
The Role of Deubiquitinating Enzyme in Head and Neck Squamous Cell Carcinoma

Department of Oral Bioscience, Tokushima University Graduate School of Biomedical Sciences, Tokushima 770-8504, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(1), 552; https://doi.org/10.3390/ijms24010552
Submission received: 7 December 2022
/
Revised: 20 December 2022
/
Accepted: 24 December 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Head and Neck Tumors: New Diagnostic, Prognostic and Therapeutic Opportunities)
Abstract
:Ubiquitination and deubiquitination are two popular ways for the post-translational modification of proteins. These two modifications affect intracellular localization, stability, and function of target proteins. The process of deubiquitination is involved in histone modification, cell cycle regulation, cell differentiation, apoptosis, endocytosis, autophagy, and DNA repair after damage. Moreover, it is involved in the processes of carcinogenesis and cancer development. In this review, we discuss these issues in understanding deubiquitinating enzyme (DUB) function in head and neck squamous cell carcinoma (HNSCC), and their potential therapeutic strategies for HNSCC patients are also discussed.
1. Ubiquitin System
Ubiquitin is a highly-conserved small regulatory protein that has been found in almost all tissues of eukaryotic organisms [1]. Ubiquitin was first identified in 1975. It performs its myriad functions through conjugation to a large range of target proteins. A variety of different modifications can occur. This discovery that ubiquitin can be attached to proteins and label them for destruction won the Nobel Prize for chemistry in 2004 [2]. The ubiquitin protein consists of 76 amino acids and has a molecular mass of about 8.5 kDa. Under the conditions where ATP provides energy, ubiquitin molecules bind to the target protein through the cascade catalytic reaction of ubiquitin-activating enzyme (E1), ubiquitin-conjugating enzyme (E2), and ubiquitin ligase (E3). The ubiquitinated target protein is recognized and degraded by 26S proteasome. At present, 2 E1s, about 50 E2s, and more than 600 E3s are known to be encoded by the human genome. The multiplexed complex cascade catalytic networks reflect the importance and complexity of the ubiquitin system [3].
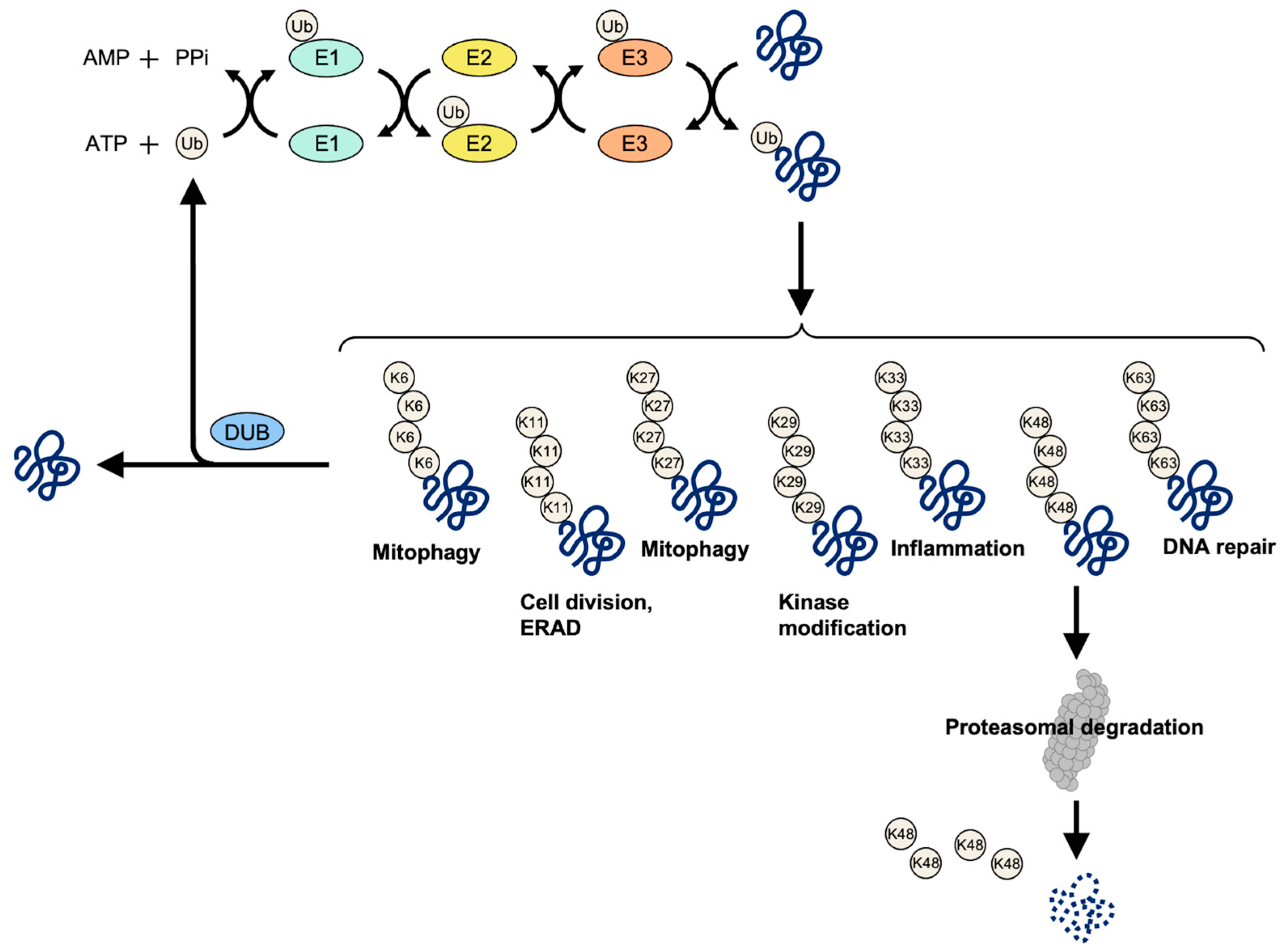
The C-terminal Gly residues of ubiquitin bind to the ε-amino groups of the target protein Lys residues via labeling amide bonds to the target protein. The ubiquitin itself also contains seven Lys residues: K6, K11, K27, K29, K33, K48, and K63. Ubiquitin can form polyubiquitin chains by connecting with their own Lys. Polymer chains are formed through a variety of connections between substrates and ubiquitin molecules. The chain can be short, containing only one ubiquitin molecule (mono-ubiquitin), or multiple sites on the substrate which are connected to a ubiquitin molecule (polyubiquitin), or it can be long. If it extends on the same Lys residue of a ubiquitin molecule, it is called a homogeneous ubiquitin chain (for example, K48- or a K63-linked polyubiquitin chain). If the ubiquitin molecule is connected with different Lys residues, the chain has a mixed topology, which is called a mixed ubiquitin chain [4]. All types of ubiquitin modifications can be detected in cells [5,6], and different types of ubiquitin modifications may lead to different results in cells, indicating that ubiquitin can be used as a code to store and transmit information. In general, polyubiquitin-labeled proteins that are linked by K48 are recognized and degraded by 26S proteasome. Additionally, the polyubiquitin of K63 connections affects the function of proteins more and participates in the process of endocytosis, DNA damage repair, activation of protein kinase, transformation of protein subcellular localization, some signal transduction, and stress response [7,8,9]. The polyubiquitination of the K6 connection can be involved in the regulation of microtubule stability and mitotic spindle orientation [10]. Polyubiquitin that is linked by K27 can participate in DNA damage repair and autoimmune regulation [11]. The polyubiquitination of K6 and K33 connections can be involved in DNA damage repair [12]. Polyubiquitin that is linked by K33 is also involved in the Type I interferon signaling pathway and innate immunity and vesicular transport [13]. The same type of ubiquitination may produce different cellular functions, and different types of ubiquitination may cooperate with each other to play a role in the same process or function, or one ubiquitination of a protein will promote the occurrence of another ubiquitination. Thus, the target protein gives a specific biological function via ubiquitination. The complex relationship between ubiquitin types and cell function leads to the abnormality of ubiquitin which may lead to complex diseases and tumors (Figure 1).
2. Deubiquitinating Enzymes (DUBs)
DUBs maintain ubiquitin system homeostasis by cleaving polyubiquitin chains or completely removing ubiquitin chains from ubiquitinated proteins. Via deubiquitination, free ubiquitins are generated and recycled [14]. Deubiquitination has important functions in regulating the ubiquitin-dependent pathways including cell cycle regulation, cell death, protein degradation, protein function, gene expression, and signal transduction [15]. So far, approximately 100 DUBs have been identified and classified as eight different families: ubiquitin-specific peptidases (UBP/USP) family, ubiquitin C-terminal hydroxylases (UCH) family, JAB1/MPN/MOV34 proteases (JAMM) family, ovarian tumor proteases (OTU) family, Machado–Joseph domain (MJD) family, motif interacting with Ub-containing novel DUB family (MINDY), monocyte chemotactic protein-induced proteins (MCPIP) family, and the Zn-finger and UFSP domain protein (ZUFSP) family [16,17] (Table 1). Among them, the USP, UCH, OTU, MJD, MCPIP, MINDY, and ZUFSP families belong to cysteine hydrolases, while the JAMM family belongs to metalloenzymes. The protein hydrolysis method that is adopted by most DUBs is to separate the ε-amino group of the target protein Lys residue from the carboxyl group of the C-terminal Gly of the ubiquitin molecule.
The USP family is the largest DUB family, which is called UBP family in yeast. The USP/UBP family is species- and tissue-specific, and the species and quantity vary across different organisms or different tissues. Members of the USP family contain three highly conserved domains: finger, palm, and thumb domains. The active catalytic center is in the palm and thumb domains, while the finger domain is used to capture the Gly residues at the end of ubiquitin molecules. Some USP domain activities are strongly inhibited by the slow dissociation of ubiquitin after substrate hydrolysis [18].
The UCH family was first discovered and reported as a member of the family of mercapto proteases, which mainly modify the ubiquitin chain. Its catalytic core region contains 230 amino acids. In the apo-enzyme structure of ubiquitin C-terminal hydrolase isozyme L3 (UCHL3), a prominent loop covers the active site. Surprisingly, in the ubiquitin aldehyde-bound structure, this active-site crossover loop straddles the C-terminal residues of the bound ubiquitin. Such an arrangement may prevent the binding of large and folded ubiquitin conjugates by fitting the substrates through this confined loop. Even polyubiquitin chains would be too big to be threaded through this confined loop [19]. Due to this unique spatial structure of UCH, it has a certain selectivity to the substrate. Therefore, members of the UCH family are only involved in the degradation of small peptides, such as those that are produced by proteasome and lysosomal pathways, or can also degrade protein precursors in the process of extension.
The JAMM/MPN family belongs to the metalloproteinase family. Members of the family usually couple two zinc ions, one in the active center and the other in the ubiquitin recognition region. The JAMM domain can recognize the tripeptide sequence Gln62-Lys63-Glu64. Therefore, the JAMM family can specifically recognize the ubiquitin chain that is connected by K63 [20]. The JAMM family is the only DUB that binds to ubiquitin molecules at the active site of the ubiquitin enzyme [21]. The JAMM family is not sensitive to DUB inhibitors but can be inhibited by metastatic cationic chelating agents.
The OTU family is divided into three subgroups: OTU, OTUB, and A20-like OTU. The members of the OTU family are not homologous to the USP and UCH families, but they have different specificities for different types of ubiquitin chains. For example, ubiquitin aldehyde binding 1 (OTUB1) can only hydrolyze the K48 ubiquitin chain. Ubiquitin aldehyde binding 2 (OTUB2) can hydrolyze the K63 and K48 ubiquitin chain. A20 can only hydrolyze K48 ubiquitin chain. Zinc finger RANBP2-type containing 1 (TRABID, also known as ZRANB1) can recognize the ubiquitin chain of K29 and K33 [22,23]. The activity conversion mode of the OTU family is similar to that of the USP family. When there is no ubiquitin binding, it is inactivated. When the ubiquitin molecule binds, it exposes the catalytic active site and activates enzyme activity [24].
The most widely studied member of the MJD family is ATXN3, a protein that is associated with degenerative diseases. ATXN3 has two ubiquitin binding sites, one of which is located at the active site and the other group is far away from the active site. Therefore, ATXN3 can hydrolyze the ubiquitin labeling of the substrate by interacting with the distal ubiquitin binding site to form a stable open concept. It recognizes the ubiquitin chains that are connected by K48 and K63, but it is more specific to K63 [25].
MCPIP is a newly discovered DUBs family. The first member of this family is zinc finger CCCH-type containing 12A (MCPIP1), At the N terminus, MCPIP1 contains a ubiquitin association domain (UBA). UBA mediates the association of MCPIP1 with ubiquitinated proteins, but it does not affect the enzymatic activity. In the middle, there is a conserved CCCH-type Zn finger domain (ZF), which is commonly responsible for its catalytic activity, and MCPIP1 contains a Pro-rich domain at the C terminus. There is no essential sequence similarity between MCPIP1 and the five DUB domains except remote homology with the UCH domain (27%), suggesting that MCPIP1 may contain a novel DUB domain [26].
MINDY and ZUFSP were discovered in 2016 and 2018, respectively [17,27,28]. Both have no homology with any known other DUB families. The MINDY family is highly selective at cleaving K48 ubiquitin chain [29]. MINDY may have specialized roles in regulating proteostasis, but ZUFSP is highly selective at cleaving the K63 ubiquitin chain and plays an important role in preventing DNA damage [30].
3. The Relationship between DUB and Tumorigenesis
Various DUBs have been reported to have connections to tumor-suppressing or oncogenic functions and may, therefore, represent potential therapeutic targets. The tumor suppressor p53 plays a critical role to preserve DNA fidelity from diverse insults through the regulation of cell-cycle checkpoints, DNA repair, senescence, and apoptosis [31]. MDM2 proto-oncogene (MDM2), an E3 ubiquitin ligase, is the main regulator of the p53 protein [32]. It can induce p53 to enucleate and degrade. Additionally, a cytoplasmic DUB, ubiquitin-specific peptidase 10 (USP10) deubiquitinates p53 and prevents Mdm2-induced p53 nuclear export and degradation. After DNA damage, USP10 is stabilized and translocated to the nucleus to activate p53 [33]. When subjected to external pressure, ubiquitin-specific peptidase 42 (USP42) can bind to p53 and stabilize its protein by removing the ubiquitin chain. Thus, the quick activation of the p53 signaling pathway causes the cell cycle arrest and carries out DNA repair for preventing carcinogenesis [34]. Ubiquitin-specific peptidase 15 (USP15) can stabilize MDM2, as well as degrade transcription factor NFATc2, to regulate T-cell activation. The deletion of USP15 activates T-cells and enhances their response to bacterial infection and tumorigenesis [35]. OTU deubiquitinase 1 (OTUD1) is a member of the OTU domain DUB family, which can directly inhibit the ubiquitination of p53 that is mediated by MDM2. It can stabilize and activate p53, resulting in p53-dependent cell proliferation inhibition and apoptosis. DNA damage enhances the interaction between OTUD1 and p53, so OTUD1 primarily plays a role in the G2/M and S checkpoints of the cell cycle [36]. OTUD1 inhibits DNA repair by inhibiting the activity of Ubc13/RNF168. This control of ring finger protein 168 (RNF168) affects the ubiquitination of chromatin. When DNA is damaged, OTUD1 is temporarily dissociated from the Ubc13/RNF168 complex, causing RNF168 to catalyze the ubiquitination of chromatin at DNA damage sites. At this site, it is associated with the UbcH5/MDM2 complex, which inhibits MDM2-dependent ubiquitination of p53, and p53 becomes stable and activated [37]. OTU deubiquitinase 5 (OTUD5) can also deubiquitinate p53 protein and stabilize it, thus affecting apoptosis that is induced by DNA damage [38].
Ubiquitin-specific peptidase 4 (USP4) is an important p53 regulatory molecule, which directly binds to ARF-BP1 and removes ubiquitin, resulting in the stabilization of the ARF-BP1 complex and subsequent decrease of p53 levels, and inhibits p53-related apoptosis. The absence of USP4 is thought to promote senescence. USP4 is highly expressed in various cancers, including bladder, prostate, and thyroid, indicating that it is a potential oncogene [39]. Ubiquitin-specific peptidase 7 (USP7) is also a p53 regulatory factor, which is able to deubiquitinate and stabilize p53 [22].
Transcription factor KLF transcription factor 5 (KLF5) is a member of the BAP1/HCF-1 complex, which is expressed in breast cancer tissues and promotes cancer cell proliferation and metastasis. BRCA1-associated protein 1 (BAP1) stabilizes KLF5 by removing the ubiquitin chain, thereby promoting the occurrence and metastasis of breast cancer. BAP1 depletion can inhibit the growth and metastasis of hepatocellular carcinoma cells [40].
Cell division cycle 25A (Cdc25A) is a phosphatase that promotes cell cycle progression by activating cyclin-dependent kinase (CDK), which has the potential to promote cancer. Cdc25A is highly expressed in tumor tissues and can be stabilized by ubiquitin-specific peptidase 17 like family member 2 (Dub3). Overexpression of Dub3 induces cell cycle arrest at S and G2 phases and activates the DNA damage response via stabilization of Cdc25A protein. Higher expression of Dub3 promotes the high levels of Cdc25A in breast cancer [41].
Phosphatase and tensin homolog (PTEN) is a tumor suppressor gene, which is deleted in many tumors. Ubiquitin-specific peptidase 13 (USP13) can deubiquitinate the PTEN protein and stabilize it. In breast cancer cells without USP13, the PTEN protein was downregulated, while Akt protein phosphorylation, cell proliferation, soft agar colony formation, glycolysis, and tumor growth were significantly enhanced [42,43]. OTU deubiquitinase 3 (OTUD3) stabilizes the PTEN protein via cleaving its ubiquitin chain. OTUD3 depletion activates the Akt signaling pathway, cell transformation, and metastasis. Reduced expression of OTUD3 is involved in carcinogenesis of breast cancer via the downregulation of PTEN protein [34,42,44,45].
Ubiquitin-specific peptidase 22 (USP22) participates in histone deubiquitination and acetylation, thus affecting gene transcription. It affects tumorigenesis and metastasis through its effects on BMI-1, MYC, FBP1, and TRF1.
Thus, there are many different functions of DUBs, which directly affect the stability of many kinds of proteins in cells and are closely related to the occurrence and development of tumors. Currently, however, the research on DUBs is not deep enough, and the function of many DUBs has not been confirmed. By comprehensively understanding the relationship between DUBs and tumorigenesis, screening DUBs that are closely related to tumor progression, and thoroughly studying the action mechanism of DUBs in tumors, the targeted therapy of DUBs can be really applied to clinical anti-tumor therapy.
4. The Role and Mechanism of DUB in HNSCC
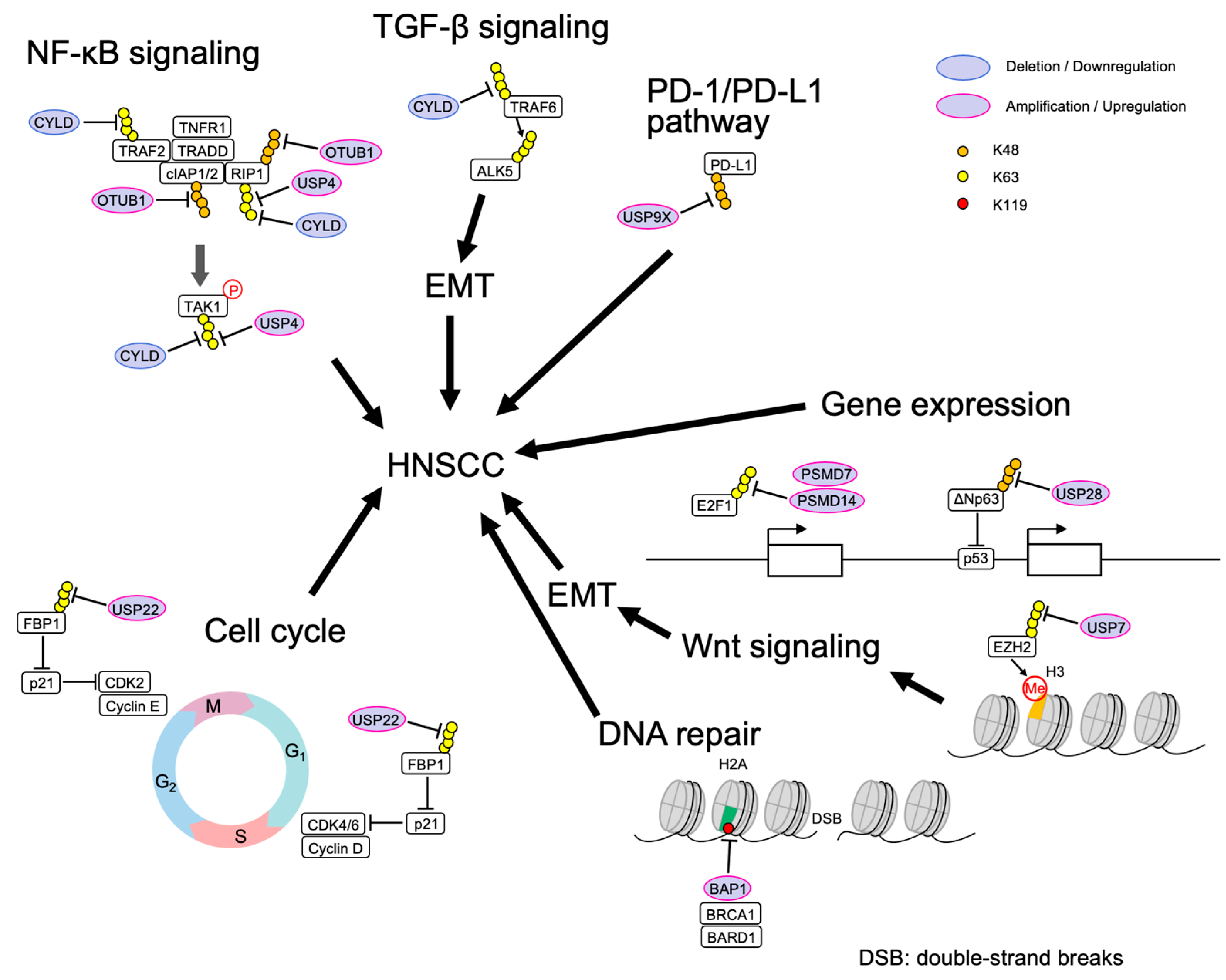
HNSCC was the seventh most common type of cancer worldwide in 2018 [46], representing about 6% of all cases and accounting for an estimated 650,000 new cancer cases and 350,000 cancer deaths worldwide every year [47]. The pathogenic factors include tobacco and alcohol intake, human papillomavirus (HPV) or Epstein Barr virus (EBV) infection, radiation, periodontal disease, vitamin deficiency, and eating habits [48]. Achieving a better understanding of molecular aberrations that are associated with HNSCC might identify new diagnostic and therapeutic strategies for this disease. (Table 2, Figure 2) [49,50,51,52,53,54,55,56,57,58,59,60,61,62].
5. The Role of DUBs in HNSCC Cell Proliferation and Apoptosis
Cell cycle regulation, uncontrolled proliferation, and inhibition of apoptosis will affect the occurrence and development of tumors. USP22, which has been described as a cancer stem cell (CSC) marker [68,69], is a member of the largest subfamily of DUBs [70]. The USP22 gene is located on chromosome 17 and consists of 14 exons, can indirectly affect chromatin structure through histone ubiquitination (H2A and H2B), thus regulating the transcriptional activation of many genes and widely affecting biological functions [66]. USP22 has the protein characteristics of CSCs and can promote the formation of various proteins for invasive tumor growth [71]. In addition, USP22 has been shown to regulate cellular growth and proliferation. Indeed, USP22 depletion reduces transformation of c-Myc and plays an important role in cell cycle progression and anchorage-independent growth [72]. Additionally, the downregulation of USP22 results in tumor cells remaining arrested at the G1 phase via p21 stabilization [73,74]. Thus, USP22 plays an important role in cell cycle regulation. The depletion of USP22 in HNSCC tissue can affect the CDK inhibitor (CDKI)/Rb signal pathway. USP22 increases the expression of cyclin-dependent kinase inhibitor 1A (CDKI) including p21 and p27, and it decreases the expression of Rb protein in HNSCC cells [74,75,76]. An immunohistochemical study showed that USP22 expression is upregulated in laryngeal carcinoma and has a close relationship with malignant behaviors including invasion, metastasis, and poor prognosis [77].
Ubiquitin-specific protease 9X (USP9X) is related to the occurrence and development of pancreatic cancer, multiple myeloma, and other tumors [78]. USP9X knockdown in HNSCC cells decreases the percentage of cells in the G0/G1 phase and increases the percentage of cells in the S and G2/M phase. The proliferation of HNSCC cells can be regulated by downstream transcription factor HES-1 (Notch pathway) through USP9X level [65]. USP9X may also regulate the proliferation of HNSCC through the mammalian target of the rapamycin (mTOR) pathway [77,79]. However, the loss of USP9X may be harmful to the growth of tumor cells, but its loss in primary tumor cells may accelerate the development of secondary tumors [80].
USP14 has been proven to be related to the occurrence and development of many cancers. In tumor xenograft mice, the growth of HNSCC with low USP14 was slower than that of the control group. The expression of USP14 in tongue carcinoma was higher than that in adjacent tissues. USP14 depletion can inhibit the proliferation and migration of HNSCC cells in vitro [81]. Moreover, USP14 can promote the proliferation and invasion of HNSCC both in vivo and in vitro.
Ubiquitin-specific peptidase 4 (USP4) has been identified to deubiquitinate K63-linked ubiquitin conjugates from TNF receptor-associated factor 2 (TRAF2), TNF receptor associated factor 6 (TRAF6), and TGF-beta activated kinase 1 (TAK1) and stabilizes molecules by deubiquitinating K48-linked ubiquitination [63]. USP4 is significantly upregulated in tumor tissues compared to matched non-tumor tissues in HNSCC. Co-immunoprecipitation showed that USP4 interacted with receptor-interacting protein 1 (RIPI) to remove K63-connected ubiquitin molecules to stabilize RIPI expression. Thus, it negatively regulates the activation of NF-κB mediated by deubiquitination of RIPI. These findings indicate that USP4 has tumor suppressor roles in HNSCC [63].
6. Relationship between DUBs and Prognosis of HNSCC
The prognosis of HNSCC is related to its stage of recurrence and lymph node metastasis. USP7 participates in intracellular tumor regulation, DNA repair, immune response, and epigenetic effects [82]. An immunohistochemical study showed that USP7 overexpression is correlated with histone-lysine N-methyltransferase (EZH2) in HNSCC [61]. EZH2 can promote the epithelial-mesenchymal transformation (EMT) in vivo and inhibits cellular senescence and differentiation [83]. EZH2 is widely expressed in various malignant tumors, including nasopharyngeal carcinoma [84]. USP7 expression in poorly differentiated HNSCC is higher than that in well differentiated HNSCC, and the high expression of USP7 was significantly related to a high degree of lymphatic invasion and high TNM stage. Survival analysis showed that USP7 overexpression is correlated with poor prognosis in HNSCC patients [61].
USP2a is one of the two splicing variants of USP2, which regulates the stability and function of many important cell growth and differentiation regulators and signal transduction factors in vivo [85]. In HNSCC, increased USP2a is associated with poor prognosis. The expression of ErbB2-interacting protein (Erbin) and USP2a increased in HNSCC, indicating that the expression of Erbin and USP2a was positively correlated with lymph node metastasis. Moreover, USP2a expression is related to epidermal growth factor (EGF) in HNSCC [86].
7. The Role of DUBs in Radiosensitivity of HNSCC
For the patients with HNSCC in the advanced stage, surgery combined with radiotherapy is needed to improve the survival rate of the patients. The radiation target and dose should be determined according to the location of the primary tumor, clinical-stage, postoperative pathological diagnosis, and postoperative imaging evaluation. The tolerance of HNSCC to radiotherapy is the main factor affecting the prognosis. BRCA1-associated protein-1 (BAP1), also known as UCHL2 can regulate various cellular processes including cell cycle, cell differentiation, transcription, DNA damage response, and resistance to tumor radiotherapy. BAP1 mutation can increase the sensitivity of HNSCC to radiotherapy and lead to tumor susceptibility syndrome, which is not related to the status of high-risk human papillomavirus (HPV) and p53 tumor suppressor protein. It was found that the survival fraction of BAP1 knockout HNSCC was significantly lower than that of HNSCC cells after radiotherapy, the sensitivity to radiotherapy was increased, and this sensitivity could be reversed by forced re-expression [67]. BAP1 can enhance the stability of histone H2A by deubiquitination and increase its expression level, thus interfering with the modification of chromatin and histone at the double strand break, reducing the rate of cell proliferation and increasing the sensitivity of radiotherapy [87]. HNSCC that is driven by HPV is more sensitive to DNA-damaging therapy than HPV-negative HNSCC. A recent study revealed that p16, the clinically used surrogate for HPV positivity, renders cells more sensitive to radiotherapy via the p16–HUWE1–USP7–TRIP12 pathway [88].
8. The Role of DUBs in Immune Mechanisms of HNSCC
Cancer is considered to be an immunosuppressive disease that is characterized by dysfunction of immune cells and disturbance of cytokine excretion [89]. EBV is closely related to the occurrence of HNSCC [90]. BPLF1 is the largest protein in EBV. There is deubiquitinating activity in the first 205 amino acids of EBV proteins. Its deubiquitinating activity can cleave K63 and K48 polyubiquitin chains. These two regulatory functions play a role in preventing virus protein degradation [91]. When the BPLF1 of EBV was knocked out, When the BPLF1 of EBV was knocked out, the replication quantity and infectivity of its genome decreased. The BPLF1 protein of EBV and its conserved deubiquitinating activity regulate cellular DNA repair enzyme polymerase and recruit it to potential virus destruction and replication sites, thus enhancing the ability to produce infectious viruses. BPLF1 can increase the fault tolerance rate of EBV virus after infecting human DNA by deubiquitinating and stabilizing EBV virus polymerase Pol, so that the DNA of the virus can continue to replicate in the injured site, thus increasing the infectivity of the virus [92]. It has been found that BPLF1 inhibits NF-κB signal transduction during lysozyme infection via deubiquitinating TNF receptor-related factor 6 (TRAF6) [93]. As HNSCC is closely related to EBV infection, the existence of BPLF1 may promote the HNSCC development.
USP9X upregulation can lead to programmed death receptor-ligand 1 (PD-L1) deubiquitination and PD-L1 stable expression in HNSCC tissues. PD-L1 inhibits T-cell activation and proliferation and allows cancer cells to escape T-cell immune surveillance. Preventing USP9X from locating PD-L1 may be an effective strategy for the treatment of HNSCC, especially metastatic tumors [64].
The conserved cylindromatosis (CYLD) belongs to the family of USPs, and its deletion can activate NF-κB and MAPK signaling pathways in HNSCC. NF-κB transcription factors can promote cell survival and induce innate and adaptive immunity to pathogens, such as viruses or bacterial pathogens, inflammatory cytokines, which are abnormally activated during the occurrence, and the development of cancer. The main target of CYLD is the classical NF-κB signal transduction that is mediated by NF-κB essential regulatory protein (NEMO) and cytokine reactive IκB kinase (IKK) complex subunit IKK-γ. CYLD deubiquitinates NEMO and prevents the phosphorylation of IκB and NF-κB activation [94]. The loss of CYLD in HNSCC can also increase the invasiveness of HNSCC by promoting transforming growth factor beta receptor 1 (ALK5) to stabilize NF-κB.
9. The Therapeutic Effect of DUBs on HNSCC
As DUBs have become a clinical target for tumor treatment, many people are paying more and more attention to the application of DUBs targeting drugs in cancer therapy. As most ubiquitin enzymes are cysteinases, it is easier to develop into a candidate drug. The effects of DUB inhibitors are as follows: (1) to increase the aggregation of polyubiquitin molecules, (2) to reduce the mono-ubiquitin part, and (3) to change some molecular activities, especially the persistence of carcinogenic proteins that are mediated by DUBs [16]. The inhibition of DUBs can lead to functional impairment of the proteasome and the accumulation of misfolded proteins, resulting in tumor cytotoxicity and death [95]. Some DUBs are associated with cancer and can be used as targets for new inhibitors [96,97]. At present, by screening the DUB inhibitor library, it was found that compound 2X-0324 had a potential radio-sensitizing effect on laryngeal cancer (RER > 1–1.86). Vif1 and Vif2 are protease inhibitors of USP7, which can regulate the anticancer effect that is mediated by p53 and have the potential to become molecular targeted drugs for the treatment of laryngeal carcinoma [98]. Azepan-4-ones and b-AP15 are protease inhibitors of USP14 and UCHL5. They can induce the accumulation of high molecular weight polyubiquitin proteins, resulting in the enhancement of the unfolded protein response (UPR). Both can inhibit the development of HNSCC [99]. In tongue cancer, b-AP15 can significantly inhibit the growth of tumor cells and increase apoptosis in a dose-dependent manner, thereby overcoming the drug resistance of bortezomib in vitro [100].
Squamous cell carcinoma of the head and neck is the most common malignant tumor of the head and neck, and its occurrence and development are related to a variety of DUBdeubiquitination enzymes [49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67]. Some DUBs are not only functional in head and neck squamous cell carcinomas but are also important in other squamous cell carcinomas. For example, CYLD is associated with HPV infection and inhibits NF-κB signaling in HNSCC, further promoting the growth and metastasis of head and neck cancer. The same functions of CYLD and HPV occur in cervical cancer cells [101]. In addition, USP46 is also associated with HPV infection in esophageal squamous cell carcinoma, but it has not been reported in HNSCC [102]. The Hippo/YAP signaling pathway plays an important role in HNSCC [103]. USP36 promotes proliferation and invasion of esophageal squamous cell carcinoma by deubiquitination of YAP [104]. USP36 may also play an important role in head and neck cancer.
10. Conclusions and Future Perspectives
In this review, we summarized the current information on the structure, regulation, and function of DUBs in tumor development, especially the association and treatment of DUBs and HNSCC. The following keywords were used to find the literature and the year in this paper: “deubiquitinating enzyme” and “head and neck cancer”. The literature was searched from 1997 to 2022 by PubMed.
DUBs can reverse the ubiquitination of the target protein, maintain the balance between ubiquitination and deubiquitination of the substrate protein, and maintain cell homeostasis by reducing the amount of deubiquitination in the target protein. DUBs have four different mechanisms: processing ubiquitin protein precursors, recovering ubiquitin molecules in the ubiquitin process, cutting ubiquitin protein chains, and reversing the binding of ubiquitin proteins to substrates [95]. DUBs regulate many cellular functions in vivo, including proteasome-dependent protein degradation [95], gene expression [105], cell cycle progression [106], chromosome segregation [107], DNA repair [108], and apoptosis [109]. The increased expression of USP22 and USP7 is related to the poor clinical prognosis of HNSCC and can be used as a biomarker to evaluate the prognosis of HNSCC. The expression of BAP1 can increase the radiotherapy tolerance in response to HNSCC and is a potential target to improve the radiosensitivity of HNSCC. The expression of USP9X, USP14, and other DUBs is increased in HNSCC, and it can promote the proliferation of cancer cells. The silencing of USP4 can inhibit the apoptosis of cancer cells and increase the persistent survival rate of HNSCC, as immunosuppressive diseases, UXP9X, CYLD1, and BPLF1 accelerate the development of HNSCC through the escape inhibition mechanism of immune cells or promoting the spread of virus infection. DUB is gradually being regarded as a therapeutic target for cancer because of its structural advantages. DUB inhibitors such as 2X-0324 and b-AP15 may serve as a new therapeutic strategy for HNSCC.
Our understanding of DUBs function, mechanism of action, regulation, and disease associations has made tremendous progress over the past decade. But DUBs-focused drug discovery has been challenging because the mechanism of action of DUB enzymes is often complex. The functions of DUBs are concentrated on the substrates that interact with DUBs. The same DUBs can have several or even dozens of substrates, and different DUBs can also have the same substrate, and the function of the substrates may interact with each other, which makes the design of predictive biochemical analysis and the development of drug-like compounds challenging. Although we can predict the existence of a wide range of therapeutic potentials for DUBs, a great deal of research is needed to successfully screen and evolve small molecule DUBs inhibitors in the future.
At present, a variety of literature has reported that DUB plays an important role in the occurrence and development of HNSCC, including abnormal expression in tumor cells and close correlation with radiotherapy and immune mechanism. Although the mechanism and downstream effects that are regulated by DUBs are not clear, as a potential target for the treatment of HNSCC, it is particularly important to better study the site of this regulatory mechanism and the side effects of targeted therapy. With the deepening of the study on the mechanism and function of DUBs in HNSCC, it is possible for DUBs to be used as a molecular target in cancer treatment.
Author Contributions
Conceptualization, S.J., Y.K., and T.H.; writing—original draft preparation, S.J.; writing—review and editing, Y.K. and T.H.; visualization, S.J. and T.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hershko, A.; Ciechanover, A. The ubiquitin system. Annu. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Wertz, I.E.; Wang, X. From Discovery to Bedside: Targeting the Ubiquitin System. Cell Chem. Biol. 2019, 26, 156–177. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The ubiquitin code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Schwartz, D.; Elias, J.E.; Thoreen, C.C.; Cheng, D.; Marsischky, G.; Roelofs, J.; Finley, D.; Gygi, S.P. A proteomics approach to understanding protein ubiquitination. Nat. Biotechnol. 2003, 21, 921–926. [Google Scholar] [CrossRef]
- Xu, D.; Yuan, L.; Liu, X.; Li, M.; Zhang, F.; Gu, X.Y.; Zhang, D.; Yang, Y.; Cui, B.; Tong, J.; et al. EphB6 overexpression and Apc mutation together promote colorectal cancer. Oncotarget 2016, 7, 31111–31121. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Riezman, H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science 2007, 315, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Gao, Y.; Li, L.; Jin, G.; Cai, Z.; Chao, J.I.; Lin, H.K. K63-linked ubiquitination in kinase activation and cancer. Front. Oncol. 2012, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.J.; Conze, D.B.; Li, T.; Srinivasula, S.M.; Ashwell, J.D. Sensing of Lys 63-linked polyubiquitination by NEMO is a key event in NF-kappaB activation [corrected]. Nat. Cell Biol. 2006, 8, 398–406. [Google Scholar] [CrossRef]
- Srivastava, D.; Chakrabarti, O. Mahogunin-mediated alpha-tubulin ubiquitination via noncanonical K6 linkage regulates microtubule stability and mitotic spindle orientation. Cell Death Dis. 2014, 5, e1064. [Google Scholar] [CrossRef]
- Liu, J.; Han, C.; Xie, B.; Wu, Y.; Liu, S.; Chen, K.; Xia, M.; Zhang, Y.; Song, L.; Li, Z.; et al. Rhbdd3 controls autoimmunity by suppressing the production of IL-6 by dendritic cells via K27-linked ubiquitination of the regulator NEMO. Nat. Immunol. 2014, 15, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.E.; Boardman, A.P.; Wang, D.C.; Huttlin, E.L.; Everley, R.A.; Dephoure, N.; Zhou, C.; Koren, I.; Gygi, S.P.; Elledge, S.J. Quantitative Proteomic Atlas of Ubiquitination and Acetylation in the DNA Damage Response. Mol. Cell 2015, 59, 867–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.; Zhao, Z.; Yang, Z.; Meng, Q.; Tan, P.; Xie, W.; Qin, Y.; Wang, R.F.; Cui, J. USP38 Inhibits Type I Interferon Signaling by Editing TBK1 Ubiquitination through NLRP4 Signalosome. Mol. Cell 2016, 64, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Farshi, P.; Deshmukh, R.R.; Nwankwo, J.O.; Arkwright, R.T.; Cvek, B.; Liu, J.; Dou, Q.P. Deubiquitinases (DUBs) and DUB inhibitors: A patent review. Expert Opin. Ther. Pat. 2015, 25, 1191–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfoh, R.; Lacdao, I.K.; Saridakis, V. Deubiquitinases and the new therapeutic opportunities offered to cancer. Endocr. Relat. Cancer 2015, 22, T35–T54. [Google Scholar] [CrossRef] [Green Version]
- D’Arcy, P.; Wang, X.; Linder, S. Deubiquitinase inhibition as a cancer therapeutic strategy. Pharmacol. Ther. 2015, 147, 32–54. [Google Scholar] [CrossRef] [Green Version]
- Abdul Rehman, S.A.; Kristariyanto, Y.A.; Choi, S.Y.; Nkosi, P.J.; Weidlich, S.; Labib, K.; Hofmann, K.; Kulathu, Y. MINDY-1 Is a Member of an Evolutionarily Conserved and Structurally Distinct New Family of Deubiquitinating Enzymes. Mol. Cell 2016, 63, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Clerici, M.; Luna-Vargas, M.P.; Faesen, A.C.; Sixma, T.K. The DUSP-Ubl domain of USP4 enhances its catalytic efficiency by promoting ubiquitin exchange. Nat. Commun. 2014, 5, 5399. [Google Scholar] [CrossRef] [Green Version]
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef]
- Hurley, J.H.; Stenmark, H. Molecular mechanisms of ubiquitin-dependent membrane traffic. Annu. Rev. Biophys 2011, 40, 119–142. [Google Scholar] [CrossRef]
- Davies, C.W.; Paul, L.N.; Kim, M.I.; Das, C. Structural and thermodynamic comparison of the catalytic domain of AMSH and AMSH-LP: Nearly identical fold but different stability. J. Mol. Biol. 2011, 413, 416–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eletr, Z.M.; Wilkinson, K.D. Regulation of proteolysis by human deubiquitinating enzymes. Biochim. Biophys. Acta 2014, 1843, 114–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Turcu, F.E.; Ventii, K.H.; Wilkinson, K.D. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu. Rev. Biochem. 2009, 78, 363–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelmann, M.J.; Iphofer, A.; Akutsu, M.; Altun, M.; di Gleria, K.; Kramer, H.B.; Fiebiger, E.; Dhe-Paganon, S.; Kessler, B.M. Structural basis and specificity of human otubain 1-mediated deubiquitination. Biochem. J. 2009, 418, 379–390. [Google Scholar] [CrossRef]
- Nicastro, G.; Masino, L.; Esposito, V.; Menon, R.P.; De Simone, A.; Fraternali, F.; Pastore, A. Josephin domain of ataxin-3 contains two distinct ubiquitin-binding sites. Biopolymers 2009, 91, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Saad, Y.; Lei, T.; Wang, J.; Qi, D.; Yang, Q.; Kolattukudy, P.E.; Fu, M. MCP-induced protein 1 deubiquitinates TRAF proteins and negatively regulates JNK and NF-kappaB signaling. J. Exp. Med. 2010, 207, 2959–2973. [Google Scholar] [CrossRef] [Green Version]
- Kwasna, D.; Abdul Rehman, S.A.; Natarajan, J.; Matthews, S.; Madden, R.; De Cesare, V.; Weidlich, S.; Virdee, S.; Ahel, I.; Gibbs-Seymour, I.; et al. Discovery and Characterization of ZUFSP/ZUP1, a Distinct Deubiquitinase Class Important for Genome Stability. Mol. Cell 2018, 70, 150–164.e6. [Google Scholar] [CrossRef] [Green Version]
- Haahr, P.; Borgermann, N.; Guo, X.; Typas, D.; Achuthankutty, D.; Hoffmann, S.; Shearer, R.; Sixma, T.K.; Mailand, N. ZUFSP Deubiquitylates K63-Linked Polyubiquitin Chains to Promote Genome Stability. Mol. Cell 2018, 70, 165–174.e6. [Google Scholar] [CrossRef] [Green Version]
- Kristariyanto, