

Detection of the Omicron SARS-CoV-2 Lineage and Its BA.1 Variant with Multiplex RT-qPCR
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
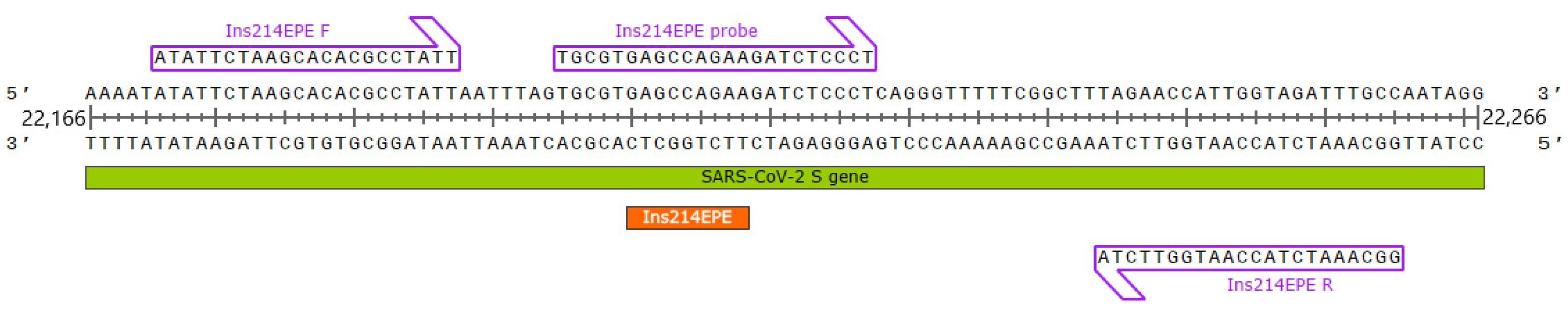
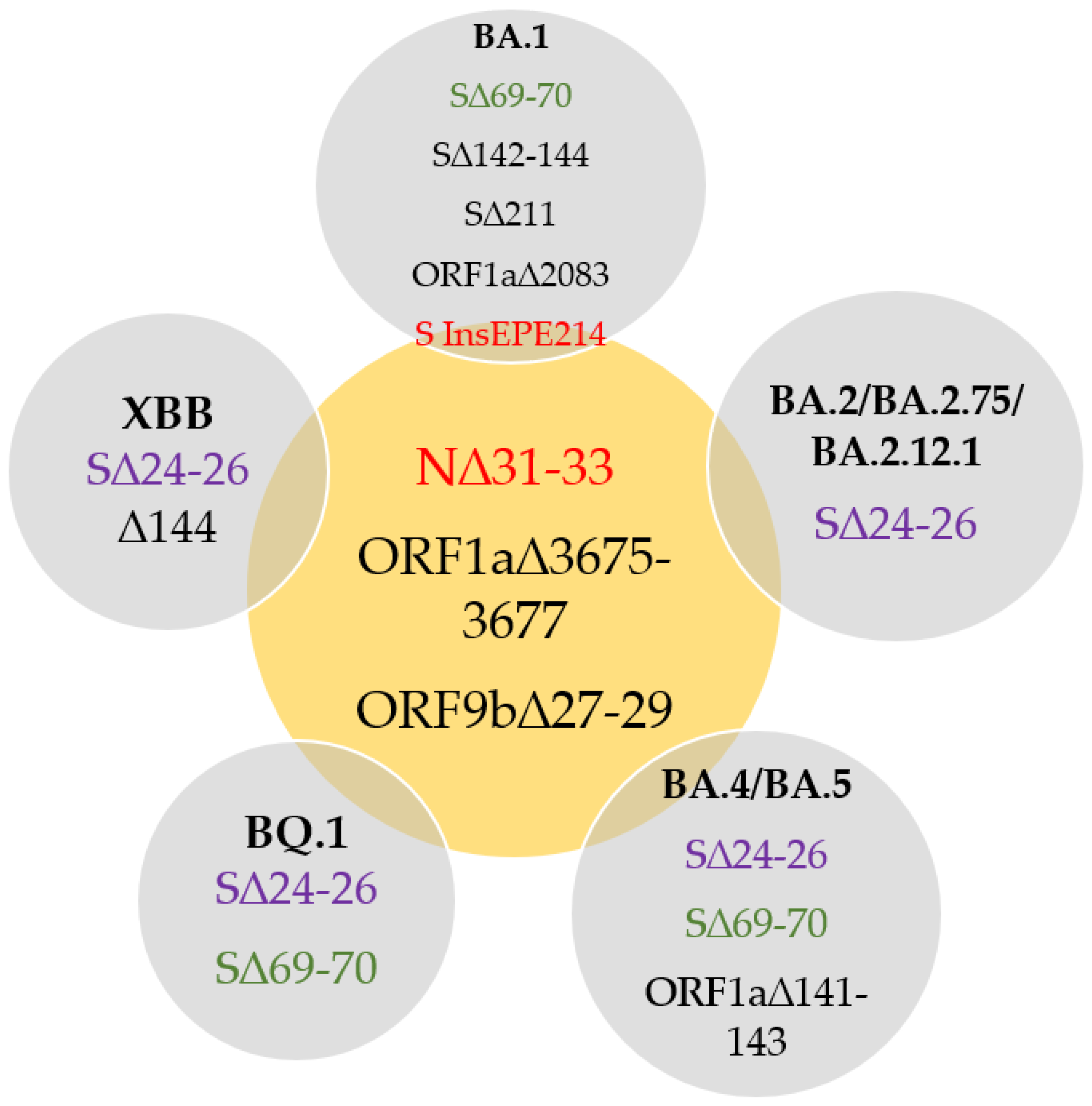
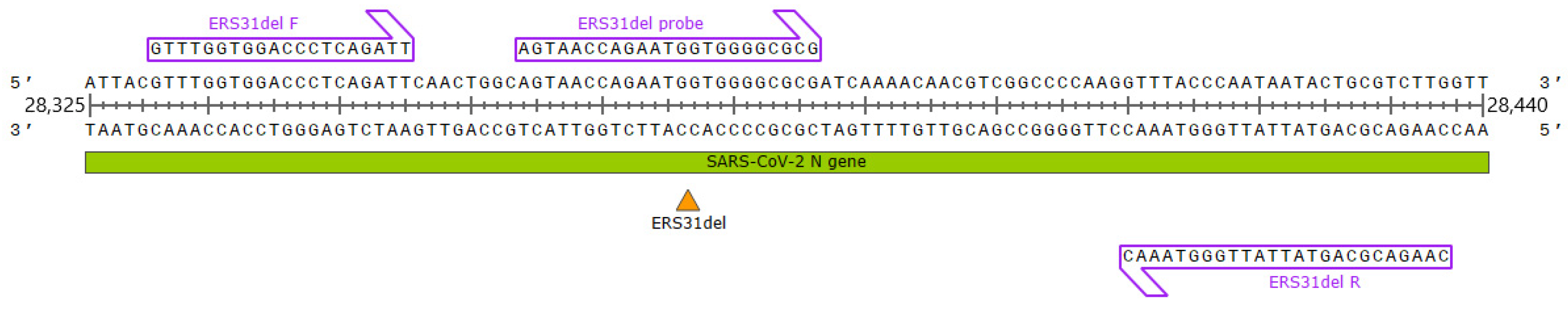
2.1. Primer and Probe Design
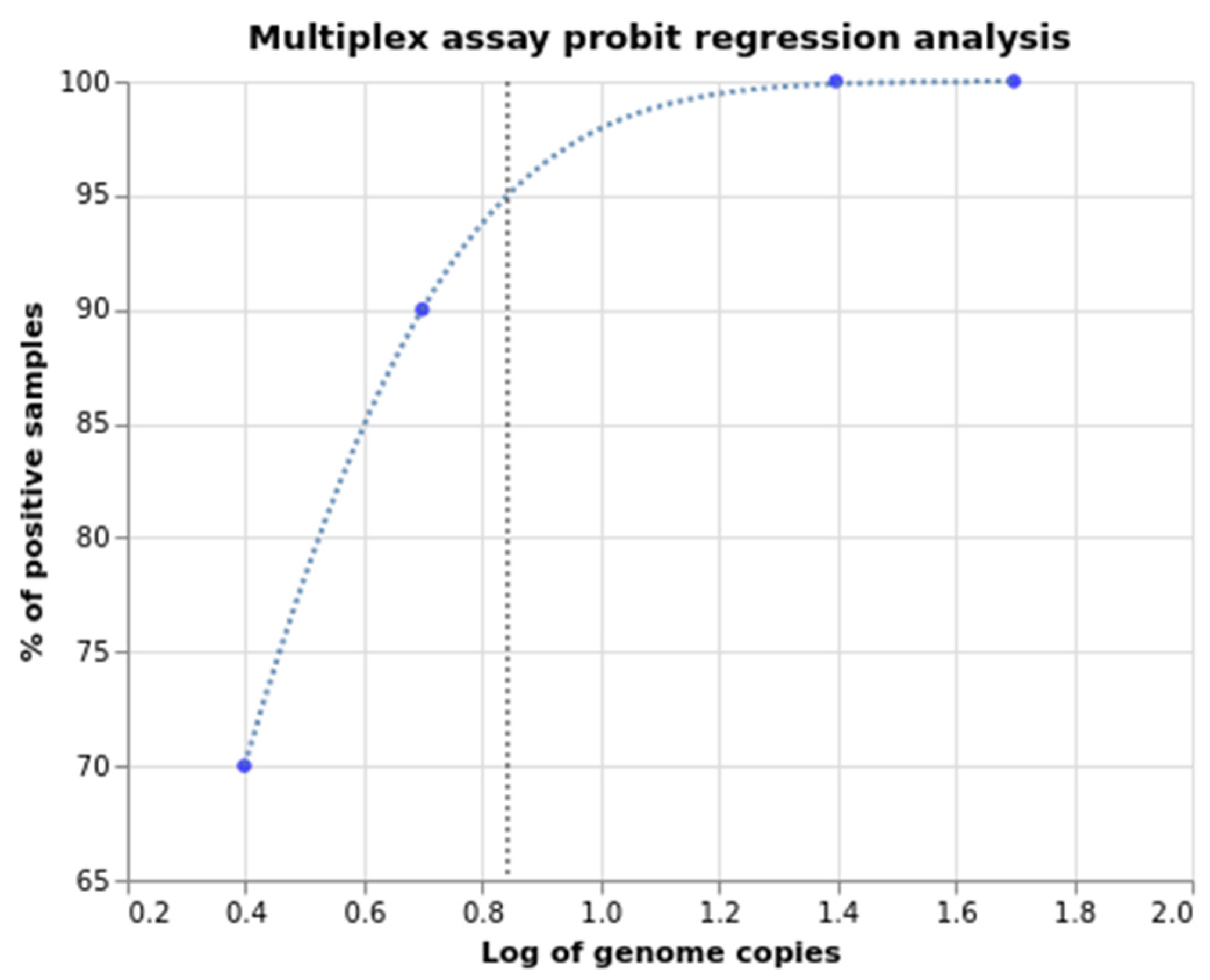
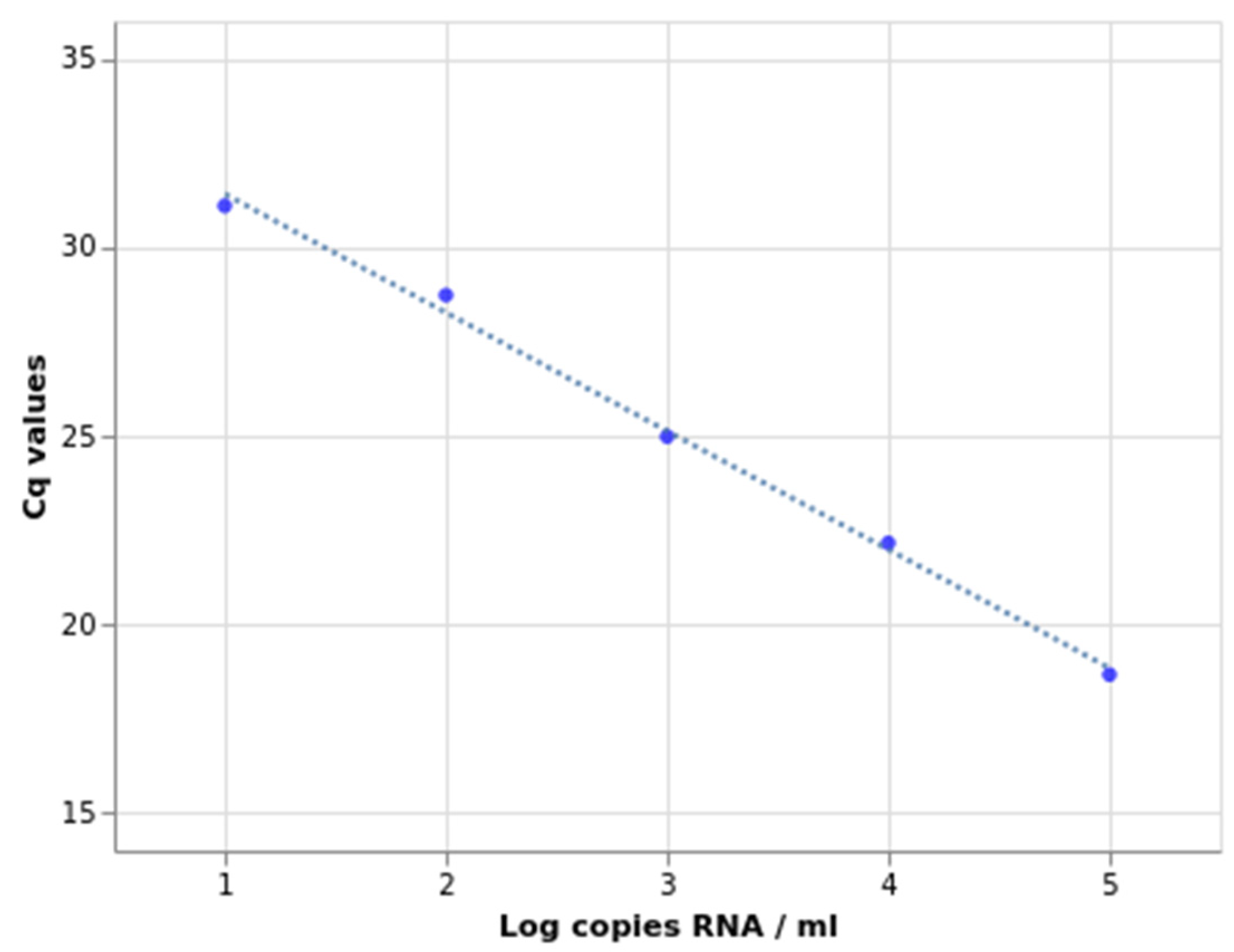
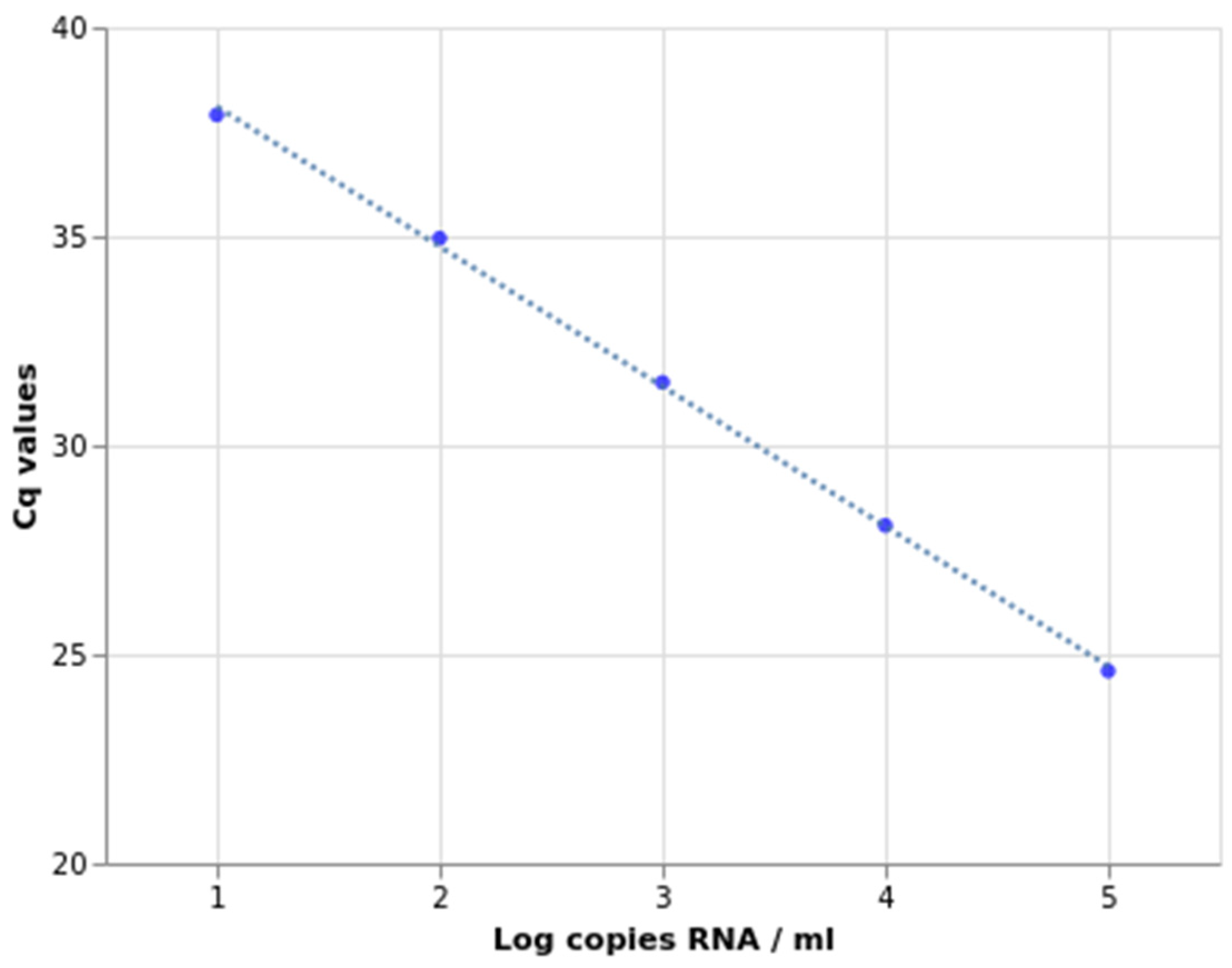
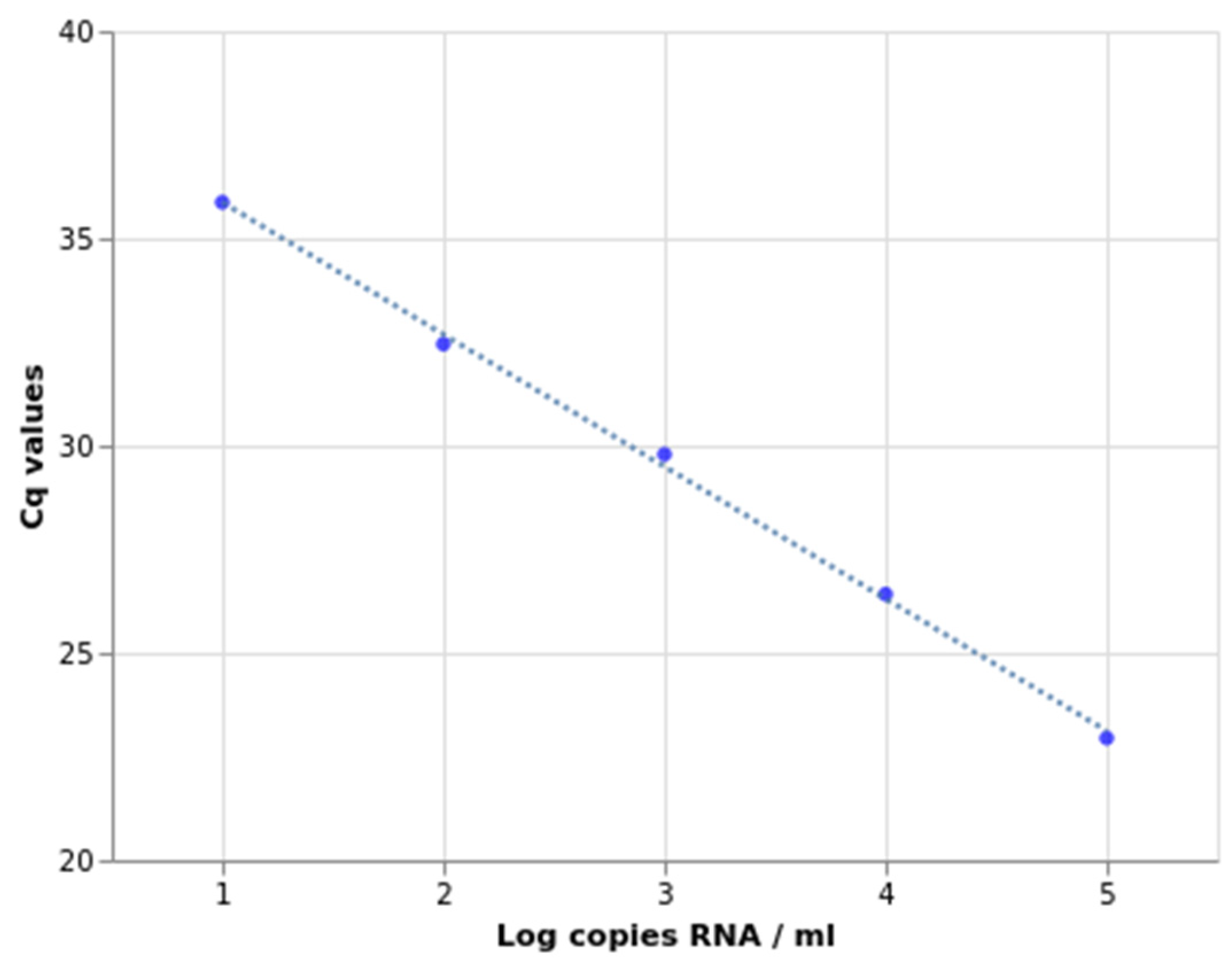
2.2. Sensitivity, Specificity, and Amplification Efficiency Assessment
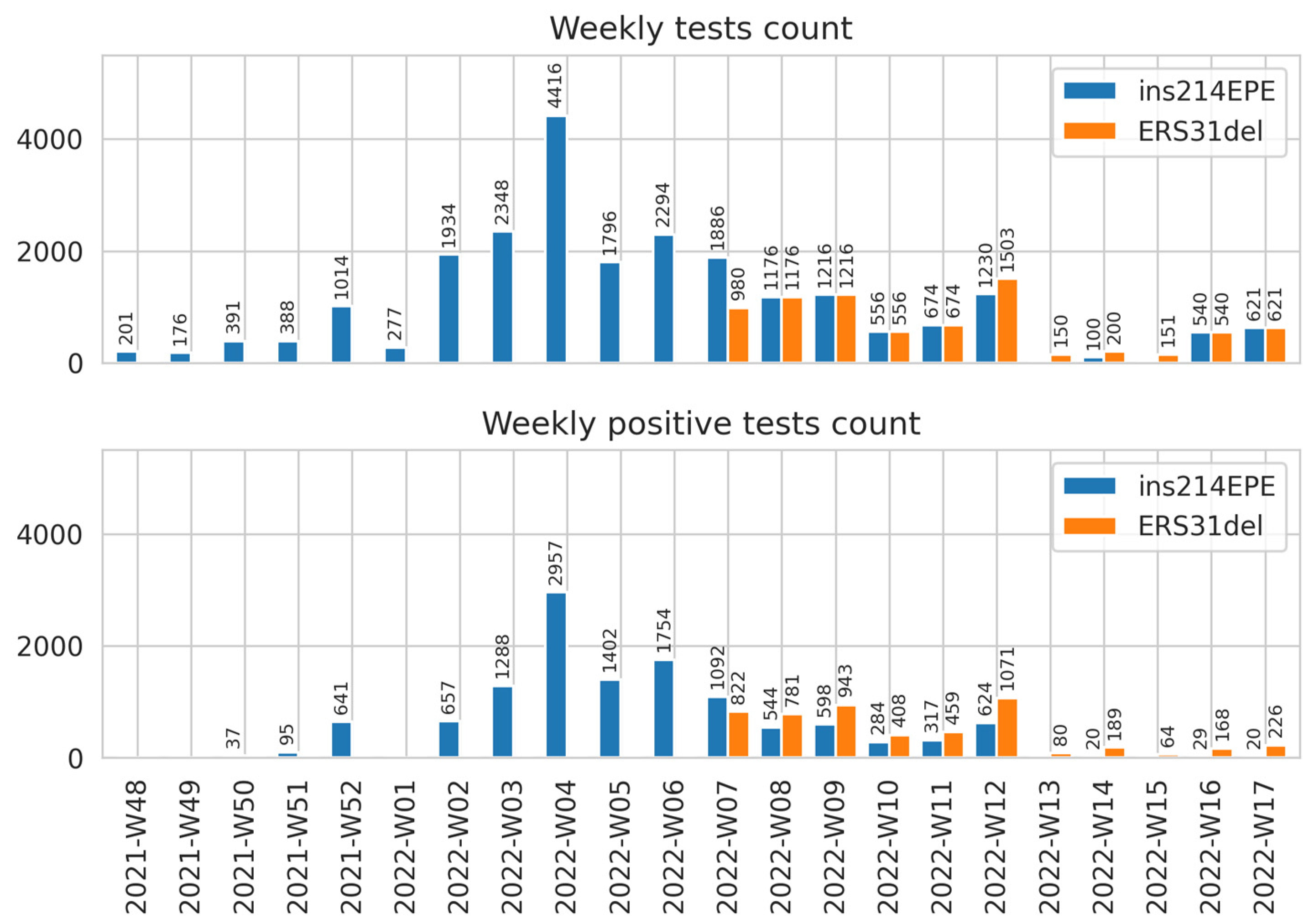
2.3. Implementing the Developed Assays in SARS-CoV-2 Surveillance in Russia
3. Discussion
4. Materials and Methods
4.1. Sample Collection and RT-PCR Testing
4.2. RT-qPCR Assay Design
4.3. Digital PCR
4.4. Genome Assembly and Consensus Correction
4.5. Molecular Cloning
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GISAID Accession Number | Pango Variant | 2-SARS-CoV-2-ORF1 Assay | ERS31del Assay | Ins214EPE Assay | RP Assay |
|---|---|---|---|---|---|
| EPI_ISL_15919776 | CL.1 | 27.65 | 26.25 | Nd | 29.26 |
| EPI_ISL_15919773 | CL.1 | 27.22 | 25.7 | Nd | 26.96 |
| EPI_ISL_15919771 | BA.5.2.6 | 25.02 | 24.91 | Nd | 30.1 |
| EPI_ISL_15919779 | CL.1 | 20.7 | 19.25 | Nd | 31.08 |
| EPI_ISL_15919777 | BA.5.2.6 | 24.69 | 24.82 | Nd | 30.52 |
| EPI_ISL_15919787 | CL.1 | 24.14 | 22.65 | Nd | 26.27 |
| EPI_ISL_15919784 | BA.5.2.30 | 27.03 | 26.92 | Nd | 29.38 |
| EPI_ISL_15919785 | BA.5.2 | 24.98 | 24.8 | Nd | 30.39 |
| EPI_ISL_15919782 | BA.5.2 | 19.87 | 19.64 | Nd | 29.05 |
| EPI_ISL_15919780 | CL.1 | 23.6 | 22.37 | Nd | 25.07 |
| EPI_ISL_15919781 | BA.5.2.1 | 24.3 | 24.19 | Nd | 25.02 |
| EPI_ISL_15920157 | BA.5.2 | 19.94 | 19.64 | Nd | 27.27 |
| EPI_ISL_15920158 | BA.5.2 | 21.35 | 21.39 | Nd | 27.96 |
| EPI_ISL_15920173 | XBB | 24.77 | 23.9 | Nd | 29 |
| EPI_ISL_15920176 | BF.5 | 29.73 | 29.58 | Nd | 32.5 |
| EPI_ISL_15920178 | BA.5.2 | 20.43 | 20.64 | Nd | 26.32 |
| EPI_ISL_15920160 | BA.5.1.10 | 20.05 | 18.63 | Nd | 24.2 |
