
May EPH/Ephrin Targeting Revolutionize Lung Cancer Treatment?

 , , , ,
, , , ,
Abstract
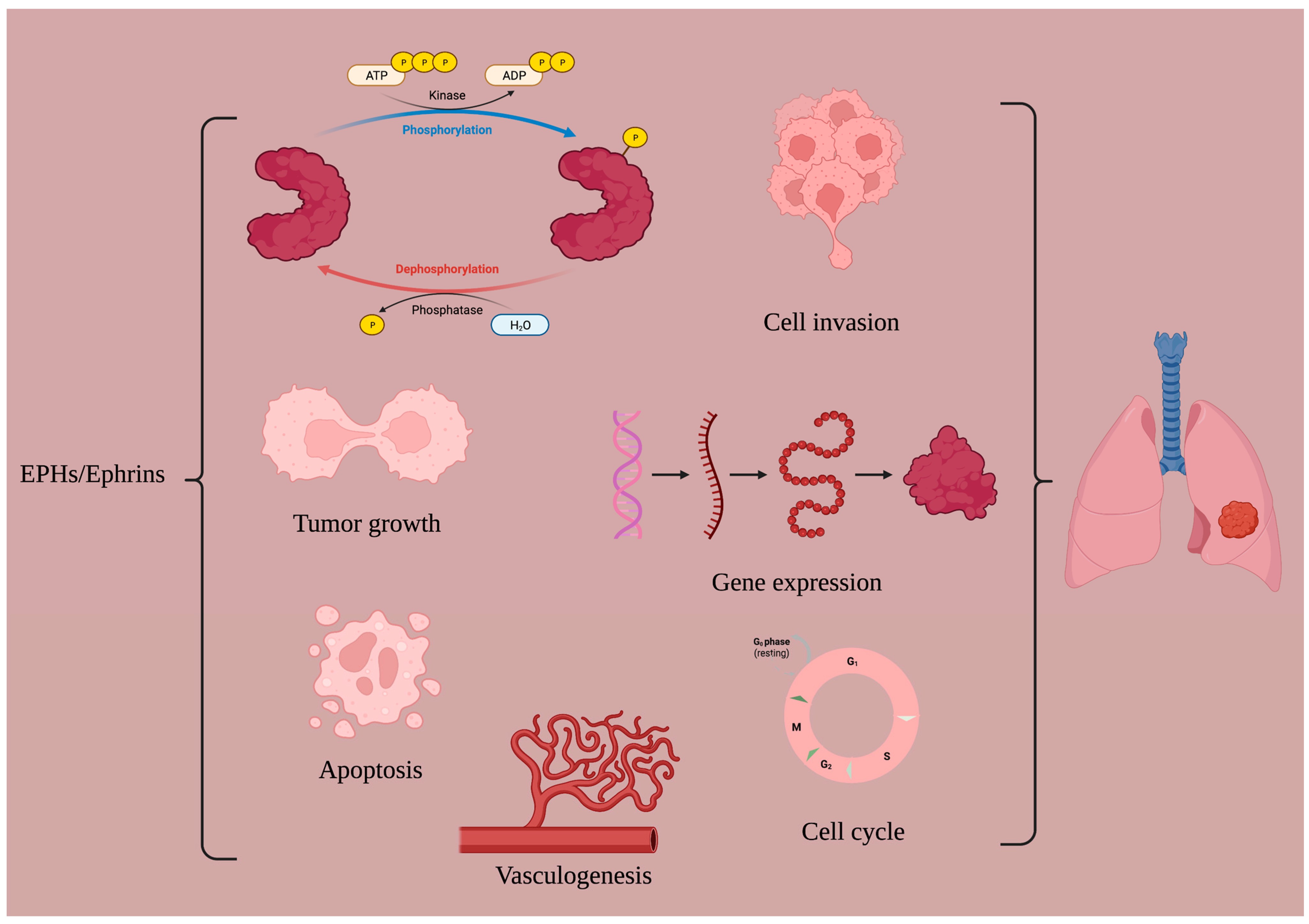
:1. Introduction
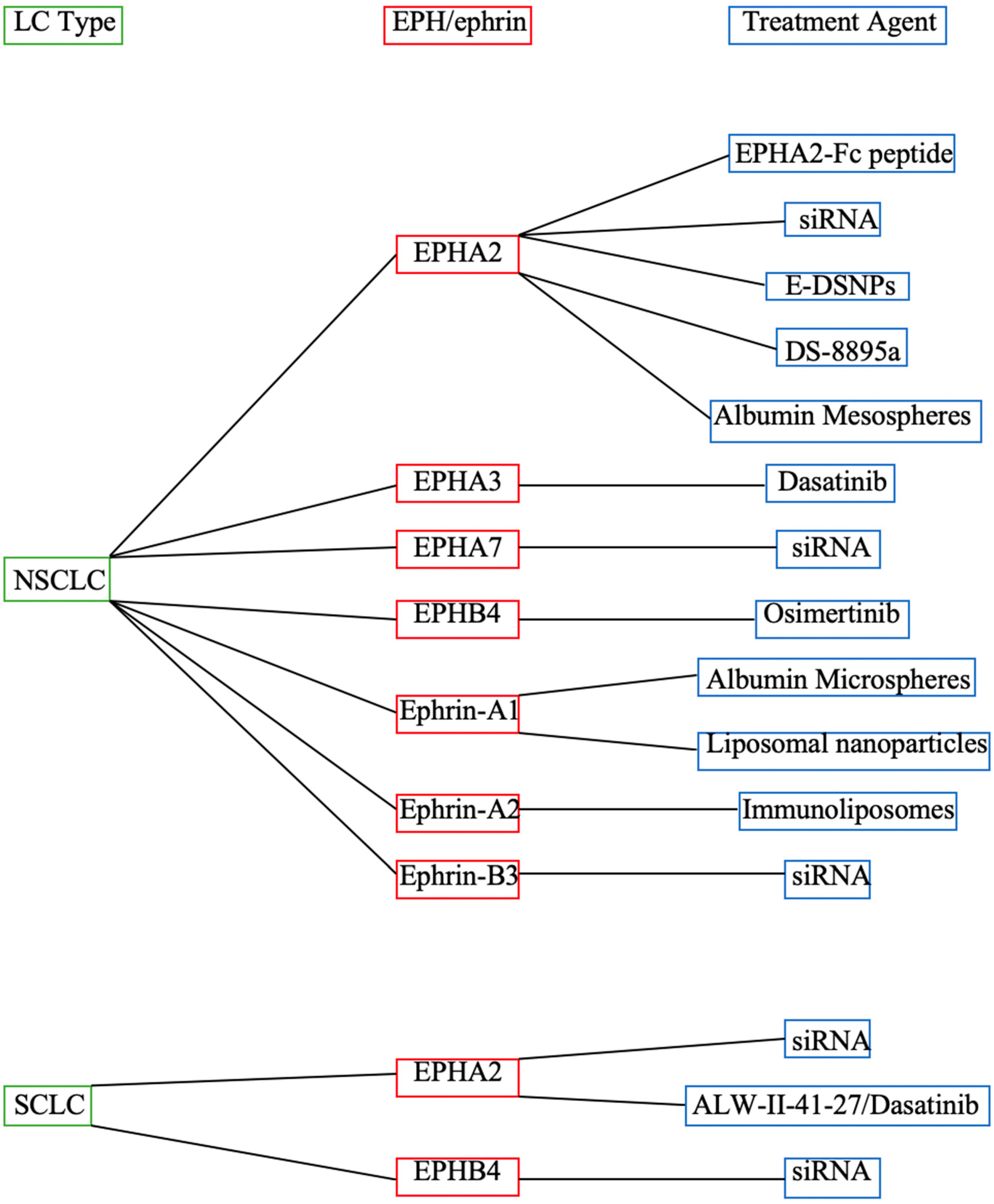
2. Available EPH/Ephrin-Targeting Agents in LC
3. EPHA-Targeting Therapeutic Agents
4. EPHB-Targeting Therapeutic Agents
5. Ephrin-Targeting Therapeutic Agents
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Key Statistics for Lung Cancer; American Cancer Society: Atlanta, GA, USA, 2022.
- What Is Lung Cancer? American Cancer Society: Atlanta, GA, USA, 2019.
- Signs and Symptoms of Lung Cancer; American Cancer Society: Atlanta, GA, USA, 2019.
- Tests for Lung Cancer; American Cancer Society: Atlanta, GA, USA, 2022.
- Treatment Choices for Non-Small Cell Lung Cancer, by Stage; American Cancer Society: Atlanta, GA, USA, 2022.
- Treatment Choices for Small Cell Lung Cancer, by Stage; American Cancer Society: Atlanta, GA, USA, 2021.
