
Mice Lacking Gpr179 with Complete Congenital Stationary Night Blindness Are a Good Model for Myopia
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
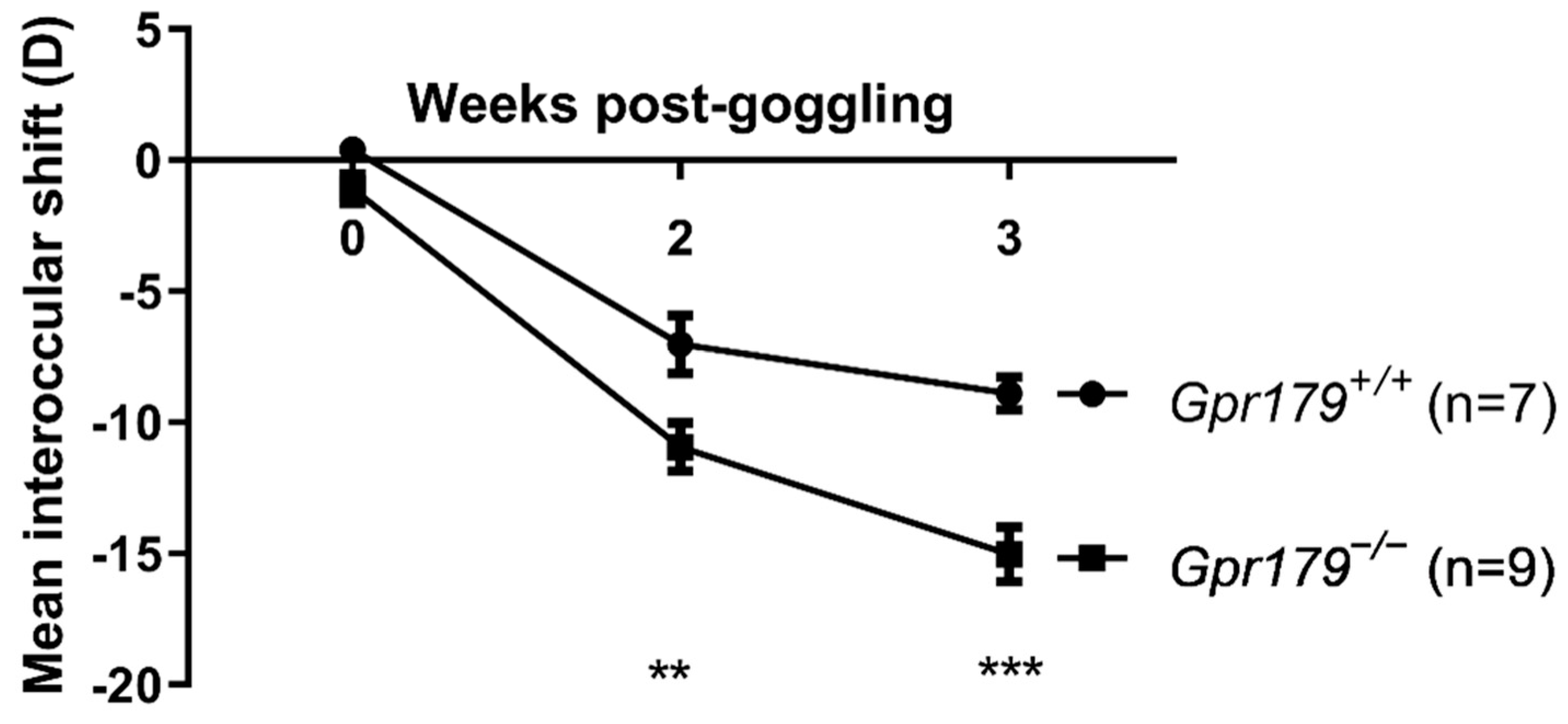
2. Results
2.1. Measurement of Retinal Levels of DA and DOPAC
2.2. Kinetics of Refractive Development
2.3. Assessment of Sensitivity to Myopia Induction
3. Discussion
4. Material and Methods
4.1. Animal Care and Ethical Statement
4.2. Polymerase Chain Reaction (PCR) Genotyping
4.3. DA and DOPAC Measurements
4.4. Lens-Induced Myopia
4.5. Refractometry
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, P.C.; Huang, H.M.; Yu, H.J.; Fang, P.C.; Chen, C.T. Epidemiology of Myopia. Asia Pac. J. Ophthalmol. 2016, 5, 386–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, J.; Tkatchenko, A.V. A Review of Current Concepts of the Etiology and Treatment of Myopia. Eye Contact Lens 2018, 44, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Dolgin, E. The myopia boom. Nat. News 2015, 519, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flitcroft, D.I.; He, M.; Jonas, J.B.; Jong, M.; Naidoo, K.; Ohno-Matsui, K.; Rahi, J.; Resnikoff, S.; Vitale, S.; Yannuzzi, L. IMI–Defining and Classifying Myopia: A Proposed Set of Standards for Clinical and Epidemiologic Studies. Investig. Ophthalmol. Vis. Sci. 2019, 60, M20–M30. [Google Scholar] [CrossRef] [Green Version]
- Ikuno, Y. Overview of the Complications of High Myopia. Retina 2017, 37, 2347–2351. [Google Scholar] [CrossRef]
- Bremond-Gignac, D. Myopia in children. Med. Sci. 2020, 36, 763–768. [Google Scholar]
- Wu, P.C.; Chen, C.T.; Lin, K.K.; Sun, C.C.; Kuo, C.N.; Huang, H.M.; Poon, Y.C.; Yang, M.L.; Chen, C.Y.; Huang, J.C.; et al. Myopia Prevention and Outdoor Light Intensity in a School-Based Cluster Randomized Trial. Ophthalmology 2018, 125, 1239–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.C.; Tsai, C.L.; Wu, H.L.; Yang, Y.H.; Kuo, H.K. Outdoor activity during class recess reduces myopia onset and progression in school children. Ophthalmology 2013, 120, 1080–1085. [Google Scholar] [CrossRef]
- Jones, L.A.; Sinnott, L.T.; Mutti, D.O.; Mitchell, G.L.; Moeschberger, M.L.; Zadnik, K. Parental history of myopia, sports and outdoor activities, and future myopia. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3524–3532. [Google Scholar] [CrossRef] [Green Version]
- Jones-Jordan, L.A.; Sinnott, L.T.; Chu, R.H.; Cotter, S.A.; Kleinstein, R.N.; Manny, R.E.; Mutti, D.O.; Twelker, J.D.; Zadnik, K. Myopia Progression as a Function of Sex, Age, and Ethnicity. Investig. Ophthalmol. Vis. Sci. 2021, 62, 36. [Google Scholar] [CrossRef]
- Mutti, D.O.; Mitchell, G.L.; Moeschberger, M.L.; Jones, L.A.; Zadnik, K. Parental myopia, near work, school achievement, and children’s refractive error. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3633–3640. [Google Scholar]
- French, A.N.; Ashby, R.S.; Morgan, I.G.; Rose, K.A. Time outdoors and the prevention of myopia. Exp. Eye Res. 2013, 114, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Rose, K.A.; Morgan, I.G.; Ip, J.; Kifley, A.; Huynh, S.; Smith, W.; Mitchell, P. Outdoor activity reduces the prevalence of myopia in children. Ophthalmology 2008, 115, 1279–1285. [Google Scholar] [CrossRef]
- Ashby, R.; Ohlendorf, A.; Schaeffel, F. The effect of ambient illuminance on the development of deprivation myopia in chicks. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5348–5354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.L., 3rd; Hung, L.F.; Huang, J. Protective effects of high ambient lighting on the development of form-deprivation myopia in rhesus monkeys. Investig. Ophthalmol. Vis. Sci. 2012, 53, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, E.G.; Park, H.N.; Chrenek, M.; He, L.; Sidhu, C.; Chakraborty, R.; Strickland, R.; Iuvone, P.M.; Pardue, M.T. Ambient Light Regulates Retinal Dopamine Signaling and Myopia Susceptibility. Investig. Ophthalmol. Vis. Sci. 2021, 62, 28. [Google Scholar] [CrossRef] [PubMed]