| EPI_ISL_15920162 | BA.5.2.7 | 20.36 | 20.17 | Nd | 24.92 |
| EPI_ISL_15920168 | BA.5.2 | 22.41 | 22.29 | Nd | 25.2 |
| EPI_ISL_15983536 | BA.5.2.6 | 18.46 | 18.1 | Nd | 25.53 |
| EPI_ISL_15983633 | BA.5.2 | 25.08 | 25.13 | Nd | 26.2 |
| EPI_ISL_15983624 | BA.5.2 | 24.73 | 24.63 | Nd | 30.28 |
| EPI_ISL_15983623 | BA.5.2 | 26.6 | 26.42 | Nd | 30.2 |
| EPI_ISL_15983622 | CL.1 | 24.58 | 23.24 | Nd | 26.31 |
| EPI_ISL_15983538 | BA.5.2.1 | 25.32 | 31.43 | Nd | 25.43 |
| EPI_ISL_15983599 | BA.5.2 | 26.34 | 26.27 | Nd | 25.38 |
| EPI_ISL_15983534 | BA.5.2 | 21.01 | 20.9 | Nd | 28.43 |
| EPI_ISL_15983596 | BA.5.2 | 25.85 | 25.81 | Nd | 29.7 |
| EPI_ISL_15983595 | BA.5.2.6 | 27.58 | 27.43 | Nd | 28.65 |
| EPI_ISL_15983592 | CL.1 | 23.87 | 22.3 | Nd | 25.03 |
| EPI_ISL_15983594 | BA.5.2.34 | 33.04 | 33.94 | Nd | 29.34 |
| EPI_ISL_15983593 | CL.1 | 28.83 | 27.65 | Nd | 30.17 |
| EPI_ISL_11503185 | BA.1 | 24.3 | 22.85 | 25.22 | 30.12 |
| EPI_ISL_11503189 | BA.1 | 25.21 | 23.68 | 26.03 | 31.63 |
| EPI_ISL_12043917 | BA.1 | 27.04 | 25.69 | 27.62 | 35.07 |
| EPI_ISL_12043919 | BA.1 | 28.5 | 27.15 | 28.84 | 30.65 |
| EPI_ISL_12043923 | BA.1 | 30.77 | 29.45 | 30.97 | 29.68 |
| EPI_ISL_12043930 | BA.1 | 27.65 | 26.27 | 28.02 | 29.08 |
| EPI_ISL_9338420 | BA.1 | 18.02 | 16.31 | 18.28 | 31.63 |
| EPI_ISL_9338432 | BA.1 | 22.56 | 21.24 | 22.76 | 23.9 |
| EPI_ISL_9338433 | BA.1 | 28.78 | 27.19 | 28.55 | 28 |
| EPI_ISL_9338434 | BA.1 | 24.09 | 22.81 | 24.26 | 26.93 |
| EPI_ISL_9338498 | BA.1 | 27.18 | 25.81 | 27.46 | 27.17 |
| EPI_ISL_9338506 | BA.1 | 21.35 | 20.04 | 22.46 | 25.32 |
| EPI_ISL_9338534 | BA.1 | 34.62 | 34.07 | 35.38 | 29.87 |
| EPI_ISL_9338538 | BA.1 | 24.18 | 22.71 | 24.77 | 27.53 |
| EPI_ISL_9338588 | BA.1 | 22.2 | 20.73 | 22.27 | 30.59 |
| EPI_ISL_9338592 | BA.1 | 19.45 | 17.85 | 19.39 | 27.42 |
| EPI_ISL_9338686 | BA.1 | 29.07 | 28.01 | 28.92 | 27.9 |
| EPI_ISL_9338701 | BA.1 | 24.26 | 22.93 | 24.27 | 27.95 |
| EPI_ISL_9338747 | BA.1 | 28.14 | 26.79 | 27.98 | 26.34 |
| EPI_ISL_9338818 | BA.1 | 21.28 | 20.07 | 21.12 | 24.44 |