- Eph Nomenclature Committee. Unified nomenclature for Eph family receptors and their ligands, the ephrins. Cell 1997, 90, 403–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Liu, J.; Wang, N.; Zhang, X.; Jin, J.; Chin-Sang, I.; Zheng, J.; Jia, Z. Structures of an Eph receptor tyrosine kinase and its potential activation mechanism. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 3135–3143. [Google Scholar] [CrossRef] [PubMed]
- Kania, A.; Klein, R. Mechanisms of ephrin-Eph signalling in development, physiology and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Yuasa, M.; Masaoka, T.; Taniyama, T.; Maehara, H.; Torigoe, I.; Yoshii, T.; Shinomiya, K.; Okawa, A.; Sotome, S. After repeated division, bone marrow stromal cells express inhibitory factors with osteogenic capabilities, and EphA5 is a primary candidate. Bone 2013, 57, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiuan, E.; Chen, J. Eph Receptor Tyrosine Kinases in Tumor Immunity. Cancer Res. 2016, 76, 6452–6457. [Google Scholar] [CrossRef] [Green Version]
- Psilopatis, I.; Pergaris, A.; Vrettou, K.; Tsourouflis, G.; Theocharis, S. The EPH/Ephrin System in Gynecological Cancers: Focusing on the Roots of Carcinogenesis for Better Patient Management. Int. J. Mol. Sci. 2022, 23, 3249. [Google Scholar] [CrossRef]
- Coulthard, M.G.; Duffy, S.; Down, M.; Evans, B.; Power, M.; Smith, F.; Stylianou, C.; Kleikamp, S.; Oates, A.; Lackmann, M.; et al. The role of the Eph-ephrin signalling system in the regulation of developmental patterning. Int. J. Dev. Biol. 2002, 46, 375–384. [Google Scholar]
- Rudno-Rudzinska, J.; Kielan, W.; Frejlich, E.; Kotulski, K.; Hap, W.; Kurnol, K.; Dzierżek, P.; Zawadzki, M.; Hałoń, A. A review on Eph/ephrin, angiogenesis and lymphangiogenesis in gastric, colorectal and pancreatic cancers. Chin. J. Cancer Res. 2017, 29, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Pergaris, A.; Danas, E.; Goutas, D.; Sykaras, A.G.; Soranidis, A.; Theocharis, S. The Clinical Impact of the EPH/Ephrin System in Cancer: Unwinding the Thread. Int. J. Mol. Sci. 2021, 22, 8412. [Google Scholar] [CrossRef]
- Papadakos, S.P.; Petrogiannopoulos, L.; Pergaris, A.; Theocharis, S. The EPH/Ephrin System in Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 2761. [Google Scholar] [CrossRef]
- Masaoutis, C.; Georgantzoglou, N.; Sarantis, P.; Theochari, I.; Tsoukalas, N.; Bobos, M.; Alexandrou, P.; Pergaris, A.; Rontogianni, D.; Theocharis, S. Ephrin Receptors (Ephs) Expression in Thymic Epithelial Tumors: Prognostic Implications and Future Therapeutic Approaches. Diagnostics 2021, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Hadjimichael, A.C.; Pergaris, A.; Kaspiris, A.; Foukas, A.F.; Kokkali, S.; Tsourouflis, G.; Theocharis, S. The EPH/Ephrin System in Bone and Soft Tissue Sarcomas’ Pathogenesis and Therapy: New Advancements and a Literature Review. Int. J. Mol. Sci. 2022, 23, 5171. [Google Scholar] [CrossRef] [PubMed]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Rev. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goutas, D.; Pergaris, A.; Goutas, N.; Theocharis, S. Utilizing Exosomal-EPHs/Ephrins as Biomarkers and as a Potential Platform for Targeted Delivery of Therapeutic Exosomes. Int. J. Mol. Sci. 2022, 23, 3551. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Li, X.; Xu, S. EphA10 drives tumor progression and immune evasion by regulating the MAPK/ERK cascade in lung adenocarcinoma. Int. Immunopharmacol. 2022, 110, 109031. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, Y.; Sun, X.; He, Z.; Wu, T.; Wu, C.; Chen, J.; Wang, J.; Diao, K.; Liu, X. Oncogenic EFNA4 Amplification Promotes Lung Adenocarcinoma Lymph Node Metastasis. Cancers 2022, 14, 4226. [Google Scholar] [CrossRef]