- Norton, T.T.; Amedo, A.O.; Siegwart, J.T., Jr. Darkness causes myopia in visually experienced tree shrews. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4700–4707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lan, W.; Yang, S.; Liao, Y.; Xu, Q.; Lin, L.; Yang, Z. The effect of spectral property and intensity of light on natural refractive development and compensation to negative lenses in guinea pigs. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6324–6332. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ding, H.; Stell, W.K.; Liu, L.; Li, S.; Liu, H.; Zhong, X. Exposure to sunlight reduces the risk of myopia in rhesus monkeys. PLoS ONE 2015, 10, e0127863. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Zhang, F.; Zhao, Y.; Sun, M.; Tao, J.; Liang, Y.; Ma, L.; Yu, Y.; Wang, J.; Hao, J. Enlargement of the Axial Length and Altered Ultrastructural Features of the Sclera in a Mutant Lumican Transgenic Mouse Model. PLoS ONE 2016, 11, e0163165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardue, M.T.; Stone, R.A.; Iuvone, P.M. Investigating mechanisms of myopia in mice. Exp. Eye Res. 2013, 114, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Qazi, Y.; Tan, C.; Jabbar, S.B.; Cao, Y.; Schmid, G.; Pardue, M.T. Assessment of axial length measurements in mouse eyes. Optom. Vis. Sci. 2012, 89, 296–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavente-Perez, A.; Nour, A.; Troilo, D. Axial eye growth and refractive error development can be modified by exposing the peripheral retina to relative myopic or hyperopic defocus. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6765–6773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmucker, C.; Schaeffel, F. In vivo biometry in the mouse eye with low coherence interferometry. Vis. Res. 2004, 44, 2445–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczerkowska, K.I.; Petrezselyova, S.; Lindovsky, J.; Palkova, M.; Dvorak, J.; Makovicky, P.; Fang, M.; Jiang, C.; Chen, L.; Shi, M.; et al. Myopia disease mouse models: A missense point mutation (S673G) and a protein-truncating mutation of the Zfp644 mimic human disease phenotype. Cell Biosci. 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Atchison, D.A.; Jones, C.E.; Schmid, K.L.; Pritchard, N.; Pope, J.M.; Strugnell, W.E.; Riley, R.A. Eye shape in emmetropia and myopia. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3380–3386. [Google Scholar] [CrossRef] [Green Version]
- Cruickshank, F.E.; Logan, N.S. Optical ‘dampening’ of the refractive error to axial length ratio: Implications for outcome measures in myopia control studies. Ophthalmic Physiol. Opt. J. Br. Coll. Ophthalmic Opt. 2018, 38, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Pardue, M.T.; Faulkner, A.E.; Fernandes, A.; Yin, H.; Schaeffel, F.; Williams, R.W.; Pozdeyev, N.; Iuvone, P.M. High susceptibility to experimental myopia in a mouse model with a retinal on pathway defect. Investig. Ophthalmol. Vis. Sci. 2008, 49, 706–712. [Google Scholar] [CrossRef]
- Park, H.; Tan, C.C.; Faulkner, A.; Jabbar, S.B.; Schmid, G.; Abey, J.; Iuvone, P.M.; Pardue, M.T. Retinal degeneration increases susceptibility to myopia in mice. Mol. Vis. 2013, 19, 2068–2079. [Google Scholar]
- Chakraborty, R.; Lacy, K.D.; Tan, C.C.; Park, H.N.; Pardue, M.T. Refractive index measurement of the mouse crystalline lens using optical coherence tomography. Exp. Eye Res. 2014, 125, 62–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffel, F.; Burkhardt, E.; Howland, H.C.; Williams, R.W. Measurement of refractive state and deprivation myopia in two strains of mice. Optom. Vis. Sci. 2004, 81, 99–110. [Google Scholar] [CrossRef]