| EPI_ISL_9861967 | BA.1 | 26.74 | 25.4 | 26.52 | 28.24 |
| EPI_ISL_9862115 | BA.1 | 27.63 | 26.48 | 27.48 | 25.9 |
| EPI_ISL_9862127 | BA.1 | 27.31 | 25.99 | 27.7 | 29.04 |
| EPI_ISL_9862130 | BA.1 | 30.94 | 29.62 | 31.25 | 34.54 |
| EPI_ISL_9862144 | BA.1 | 25.8 | 24.26 | 26.15 | 28.42 |
| EPI_ISL_9862153 | BA.1 | 23.95 | 22.57 | 23.96 | 29.04 |
| EPI_ISL_9862158 | BA.1 | 22.84 | 21.5 | 22.89 | 27.67 |
| EPI_ISL_9862159 | BA.1 | 29.65 | 28.33 | 29.47 | 33.28 |
| EPI_ISL_9862161 | BA.1 | 26.05 | 24.59 | 26.1 | 29.05 |
| EPI_ISL_9862162 | BA.1 | 26.56 | 25.16 | 26.85 | 28.19 |
| EPI_ISL_9862172 | BA.1 | 23.32 | 21.82 | 23.25 | 27.38 |
| EPI_ISL_9862177 | BA.1 | 23.36 | 22.17 | 22.91 | 25.54 |
| EPI_ISL_3122915 | B.1.617.2 | 24.87 | Nd | Nd | 26.64 |
| EPI_ISL_3122916 | B.1.617.2 | 33.94 | Nd | Nd | 28.28 |
| EPI_ISL_3122917 | AY.122 | 25.77 | Nd | Nd | 29.62 |
| EPI_ISL_3122923 | AY.122 | 25.72 | Nd | Nd | 27.69 |
| EPI_ISL_3122927 | AY.122 | 26.36 | Nd | Nd | 27.12 |
| EPI_ISL_3122928 | AY.122 | 22.99 | Nd | Nd | 26.28 |
| EPI_ISL_3122931 | AY.122 | 30.06 | Nd | Nd | 29.54 |
| EPI_ISL_3122933 | AY.122 | 22.81 | Nd | Nd | 26.56 |
| EPI_ISL_3122971 | AY.122 | 31.27 | Nd | Nd | 26.36 |
| EPI_ISL_3122972 | AY.122 | 29.13 | Nd | Nd | 32.05 |
| EPI_ISL_3122973 | AY.122 | 23.03 | Nd | Nd | 27.8 |
| EPI_ISL_3122974 | AY.122 | 30.12 | Nd | Nd | 28.77 |
| EPI_ISL_3122975 | B.1.617.2 | 26.55 | Nd | Nd | 24.21 |
| EPI_ISL_3122976 | AY.122 | 36.43 | Nd | Nd | 28.2 |
| EPI_ISL_3122977 | B.1.1.7 | 28.96 | Nd | Nd | 28.26 |
| EPI_ISL_3122978 | AY.122 | 30.76 | Nd | Nd | 31.7 |
| EPI_ISL_3122979 | AY.122 | 34.45 | Nd | Nd | 32.33 |
| EPI_ISL_3122980 | AY.122 | 25.22 | Nd | Nd | 28.57 |
| EPI_ISL_3122981 | AY.122 | 32.53 | Nd | Nd | 26.87 |
| EPI_ISL_3122982 | B.1.617.2 | 31.29 | Nd | Nd | 26.91 |
| EPI_ISL_3122986 | AY.122 | 22.96 | Nd | Nd | 29.73 |
| EPI_ISL_3122987 | AY.122 | 23.33 | Nd | Nd | 26.07 |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
| Negative control | - | Nd | Nd | Nd | Nd |