- Yiminniyaze, R.; Zhang, X.; Zhu, N.; Wang, J.; Li, C.; Wumaier, G.; Zhou, D.; Li, J.; Xia, J.; Zhang, Y.; et al. EphrinA3 is a key regulator of malignant behaviors and a potential prognostic factor in lung adenocarcinoma. Cancer Med. 2022. [Google Scholar] [CrossRef]
- Deng, M.; Tong, R.; Zhang, Z.; Wang, T.; Liang, C.; Zhou, X.; Hou, G. EFNA3 as a predictor of clinical prognosis and immune checkpoint therapy efficacy in patients with lung adenocarcinoma. Cancer Cell Int. 2021, 21, 535. [Google Scholar] [CrossRef]
- Wang, L.; Peng, Q.; Sai, B.; Zheng, L.; Xu, J.; Yin, N.; Feng, X.; Xiang, J. Ligand-independent EphB1 signaling mediates TGF-beta-activated CDH2 and promotes lung cancer cell invasion and migration. J. Cancer 2020, 11, 4123–4131. [Google Scholar] [CrossRef] [Green Version]
- Volz, C.; Breid, S.; Selenz, C.; Zaplatina, A.; Golfmann, K.; Meder, L.; Dietlein, F.; Borchmann, S.; Chatterjee, S.; Siobal, M.; et al. Inhibition of Tumor VEGFR2 Induces Serine 897 EphA2-Dependent Tumor Cell Invasion and Metastasis in NSCLC. Cell Rep. 2020, 31, 107568. [Google Scholar] [CrossRef]
- Li, J.; Sun, Y.; Wang, X.; Wang, J.; Zhu, Y. The expressions of EphB4 and ephrinB2 in lung adenocarcinomas: A high level of the EphB4 protein is associated with lymph node metastasis. Int. J. Clin. Exp. Pathol. 2019, 12, 3447–3452. [Google Scholar] [PubMed]
- Zhao, C.; Wang, A.; Lu, F.; Chen, H.; Fu, P.; Zhao, X.; Chen, H. Overexpression of junctional adhesion molecule-A and EphB2 predicts poor survival in lung adenocarcinoma patients. Tumour Biol. 2017, 39, 1010428317691000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efazat, G.; Novak, M.; Kaminskyy, V.O.; De Petris, L.; Kanter, L.; Juntti, T.; Bergman, P.; Zhivotovsky, B.; Lewensohn, R.; Hååg, P.; et al. Ephrin B3 interacts with multiple EphA receptors and drives migration and invasion in non-small cell lung cancer. Oncotarget 2016, 7, 60332–60347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaginis, C.; Tsoukalas, N.; Bournakis, E.; Alexandrou, P.; Kavantzas, N.; Patsouris, E.; Theocharis, S. Ephrin (Eph) receptor A1, A4, A5 and A7 expression in human non-small cell lung carcinoma: Associations with clinicopathological parameters, tumor proliferative capacity and patients’ survival. BMC Clin. Pathol. 2014, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Saintigny, P.; Peng, S.; Zhang, L.; Sen, B.; Wistuba, I.I.; Lippman, S.M.; Girard, L.; Minna, J.D.; Heymach, J.V.; Johnson, F.M. Global evaluation of Eph receptors and ephrins in lung adenocarcinomas identifies EphA4 as an inhibitor of cell migration and invasion. Mol. Cancer Ther. 2012, 11, 2021–2032. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Ji, X.D.; Gao, H.; Zhao, J.S.; Xu, J.F.; Sun, Z.J.; Deng, Y.Z.; Shi, S.; Feng, Y.X.; Zhu, Y.Q.; et al. EphB3 suppresses non-small-cell lung cancer metastasis via a PP2A/RACK1/Akt signalling complex. Nat. Commun. 2012, 3, 667. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Miyahara, R.; Sonobe, M.; Horiuchi, M.; Mennju, T.; Nakayama, E.; Kobayashi, M.; Kikuchi, R.; Kitamura, J.; Imamura, N.; et al. Higher expression of EphA2 and ephrin-A1 is related to favorable clinicopathological features in pathological stage I non-small cell lung carcinoma. Lung Cancer 2012, 76, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Bulk, E.; Yu, J.; Hascher, A.; Koschmieder, S.; Wiewrodt, R.; Krug, U.; Timmermann, B.; Marra, A.; Hillejan, L.; Wiebe, K.; et al. Mutations of the EPHB6 receptor tyrosine kinase induce a pro-metastatic phenotype in non-small cell lung cancer. PLoS ONE 2012, 7, e44591. [Google Scholar] [CrossRef] [Green Version]