- Tejedor, J.; de la Villa, P. Refractive changes induced by form deprivation in the mouse eye. Investig. Ophthalmol. Vis. Sci. 2003, 44, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Tkatchenko, A.V.; Tkatchenko, T.V.; Guggenheim, J.A.; Verhoeven, V.J.; Hysi, P.G.; Wojciechowski, R.; Singh, P.K.; Kumar, A.; Thinakaran, G.; Williams, C.; et al. APLP2 Regulates Refractive Error and Myopia Development in Mice and Humans. PLoS Genet 2015, 11, e1005432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhi, Z.; Ruan, Q.; Liu, Q.; Li, F.; Wan, F.; Reinach, P.S.; Chen, J.; Qu, J.; Zhou, X. Bright Light Suppresses Form-Deprivation Myopia Development With Activation of Dopamine D1 Receptor Signaling in the ON Pathway in Retina. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2306–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, Y.; Peleg, E.; Belkin, M.; Polat, U.; Solomon, A.S. Ambient illuminance, retinal dopamine release and refractive development in chicks. Exp. Eye Res. 2012, 103, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Schaeffel, F.; Kohler, K.; Zrenner, E. Dose-dependent effects of 6-hydroxy dopamine on deprivation myopia, electroretinograms, and dopaminergic amacrine cells in chickens. Vis. Neurosci. 1992, 9, 483–492. [Google Scholar] [CrossRef]
- Wu, X.H.; Qian, K.W.; Xu, G.Z.; Li, Y.Y.; Ma, Y.Y.; Huang, F.; Wang, Y.Q.; Zhou, X.; Qu, J.; Yang, X.L.; et al. The Role of Retinal Dopamine in C57BL/6 Mouse Refractive Development as Revealed by Intravitreal Administration of 6-Hydroxydopamine. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5393–5404. [Google Scholar] [CrossRef]
- Stone, R.A.; Lin, T.; Laties, A.M.; Iuvone, P.M. Retinal dopamine and form-deprivation myopia. Proc. Natl. Acad. Sci. USA 1989, 86, 704–706. [Google Scholar] [CrossRef] [Green Version]
- Nickla, D.L.; Totonelly, K. Dopamine antagonists and brief vision distinguish lens-induced- and form-deprivation-induced myopia. Exp. Eye Res. 2011, 93, 782–785. [Google Scholar] [CrossRef] [Green Version]
- Hirasawa, H.; Contini, M.; Raviola, E. Extrasynaptic release of GABA and dopamine by retinal dopaminergic neurons. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140186. [Google Scholar] [CrossRef] [Green Version]
- Boatright, J.H.; Hoel, M.J.; Iuvone, P.M. Stimulation of endogenous dopamine release and metabolism in amphibian retina by light- and K+-evoked depolarization. Brain Res. 1989, 482, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Feldkaemper, M.; Schaeffel, F. An updated view on the role of dopamine in myopia. Exp. Eye Res. 2013, 114, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Pardue, M.T.; Iuvone, P.M.; Qu, J. Dopamine signaling and myopia development: What are the key challenges. Prog. Retin. Eye Res. 2017, 61, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Park, H.N.; Hanif, A.M.; Sidhu, C.S.; Iuvone, P.M.; Pardue, M.T. ON pathway mutations increase susceptibility to form-deprivation myopia. Exp. Eye Res. 2015, 137, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Kurihara, T.; Kunimi, H.; Miyauchi, M.; Ikeda, S.-I.; Mori, K.; Tsubota, K.; Torii, H.; Tsubota, K. A highly efficient murine model of experimental myopia. Sci. Rep. 2018, 8, 2026. [Google Scholar] [CrossRef] [Green Version]
- Morgan, I.G.; Ashby, R.S.; Nickla, D.L.; Guggenheim, J.A. Form deprivation and lens-induced myopia: Are they different? Ophthalmic Physiol. Opt. J. Br. Coll. Ophthalmic Opt. 2013, 33, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, K.; Karouta, C.; Ashby, R. Form-Deprivation and Lens-Induced Myopia Are Similarly Affected by Pharmacological Manipulation of the Dopaminergic System in Chicks. Investig. Ophthalmol. Vis. Sci. 2020, 61, 4. [Google Scholar] [CrossRef] [PubMed]
- Thomson, K.; Morgan, I.; Karouta, C.; Ashby, R. Levodopa inhibits the development of lens-induced myopia in chicks. Sci. Rep. 2020, 10, 13242. [Google Scholar] [CrossRef] [PubMed]
- Zeitz, C.; Robson, A.G.; Audo, I. Congenital stationary night blindness: An analysis and update of genotype–phenotype correlations and pathogenic mechanisms. Prog. Retin. Eye Res. 2015, 45, 58–110. [Google Scholar] [CrossRef]
- Bijveld, M.M.; Florijn, R.J.; Bergen, A.A.; van den Born, L.I.; Kamermans, M.; Prick, L.; Riemslag, F.C.; van Schooneveld, M.J.; Kappers, A.M.; van Genderen, M.M. Genotype and phenotype of 101 dutch patients with congenital stationary night blindness. Ophthalmology 2013, 120, 2072–2081. [Google Scholar] [CrossRef]
- Zeitz, C. Molecular genetics and protein function involved in nocturnal vision. Expert Rev. Ophthalmol. 2007, 2, 467–485. [Google Scholar] [CrossRef]
- Dry, K.L.; Van Dorp, D.B.; Aldred, M.A.; Brown, J.; Hardwick, L.J.; Wright, A.F. Linkage analysis in a family with complete type congenital stationary night blindness with and without myopia. Clin. Genet. 1993, 43, 250–254. [Google Scholar] [CrossRef]
- Zeitz, C.; Labs, S.; Lorenz, B.; Forster, U.; Uksti, J.; Kroes, H.Y.; De Baere, E.; Leroy, B.P.; Cremers, F.P.; Wittmer, M.; et al. Genotyping microarray for CSNB-associated genes. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5919–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, Y.; Yagasaki, K.; Horiguchi, M.; Kawase, Y.; Kanda, T. Congenital stationary night blindness with negative electroretinogram. A new classification. Arch. Ophthalmol. 1986, 104, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Bijveld, M.M.; van Genderen, M.M.; Hoeben, F.P.; Katzin, A.A.; van Nispen, R.M.; Riemslag, F.C.; Kappers, A.M. Assessment of night vision problems in patients with congenital stationary night blindness. PLoS ONE 2013, 8, e62927. [Google Scholar] [CrossRef] [Green Version]
- Audo, I.; Robson, A.G.; Holder, G.E.; Moore, A.T. The negative ERG: Clinical phenotypes and disease mechanisms of inner retinal dysfunction. Surv. Ophthalmol. 2008, 53, 16–40. [Google Scholar] [CrossRef] [PubMed]
- Audo, I.; Bujakowska, K.; Orhan, E.; Poloschek, C.M.; Defoort-Dhellemmes, S.; Drumare, I.; Kohl, S.; Luu, T.D.; Lecompte, O.; Zrenner, E.; et al. Whole-exome sequencing identifies mutations in GPR179 leading to autosomal-recessive complete congenital stationary night blindness. Am. J. Hum. Genet. 2012, 90, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Audo, I.; Kohl, S.; Leroy, B.P.; Munier, F.L.; Guillonneau, X.; Mohand-Saïd, S.; Bujakowska, K.; Nandrot, E.F.; Lorenz, B.; Preising, M.; et al. TRPM1 is mutated in patients with autosomal-recessive complete congenital stationary night blindness. Am. J. Hum. Genet. 2009, 85, 720–729. [Google Scholar] [CrossRef] [Green Version]
- Zeitz, C.; Jacobson, S.G.; Hamel, C.P.; Bujakowska, K.; Neuillé, M.; Orhan, E.; Zanlonghi, X.; Lancelot, M.E.; Michiels, C.; Schwartz, S.B.; et al. Whole-exome sequencing identifies LRIT3 mutations as a cause of autosomal-recessive complete congenital stationary night blindness. Am. J. Hum. Genet. 2013, 92, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Bech-Hansen, N.T.; Naylor, M.J.; Maybaum, T.A.; Sparkes, R.L.; Koop, B.; Birch, D.G.; Bergen, A.A.; Prinsen, C.F.; Polomeno, R.C.; Gal, A.; et al. Mutations in NYX, encoding the leucine-rich proteoglycan nyctalopin, cause X-linked complete congenital stationary night blindness. Nat. Genet. 2000, 26, 319–323. [Google Scholar] [CrossRef]
- Peachey, N.S.; Ray, T.A.; Florijn, R.; Rowe, L.B.; Sjoerdsma, T.; Contreras-Alcantara, S.; Baba, K.; Tosini, G.; Pozdeyev, N.; Iuvone, P.M.; et al. GPR179 is required for depolarizing bipolar cell function and is mutated in autosomal-recessive complete congenital stationary night blindness. Am. J. Hum. Genet. 2012, 90, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Sergouniotis, P.I.; Michaelides, M.; Mackay, D.S.; Wright, G.A.; Devery, S.; Moore, A.T.; Holder, G.E.; Robson, A.G.; Webster, A.R. Recessive mutations of the gene TRPM1 abrogate ON bipolar cell function and cause complete congenital stationary night blindness in humans. Am. J. Hum. Genet. 2009, 85, 711–719. [Google Scholar] [CrossRef]
- Van Genderen, M.M.; Bijveld, M.M.; Claassen, Y.B.; Florijn, R.J.; Pearring, J.N.; Meire, F.M.; McCall, M.A.; Riemslag, F.C.; Gregg, R.G.; Bergen, A.A.; et al. Mutations in TRPM1 are a common cause of complete congenital stationary night blindness. Am. J. Hum. Genet. 2009, 85, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryja, T.P.; McGee, T.L.; Berson, E.L.; Fishman, G.A.; Sandberg, M.A.; Alexander, K.R.; Derlacki, D.J.; Rajagopalan, A.S. Night blindness and abnormal cone electroretinogram ON responses in patients with mutations in the GRM6 gene encoding mGluR6. Proc. Natl. Acad. Sci. USA 2005, 102, 4884–4889. [Google Scholar] [CrossRef] [Green Version]