Appendix B



Appendix C
| Clinical Sample Positive for | RP Assay, Cq | SARS-CoV-2 Assay, Cq | Ins214EPE, Cq |
|---|---|---|---|
| RSV A | 28.76 | Nd | Nd |
| RSV A | 30.56 | Nd | Nd |
| RSV A | 27.70 | Nd | Nd |
| RSV B | 31.49 | Nd | Nd |
| RSV B | 30.98 | Nd | Nd |
| RSV B | 32.33 | Nd | Nd |
| NL63 | 32.20 | Nd | Nd |
| NL63 | 30.42 | Nd | Nd |
| NL63 | 24.95 | Nd | Nd |
| Oc43 | 30.34 | Nd | Nd |
| Oc43 | 30.69 | Nd | Nd |
| Oc43 | 28.64 | Nd | Nd |
| HKU1 | 30.06 | Nd | Nd |
| HKU1 | 28.30 | Nd | Nd |
| HKU1 | 30.73 | Nd | Nd |
| 229E | 29.11 | Nd | Nd |
| 229E | 32.52 | Nd | Nd |
| 229E | 29.37 | Nd | Nd |
| BoV | 32.26 | Nd | Nd |
| BoV | 30.75 | Nd | Nd |
| BoV | 27.25 | Nd | Nd |
| Rv | 32.85 | Nd | Nd |
| Rv | 33.76 | Nd | Nd |
| Rv | 27.75 | Nd | Nd |
| Piv1 | 28.63 | Nd | Nd |
| Piv2 | 24.72 | Nd | Nd |
| Piv3 | 27.01 | Nd | Nd |
| Piv4 | 23.90 | Nd | Nd |
| Adv | 29.47 | Nd | Nd |
| MPV | 30.12 | Nd | Nd |
| HIV A | 29.13 | Nd | Nd |
| HIV A | 28.45 | Nd | Nd |
| HIV A | 28.16 | 39.06 | Nd |
| SARS-CoV-2 B.1.1.529 | 34.15 | 26.61 | 28.44 |
Appendix D
| Multiplex RT-qPCR Results | ||||
| dilution | 2-SARS-CoV-2 | ERS31del | Ins214EPE | RP |
| 1x | 19.01 | 21.00 | 19.38 | 29.38 |
| 10x | 22.22 | 24.14 | 22.44 | 33.19 |
| 100x | 25.56 | 27.78 | 26.13 | 34.51 |
| 1000x | 28.96 | 31.05 | 29.37 | 37.74 |
| 10,000x | 31.44 | 34.04 | 32.25 | Nd |
| 100,000x | 34.55 | 37.69 | 35.52 | Nd |
| Singleplex assay results | ||||
| 2-SARS-CoV-2 | ERS31del | Ins214EPE | RP | |
| 1x | 19.03 | 19.47 | 18.94 | 29.46 |
| 10x | 22.04 | 22.95 | 22.61 | 33.15 |
| 100x | 25.34 | 26.42 | 26.01 | 36.11 |
| 1000x | 28.44 | 29.79 | 29.32 | 38.42 |
| 10,000x | 31.49 | 32.45 | 31.87 | Nd |
| 100,000x | 34.16 | 35.87 | 35.64 | Nd |
References
- Worldometers. Available online: https://www.worldometers.info/coronavirus/ (accessed on 12 November 2022).
- WHO. Weekly Epidemiological Update on COVID-19. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 12 November 2022).
- Mistry, P.; Barmania, F.; Mellet, J.; Peta, K.; Strydom, A.; Viljoen, I.M.; James, W.; Gordon, S.; Pepper, M.S. SARS-CoV-2 Variants, Vaccines, and Host Immunity. Front Immunol. 2022, 12, 809244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiao, T.; Cai, Y.; Lavine, C.L.; Peng, H.; Zhu, H.; Anand, K.; Tong, P.; Gautam, A.; Mayer, M.L.; et al. Membrane fusion and immune evasion by the spike protein of SARS-CoV-2 Delta variant. Science 2021, 374, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Gobeil, S.M.-C.; Janowska, K.; McDowell, S.; Mansouri, K.; Parks, R.; Manne, K.; Stalls, V.; Kopp, M.F.; Henderson, R.; Edwards, R.J.; et al. D614G Mutation Alters SARS-CoV-2 Spike Conformation and Enhances Protease Cleavage at the S1/S2 Junction. Cell Rep. 2021, 34, 108630. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef]
- Schmidt, F.; Muecksch, F.; Weisblum, Y.; Da Silva, J.; Bednarski, E.; Cho, A.; Wang, Z.; Gaebler, C.; Caskey, M.; Nussenzweig, M.C.; et al. Plasma Neutralization of the SARS-CoV-2 Omicron Variant. N. Engl. J. Med. 2022, 386, 599–601. [Google Scholar] [CrossRef]
- Lu, L.; Mok, B.W.Y.; Chen, L.L.; Chan, J.M.C.; Tsang, O.T.Y.; Lam, B.H.S.; Chuang, V.W.M.; Chu, A.W.H.; Chan, W.M.; Ip, J.D.; et al. Neutralization of Severe Acute Respiratory Syndrome Coronavirus 2 Omicron Variant by Sera From BNT162b2 or CoronaVac Vaccine Recipients. Clin. Infect. Dis. 2021, 75, e822–e826. [Google Scholar] [CrossRef]
- Shu, Y.; McCauley, J. GISAID: Global initiative on sharing all influenza data—From vision to reality. Eurosurveillance 2017, 22, 30494. [Google Scholar] [CrossRef] [Green Version]
- Knyazev, S.; Chhugani, K.; Sarwal, V.; Ayyala, R.; Singh, H.; Karthikeyan, S.; Deshpande, D.; Baykal, P.I.; Comarova, Z.; Lu, A.; et al. Unlocking capacities of genomics for the COVID-19 response and future pandemics. Nat. Methods 2022, 19, 374–380. [Google Scholar] [CrossRef]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Ferguson, N.M.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; et al. Transmission of SARS-CoV-2 lineage B.1.1.7 in England: Insights from linking epidemiological and genetic data. MedRxiv 2021. [Google Scholar] [CrossRef]
- Subramoney, K.; Mtileni, N.; Bharuthram, A.; Davis, A.; Kalenga, B.; Rikhotso, M.; Maphahlele, M.; Giandhari, J.; Naidoo, Y.; Pillay, S.; et al. Identification of SARS-CoV-2 Omicron variant using spike gene target failure and genotyping assays, Gauteng, South Africa, 2021. J. Med. Virol. 2022, 94, 3676–3684. [Google Scholar] [CrossRef]
- Kalthoff, D.; Beer, M.; Hoffmann, B. High resolution melting analysis: Rapid and precise characterisation of recombinant influenza A genomes. Virol. J. 2013, 10, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alisoltani, A.; Jaroszewski, L.; Iyer, M.; Iranzadeh, A.; Godzik, A. Increased Frequency of Indels in Hypervariable Regions of SARS-CoV-2 Proteins—A Possible Signature of Adaptive Selection. Front. Genet. 2022, 13, 875406. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M.; Dishnica, K.; Giorgetti, A. Emergence of a recurrent insertion in the N-terminal domain of the SARS-CoV-2 spike glycoprotein. Virus Res. 2022, 310, 198674. [Google Scholar] [CrossRef] [PubMed]
- Garushyants, S.K.; Rogozin, I.B.; Koonin, E.V. Template switching and duplications in SARS-CoV-2 genomes give rise to insertion variants that merit monitoring. Commun. Biol. 2021, 4, 1343. [Google Scholar] [CrossRef]
- Albers, C.A.; Lunter, G.; MacArthur, D.G.; McVean, G.; Ouwehand, W.H.; Durbin, R. Dindel: Accurate indel calls from short-read data. Genome Res. 2011, 21, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Yolshin, N.; Varchenko, K.; Komissarova, K.; Danilenko, D.; Komissarov, A.; Lioznov, D. One-step RT-PCR Ins214EPE assay for Omicron (B.1.1.529) variant detection. Protocols.Io 2021. [Google Scholar] [CrossRef]
- Yolshin, N.; Varchenko, K.; Komissarov, A. SARS-CoV-2 Omicron detection RT-qPCR assay with BA.1 and BA.2/BA.3 differentitation. Protocols.Io 2022. [Google Scholar] [CrossRef]
- Harper, H.; Burridge, A.; Winfield, M.; Finn, A.; Davidson, A.; Matthews, D.; Hutchings, S.; Vipond, B.; Jain, N.; The COVID-19 Genomics UK (COG-UK) Consortium; et al. Detecting SARS-CoV-2 variants with SNP genotyping. PLoS ONE 2022, 16, e0243185. [Google Scholar] [CrossRef]
- Gene-Quantification. Available online: https://www.gene-quantification.de/ab-hrm-guide.pdf (accessed on 12 November 2022).
- Corbisier, P.; Petrillo, M.; Marchini, A.; Querci, M.; Buttinger, G.; Bekliz, M.; Spiess, K.; Polacek, C.; Fomsgaard, A.; Eede, G.V.D. A qualitative RT-PCR assay for the specific identification of the SARS-CoV-2 B.1.1.529 (Omicron) Variant of Concern. J. Clin. Virol. 2022, 152, 105191. [Google Scholar] [CrossRef]
- Bloemen, M.; Rector, A.; Swinnen, J.; Van Ranst, M.; Maes, P.; Vanmechelen, B.; Wollants, E. Fast detection of SARS-CoV-2 variants including Omicron using one-step RT-PCR and Sanger sequencing. J. Virol. Methods 2022, 304, 114512. [Google Scholar] [CrossRef]
- Phan, T.; Boes, S.; McCullough, M.; Gribschaw, J.; Marsh, J.; Harrison, L.H.; Wells, A. Development of a One-Step Qualitative RT-PCR Assay to Detect the SARS-CoV-2 Omicron (B.1.1.529) Variant in Respiratory Specimens. J. Clin. Microbiol. 2022, 60, e0002422. [Google Scholar] [CrossRef] [PubMed]
- Chassalevris, T.; Chaintoutis, S.C.; Koureas, M.; Petala, M.; Moutou, E.; Beta, C.; Kyritsi, M.; Hadjichristodoulou, C.; Kostoglou, M.; Karapantsios, T.; et al. SARS-CoV-2 wastewater monitoring using a novel PCR-based method rapidly captured the Delta-to-Omicron ΒA.1 transition patterns in the absence of conventional surveillance evidence. Sci. Total Environ. 2022, 844, 156932. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Pan, Y.; Cheng, S.M.S.; Hui, K.P.Y.; Krishnan, P.; Liu, Y.; Ng, D.Y.M.; Wan, C.K.C.; Yang, P.; Wang, Q.; et al. Molecular Diagnosis of a Novel Coronavirus (2019-nCoV) Causing an Outbreak of Pneumonia. Clin. Chem. 2020, 66, 549–555. [Google Scholar] [CrossRef] [Green Version]
- CDC. Available online: https://www.cdc.gov/coronavirus/2019-ncov/lab/rt-pcr-panel-primer-probes.html (accessed on 12 November 2022).