- Anderton, M.; van der Meulen, E.; Blumenthal, M.J.; Schafer, G. The Role of the Eph Receptor Family in Tumorigenesis. Cancers 2021, 13, 206. [Google Scholar] [CrossRef]
- Kou, C.J.; Kandpal, R.P. Differential Expression Patterns of Eph Receptors and Ephrin Ligands in Human Cancers. Biomed Res. Int. 2018, 2018, 7390104. [Google Scholar] [CrossRef] [Green Version]
- Pottier, C.; Fresnais, M.; Gilon, M.; Jerusalem, G.; Longuespee, R.; Sounni, N.E. Tyrosine Kinase Inhibitors in Cancer: Breakthrough and Challenges of Targeted Therapy. Cancers 2020, 12, 731. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.Y.; Sparreboom, A.; Baker, S.D. Kinase Inhibitors: The Reality Behind the Success. Clin. Pharmacol. Ther. 2017, 102, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Noor, Z.S.; Cummings, A.L.; Johnson, M.M.; Spiegel, M.L.; Goldman, J.W. Targeted Therapy for Non-Small Cell Lung Cancer. Semin. Respir. Crit. Care Med. 2020, 41, 409–434. [Google Scholar] [CrossRef]
- Sun, B.K.; Tsao, H. Small RNAs in development and disease. J. Am. Acad. Dermatol. 2008, 59, 725–737, quiz 38–40. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Liang, G.; Cui, K.; Liang, Y.; Wang, Q.; Lv, S.; Cheng, X.; Zhang, L. Insight Into the Prospects for RNAi Therapy of Cancer. Front. Pharmacol. 2021, 12, 644718. [Google Scholar] [CrossRef] [PubMed]
- Khan, P.; Siddiqui, J.A.; Lakshmanan, I.; Ganti, A.K.; Salgia, R.; Jain, M.; Batra, S.K.; Nasser, M.W. RNA-based therapies: A cog in the wheel of lung cancer defense. Mol. Cancer 2021, 20, 54. [Google Scholar] [CrossRef]
- Zahavi, D.; Weiner, L. Monoclonal Antibodies in Cancer Therapy. Antibodies 2020, 9, 34. [Google Scholar] [CrossRef]
- Paszko, E.; Senge, M.O. Immunoliposomes. Curr. Med. Chem. 2012, 19, 5239–5277. [Google Scholar] [CrossRef]
- Drugs Approved for Lung Cancer; National Cancer Institute: Bethesda, MD, USA, 2022.
- Cho, H.; Jeon, S.I.; Ahn, C.H.; Shim, M.K.; Kim, K. Emerging Albumin-Binding Anticancer Drugs for Tumor-Targeted Drug Delivery: Current Understandings and Clinical Translation. Pharmaceutics 2022, 14, 728. [Google Scholar] [CrossRef]
- Kundranda, M.N.; Niu, J. Albumin-bound paclitaxel in solid tumors: Clinical development and future directions. Drug Des. Dev. Ther. 2015, 9, 3767–3777. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, H.; Chen, G. Core-shell nanoparticles for cancer imaging and therapy. In Core-Shell Nanostructures for Drug Delivery and Theranostics; Focarete, M.L., Tampieri, A., Eds.; Woodhead Publishing: Cambridge, UK, 2018; pp. 143–175. [Google Scholar]
- Yu, H.; Wang, Y.; Wang, S.; Li, X.; Li, W.; Ding, D.; Gong, X.; Keidar, M.; Zhang, W. Paclitaxel-Loaded Core-Shell Magnetic Nanoparticles and Cold Atmospheric Plasma Inhibit Non-Small Cell Lung Cancer Growth. ACS Appl. Mater. Interfaces 2018, 10, 43462–43471. [Google Scholar] [CrossRef] [PubMed]
- Menon, J.U.; Kuriakose, A.; Iyer, R.; Hernandez, E.; Gandee, L.; Zhang, S.; Takahashi, M.; Zhang, Z.; Saha, D.; Nguyen, K.T. Dual-Drug Containing Core-Shell Nanoparticles for Lung Cancer Therapy. Sci. Rep. 2017, 7, 13249. [Google Scholar] [CrossRef] [Green Version]