- Pusch, C.M.; Zeitz, C.; Brandau, O.; Pesch, K.; Achatz, H.; Feil, S.; Scharfe, C.; Maurer, J.; Jacobi, F.K.; Pinckers, A.; et al. The complete form of X-linked congenital stationary night blindness is caused by mutations in a gene encoding a leucine-rich repeat protein. Nat. Genet. 2000, 26, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Zeitz, C.; van Genderen, M.; Neidhardt, J.; Luhmann, U.F.; Hoeben, F.; Forster, U.; Wycisk, K.; Mátyás, G.; Hoyng, C.B.; Riemslag, F.; et al. Mutations in GRM6 cause autosomal recessive congenital stationary night blindness with a distinctive scotopic 15-Hz flicker electroretinogram. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4328–4335. [Google Scholar] [CrossRef] [Green Version]
- Gregg, R.G.; Mukhopadhyay, S.; Candille, S.I.; Ball, S.L.; Pardue, M.T.; McCall, M.A.; Peachey, N.S. Identification of the gene and the mutation responsible for the mouse nob phenotype. Investig. Ophthalmol. Vis. Sci. 2003, 44, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Pinto, L.H.; Vitaterna, M.H.; Shimomura, K.; Siepka, S.M.; McDearmon, E.L.; Balannik, V.; Omura, C.; Lumayag, S.; Invergo, B.M.; Glawe, B.; et al. Generation, Identification and Functional Characterization of the nob4 Mutation of Grm6 in the Mouse. Vis. Neurosci. 2007, 24, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Masu, M.; Iwakabe, H.; Tagawa, Y.; Miyoshi, T.; Yamashita, M.; Fukuda, Y.; Sasaki, H.; Hiroi, K.; Nakamura, Y.; Shigemoto, R.; et al. Specific deficit of the ON response in visual transmission by targeted disruption of the mGluR6 gene. Cell 1995, 80, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Morgans, C.W.; Brown, R.L.; Duvoisin, R.M. TRPM1: The endpoint of the mGluR6 signal transduction cascade in retinal ON-bipolar cells. BioEssays News Rev. Mol. Cell. Dev. Biol. 2010, 32, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Morgans, C.W.; Zhang, J.; Jeffrey, B.G.; Nelson, S.M.; Burke, N.S.; Duvoisin, R.M.; Brown, R.L. TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proc. Natl. Acad. Sci. USA 2009, 106, 19174–19178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, M.; Sanuki, R.; Tetsuhiro, R.Y.; Onishi, A.; Nishiguchi, K.M.; Koike, C.; Kadowaki, M.; Kondo, M.; Miyake, Y.; Furukawa, T. TRPM1 mutations are associated with the complete form of congenital stationary night blindness. Mol. Vis. 2010, 16, 425–437. [Google Scholar] [PubMed]
- Orhan, E.; Neuillé, M.; de Sousa Dias, M.; Pugliese, T.; Michiels, C.; Condroyer, C.; Antonio, A.; Sahel, J.A.; Audo, I.; Zeitz, C. A New Mouse Model for Complete Congenital Stationary Night Blindness Due to Gpr179 Deficiency. Int. J. Mol. Sci. 2021, 22, 4424. [Google Scholar] [CrossRef] [PubMed]
- Orhan, E.; Prézeau, L.; El Shamieh, S.; Bujakowska, K.M.; Michiels, C.; Zagar, Y.; Vol, C.; Bhattacharya, S.S.; Sahel, J.A.; Sennlaub, F.; et al. Further insights into GPR179: Expression, localization, and associated pathogenic mechanisms leading to complete congenital stationary night blindness. Investig. Ophthalmol. Vis. Sci. 2013, 54, 8041–8050. [Google Scholar] [CrossRef] [Green Version]
- Peachey, N.S.; Pearring, J.N.; Bojang, P., Jr.; Hirschtritt, M.E.; Sturgill-Short, G.; Ray, T.A.; Furukawa, T.; Koike, C.; Goldberg, A.F.; Shen, Y.; et al. Depolarizing bipolar cell dysfunction due to a Trpm1 point mutation. J. Neurophysiol. 2012, 108, 2442–2451. [Google Scholar] [CrossRef] [Green Version]
- Neuillé, M.; Cao, Y.; Caplette, R.; Guerrero-Given, D.; Thomas, C.; Kamasawa, N.; Sahel, J.A.; Hamel, C.P.; Audo, I.; Picaud, S.; et al. LRIT3 Differentially Affects Connectivity and Synaptic Transmission of Cones to ON- and OFF-Bipolar Cells. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1768–1778. [Google Scholar] [CrossRef] [Green Version]
- Neuillé, M.; El Shamieh, S.; Orhan, E.; Michiels, C.; Antonio, A.; Lancelot, M.-E.; Condroyer, C.; Bujakowska, K.; Poch, O.; Sahel, J.-A.; et al. Lrit3 Deficient Mouse (nob6): A Novel Model of Complete Congenital Stationary Night Blindness (cCSNB). PLoS ONE 2014, 9, e90342. [Google Scholar] [CrossRef] [Green Version]
