
Freshwater Macroalgae, Oedogonium, Grown in Wastewater Reduce Diet-Induced Metabolic Syndrome in Rats

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Composition of Macroalgae from Different Sources
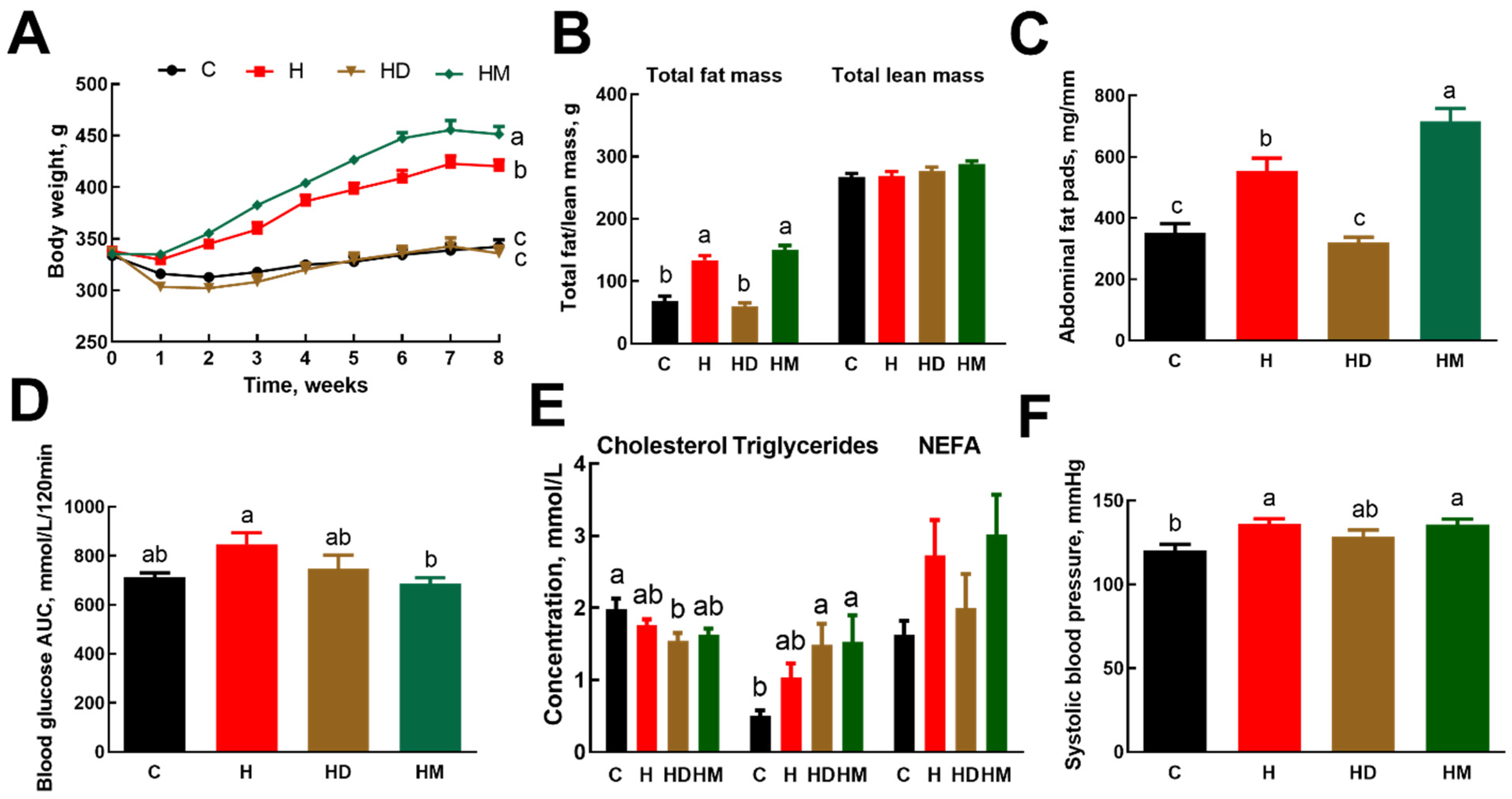
2.2. Metabolic Parameters and Body Composition
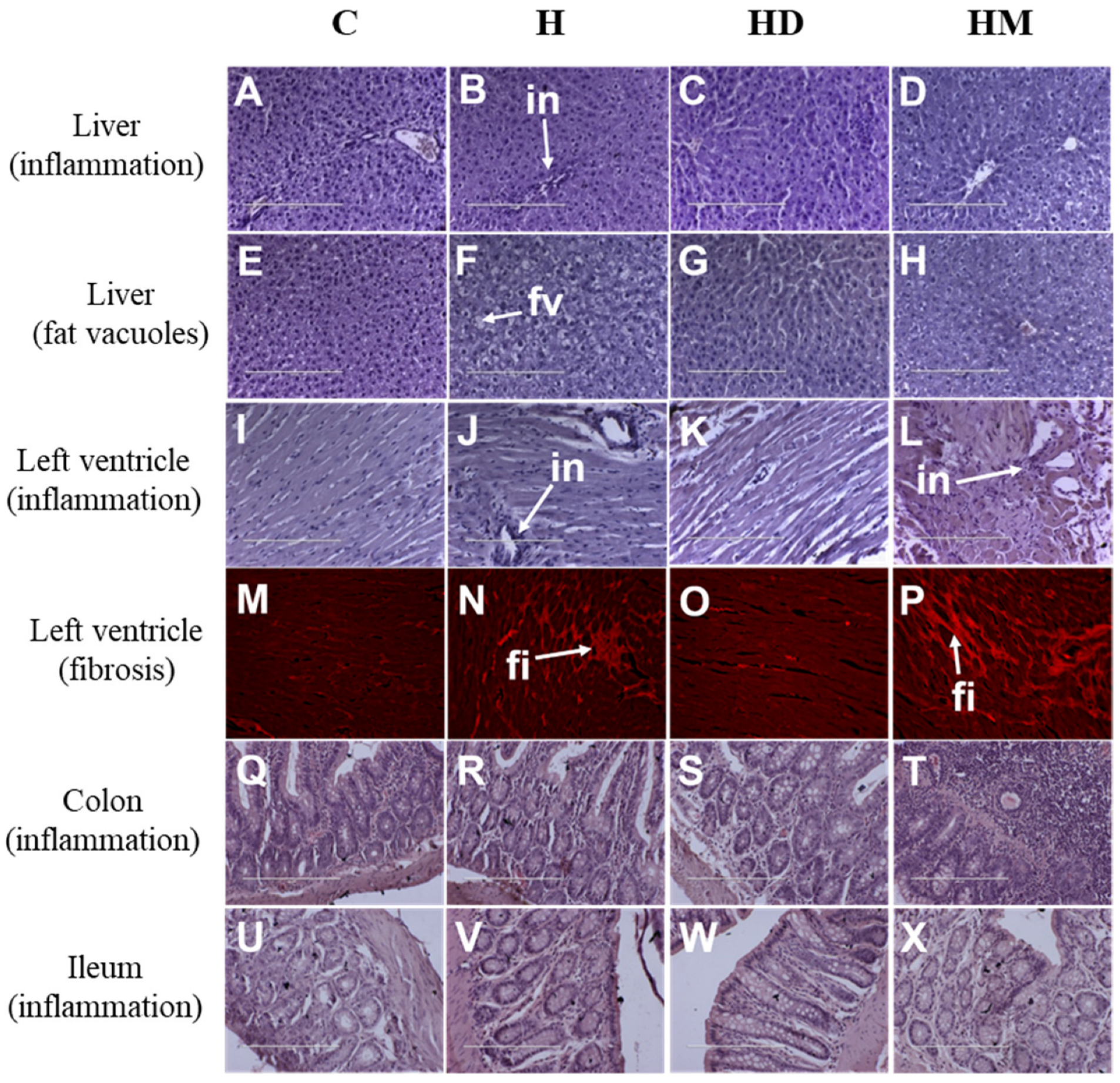
2.3. Liver, Heart and Gastrointestinal Parameters
3. Discussion
4. Materials and Methods
4.1. Sources of Freshwater Macroalgae
4.2. Chemical Analysis of Macroalgae
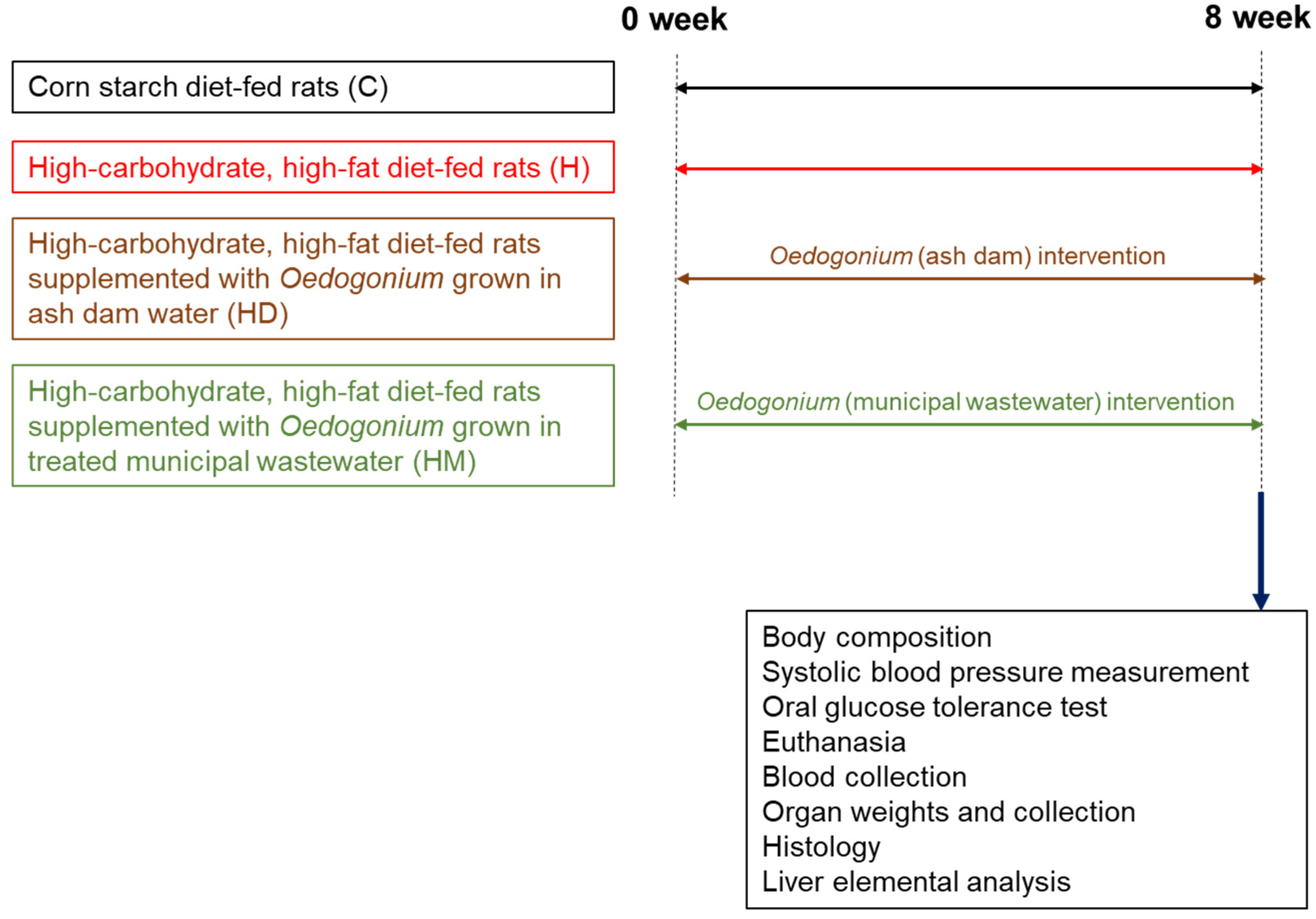
4.3. Studies in Live Rats
4.4. Studies after Euthanasia
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mouritsen, O.G.; Rhatigan, P.; Cornish, M.L.; Critchley, A.T.; Pérez-Lloréns, J.L. Saved by seaweeds: Phyconomic contributions in times of crises. J. Appl. Phycol. 2021, 33, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Lloréns, J.L.; Mouritsen, O.G.; Rhatigan, P.; Cornish, M.L.; Critchley, A.T. Seaweeds in mythology, folklore, poetry, and life. J. Appl. Phycol. 2020, 32, 3157–3182. [Google Scholar] [CrossRef]
- Raja, K.; Kadirvel, V.; Subramaniyan, T. Seaweeds, an aquatic plant-based protein for sustainable nutrition—A review. Future Foods 2022, 5, 100142. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef]
- Granato, D.; Barba, F.J.; Bursać Kovačević, D.; Lorenzo, J.M.; Cruz, A.G.; Putnik, P. Functional foods: Product development, technological trends, efficacy testing, and safety. Annu. Rev. Food Sci. Technol. 2020, 11, 93–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, S.; O’Driscoll, L. Metabolic syndrome: A closer look at the growing epidemic and its associated pathologies. Obes. Rev. 2015, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.A.; Magnusson, M.; Ward, L.C.; Paul, N.A.; Brown, L. Seaweed supplements normalise metabolic, cardiovascular and liver responses in high-carbohydrate, high-fat fed rats. Mar. Drugs 2015, 13, 788–805. [Google Scholar] [CrossRef] [PubMed]
- du Preez, R.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Caulerpa lentillifera (sea grapes) improves cardiovascular and metabolic health of rats with diet-induced metabolic syndrome. Metabolites 2020, 10, 500. [Google Scholar] [CrossRef]
- du Preez, R.; Magnusson, M.; Majzoub, M.E.; Thomas, T.; Praeger, C.; Glasson, C.R.K.; Panchal, S.K.; Brown, L. Brown seaweed Sargassum siliquosum as an intervention for diet-induced obesity in male Wistar rats. Nutrients 2021, 13, 1754. [Google Scholar] [CrossRef]
- du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the red seaweed Sarconema filiforme attenuate symptoms of diet-induced metabolic syndrome in rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanyonyi, S.; du Preez, R.; Brown, L.; Paul, N.A.; Panchal, S.K. Kappaphycus alvarezii as a food supplement prevents diet-induced metabolic syndrome in rats. Nutrients 2017, 9, 1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, O.D.; du Preez, R.; Panchal, S.K.; Brown, L. Tropical foods as functional foods for metabolic syndrome. Food Funct. 2020, 11, 6946–6960. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. The State of the World’s Fisheries and Aquaculture 2020. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; p. 244. Available online: https://www.fao.org/3/ca9229en/ca9229en.pdf (accessed on 22 August 2022).
