
Characterisation of Trichomonas vaginalis Isolates Collected from Patients in Vienna between 2019 and 2021

Abstract
:1. Introduction
2. Results
2.1. Collection and Characteristic of TV Isolates
2.2. Detection of TVVs in Clinical Isolates
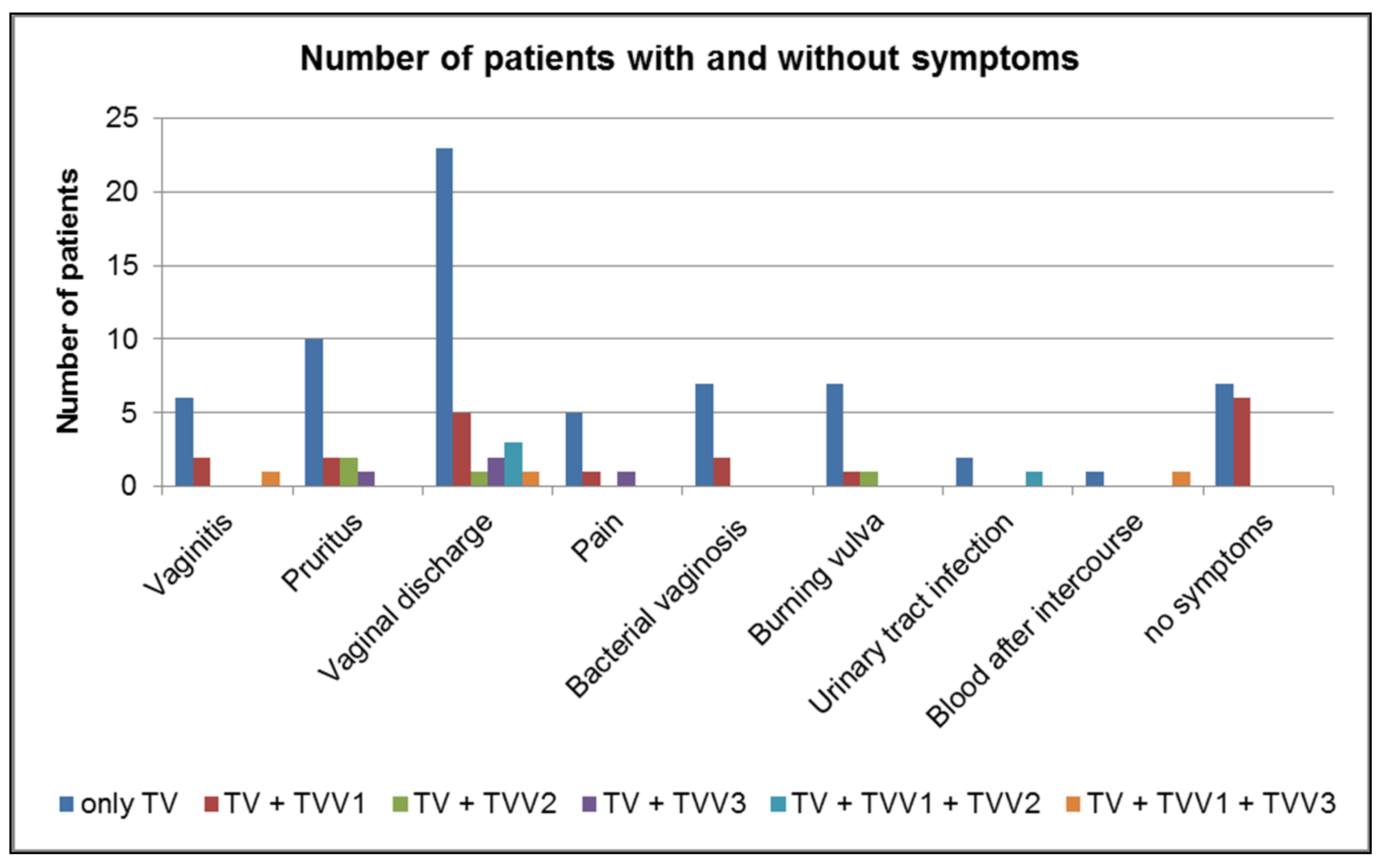
2.3. Prevalence of TVVs in Symptomatic and Asymptomatic Patients
2.4. Detection of M. hominis in TV and Vaginal/Urethral Swabs
2.5. Prevalence of M. hominis in Symptomatic and Asymptomatic Patients
2.6. MICs
2.7. LDH Assay
3. Discussion
4. Materials and Methods
4.1. Collection and Culture of Trichomonas vaginalis Isolates
4.2. RNA Extraction and First-Strand cDNA Synthesis
4.3. PCR for TVVs
4.4. Detection of M. hominis
4.5. Determination of Minimal Inhibitory Concentrations (MICs)
4.6. LDH Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rowley, J.; Hoorn, S.V.; Korenromp, E.; Low, N.; Unemo, M.; Abu-Raddad, L.J.; Chico, R.M.; Smolak, A.; Newman, L.; Gottlieb, S.; et al. Chlamydia, gonorrhoea, trichomoniasis and syphilis: Global prevalence and incidence estimates. Bull. World Health Organ. 2016, 97, 548–562. [Google Scholar] [CrossRef]
- Sutton, M.; Sternberg, M.; Koumans, E.H.; McQuillan, G.; Berman, S.; Markowitz, L. The prevalence of Trichomonas vaginalis infection among reproductive-age women in the United States, 2001–2004. Clin. Infect. Dis. 2007, 45, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Depuydt, C.E.; Leuridan, E.; Van Damme, P.; Bogers, J.; Vereecken, A.J.; Donders, G.G. Epidemiology of Trichomonas vaginalis and human papillomavirus infection detected by real-time PCR in flanders. Gynecol Obstet Investig. 2010, 70, 273–280. [Google Scholar] [CrossRef]
- Petrin, D.; Delgaty, K.; Bhatt, R.; Garber, G. Clinical and microbial aspects of Trichomonas vaginalis. Clin. Microbiol Rev. 1998, 11, 300–317. [Google Scholar] [CrossRef] [Green Version]
- Edwards, T.; Burke, P.; Smalley, H.; Hobbs, G. Trichomonas vaginalis: Clinical relevance, pathogenicity and diagnosis. Crit Rev. Microbiol. 2016, 42, 406–417. [Google Scholar] [CrossRef] [PubMed]
- McClelland, R.S.; Sangare, L.; Hassan, W.M.; Lavreys, I.; Mandaliya, K.; Kiarie, J.; Ndinya-Achola, J.; Jaoko, W.; Baeten, J.M. Infections with Trichomonas vaginalis increases the risk of HIV-1 acquisition. J. Infect. Dis. 2007, 195, 698–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Icho, T.; Wickner, R.B. The double-stranded RNA genome of yeast virus L-A encodes its own putative RNA polymerase by fusing two open reading frames. J. Biol. Chem. 1989, 264, 6717–6723. [Google Scholar] [CrossRef]
- Janssen, M.E.W.; Takagi, Y.; Parent, K.N.; Cardone, G.; Nibert, M.L.; Baker, T.S. Three-dimensional structure of a protozoal double-stranded RNA virus that infects the enteric pathogen Giardia lamblia. J. Virol. 2015, 89, 1182–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartley, M.A.; Ronet, C.; Zangger, H.; Beverley, S.M.; Fasel, N. Leishmania RNA virus: When the host pays the toll. Front. Cell Infect. Microbiol. 2012, 2, 99. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.L.; Wang, C.C. Viruses of the protozoa. Annu. Rev. Microbiol. 1991, 45, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Graves, K.J.; Ghosh, A.P.; Schmidt, N.; Augostini, P.; Secor, W.E.; Schwebke, J.R.; Martin, D.H.; Kissinger, P.J.; Muzny, C.A. Trichomonas vaginalis virus among women with trichomoniasis and association with demographics, clinical outcomes, and metronidazole resistance. Clin. Infect. Dis. 2019, 69, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Benchimol, M.; Chang, T.H.; Alderete, J.F. Trichomonas vaginalis: Observation of coexistence of multiple viruses in the same isolate. FEMS Microbiol. Lett. 2002, 215, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, D.; Khoshnan, A.; Alderete, J.F. Involvement of dsRNA virus in the protein composition and growth kinetics of host Trichomonas vaginalis. Arch. Virol. 1997, 142, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Razin, S. Peculiar properties of Mycoplasmas: The smallest self-replicating prokaryotes. FEMS Microbiol. Lett. 1992, 100, 423–431. [Google Scholar] [CrossRef]
- Razin, S.; Yogev, D.; Naot, Y. Molecular biology and pathogenicity of Mycoplasmas. Microbiol. Mol. Biol. Rev. 1998, 62, 1094–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Sun, C.L.; Carlisle, E.M.; Morowitz, M.J.; Banfield, J.F.; Relman, D.A. Candidatus Mycoplasma girerdii replicates, diversifies, and co-occurs with Trichomonas vaginalis in the oral cavity of a premature infant. Sci. Rep. 2017, 7, 3764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappelli, P.; Addis, M.F.; Carta, F.; Fiori, P.L. Mycoplasma hominis parasitism of Trichomonas vaginalis. Lancet 1998, 352, 1286. [Google Scholar] [CrossRef]
- Meyer, R.D.; Clough, W. Extragenital Mycoplasma hominis infections in adults: Emphasis on immunosuppression. Clin. Infect. Dis. 1993, 17 (Suppl. S1), 243–249. [Google Scholar] [CrossRef] [PubMed]
- Cosar, C.; Julou, L. Activity of 1-(2-hydroxyethyl)-2-methyl-5-nitroimidazole (8823 RP) against experimental Trichomonas vaginalis infection. Ann. Inst. Pasteur 1959, 96, 238–241. [Google Scholar]
- Robinson, S.C. Trichomonal vaginitis resistance to metronidazole. Canadian Med. Assoc. J. 1962, 86, 665. [Google Scholar]
- Muzny, C.; Van Gerwen, O.T.; Kissinger, P. Updates in Trichomonas treatment including persistent infection and 5-nitroimidazole hypersensitivity. Curr. Opin. Infect. Dis. 2020, 33, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Margarita, V.; Marongiu, A.; Diaz, N.; Dessi, D.; Fiori, P.L.; Rappelli, P. Prevalence of double-stranded RNA virus in Trichomonas vaginalis isolated in Italy and association with the symbiont Mycoplasma hominis. Parasitol. Res. 2019, 118, 3565–3570. [Google Scholar] [CrossRef] [PubMed]
- Masha, S.C.; Cools, P.; Crucitti, T.; Sanders, E.J.; Vaneechouette, M. Molecular typing of Trichomonas vaginalis isolates by actin gene sequence analysis and carriage of T. vaginalis viruses. Parasit. Vectors 2017, 10, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Luz Becker, D.; dos Santos, O.; Frasson, A.P.; de Vargas Rigo, G.; Macedo, A.J.; Tasca, T. High rates of double-stranded RNA viruses and Mycoplasma hominis in Trichomonas vaginalis clinical isolates in South Brazil. Infect. Genet. Evol. 2015, 34, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichorova, R.N.; Lee, Y.; Yamamoto, H.S.; Takagi, Y.; Hayes, G.R.; Goodman, R.P.; Chepa-Lotrea, X.; Buck, O.R.; Murray, R.; Kula, T. Endobiont viruses sensed by the human host—Beyond conventional antiparasitic therapy. PLoS ONE 2012, 7, e48418. [Google Scholar] [CrossRef] [PubMed]
- Nalçaci, M.; Karakuş, M.; Yılmaz, B.; Demir, S.; Özbilgin, A.; Özbel, Y.; Töz, S. Detection of Leishmania RNA virus 2 in Leishmania species from Turkey. Trop. Med. Hyg. 2019, 113, 410–417. [Google Scholar] [CrossRef]
- Fermin, G.; Mazumdar-Leighton, S.; Tennant, P. Viruses of Prokaryotes, Protozoa, Fungi, and Chromista. Viruses 2018, 9, 217–244. [Google Scholar]
- Fraga, J.; Rojas, L.; Sariego, I.; Fernández-Calienes, A.; Nuñez, F.A. Species typing of Cuban Trichomonas vaginalis virus by RT-PCR, and association of TVV-2 with high parasite adhesion levels and high pathogenicity in patients. Arch. Virol. 2012, 157, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Alderete, J.F.; Pearlmann, E. Pathogenic Trichomonas vaginalis cytotoxicity to cell culture monolayers. Br. J. Vener Dis. 1984, 60, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.B.; Piccoli Frasson, A.; Meirelles, L.C.; Tasca, T. Adenosine, but not guanosine, protects vaginal epithelial cells from Trichomonas vaginalis cytotoxicity. Microbes Infect. 2017, 9, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.E.; Nielsen, N.H.; Lind, I.; Rhodes, J.M. Morphological studies of the cytotoxicity of Trichomonas vaginalis to normal human vaginal epithelial cells in vitro. Genitourin Med. 1986, 62, 240–246. [Google Scholar]
- Gilbert, R.O.; Elia, G.; Beach, D.H.; Klaessig, S.; Singh, B.N. Cytopathogenic effect of Trichomonas vaginalis on human vaginal epithelial cells cultured in vitro. Infect. Immun. 2000, 68, 4200–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narayanasamy, R.K.; Rada, P.; Zdrha, A.; van Ranst, M.; Neyts, J.; Tachezy, J. Cytidine nucleoside analog is an effective antiviral drug against Trichomonasvirus. J. Microbiol. Immunol. Infect. 2022, 55, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Schwebke, J.R.; Barrientes, F. Prevalence of Trichomonas vaginalis isolates with resistance to metronidazole and tinidazole. Antimicrob. Agents Chemother. 2006, 50, 4209–4210. [Google Scholar] [CrossRef]
- Upcroft, J.A.; Dunn, L.; Wal, T.; Tabrizi, S.; Delgadillo-Correa, M.G.; Johnson, J.P.; Garland, S.; Siba, P.; Upcroft, P. Metronidazole resistance in Trichomonas vaginalis from highland women in Papua New Guinea. Sex. Health 2009, 6, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Kulda, J.; Tachezy, J.; Cerkasovova, A. In vitro induced anaerobic resistance to metronidazole in Trichomonas vaginalis. J. Eukaryot. Microbiol. 1993, 40, 262–269. [Google Scholar] [CrossRef]
- Tachezy, J.; Kulda, J.; Tomkova, E. Aerobic resistance of Trichomonas vaginalis to metronidazole induced in vitro. Parasitology 1993, 106, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Lossick, J.G.; Gorrell, T.E. In vitro susceptibility of Trichomonas vaginalis to metronidazole and treatment outcome in vaginal trichomoniasis. Sex Transm. Dis. 1988, 15, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Malla, N.; Kaul, P.; Sehgal, R.; Gupta, I. The presence of dsRNA virus in Trichomonas vaginalis clinical isolates from symptomatic and asymptomatic Indian women and its correlation with in vitro metronidazole sensitivity. Indian J. Med. Microbiol. 2011, 29, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Snipes, L.G.; Gamard, P.M.; Narcisi, E.M.; Beard, C.B.; Lehmann, T.; Secor, W.E. Molecular epidemiology of metronidazole resistance in a population of Trichomonas vaginalis clinical isolates. J. Clin. Microbiol. 2000, 38, 3004–3009. [Google Scholar] [CrossRef] [Green Version]
- Gordts, B.; De Jonckheere, J.; Kasprzak, W.; Majewska, A.C.; Butzler, J.P. In vitro activity of antiprotozoal drugs against Giardia intestinalis of human origin. Antimicrob. Agents Chemother. 1987, 31, 672–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiori, P.L.; Diaz, N.; Cocco, A.R.; Rappelli, P.; Dessì, D. Association of Trichomonas vaginalis with its symbiont Mycoplasma hominis synergistically upregulates the in vitro proinflammatory response of human monocytes. Sex Transm. Infect. 2013, 89, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, A.; Papaioannou, P.; Magiorkinis, E.; Magana, M.; Ioannidou, V.; Tzanetou, K.; Burriel, A.R.; Tsironi, M.; Chatzipanagiotou, S. Detecting the Diversity of Mycoplasma and Ureaplasma Endosymbionts Hosted by Trichomonas vaginalis Isolates. Front. Microbiol. 2017, 28, 1188. [Google Scholar] [CrossRef] [PubMed]
- Diaz, N.; Dessi, D.; Dessole, S.; Fiori, P.L.; Rappelli, P. Rapid detection of coinfections by Trichomonas vaginalis, Mycoplasma hominis, and Ureaplasma urealiticum by a new multiplex polymerase chain reaction. Diagn. Microbiol. Infect. Dis. 2010, 67, 30–36. [Google Scholar] [CrossRef]
- van Belkum, A.; van der Schee, C.; van der Meijden, W.I.; Verbrugh, H.A.; Sluiters, H.J. A clinical study on the association of Trichomonas vaginalis and Mycoplasma hominis infections in women attending a sexually transmitted disease (STD) outpatient clinic. FEMS Immunol. Med. Microbiol. 2001, 32, 27–32. [Google Scholar] [CrossRef]
- Rappelli, P.; Carta, F.; Delogu, G.; Addis, M.F.; Dessì, D.; Cappuccinelli, P. Mycoplasma hominis and Trichomonas vaginalis symbiosis: Multiplicity of infection and transmissibility of M. hominis to human cells. Arch. Microbiol. 2001, 175, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Butler, S.E.; Augostini, P.; Secor, W.E. Mycoplasma hominis infection of Trichomonas vaginalis in clinical isolates from the United States. Parasitol. Res. 2010, 107, 1023–1027. [Google Scholar] [CrossRef]
- Dessi, D.; Rappelli, P.; Diaz, N.; Cappuccinelli, P.; Fiori, P.L. Mycoplasma hominis and Trichomonas vaginalis: A unique case of symbiotic relationship between two obligate human parasites. Front. Biosci. J. Virtual Libr. 2006, 11, 2028–2034. [Google Scholar] [CrossRef] [Green Version]
- Thi Trung Thu, T.; Margarita, V.; Cocco, A.R.; Marongiu, A.; Dessì, D.; Rappelli, P.; Fiori, P.L. Trichomonas vaginalis Transports Virulent Mycoplasma hominis and Transmits the Infection to Human Cells after Metronidazole Treatment: A Potential Role in Bacterial Invasion of Fetal Membranes and Amniotic Fluid. J. Pregnancy 2018, 2, 5037181. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.C.; Xie, L.F.; Fang, S.L.; Gao, M.Y.; Zhu, Y.; Song, L.Y.; Zhong, H.M.; Lun, Z.R. Symbiosis of Mycoplasma hominis in Trichomonas vaginalis may link metronidazole resistance in vitro. Parasitol. Res. 2006, 100, 123–130. [Google Scholar] [CrossRef]
- Fürnkranz, U.; Henrich, B.; Walochnik, J. Mycoplasma hominis impacts gene expression expression in Trichomonas vaginalis. Parasitol. Res. 2018, 117, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Margarita, V.; Cao, L.C.; Bailey, N.P.; Ngoc, T.H.T.; Ngo, T.M.C.; Nu, P.A.T.; Diaz, N.; Dessì, D.; Hirt, R.P.; Fiori, P.L.; et al. Effect of the Symbiosis with Mycoplasma hominis and Candidatus Mycoplasma Girerdii on Trichomonas vaginalis Metronidazole Susceptibility. Antibiotics 2022, 16, 812. [Google Scholar] [CrossRef] [PubMed]
- Margarita, V.; Bailey, N.P.; Rappelli, P.; Diaz, N.; Fettweis, J.M.; Hirt, R.P.; Fiori, P.L. Two Different Species of Mycoplasma Endosymbionts Can Influence Trichomonas vaginalis Pathophysiology. mBio 2022, 28, e0091822. [Google Scholar] [CrossRef] [PubMed]
- Plummer, E.L.; Vodstrcil, L.A.; Bodiyabadu, K.; Murray, G.L.; Doyle, M.; Latimer, R.L.; Fairley, C.K.; Payne, M.; Chow, E.P.F.; Garland, S.M.; et al. Are Mycoplasma hominis, Ureaplasma urealyticum and Ureaplasma parvum associated with specific genital symptoms and clinical signs in nonpregnant women? Clin. Infect. Dis. 2021, 16, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Arya, O.P.; Tong, C.Y.; Hart, C.A.; Pratt, B.C.; Hughes, S.; Roberts, P.; Kirby, P.; Howel, J.; McCormick, A.; Goddard, A.D. Is mycoplasma hominis a vaginal pathogen? Sex Transm. Infect. 2001, 77, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Horner, P.; Donders, G.; Cusini, M.; Gomberg, M.; Jensen, J.S.; Unemo, M. Should we be testing for urogenital Mycoplasma hominis, Ureaplasma parvum and Ureaplasma urealiticum in men and women?—A position statement from the European STI Guidelines Editorial Board. JEADV 2018, 32, 1845–1852. [Google Scholar]
- Saadat, S.; Karami, P.; Jafari, M.; Kholoujini, M.; Rikhtegaran Tehrani, Z.; Mohammadi, Y.; Alikhani, M.Y. The silent presence of Mycoplasma hominis in patients with prostate cancer. Pathog. Dis. 2020, 78, ftaa037. [Google Scholar] [CrossRef]
- Stary, A.; Kuchinka-Koch, A.; Teodorowicz, L. Detection of Trichomonas vaginalis on Modified Columbia Agar in the Routine Laboratory. J. Clin. Microbiol. 2002, 40, 3277–3280. [Google Scholar] [CrossRef] [Green Version]
- Diamond, L.S. The establishment of various trichomonads of animals and man in axenic cultures. J. Parasitol. 1957, 3, 488–490. [Google Scholar] [CrossRef]
- Goodman, R.P.; Freret, S.; Kula, T.; Geller, A.M.; Talkington, M.W.T.; Tang-Fernandez, V.; Suciu, O.; Demidenko, A.A.; Ghabrial, S.A.; Beach, D.H.; et al. Clinical isolates of Trichomonas vaginalis concurrently infected by strains of up to four Trichomonas virus Species (Familiy Totiviridae). J. Virol. 2011, 85, 4258–4270. [Google Scholar] [CrossRef] [Green Version]
- Van Kuppeveld, F.J.M.; van der Logt, J.T.M.; Angulo, A.F.; van Zoest, M.J.; Quint, G.V.; Niesters, H.G.; Galama, J.M.; Melchers, W.J. Genus- and species-specific identification of Mycoplasma by 16S rRNA amplification. Appl. Environ. Microbiol. 1992, 58, 2606–2615. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Number of Patients with TV | 82 | |
|---|---|---|
| Sex | 72 female (88%) | 10 male (12%) |
| Age (mean age) | 23–64 (41.2) | 34–58 (46) |
| No clinical symptoms reported | 7/72 (9.7%) | 6/10 (60%) |
| No other pathogens diagnosed | 4/72 (5.5%) | 0/10 (0%) |
| Number of Positive Patients and Cultured TV Isolates | 82 | |
|---|---|---|
| Sex | 72 female | 10 male |
| M. hominis in patients (culture) | 27/72 (36.5%) | 3/10 (30%) |
| M. hominis in TV (PCR) | 20/72 (27.8%) | 3/10 (30%) |
| M. hominis in patients and in TV (culture and PCR) | 10/72 (14%) | 2/10 (20%) |
| M. hominis only in patients (culture), not in TV | 17/72 (24%) | 1/10 (10%) |
| M. hominis only in TV (PCR), not in patients (culture) | 10/72 (14%) | 1/10 (10%) |
| TV | Metronidazole Mean Concentration (Min–Max) | |
|---|---|---|
| Anaerobic Conditions | Aerobic Conditions | |
| no TVVs (n = 28) | 0.8 µg/mL (0.5–2 µg/mL) | 3.3 µg/mL (0.5–32 µg/mL) |
| +TVV1 (n = 12) | 0.65 µg/mL (0.5–1 µg/mL) | 3.3 µg/mL (0.5–32 µg/mL) |
| +TVV2 (n = 2) | 0.67 µg/mL (0.5–1 µg/mL) | 5.5 µg/mL (0.5–8 µg/mL) |
| +TVV3 (n = 5) | 0.5 µg/mL (0.5 µg/mL) | 0.93 µg/mL (0.5–2 µg/mL) |
| +TVV1 + TVV2 (n = 3) | 1.13 µg/mL (0.5–2 µg/mL) | 4.16 µg/mL (0.5–8 µg/mL) |
| +TVV1 + TVV3 (n = 2) | 1.17 µg/mL (0.5–2 µg/mL) | 1.5 µg/mL (0.5–2 µg/mL) |
| +M. hominis (n = 12) | 0.63 µg/mL (0.5–1 µg/mL) | 2.58 µg/mL (0.5–32 µg/mL) |
| +M. hominis + TVVs (n = 7) | 0.62 µg/mL (0.5–1 µg/mL) | 4.65 µg/mL (0.5–32 µg/mL) |
| T. vaginalis Virus | Primer Name | Primer Sequence (5′→3′) | Expected Product Size |
|---|---|---|---|
| TVV1 | TVV1F2875 TVV1R3443 | ATTAGCGGTGTTTGTGATGCA CTATCTTGCCATCCTGACTC | 569 bp |
| TVV2 | TVV2F2456 TVV2R3080 | GCTTGAGCACTGCTCGCG TCTCTTTTGGCATCGCTT | 625 bp |
| TVV3 | TVV3F1474 TVV3R2025 | TGGAGTTAAATGGTCTCGAGC GATTTCATCTTGTTCAATTGCA | 552 bp |
| TVV4 | TVV4F1338 TVV4R1834 | ATGCCAGTTGCTTTCCG TTCCCCAATAGTTATCAG | 514 bp |
| Primer Name | Target Gene | Primer Sequence (5′→3′) | Expected Product Size |
|---|---|---|---|
| GPOI MGSO | Mycoplasma genus specific 16S rRNA | ACTCCTACGGGAGGCAGCAGTA TGCACCATCTGTCACTCTGTTAACCTC | 717 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lesiak-Markowicz, I.; Walochnik, J.; Stary, A.; Fürnkranz, U. Characterisation of Trichomonas vaginalis Isolates Collected from Patients in Vienna between 2019 and 2021. Int. J. Mol. Sci. 2022, 23, 12422. https://doi.org/10.3390/ijms232012422
Lesiak-Markowicz I, Walochnik J, Stary A, Fürnkranz U. Characterisation of Trichomonas vaginalis Isolates Collected from Patients in Vienna between 2019 and 2021. International Journal of Molecular Sciences. 2022; 23(20):12422. https://doi.org/10.3390/ijms232012422
Chicago/Turabian StyleLesiak-Markowicz, Iwona, Julia Walochnik, Angelika Stary, and Ursula Fürnkranz. 2022. "Characterisation of Trichomonas vaginalis Isolates Collected from Patients in Vienna between 2019 and 2021" International Journal of Molecular Sciences 23, no. 20: 12422. https://doi.org/10.3390/ijms232012422