- Neuillé, M.; Morgans, C.W.; Cao, Y.; Orhan, E.; Michiels, C.; Sahel, J.A.; Audo, I.; Duvoisin, R.M.; Martemyanov, K.A.; Zeitz, C. LRIT3 is essential to localize TRPM1 to the dendritic tips of depolarizing bipolar cells and may play a role in cone synapse formation. Eur. J. Neurosci. 2015, 42, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Hasan, N.; Pangeni, G.; Ray, T.A.; Fransen, K.M.; Noel, J.; Borghuis, B.G.; McCall, M.A.; Gregg, R.G. LRIT3 is Required for Nyctalopin Expression and Normal ON and OFF Pathway Signaling in the Retina. eNeuro 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, J.; Hasan, N.; Bell, B.A.; Peachey, N.S. Reduced expression of the nob8 gene does not normalize the distribution or function of mGluR6 in the mouse retina. Mol. Vis. 2019, 25, 890–901. [Google Scholar] [PubMed]
- Orlandi, C.; Omori, Y.; Wang, Y.; Cao, Y.; Ueno, A.; Roux, M.J.; Condomitti, G.; de Wit, J.; Kanagawa, M.; Furukawa, T.; et al. Transsynaptic Binding of Orphan Receptor GPR179 to Dystroglycan-Pikachurin Complex Is Essential for the Synaptic Organization of Photoreceptors. Cell Rep. 2018, 25, 130–145.e5. [Google Scholar] [CrossRef] [Green Version]
- Koike, C.; Obara, T.; Uriu, Y.; Numata, T.; Sanuki, R.; Miyata, K.; Koyasu, T.; Ueno, S.; Funabiki, K.; Tani, A.; et al. TRPM1 is a component of the retinal ON bipolar cell transduction channel in the mGluR6 cascade. Proc. Natl. Acad. Sci. USA 2010, 107, 332–337. [Google Scholar] [CrossRef]
- Maddox, D.M.; Vessey, K.A.; Yarbrough, G.L.; Invergo, B.M.; Cantrell, D.R.; Inayat, S.; Balannik, V.; Hicks, W.L.; Hawes, N.L.; Byers, S.; et al. Allelic variance between GRM6 mutants, Grm6nob3 and Grm6nob4 results in differences in retinal ganglion cell visual responses. J. Physiol. 2008, 586, 4409–4424. [Google Scholar] [CrossRef]
- Qian, H.; Ji, R.; Gregg, R.G.; Peachey, N.S. Identification of a new mutant allele, Grm6(nob7), for complete congenital stationary night blindness. Vis. Neurosci. 2015, 32, E004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardue, M.T.; McCall, M.A.; LaVail, M.M.; Gregg, R.G.; Peachey, N.S. A naturally occurring mouse model of X-linked congenital stationary night blindness. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2443–2449. [Google Scholar]
- Chakraborty, R.; Yang, V.; Park, H.N.; Landis, E.G.; Dhakal, S.; Bergen, M.A.; Iuvone, P.M.; Pardue, M.T. Lack of cone mediated retinal function increases susceptibility to form-deprivation myopia in mice. Exp. Eye Res. 2019, 180, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Fain, G.; Sampath, A.P. Rod and cone interactions in the retina. F1000Research 2018, 7, F1000. [Google Scholar] [CrossRef]
- Tsukamoto, Y.; Omi, N. Classification of Mouse Retinal Bipolar Cells: Type-Specific Connectivity with Special Reference to Rod-Driven AII Amacrine Pathways. Front. Neuroanat. 2017, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Field, G.D. Dopaminergic modulation of retinal processing from starlight to sunlight. J. Pharmacol. Sci. 2019, 140, 86–93. [Google Scholar] [CrossRef]
- Iuvone, P.M.; Tigges, M.; Fernandes, A.; Tigges, J. Dopamine synthesis and metabolism in rhesus monkey retina: Development, aging, and the effects of monocular visual deprivation. Vis. Neurosci. 1989, 2, 465–471. [Google Scholar] [CrossRef]
- Stone, R.A.; Lin, T.; Laties, A.M. Muscarinic antagonist effects on experimental chick myopia. Exp. Eye Res. 1991, 52, 755–758. [Google Scholar] [CrossRef]
- Feldkaemper, M.P.; Schaeffel, F. Evidence for a potential role of glucagon during eye growth regulation in chicks. Vis. Neurosci. 2002, 19, 755–766. [Google Scholar] [CrossRef]
- Troilo, D.; Nickla, D.L.; Mertz, J.R.; Summers Rada, J.A. Change in the synthesis rates of ocular retinoic acid and scleral glycosaminoglycan during experimentally altered eye growth in marmosets. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Fujikado, T.; Kawasaki, Y.; Fujii, J.; Taniguchi, N.; Okada, M.; Suzuki, A.; Ohmi, G.; Tano, Y. The effect of nitric oxide synthase inhibitor on form-deprivation myopia. Curr. Eye Res. 1997, 16, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Mertz, J.R.; Wallman, J. Choroidal retinoic acid synthesis: A possible mediator between refractive error and compensatory eye growth. Exp. Eye Res. 2000, 70, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, E.G.; Chrenek, M.A.; Chakraborty, R.; Strickland, R.; Bergen, M.; Yang, V.; Iuvone, P.M.; Pardue, M.T. Increased endogenous dopamine prevents myopia in mice. Exp. Eye Res. 2020, 193, 107956. [Google Scholar] [CrossRef] [PubMed]
- Boatright, J.H.; Gordon, J.R.; Iuvone, P.M. Inhibition of endogenous dopamine release in amphibian retina by L-2-amino-4-phosphonobutyric acid (L-AP4) and trans-2-aminocyclopentane-1,3-dicarboxylate (ACPD). Brain Res. 1994, 649, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.H.; Hogan, K.; Mazade, R.E.; Park, H.N.; Sidhu, C.S.; Iuvone, P.M.; Pardue, M.T. ON than OFF pathway disruption leads to greater deficits in visual function and retinal dopamine signaling. Exp. Eye Res. 2022, 220, 109091. [Google Scholar] [CrossRef]
- Park, H.; Jabbar, S.B.; Tan, C.C.; Sidhu, C.S.; Abey, J.; Aseem, F.; Schmid, G.; Iuvone, P.M.; Pardue, M.T. Visually-driven ocular growth in mice requires functional rod photoreceptors. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6272–6279. [Google Scholar] [CrossRef]
- Leifert, D.; Todorova, M.G.; Prünte, C.; Palmowski-Wolfe, A.M. LED-generated multifocal ERG on- and off-responses in complete congenital stationary night blindness—A case report. Doc. Ophthalmol. Adv. Ophthalmol. 2005, 111, 1–6. [Google Scholar] [CrossRef]
- Takeuchi, H.; Horie, S.; Moritoh, S.; Matsushima, H.; Hori, T.; Kimori, Y.; Kitano, K.; Tsubo, Y.; Tachibana, M.; Koike, C. Different Activity Patterns in Retinal Ganglion Cells of TRPM1 and mGluR6 Knockout Mice. BioMed Res. Int. 2018, 2018, 2963232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtoshi, A.; Wang, S.W.; Maeda, H.; Saszik, S.M.; Frishman, L.J.; Klein, W.H.; Behringer, R.R. Regulation of retinal cone bipolar cell differentiation and photopic vision by the CVC homeobox gene Vsx1. Curr. Biol. 2004, 14, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, R.; Park, H.; Aung, M.H.; Tan, C.C.; Sidhu, C.S.; Iuvone, P.M.; Pardue, M.T. Comparison of refractive development and retinal dopamine in OFF pathway mutant and C57BL/6J wild-type mice. Mol. Vis. 2014, 20, 1318–1327. [Google Scholar] [PubMed]
- Chakraborty, R.; Pardue, M.T. Molecular and Biochemical Aspects of the Retina on Refraction. Prog. Mol. Biol. Transl. Sci. 2015, 134, 249–267. [Google Scholar]
- Tkatchenko, T.V.; Shen, Y.; Tkatchenko, A.V. Analysis of postnatal eye development in the mouse with high-resolution small animal magnetic resonance imaging. Investig. Ophthalmol. Vis. Sci. 2010, 51, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, L.; Chen, R.; Zhu, Y.; Wang, S.; Hou, X.; Wei, B.; Wang, Q.; Liu, Y.; Qu, J.; et al. A role of color vision in emmetropization in C57BL/6J mice. Sci. Rep. 2020, 10, 14895. [Google Scholar] [CrossRef]
- Tkatchenko, T.V.; Shah, R.L.; Nagasaki, T.; Tkatchenko, A.V. Analysis of genetic networks regulating refractive eye development in collaborative cross progenitor strain mice reveals new genes and pathways underlying human myopia. BMC Med. Genom. 2019, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, R.; Ostrin, L.A.; Nickla, D.L.; Iuvone, P.M.; Pardue, M.T.; Stone, R.A. Circadian rhythms, refractive development, and myopia. Ophthalmic Physiol. Opt. J. Br. Coll. Ophthalmic Opt. 2018, 38, 217–245. [Google Scholar] [CrossRef]
- Tkatchenko, T.V.; Tkatchenko, A.V. Ketamine-xylazine anesthesia causes hyperopic refractive shift in mice. J. Neurosci. Methods 2010, 193, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Glickstein, M.; Millodot, M. Retinoscopy and eye size. Science 1970, 168, 605–606. [Google Scholar] [CrossRef] [PubMed]