- Duarte, C.M.; Gattuso, J.-P.; Hancke, K.; Gundersen, H.; Filbee-Dexter, K.; Pedersen, M.F.; Middelburg, J.J.; Burrows, M.T.; Krumhansl, K.A.; Wernberg, T.; et al. Global estimates of the extent and production of macroalgal forests. Glob. Ecol. Biogeogr. 2022, 31, 1422–1439. [Google Scholar] [CrossRef]
- Yun, J.-H.; Smith, V.H.; deNoyelles, F.J.; Roberts, G.W.; Stagg-Williams, S.M. Freshwater macroalgae as a biofuels feedstock: Mini-review and assessment of their bioenergy potential. Indus. Biotechnol. 2014, 10, 212–220. [Google Scholar] [CrossRef]
- Lawton, R.J.; de Nys, R.; Paul, N.A. Selecting reliable and robust freshwater macroalgae for biomass applications. PLoS ONE 2013, 8, e64168. [Google Scholar] [CrossRef]
- Arumugam, N.; Chelliapan, S.; Kamyab, H.; Thirugnana, S.; Othman, N.; Nasri, N.S. Treatment of wastewater using seaweed: A review. Int. J. Environ. Res. Public Health 2018, 15, 2851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deniz, F.; Ersanli, E.T. An ecofriendly approach for bioremediation of contaminated water environment: Potential contribution of a coastal seaweed community to environmental improvement. Int. J. Phytoremediat. 2018, 20, 256–263. [Google Scholar] [CrossRef]
- Kidgell, J.T.; de Nys, R.; Hu, Y.; Paul, N.A.; Roberts, D.A. Bioremediation of a complex industrial effluent by biosorbents derived from freshwater macroalgae. PLoS ONE 2014, 9, e94706. [Google Scholar] [CrossRef] [Green Version]
- Rajasulochana, P.; Preethy, V. Comparison on efficiency of various techniques in treatment of waste and sewage water—A comprehensive review. Resour.-Effic. Technol. 2016, 2, 175–184. [Google Scholar] [CrossRef]
- Han, W.; Clarke, W.; Pratt, S. Composting of waste algae: A review. Waste Manag. 2014, 34, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Michalak, I.; Chojnacka, K.; Saeid, A. Plant growth biostimulants, dietary feed supplements and cosmetics formulated with supercritical CO2 algal extracts. Molecules 2017, 22, 66. [Google Scholar] [CrossRef]
- Neveux, N.; Nugroho, A.A.; Roberts, D.A.; Vucko, M.J.; de Nys, R. Selecting extraction conditions for the production of liquid biostimulants from the freshwater macroalga Oedogonium intermedium. J. Appl. Phycol. 2020, 32, 539–551. [Google Scholar] [CrossRef]
- Roberts, D.A.; Paul, N.A.; Bird, M.I.; de Nys, R. Bioremediation for coal-fired power stations using macroalgae. J. Environ. Manag. 2015, 153, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.J.; Neveux, N.; Whelan, A.; Morton, J.; Vis, M.; de Nys, R.; Paul, N.A. Adding value to the treatment of municipal wastewater through the intensive production of freshwater macroalgae. Algal Res. 2016, 20, 100–109. [Google Scholar] [CrossRef]
