Very high aquaporin-1 facilitated water permeability in mouse gallbladder. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G816–G822. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.E.S.; Clementino-Neto, J.; Mendes, C.B.; Franzon, N.H.; de Oliveira Costa, E.; Moura-Neto, V.; Ximenes-da-Silva, A. Evidence of Aquaporin 4 Regulation by Thyroid Hormone During Mouse Brain Development and in Cultured Human Glioblastoma Multiforme Cells. Front. Neurosci. 2019, 13, 317. [Google Scholar] [CrossRef]
- Jaruvongvanich, V.; Sanguankeo, A.; Upala, S. Significant Association Between Gallstone Disease and Nonalcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. Dig. Dis. Sci. 2016, 61, 2389–2396. [Google Scholar] [CrossRef]
- Rodriguez, A.; Gena, P.; Mendez-Gimenez, L.; Rosito, A.; Valenti, V.; Rotellar, F.; Sola, I.; Moncada, R.; Silva, C.; Svelto, M.; et al. Reduced hepatic aquaporin-9 and glycerol permeability are related to insulin resistance in non-alcoholic fatty liver disease. Int. J. Obes. 2014, 38, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Marinelli, R.A.; Tesse, A.; Frühbeck, G.; Calamita, G. Sexual Dimorphism of Adipose and Hepatic Aquaglyceroporins in Health and Metabolic Disorders. Front. Endocrinol. 2015, 6, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iena, F.M.; Jul, J.B.; Vegger, J.B.; Lodberg, A.; Thomsen, J.S.; Brüel, A.; Lebeck, J. Sex-Specific Effect of High-Fat Diet on Glycerol Metabolism in Murine Adipose Tissue and Liver. Front. Endocrinol. 2020, 11, 577650. [Google Scholar] [CrossRef] [PubMed]
- Marrone, J.; Danielli, M.; Gaspari, C.I.; Marinelli, R.A. Adenovirus-Mediated Human Aquaporin-1 Expression in Hepatocytes Improves Lipopolysaccharide-Induced Cholestasis. IUBMB 2017, 69, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Danielli, M.; Capiglioni, A.M.; Marrone, J.; Calamita, G.; Marinelli, R.A. Cholesterol Can Modulate Mitochondrial Aquaporin-8 Expression in Human Hepatic Cells. IUBMB 2017, 69, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Moreno, N.R.; Balaguer, I.; Mendez-Gimenez, L.; Becerril, S.; Catalan, V.; Gomez-Ambrosi, J.; Portincasa, P.; Calamita, G.; Soveral, G.; et al. Leptin administration restores the altered adipose and hepatic expression of aquaglyceroporins improving the non-alcoholic fatty liver of ob/ob mice. Sci. Rep. 2015, 5, 12067. [Google Scholar] [CrossRef] [Green Version]
- Zwanziger, D.; Rakov, H.; Engels, K.; Moeller, L.C.; Führer, D. Sex-Dependent Claudin-1 Expression in the Liver of Euthyroid and Hypothyroid Mice. Eur. Thyroid J. 2015, 4 (Suppl. 1), 67–73. [Google Scholar] [CrossRef]
- Ghallab, A.; Hassan, R.; Hofmann, U.; Friebel, A.; Hobloss, Z.; Brackhagen, L.; Begher-Tibbe, B.; Myllys, M.; Reinders, J.; Overbeck, N.; et al. Interruption of bile acid uptake by hepatocytes after acetaminophen overdose ameliorates hepatotoxicity. J. Hepatol. 2022, 77, 71–83. [Google Scholar] [CrossRef]
- Maier, K.; Hofmann, U.; Bauer, A.; Niebel, A.; Vacun, G.; Reuss, M.; Mauch, K. Quantification of statin effects on hepatic cholesterol synthesis by transient (13)C-flux analysis. Metab. Eng. 2009, 11, 292–309. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kube, I.; Kowalczyk, M.; Hofmann, U.; Ghallab, A.; Hengstler, J.G.; Führer, D.; Zwanziger, D. Hepatobiliary Thyroid Hormone Deficiency Impacts Bile Acid Hydrophilicity and Aquaporins in Cholestatic C57BL/6J Mice. Int. J. Mol. Sci. 2022, 23, 12355. https://doi.org/10.3390/ijms232012355
Kube I, Kowalczyk M, Hofmann U, Ghallab A, Hengstler JG, Führer D, Zwanziger D. Hepatobiliary Thyroid Hormone Deficiency Impacts Bile Acid Hydrophilicity and Aquaporins in Cholestatic C57BL/6J Mice. International Journal of Molecular Sciences. 2022; 23(20):12355. https://doi.org/10.3390/ijms232012355
Chicago/Turabian StyleKube, Irina, Manuela Kowalczyk, Ute Hofmann, Ahmed Ghallab, Jan Georg Hengstler, Dagmar Führer, and Denise Zwanziger. 2022. "Hepatobiliary Thyroid Hormone Deficiency Impacts Bile Acid Hydrophilicity and Aquaporins in Cholestatic C57BL/6J Mice" International Journal of Molecular Sciences 23, no. 20: 12355. https://doi.org/10.3390/ijms232012355