- Rodríguez, A.; Rodríguez, M.; Córdoba, J.J.; Andrade, M.J. Design of primers and probes for quantitative real-time PCR methods. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: New York, NY, USA, 2015; Volume 1275, pp. 31–56. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| Copies per Reaction | Log | % Positives | Signal Ratio |
|---|---|---|---|
| 50 | 1.699 | 100 | 20/20 |
| 25 | 1.398 | 100 | 20/20 |
| 5 | 0.699 | 90 | 18/20 |
| 2.5 | 0.398 | 70 | 14/20 |
| LOD (C95) = 0.84 log | LOD (C95) = 7 copies per reaction | ||
| GISAID # | PANGO Lineage | RP Assay, Cq | SARS-CoV-2 Assay, Cq | ERS31del Assay, Cq | Ins214EPE Assay, Cq |
|---|---|---|---|---|---|
| P.1 | 35.35 | 15.45 | Nd | Nd | |
| EPI_ISL_1257814 | B.1.351 | 39.91 | Nd | Nd | Nd |
| EPI_ISL_415710 | B.1 | Nd | 15.69 | Nd | Nd |
| EPI_ISL_1652610 | B.1 | 33.84 | 26.69 | Nd | Nd |
| EPI_ISL_1919527 | B.1 | 24.26 | 26.98 | Nd | Nd |
| EPI_ISL_2698439 | B.1.1.7 | 25.27 | 20.64 | Nd | Nd |
| EPI_ISL_2523457 | B.1.1.7 | 30.54 | 28.05 | Nd | Nd |
| EPI_ISL_2450515 | B.1.1.7 | 26.99 | 31.25 | Nd | Nd |
| EPI_ISL_3454801 | AT.1 | 30.30 | 33.07 | Nd | Nd |
| EPI_ISL_3454797 | AT.1 | 30.02 | 31.33 | Nd | Nd |
| EPI_ISL_3454782 | AT.1 | 28.00 | 33.00 | Nd | Nd |
| EPI_ISL_2698481 | B.1.1.523 | 29.12 | 27.09 | Nd | Nd |
| EPI_ISL_2698509 | B.1.1.523 | 27.39 | 23.46 | Nd | Nd |
| EPI_ISL_2523540 | B.1.1.523 | 30.02 | 27.09 | Nd | Nd |
| EPI_ISL_2698496 | B.1.1.317 | 25.22 | 25.31 | Nd | Nd |
| EPI_ISL_2523528 | B.1.1.317 | 27.58 | 22.71 | Nd | Nd |
| EPI_ISL_2450468 | B.1.1.317 | 28.70 | 23.91 | Nd | Nd |
| EPI_ISL_4563892 | B.1.617.2 | 26.66 | 34.03 | Nd | Nd |
| EPI_ISL_5263415 | B.1.617.2 | 25.72 | 28.43 | Nd | Nd |
| EPI_ISL_4563887 | AY.129 | 26.78 | 25.42 | Nd | Nd |
| EPI_ISL_6831894 | AY.122 | 23.96 | 28.95 | Nd | Nd |
| EPI_ISL_6831897 | AY.122 | 31.16 | 35.05 | Nd | Nd |
| EPI_ISL_6831903 | AY.122 | 24.19 | 34.93 | Nd | Nd |
| EPI_ISL_11503033 | BA.1 | 27.98 | 22.20 | 23.57 | 22.08 |
| EPI_ISL_10931393 | BA.1 | 27.08 | 24.93 | 27.16 | 24.65 |
| EPI_ISL_10931401 | BA.1 | 25.19 | 27.48 | 29.83 | 25.57 |
| EPI_ISL_9338305 | BA.2 | 26.50 | 19.64 | 21.07 | Nd |
| EPI_ISL_9338650 | BA.2 | 24.70 | 20.81 | 22.70 | Nd |
| Omicron BA* Non-BA.1 (32) | Omicron BA.1 (32) | Non-Omicron (22) | NTC (10) | |
|---|---|---|---|---|
| 2-SARS-CoV-2 ORF1 assay | True positive (32) False negative (0) | True positive (32) False negative (0) | True positive (22) False negative (0) | True negative (10) False positive (0) |
| ERS31del assay | True positive (32) False negative (0) | True positive (32) False negative (0) | True negative (22) False positive (0) | True negative (10) False positive (0) |
| Ins214EPE assay | True negative (32) False positive (0) | True positive (32) False negative (0) | True negative (22) False positive (0) | True negative (10) False positive (0) |
| 2-SARS-CoV-2 Assay | ERS31del Assay | Ins214EPE Assay | RP Assay | |
|---|---|---|---|---|
| FAM | HEX | ROX | Cy5 | |
| + | − | − | + | SARS-CoV-2, non-Omicron |
| + | + | − | + | SARS-CoV-2 Omicron lineage, non-BA.1 |
| + | + | + | + | SARS-CoV-2 Omicron lineage, BA.1 |
| − | − | − | + | No SARS-CoV-2 RNA in the sample |
| − | − | − | − | No RNA in the reaction/ RNA extraction failed |
| Target | Name | Sequence |
|---|---|---|
| ORF1 | 2-SARS-CoV-2-ORF-1-F | AGAGCTATGAATTGCAGAC |
| ORF1 | 2-SARS-CoV-2-ORF-1-R | GGGAAATACAAAATTTGGACA |
| ORF1 | 2-SARS-CoV-2-ORF-1-P | FAM-AATTGGCAAAGAAATTTGACACCTTCA-BHQ1 |
| ERS31del | N31-33del F | GTTTGGTGGACCCTCAGATT |
| ERS31del | N31-33del R | CAAGACGCAGTATTATTGGGTAAAC |
| ERS31del | N31-33del P | HEX-AGTAACCAGAATGGTGGGGCGCG-BHQ1 |
| Ins214EPE | Ins214EPE F | ATATTCTAAGCACACGCCTATT |
| Ins214EPE | Ins214EPE R | GGCAAATCTACCAATGGTTCTA |
| Ins214EPE | Ins214EPE P | ROX-TGCGTGAGCCAGAAGATCTCCCT-BHQ2 |
| Human RP | RP-CB-F | AGATTTGGACCTGCGAGCG |
| Human RP | RP-CB-R | GAGCGGCTGTCTCCACAAGT |
| Human RP | RP-CB-P | Cy5-TTCTGACCTGAAGGCTCTGCGCG-BHQ2 |
| Reagents | μL | Concentration | Quantity |
|---|---|---|---|
| 2-SARS-CoV-2-ORF-1-F | 7.5 | 100 pmol/μL | 300 nM |
| 2-SARS-CoV-2-ORF-1-R | 7.5 | 100 pmol/μL | 300 nM |
| 2-SARS-CoV-2-ORF-1-P | 5 | 100 pmol/μL | 200 nM |
| N31-33del F | 2.5 | 100 pmol/μL | 100 nM |
| N31-33del R | 2.5 | 100 pmol/μL | 100 nM |
| N31-33del P | 1.25 | 100 pmol/μL | 50 nM |
| Ins214EPE F | 10 | 100 pmol/μL | 400 nM |
| Ins214EPE R | 10 | 100 pmol/μL | 400 nM |
| Ins214EPE P | 5 | 100 pmol/μL | 200 nM |
| RP F | 5 | 100 pmol/μL | 200 nM |
| RP R | 5 | 100 pmol/μL | 200 nM |
| RP P | 5 | 100 pmol/μL | 200 nM |
| water | 53.75 | ||
| Total volume | 120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yolshin, N.D.; Komissarov, A.B.; Varchenko, K.V.; Musaeva, T.D.; Fadeev, A.V.; Lioznov, D.A. Detection of the Omicron SARS-CoV-2 Lineage and Its BA.1 Variant with Multiplex RT-qPCR. Int. J. Mol. Sci. 2022, 23, 16153. https://doi.org/10.3390/ijms232416153
Yolshin ND, Komissarov AB, Varchenko KV, Musaeva TD, Fadeev AV, Lioznov DA. Detection of the Omicron SARS-CoV-2 Lineage and Its BA.1 Variant with Multiplex RT-qPCR. International Journal of Molecular Sciences. 2022; 23(24):16153. https://doi.org/10.3390/ijms232416153
Chicago/Turabian StyleYolshin, Nikita D., Andrey B. Komissarov, Kirill V. Varchenko, Tamila D. Musaeva, Artem V. Fadeev, and Dmitry A. Lioznov. 2022. "Detection of the Omicron SARS-CoV-2 Lineage and Its BA.1 Variant with Multiplex RT-qPCR" International Journal of Molecular Sciences 23, no. 24: 16153. https://doi.org/10.3390/ijms232416153