- Neveux, N.; Magnusson, M.; Mata, L.; Whelan, A.; de Nys, R.; Paul, N.A. The treatment of municipal wastewater by the macroalga Oedogonium sp. and its potential for the production of biocrude. Algal Res. 2016, 13, 284–292. [Google Scholar] [CrossRef]
- Roberts, D.A.; Shiels, L.; Tickle, J.; de Nys, R.; Paul, N.A. Bioremediation of aluminium from the waste water of a conventional water treatment plant using the freshwater macroalga Oedogonium. Water 2018, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Kraak, V.I.; Allender, S.; Atkins, V.J.; Baker, P.I.; Bogard, J.R.; Brinsden, H.; Calvillo, A.; De Schutter, O.; Devarajan, R.; et al. The global syndemic of obesity, undernutrition, and climate change: The Lancet Commission report. Lancet 2019, 393, 791–846. [Google Scholar] [CrossRef]
- NCD Risk Factor Collaboration (NCD-RisC). Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef]
- Kearney, J. Food consumption trends and drivers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 2793–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, G.A.; Kim, K.K.; Wilding, J.P.H.; World Obesity, F. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunnane, S.C.; Crawford, M.A. Energetic and nutritional constraints on infant brain development: Implications for brain expansion during human evolution. J. Hum. Evol. 2014, 77, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Lange, K.W.; Hauser, J.; Nakamura, Y.; Kanaya, S. Dietary seaweeds and obesity. Food Sci. Hum. Wellness 2015, 4, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Tipnee, S.; Ramaraj, R.; Unpaprom, Y. Nutritional evaluation of edible freshwater green macroalga Spirogyra varians. Emer. Life Sci. Res. 2015, 1, 1–7. [Google Scholar]
- Santos-Marcos, J.A.; Perez-Jimenez, F.; Camargo, A. The role of diet and intestinal microbiota in the development of metabolic syndrome. J. Nutr. Biochem. 2019, 70, 1–27. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Miranda, J.M.; Mondragon, A.D.C.; Lamas, A.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Potential use of marine seaweeds as prebiotics: A review. Molecules 2020, 25, 1004. [Google Scholar] [CrossRef] [Green Version]
- Gotteland, M.; Riveros, K.; Gasaly, N.; Carcamo, C.; Magne, F.; Liabeuf, G.; Beattie, A.; Rosenfeld, S. The pros and cons of using algal polysaccharides as prebiotics. Front. Nutr. 2020, 7, 163. [Google Scholar] [CrossRef]
- Arata, P.X.; Alberghina, J.; Confalonieri, V.; Errea, M.I.; Estevez, J.M.; Ciancia, M. Sulfated polysaccharides in the freshwater green macroalga Cladophora surera not linked to salinity adaptation. Front. Plant Sci. 2017, 8, 1927. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, P.T.; Misra, S.R.; Hussain, M. Nutritional aspects of essential trace elements in oral health and disease: An extensive review. Scientifica 2016, 2016, 5464373. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Pan, X.D.; Huang, B.F.; Han, J.L. Distribution of metals and metalloids in dried seaweeds and health risk to population in southeastern China. Sci. Rep. 2018, 8, 3578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamshad, I.; Khan, S.; Waqas, M.; Asma, M.; Nawab, J.; Gul, N.; Raiz, A.; Li, G. Heavy metal uptake capacity of fresh water algae (Oedogonium westti) from aqueous solution: A mesocosm research. Int. J. Phytoremediat. 2016, 18, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Xie, S.; Feng, J.; Li, Y.; Chen, L. Heavy metal uptake capacities by the common freshwater green alga Cladophora fracta. J. Appl. Phycol. 2012, 24, 979–983. [Google Scholar] [CrossRef]
- Lewicki, S.; Zdanowski, R.; Krzyzowska, M.; Lewicka, A.; Debski, B.; Niemcewicz, M.; Goniewicz, M. The role of chromium III in the organism and its possible use in diabetes and obesity treatment. Ann. Agric. Environ. Med. 2014, 21, 331–335. [Google Scholar] [CrossRef]
- Rehder, D. The future of/for vanadium. Dalton Trans. 2013, 42, 11749–11761. [Google Scholar] [CrossRef]
- Rehder, D. The role of vanadium in biology. Metallomics 2015, 7, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Tinkov, A.A.; Popova, E.V.; Polyakova, V.S.; Kwan, O.V.; Skalny, A.V.; Nikonorov, A.A. Adipose tissue chromium and vanadium disbalance in high-fat fed Wistar rats. J. Trace Elem. Med. Biol. 2015, 29, 176–181. [Google Scholar] [CrossRef]
- Tinkov, A.A.; Sinitskii, A.I.; Popova, E.V.; Nemereshina, O.N.; Gatiatulina, E.R.; Skalnaya, M.G.; Skalny, A.V.; Nikonorov, A.A. Alteration of local adipose tissue trace element homeostasis as a possible mechanism of obesity-related insulin resistance. Med. Hypotheses 2015, 85, 343–347. [Google Scholar] [CrossRef]
- Chimienti, F. Zinc, pancreatic islet cell function and diabetes: New insights into an old story. Nutr. Res. Rev. 2013, 26, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Koliaki, C.; Katsilambros, N. Dietary sodium, potassium, and alcohol: Key players in the pathophysiology, prevention, and treatment of human hypertension. Nutr. Rev. 2013, 71, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Wanyonyi, S.; Brown, L. Selenium, vanadium, and chromium as micronutrients to improve metabolic syndrome. Curr. Hypertens. Rep. 2017, 19, 10. [Google Scholar] [CrossRef] [PubMed]
- Tuzcu, M.; Sahin, N.; Orhan, C.; Agca, C.A.; Akdemir, F.; Tuzcu, Z.; Komorowski, J.; Sahin, K. Impact of chromium histidinate on high fat diet induced obesity in rats. Nutr. Metab. 2011, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexanian, I.; Parissis, J.; Farmakis, D.; Pantziou, C.; Ikonomidis, I.; Paraskevaidis, I.; Ioannidou, S.; Sideris, A.; Kremastinos, D.; Lekakis, J.; et al. Selenium contributes to myocardial injury and cardiac remodeling in heart failure. Int. J. Cardiol. 2014, 176, 272–273. [Google Scholar] [CrossRef]
- Chaabane, M.; Tir, M.; Hamdi, S.; Boudawara, O.; Jamoussi, K.; Boudawara, T.; Ghorbel, R.E.; Zeghal, N.; Soudani, N. Improvement of heart redox states contributes to the beneficial effects of selenium against penconazole-induced cardiotoxicity in adult rats. Biol. Trace Elem. Res. 2016, 169, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Metelo, A.M.; Perez-Carro, R.; Castro, M.M.; Lopez-Larrubia, P. VO(dmpp)2 normalizes pre-diabetic parameters as assessed by in vivo magnetic resonance imaging and spectroscopy. J. Inorg. Biochem. 2012, 115, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Youn, C.K.; Hyun, J.W.; You, H.J. The anti-obesity effect of natural vanadium-containing Jeju ground water. Biol. Trace Elem. Res. 2013, 151, 294–300. [Google Scholar] [CrossRef]
- Nigra, A.E.; Olmedo, P.; Grau-Perez, M.; O’Leary, R.; O’Leary, M.; Fretts, A.M.; Umans, J.G.; Best, L.G.; Francesconi, K.A.; Goessler, W.; et al. Dietary determinants of inorganic arsenic exposure in the Strong Heart Family Study. Environ. Res. 2019, 177, 108616. [Google Scholar] [CrossRef]
- Spratlen, M.J.; Grau-Perez, M.; Best, L.G.; Yracheta, J.; Lazo, M.; Vaidya, D.; Balakrishnan, P.; Gamble, M.V.; Francesconi, K.A.; Goessler, W.; et al. The association of arsenic exposure and arsenic metabolism with the metabolic syndrome and its individual components: Prospective evidence from the Strong Heart Family Study. Am. J. Epidemiol. 2018, 187, 1598–1612. [Google Scholar] [CrossRef] [Green Version]
- Moon, K.A.; Oberoi, S.; Barchowsky, A.; Chen, Y.; Guallar, E.; Nachman, K.E.; Rahman, M.; Sohel, N.; D’Ippoliti, D.; Wade, T.J.; et al. A dose-response meta-analysis of chronic arsenic exposure and incident cardiovascular disease. Int. J. Epidemiol. 2018, 47, 1013. [Google Scholar] [CrossRef]
- Khairul, I.; Wang, Q.Q.; Jiang, Y.H.; Wang, C.; Naranmandura, H. Metabolism, toxicity and anticancer activities of arsenic compounds. Oncotarget 2017, 8, 23905–23926. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.; Goodale, B.; Raab, A.; Schwerdtle, T.; Reimer, K.; Conklin, S.; Karagas, M.R.; Francesconi, K.A. Human exposure to organic arsenic species from seafood. Sci. Total Environ. 2017, 580, 266–282. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Pal, N.; Sharma, P.; Kumawat, M.; Sarma, D.K.; Nabi, B.; Verma, V.; Tiwari, R.R.; Shubham, S.; Arjmandi, B.; et al. Omega-3 fatty acids and their interaction with the gut microbiome in the prevention and amelioration of type-2 diabetes. Nutrients 2022, 14, 1723. [Google Scholar] [CrossRef] [PubMed]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Rivera-Madrid, R.; Carballo-Uicab, V.M.; Cárdenas-Conejo, Y.; Aguilar-Espinosa, M.; Siva, R. Overview of carotenoids and beneficial effects on human health. In Carotenoids: Properties, Processing and Applications; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–40. [Google Scholar] [CrossRef]
- Lawton, R.J. Oedogonium sp. Tsv2 Internal Transcribed Spacer 1, Partial Sequence; 5.8S Ribosomal RNA Gene, Complete Sequence; and Internal Transcribed Spacer 2, Partial Sequence. GenBank: KF606977.1. 2014. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KF606977 (accessed on 14 October 2022).
- Lawton, R.J.; de Nys, R.; Skinner, S.; Paul, N.A. Isolation and identification of Oedogonium species and strains for biomass applications. PLoS ONE 2014, 9, e90223. [Google Scholar] [CrossRef] [Green Version]
- Lawton, R.J. Oedogonium sp. Tar1 Internal Transcribed Spacer 1, Partial Sequence; 5.8S Ribosomal RNA Gene, Complete Sequence; and Internal Transcribed Spacer 2, Partial Sequence. GenBank: KF606974.1. 2014. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KF606974 (accessed on 14 October 2022).
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Gosch, B.J.; Magnusson, M.; Paul, N.A.; de Nys, R. Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012, 4, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Channiwala, S.A.; Parikh, P.P. A unified correlation for estimating HHV of solid, liquid and gaseous fuels. Fuel 2002, 81, 1051–1063. [Google Scholar] [CrossRef]
- Saunders, R.J.; Paul, N.A.; Hu, Y.; de Nys, R. Sustainable sources of biomass for bioremediation of heavy metals in waste water derived from coal-fired power generation. PLoS ONE 2012, 7, e36470. [Google Scholar] [CrossRef]
- Ritchie, R.J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Weast, R.C. CRC—Handbook of Chemistry and Physics, 56th ed.; Chemical Rubber Publishing Co.: Cleveland, OH, USA, 1975–1976. [Google Scholar]
- Poudyal, H.; Panchal, S.K.; Waanders, J.; Ward, L.; Brown, L. Lipid redistribution by α-linolenic acid-rich chia seed inhibits stearoyl-CoA desaturase-1 and induces cardiac and hepatic protection in diet-induced obese rats. J. Nutr. Biochem. 2012, 23, 153–162. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Oedogonium Cultivated in Ash Dam Water (D) | Oedogonium Cultivated in Treated Municipal Wastewater (M) | |
|---|---|---|
| Elements and proximate composition (% dry weight) | ||
| C | 32.41 ± 0.29 | 38.90 ± 0.05 |
| H | 5.31 ± 0.14 | 6.09 ± 0.01 |
| O | 19.64 ± 0.024 | 24.03 ± 1.68 |
| N | 5.15 ± 0.02 | 5.37 ± 0.11 |
| S | 0.33 ± 0.04 | 0.30 ± 0.015 |
| F | 0.035 ± 0.0002 | 0.0006 ± 0.0 |
| Cl | 0.209 ± 0.002 | 0.814 ± 0.012 |
| Br | 0.0032 ± 0.0001 | 0.0021 ± 0.00005 |
| I | 0.005 ± 0.0001 | <0.0016 |
| Moisture | 5.16 ± 0.35 | 6.41 ± 1.38 |
| Ash | 32.00 ± 0.06 | 18.90 ± 0.20 |
| Protein (sum of amino acids) | 21.44 | 23.87 |
| Total lipids | 3.40 ± 0.25 | 4.82 ± 0.24 |
| Carbohydrate (by difference) | 37.99 | 46.01 |
| Metals, non-metals and metalloids (mg/kg dry weight) | ||
| Al | 1315.0 ± 35.4 | 276.5 ± 2.1 |
| As | 99.95 ± 1.49 | 1.26 ± 0.014 |
| B | 9.61 ± 0.16 | 7.63 ± 0.09 |
| Ba | 209.5 ± 0.71 | 77.85 ± 0.50 |
| Ca | 9010 ± 141 | 14,450 ± 212 |
| Cd | 2.20 ± 0.02 | 0.11 ± 0.009 |
| Co | 5.75 ± 0.099 | 0.84 ± 0.008 |
| Cr | 16.35 ± 0.071 | 1.32 ± 0.04 |
| Cu | 48.70 ± 0.85 | 13.35 ± 0.21 |
| Fe | 14,500 ± 141 | 423.5 ± 6.4 |
| Hg | <0.5 | <0.5 |
| K | 10,550 ± 71 | 28,400 ± 1839 |
| Mg | 2485 ± 35 | 6165 ± 35 |
| Mn | 737.0 ± 9.9 | 184.0 ± 2.8 |
| Mo | 10.10 ± 0.00 | 0.82 ± 0.38 |
| Na | 1550 ± 14 | 1990 ± 14 |
| Ni | 42.60 ± 0.57 | 2.05 ± 0.02 |
| P | 10,900 ± 0 | 13,900 ± 141 |
| Pb | 0.83 ± 0.016 | 2.15 ± 0.134 |
| S | 3975 ± 219 | 3080 ± 141 |
| Se | 13.60 ± 0.28 | <1 |
| Sr | 124.50 ± 0.71 | 180.5 ± 0.71 |
| V | 723.0 ± 4.24 | 0.82 ± 0.02 |
| Zn | 258.0 ± 4.24 | 42.15 ± 1.91 |
| Fatty acids (% of total fatty acids) | ||
| C14:0 | 9.08 ± 0.35 | 0.91 ± 0.01 |
| C15:0 | 0.44 ± 0.02 | 0.12 ± 0.01 |
| C16:0 | 50.84 ± 0.27 | 26.50 ± 0.09 |
| C16:1n-7 | 22.90 ± 0.38 | 4.69 ± 0.13 |
| C16:2n-6 | 0.11 ± 0.06 | 0.94 ± 0.04 |
| C17:0 | 0.26 ± 0.09 | 0.32 ± 0.04 |
| C16:3n-3 | 0.59 ± 0.38 | 18.28 ± 0.30 |
| C18:0 | 1.23 ± 0.04 | 0.80 ± 0.01 |
| C18:1n-9 | 5.24 ± 0.21 | 1.75 ± 0.04 |
| C18:2n-6 | 1.90 ± 0.07 | 4.59 ± 0.04 |
| C18:3n-6 | ND | 2.97 ± 0.10 |
| C18:3n-3 | 5.63 ± 0.05 | 30.82 ± 0.18 |
| C18:4n-3 | ND | 3.81 ± 0.05 |
| C20:1n-9 | 0.62 ± 0.05 | ND |
| C20:2n-6 | 1.20 ± 0.18 | 0.45 ± 0.10 |
| C20:3n-6 | ND | 1.99 ± 0.07 |
| C22:0 | ND | 1.05 ± 0.06 |
| Total SFA | 61.85 ± 0.54 | 29.71 ± 0.06 |
| Total MUFA | 28.75 ± 0.41 | 6.44 ± 0.12 |
| Total PUFA | 9.40 ± 0.49 | 63.85 ± 0.17 |
| Total n-3 PUFA | 6.22 ± 0.35 | 52.90 ± 0.40 |
| Total n-6 PUFA | 3.18 ± 0.26 | 10.95 ± 0.26 |
| n-3:n-6 | 5.63 ± 0.05 | 30.82 ± 0.18 |
| Dietary fibre (% dry weight) | ||
| Total dietary fibre | 23.8 | 35.3 |
| Insoluble fibre | 21.4 | 34.5 |
| Vitamins | ||
| α-carotene (μg/100 g) | <5 | <5 |
| Ascorbic acid (mg/100 g) | <1 | <1 |
| β-carotene (μg/100 g) | 320 | 6400 |
| Vitamin K1 (μg/100 g) | 37 | 12 |
| Vitamin B12 (μg/100 g) | 218 | 634 |
| Chlorophyll (% dry weight) | ||
| Chlorophyll a | 0.20 ± 0.01 | 0.59 ± 0.01 |
| Chlorophyll b | 0.50 ± 0.01 | 1.43 ± 0.04 |
| Amino acids (mg/g dry weight) | ||
| Histidine | 3.7 | 4.6 |
| Serine | 10.5 | 12.3 |
| Arginine | 14.5 | 14.3 |
| Glycine | 12.6 | 13.0 |
| Aspartic acid | 24.8 | 28.7 |
| Glutamic acid | 28.2 | 32.1 |
| Threonine | 11.9 | 13.2 |
| Alanine | 15.1 | 16.5 |
| Proline | 10.3 | 12.0 |
| Lysine | 12.9 | 15.0 |
| Tyrosine | 8.3 | 9.6 |
| Methionine | 4.0 | 4.1 |
| Valine | 14.3 | 15.9 |
| Isoleucine | 10.9 | 11.3 |
| Leucine | 19.9 | 21.9 |
| Phenylalanine | 12.5 | 14.2 |
| Variables | C | H | HD | HM |
|---|---|---|---|---|
| Body weight gain, g | 17.0 ± 7.0 c | 81.0 ± 5.0 b | −1.0 ± 5.6 c | 124.0 ± 6.6 a |
| Food intake, g/day | 36.7 ± 1.3 a | 21.7 ± 0.4 b | 22.8 ± 1.5 b | 22.6 ± 0.4 b |
| Water intake, g/day | 28.9 ± 2.5 | 20.3 ± 1.6 | 24.2 ± 1.8 | 26.4 ± 1.3 |
| Energy intake, kJ/day | 411.6 ± 14.6 b | 465.7 ± 7.6 a | 515.6 ± 28.5 a | 524.8 ± 10.0 a |
| Feed efficiency, g/kJ | 0.04 ± 0.02 c | 0.17 ± 0.01 b | 0.00 ± 0.01 d | 0.24 ± 0.01 a |
| Abdominal circumference, cm | 18.2 ± 0.3 c | 21.1 ± 0.2 b | 18.6 ± 0.3 c | 22.4 ± 0.2 a |
| Bone mineral density, g/cm2 | 0.163 ± 0.003 ab | 0.170 ± 0.004 a | 0.158 ± 0.003 b | 0.172 ± 0.002 a |
| Bone mineral content, g | 10.0 ± 0.3 b | 11.8 ± 0.3 a | 9.3 ± 0.3 b | 12.6 ± 0.2 a |
| Basal blood glucose concentrations (week 8), mmol/L | 4.1 ± 0.3 b | 5.2 ± 0.2 a | 4.5 ± 0.2 b | 4.3 ± 0.1 b |
| Blood glucose 120 min (week 8), mmol/L | 4.7 ± 0.2 b | 6.2 ± 0.4 a | 4.7 ± 0.4 b | 4.5 ± 0.2 b |
| Retroperitoneal fat, mg/mm | 154.0 ± 13.3 c | 253.8 ± 26.7 b | 130.1 ± 10.6 c | 338.9 ± 24.9 a |
| Epididymal fat, mg/mm | 89.7 ± 8.1 c | 131.2 ± 10.5 b | 70.4 ± 3.6 c | 182.5 ± 15.3 a |
| Omental fat, mg/mm | 108.3 ± 11.2 b | 169.6 ± 8.4 a | 120.6 ± 9.1 b | 194.4 ± 9.9 a |