- Vaisberg, A.J.; Milla, M.; Planas, M.C.; Cordova, J.L.; De Agusti, E.R.; Ferreyra, R.; Mustiga, M.C.; Carlin, L.; Hammond, G.B. Taspine is the cicatrizant principle in Sangre de Grado extracted from Croton lechleri. Planta Med. 1989, 55, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, H.; Qi, G.; Gu, X.; Zhao, Y.; Zhang, J. TPD7 inhibits the non-small cell lung cancer HCC827 cell growth by regulating EGFR signalling pathway. J. Chemother. 2022, 34, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Dai, B.; Ma, W.; Zhang, Y. A novel taspine analog, HMQ1611, inhibits growth of non-small cell lung cancer by inhibiting angiogenesis. Oncol. Lett. 2012, 4, 1109–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montopoli, M.; Bertin, R.; Chen, Z.; Bolcato, J.; Caparrotta, L.; Froldi, G. Croton lechleri sap and isolated alkaloid taspine exhibit inhibition against human melanoma SK23 and colon cancer HT29 cell lines. J. Ethnopharmacol. 2012, 144, 747–753. [Google Scholar] [CrossRef]
- Zhan, Y.; Zhang, Y.; Liu, C.; Zhang, J.; Smith, W.W.; Wang, N.; Chen, Y.; Zheng, L.; He, L. A novel taspine derivative, HMQ1611, inhibits breast cancer cell growth via estrogen receptor alpha and EGF receptor signaling pathways. Cancer Prev. Res. 2012, 5, 864–873. [Google Scholar] [CrossRef] [Green Version]
- Dai, B.; Wang, W.; Ma, Y.; Liu, R.; Zhang, Y. A taspine derivative supresses Caco-2 cell growth by competitively targeting EphrinB2 and regulating its pathway. Oncol. Rep. 2016, 36, 1526–1534. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Duan, J.; Li, C.; Xie, W.; Fang, W.; Xu, Y.; Wang, G.; Wan, R.; Sun, J.; Xu, J.; et al. EPHA mutation as a predictor of immunotherapeutic efficacy in lung adenocarcinoma. J. Immunother. Cancer 2020, 8, e001315. [Google Scholar] [CrossRef]
- Li, J.; Rix, U.; Fang, B.; Bai, Y.; Edwards, A.; Colinge, J.; Bennett, K.L.; Gao, J.; Song, L.; Eschrich, S.; et al. A chemical and phosphoproteomic characterization of dasatinib action in lung cancer. Nat. Chem. Biol. 2010, 6, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Creighton, C.J.; Stivers, D.; Fujimoto, N.; Kurie, J.M. Transcriptional profiling of non-small cell lung cancer cells with activating EGFR somatic mutations. PLoS ONE 2007, 2, e1226. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.; Busto, M.E.; Kramer, K.; Medard, G.; Kuster, B. Chemical Proteomics Uncovers EPHA2 as a Mechanism of Acquired Resistance to Small Molecule EGFR Kinase Inhibition. J. Proteome Res. 2015, 14, 2617–2625. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Pang, K.M.; Iida, M.; Nelson, M.S.; Liu, J.; Nam, A.; Wang, J.; Mambetsariev, I.; Pillai, R.; Mohanty, A.; et al. Activation of EPHA2-ROBO1 Heterodimer by SLIT2 Attenuates Non-canonical Signaling and Proliferation in Squamous Cell Carcinomas. iScience 2020, 23, 101692. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R.; Wang, S.; Tan, L.; Hastings, A.K.; Song, W.; Lovly, C.M.; Meador, C.B.; Ye, F.; Lu, P.; Balko, J.M.; et al. EPHA2 Blockade Overcomes Acquired Resistance to EGFR Kinase Inhibitors in Lung Cancer. Cancer Res. 2016, 76, 305–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishigaki, H.; Minami, T.; Morimura, O.; Kitai, H.; Horio, D.; Koda, Y.; Fujimoto, E.; Negi, Y.; Nakajima, Y.; Niki, M.; et al. EphA2 inhibition suppresses proliferation of small-cell lung cancer cells through inducing cell cycle arrest. Biochem. Biophys. Res. Commun. 2019, 519, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Kaminskyy, V.O.; Haag, P.; Novak, M.; Végvári, Á.; Arapi, V.; Lewensohn, R.; Viktorsson, K. EPHA2 Interacts with DNA-PKcs in Cell Nucleus and Controls Ionizing Radiation Responses in Non-Small Cell Lung Cancer Cells. Cancers 2021, 13, 1010. [Google Scholar] [CrossRef]