- Tkatchenko, T.V.; Shen, Y.; Tkatchenko, A.V. Mouse Experimental Myopia Has Features of Primate Myopia. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Ward, A.H.; Siegwart, J.T.; Frost, M.R.; Norton, T.T. Intravitreally-administered dopamine D2-like (and D4), but not D1-like, receptor agonists reduce form-deprivation myopia in tree shrews. Vis. Neurosci. 2017, 34, E003. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, J.; Reinach, P.S.; Wang, F.; Zhang, L.; Fan, M.; Ying, H.; Pan, M.; Qu, J.; Zhou, X. Dopamine Receptor Subtypes Mediate Opposing Effects on Form Deprivation Myopia in Pigmented Guinea Pigs. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4441–4448. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Zhi, Z.; Pan, M.; Xie, R.; Qin, X.; Lu, R.; Mao, X.; Chen, J.F.; Willcox, M.D.; Qu, J.; et al. Inhibition of experimental myopia by a dopamine agonist: Different effectiveness between form deprivation and hyperopic defocus in guinea pigs. Mol. Vis. 2011, 17, 2824–2834. [Google Scholar] [PubMed]
- Feldkaemper, M.; Diether, S.; Kleine, G.; Schaeffel, F. Interactions of spatial and luminance information in the retina of chickens during myopia development. Exp. Eye Res. 1999, 68, 105–115. [Google Scholar] [CrossRef]
- Wu, X.H.; Li, Y.Y.; Zhang, P.P.; Qian, K.W.; Ding, J.H.; Hu, G.; Weng, S.J.; Yang, X.L.; Zhong, Y.M. Unaltered retinal dopamine levels in a C57BL/6 mouse model of form-deprivation myopia. Investig. Ophthalmol. Vis. Sci. 2015, 56, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickla, D.L. Ocular diurnal rhythms and eye growth regulation: Where we are 50 years after Lauber. Exp. Eye Res. 2013, 114, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Qian, K.W.; Li, Y.Y.; Wu, X.H.; Gong, X.; Liu, A.L.; Chen, W.H.; Yang, Z.; Cui, L.J.; Liu, Y.F.; Ma, Y.Y.; et al. Altered Retinal Dopamine Levels in a Melatonin-proficient Mouse Model of Form-deprivation Myopia. Neurosci. Bull. 2022, 38, 992–1006. [Google Scholar] [CrossRef]
- Fujikado, T.; Hosohata, J.; Omoto, T. ERG of form deprivation myopia and drug induced ametropia in chicks. Curr. Eye Res. 1996, 15, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Crewther, D.P.; Crewther, S.G.; Xie, R.Z. Changes in eye growth produced by drugs which affect retinal ON or OFF responses to light. J. Ocul. Pharmacol. Ther. Off. J. Assoc. Ocul. Pharmacol. Ther. 1996, 12, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Gregg, R.G.; Kamermans, M.; Klooster, J.; Lukasiewicz, P.D.; Peachey, N.S.; Vessey, K.A.; McCall, M.A. Nyctalopin expression in retinal bipolar cells restores visual function in a mouse model of complete X-linked congenital stationary night blindness. J. Neurophysiol. 2007, 98, 3023–3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamache, P.H.; Kingery, M.L.; Acworth, I.N. Urinary metanephrine and normetanephrine determined without extraction by using liquid chromatography and coulometric array detection. Clin. Chem. 1993, 39, 1825–1830. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilmet, B.; Callebert, J.; Duvoisin, R.; Goulet, R.; Tourain, C.; Michiels, C.; Frederiksen, H.; Schaeffel, F.; Marre, O.; Sahel, J.A.; et al. Mice Lacking Gpr179 with Complete Congenital Stationary Night Blindness Are a Good Model for Myopia. Int. J. Mol. Sci. 2023, 24, 219. https://doi.org/10.3390/ijms24010219
Wilmet B, Callebert J, Duvoisin R, Goulet R, Tourain C, Michiels C, Frederiksen H, Schaeffel F, Marre O, Sahel JA, et al. Mice Lacking Gpr179 with Complete Congenital Stationary Night Blindness Are a Good Model for Myopia. International Journal of Molecular Sciences. 2023; 24(1):219. https://doi.org/10.3390/ijms24010219
Chicago/Turabian StyleWilmet, Baptiste, Jacques Callebert, Robert Duvoisin, Ruben Goulet, Christophe Tourain, Christelle Michiels, Helen Frederiksen, Frank Schaeffel, Olivier Marre, José Alain Sahel, and et al. 2023. "Mice Lacking Gpr179 with Complete Congenital Stationary Night Blindness Are a Good Model for Myopia" International Journal of Molecular Sciences 24, no. 1: 219. https://doi.org/10.3390/ijms24010219