| Left ventricle + septum, mg/mm | 18.6 ± 0.7 b | 20.0 ± 0.8 ab | 19.2 ± 0.9 b | 22.1 ± 0.7 a |
| Right ventricle, mg/mm | 4.25 ± 0.19 | 4.08 ± 0.46 | 3.40 ± 0.22 | 4.38 ± 0.32 |
| Liver, mg/mm | 223 ± 10 b | 279 ± 4 a | 323 ± 19 a | 323 ±13 a |
| Kidneys, mg/mm | 47.5 ± 1.8 c | 55.2 ± 1.1 b | 58.4 ± 3.2 ab | 63.6 ± 1.8 a |
| Spleen, mg/mm | 14.6 ± 0.7 b | 16.9 ± 0.9 b | 15.9 ± 0.6 b | 20.2 ± 1.0 a |
| Plasma alanine transaminase activity, U/L | 31.5 ± 6.6 | 39.5 ± 3.2 | 47.0 ± 3.0 | 31.0 ± 2.0 |
| Plasma aspartate transaminase activity, U/L | 82.0 ± 11.2 | 75.5 ± 6.8 | 72.0 ± 3.0 | 65.0 ± 4.0 |
| Metals (mg/kg Dry Weight) | C | H | HD | HM |
|---|---|---|---|---|
| Al | 1.99 ± 0.89 | <0.5 | 0.53 ± 0.24 | 1.93 ± 0.86 |
| As | 4.94 ± 2.21 | 2.60 ± 1.16 | 17.34 ± 7.76 | 2.53 ± 1.13 |
| B | 28.20 ± 12.61 a | 0.24 ± 0.11 b | 11.62 ± 5.20 ab | 4.36 ± 1.95 ab |
| Ba | <0.1 | 0.08 ± 0.03 | 0.06 ± 0.03 | 0.11 ± 0.05 |
| Ca | 135 ± 60 | 129 ± 58 | 108 ± 48 | 108 ± 48 |
| Cd | <0.05 | <0.05 | <0.05 | <0.05 |
| Co | 0.05 ± 0.02 | <0.1 | 0.21 ± 0.10 | 0.07 ± 0.03 |
| Cr | 0.25 ± 0.11 | 0.18 ± 0.08 | 0.16 ± 0.07 | 0.14 ± 0.06 |
| Cu | 15.24 ± 6.82 | 16.55 ± 7.40 | 11.92 ± 5.33 | 13.92 ± 6.23 |
| Fe | 781 ± 349 | 543 ± 243 | 762 ± 341 | 526 ± 235 |
| Hg | 2.67 ± 1.19 | 1.75 ± 0.78 | 1.62 ± 0.72 | 1.81 ± 0.81 |
| K | 10,378 ± 4641 | 10,416 ± 4658 | 8622 ± 3856 | 8197 ± 3666 |
| Mg | 717 ± 321 | 714 ± 319 | 611 ± 273 | 655 ± 293 |
| Mn | 6.24 ± 2.79 | 5.49 ± 2.46 | 5.57 ± 2.49 | 6.37 ± 2.85 |
| Mo | 0.87 ± 0.39 | 1.03 ± 0.46 | 1.00 ± 0.45 | 0.96 ± 0.43 |
| Na | 3052 ± 1365 | 2800 ± 1252 | 1373 ± 614 | 1143 ± 511 |
| Ni | 0.08 ± 0.03 | 0.05 ± 0.02 | 0.08 ± 0.04 | 0.07 ± 0.03 |
| P | 9048 ± 4046 | 9342 ± 4178 | 7026 ± 3142 | 7590 ± 3394 |
| Pb | 0.08 ± 0.04 | 0.07 ± 0.03 | 0.07 ± 0.03 | 0.07 ± 0.03 |
| S | 5232 ± 2340 | 5146 ± 2301 | 4060 ± 1816 | 4218 ± 1886 |
| Se | 1.76 ± 0.79 | 1.81 ± 0.81 | 2.99 ± 1.34 | 2.27 ±1.01 |
| Sr | 0.12 ± 0.05 | 0.18 ± 0.08 | 0.22 ± 0.10 | 0.19 ± 0.08 |
| V | 0.06 ± 0.03 b | 0.09 ± 0.04 b | 4.07 ± 1.82 a | 0.18 ± 0.08 b |
| Zn | 66.5 ± 29.7 | 69.7 ± 31.2 | 57.3 ± 25.6 | 66.3 ± 29.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panchal, S.K.; Ghattamaneni, N.K.R.; Magnusson, M.; Cole, A.; Roberts, D.; Neveux, N.; Brown, L.; Paul, N.A. Freshwater Macroalgae, Oedogonium, Grown in Wastewater Reduce Diet-Induced Metabolic Syndrome in Rats. Int. J. Mol. Sci. 2022, 23, 13811. https://doi.org/10.3390/ijms232213811
Panchal SK, Ghattamaneni NKR, Magnusson M, Cole A, Roberts D, Neveux N, Brown L, Paul NA. Freshwater Macroalgae, Oedogonium, Grown in Wastewater Reduce Diet-Induced Metabolic Syndrome in Rats. International Journal of Molecular Sciences. 2022; 23(22):13811. https://doi.org/10.3390/ijms232213811
Chicago/Turabian StylePanchal, Sunil K., Naga K. R. Ghattamaneni, Marie Magnusson, Andrew Cole, David Roberts, Nicolas Neveux, Lindsay Brown, and Nicholas A. Paul. 2022. "Freshwater Macroalgae, Oedogonium, Grown in Wastewater Reduce Diet-Induced Metabolic Syndrome in Rats" International Journal of Molecular Sciences 23, no. 22: 13811. https://doi.org/10.3390/ijms232213811