- Iyer, R.; Ramachandramoorthy, H.; Nguyen, T.; Xu, C.; Fu, H.; Kotadia, T.; Chen, B.; Hong, Y.; Saha, D.; Nguyen, K.T. Lung Cancer Targeted Chemoradiotherapy via Dual-Stimuli Responsive Biodegradable Core-Shell Nanoparticles. Pharmaceutics 2022, 14, 1525. [Google Scholar] [CrossRef]
- Lee, H.Y.; Mohammed, K.A.; Kaye, F.; Moudgil, B.M.; Nasreen, N. EphA2 targeted intratumoral therapy for non-small cell lung cancer using albumin mesospheres. Am. J. Transl. Res. 2017, 9, 3293–3303. [Google Scholar]
- Gan, H.K.; Parakh, S.; Lee, F.T.; Tebbutt, N.C.; Ameratunga, M.; Lee, S.T.; O’Keefe, G.J.; Gong, S.J.; Vanrenen, C.; Caine, J.; et al. A phase 1 safety and bioimaging trial of antibody DS-8895a against EphA2 in patients with advanced or metastatic EphA2 positive cancers. Investig. New Drugs 2022, 40, 747–755. [Google Scholar] [CrossRef]
- Peng, J.; Wang, Q.; Liu, H.; Ye, M.; Wu, X.; Guo, L. EPHA3 regulates the multidrug resistance of small cell lung cancer via the PI3K/BMX/STAT3 signaling pathway. Tumour Biol. 2016, 37, 11959–11971. [Google Scholar] [CrossRef] [Green Version]
- Sos, M.L.; Michel, K.; Zander, T.; Weiss, J.; Frommolt, P.; Peifer, M.; Li, D.; Ullrich, R.; Koker, M.; Fischer, F.; et al. Predicting drug susceptibility of non-small cell lung cancers based on genetic lesions. J. Clin. Investig. 2009, 119, 1727–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staquicini, F.I.; Qian, M.D.; Salameh, A.; Dobroff, A.S.; Edwards, J.K.; Cimino, D.F.; Moeller, B.J.; Kelly, P.; Nunez, M.I.; Tang, X.; et al. Receptor tyrosine kinase EphA5 is a functional molecular target in human lung cancer. J. Biol. Chem. 2015, 290, 7345–7359. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Sun, Y.; Jiang, A.; Wu, Y.; Li, C.; Jin, M.; Yan, H.; Jin, H. Knockdown of ephrin receptor A7 suppresses the proliferation and metastasis of A549 human lung cancer cells. Mol. Med. Rep. 2016, 13, 3190–3196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.D.; Liu, R.; Rolle, C.E.; Tan, Y.H.C.; Krasnoperov, V.; Kanteti, R.; Tretiakova, M.S.; Cervantes, G.M.; Hasina, R.; Hseu, R.D.; et al. The EphB4 receptor tyrosine kinase promotes lung cancer growth: A potential novel therapeutic target. PLoS ONE 2013, 8, e67668. [Google Scholar] [CrossRef] [Green Version]
- Nanamiya, R.; Saito-Koyama, R.; Miki, Y.; Inoue, C.; Asavasupreechar, T.; Abe, J.; Sato, I.; Sasano, H. EphB4 as a Novel Target for the EGFR-Independent Suppressive Effects of Osimertinib on Cell Cycle Progression in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 8522. [Google Scholar] [CrossRef]
- Yoon, S.; Choi, J.H.; Kim, S.J.; Lee, E.J.; Shah, M.; Choi, S.; Woo, H.G. EPHB6 mutation induces cell adhesion-mediated paclitaxel resistance via EPHA2 and CDH11 expression. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Mohammed, K.A.; Peruvemba, S.; Goldberg, E.P.; Nasreen, N. Targeted lung cancer therapy using ephrinA1-loaded albumin microspheres. J. Pharm. Pharmacol. 2011, 63, 1401–1410. [Google Scholar] [CrossRef]
- Lee, H.Y.; Mohammed, K.A.; Kaye, F.; Sharma, P.; Moudgil, B.M.; Clapp, W.L.; Nasreen, N. Targeted delivery of let-7a microRNA encapsulated ephrin-A1 conjugated liposomal nanoparticles inhibit tumor growth in lung cancer. Int. J. Nanomed. 2013, 8, 4481–4494. [Google Scholar]
- Murugan, D.; Rangasamy, L. A perspective to weaponize microRNAs against lung cancer. Noncoding RNA Res. 2022, 8, 18–32. [Google Scholar] [CrossRef]
- Huang, Z.R.; Tipparaju, S.K.; Kirpotin, D.B.; Pien, C.; Kornaga, T.; O Noble, C.; Koshkaryev, A.; Tran, J.; Kamoun, W.S.; Drummond, D.C. Formulation optimization of an ephrin A2 targeted immunoliposome encapsulating reversibly modified taxane prodrugs. J. Control. Release 2019, 310, 47–57. [Google Scholar] [CrossRef]
- Dai, B.; Ma, Y.; Yang, T.; Wang, W.; Zhang, Y. Taspine derivative 12k suppressed A549 cell migration through the Wnt/beta-catenin and EphrinB2 signaling pathway. Biomed. Pharmacother. 2017, 87, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, S.; Branca, R.M.; Efazat, G.; Ruzzene, M.; Zhivotovsky, B.; Lewensohn, R.; Viktorsson, K.; Lehtiö, J. Phosphoproteomic profiling of NSCLC cells reveals that ephrin B3 regulates pro-survival signaling through Akt1-mediated phosphorylation of the EphA2 receptor. J. Proteome Res. 2011, 10, 2566–2578. [Google Scholar] [CrossRef] [PubMed]
- Lung Cancer Survival Rates; American Cancer Society: Atlanta, GA, USA, 2022.



{kind=link}
{kind=link}
| Targeting Strategy | Mechanism of Action |
|---|---|
| Small-molecule drugs | Tyrosine kinase inhibitors |
| Regulatory small RNA molecules | Short interfering RNAs |
| Monoclonal antibodies | Anti-EPH/ephrin monoclonal antibodies, antibody–drug conjugations |
| Immunoliposomes | Antibodies coupled to the liposomal surface |
| Nanoparticles | Albumin microspheres, core–shell nanoparticles |
| Natural compounds | Taspine derivatives |
| Targeted EPH/Ephrin | Therapeutic Agent | Mechanism of Action | References |
|---|---|---|---|
| EPHA2 | TKIs |
regulation;
| [58,60,61,62] |
| EPHA2-Fc peptide |
inhibition. | [59] | |
| siRNA |
amplification. | [63,64] | |
| E-DSNPs |
concurrent radiation therapy;
| [65] | |
| Albumin mesosphere-conjugated ephrin-A1 |
| [66] | |
| DS-8895a |
| [67] | |
| EPHA3 | Dasatinib |
| [69] |
| EPHA5 | 11C12 |
effect;
| [70] |
| EPHA7 | siRNA |
expression levels;
| [71] |
| EPHB4 | siRNA |
| [72] |
| Osimertinib |
| [73] | |
| EPHB6 | ALW-II-41-27 |
expression downregulation;
| [74] |
| Ephrin-A1 | Ephrin-A1-loaded albumin microspheres |
| [75] |
| let-7a microRNA-encapsulated ephrin-A1-conjugated liposomal nanoparticles |
| [76] | |
| Ephrin-A2 | Ephrin-A2-targeted taxane liposomal prodrug 2 |
| [78] |
| Ephrin-B2 | 12k |
| [79] |
| Ephrin-B3 | siRNA |
| [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Psilopatis, I.; Karniadakis, I.; Danos, K.S.; Vrettou, K.; Michaelidou, K.; Mavridis, K.; Agelaki, S.; Theocharis, S. May EPH/Ephrin Targeting Revolutionize Lung Cancer Treatment? Int. J. Mol. Sci. 2023, 24, 93. https://doi.org/10.3390/ijms24010093
Psilopatis I, Karniadakis I, Danos KS, Vrettou K, Michaelidou K, Mavridis K, Agelaki S, Theocharis S. May EPH/Ephrin Targeting Revolutionize Lung Cancer Treatment? International Journal of Molecular Sciences. 2023; 24(1):93. https://doi.org/10.3390/ijms24010093
Chicago/Turabian StylePsilopatis, Iason, Ioannis Karniadakis, Konstantinos Stylianos Danos, Kleio Vrettou, Kleita Michaelidou, Konstantinos Mavridis, Sofia Agelaki, and Stamatios Theocharis. 2023. "May EPH/Ephrin Targeting Revolutionize Lung Cancer Treatment?" International Journal of Molecular Sciences 24, no. 1: 93. https://doi.org/10.3390/ijms24010093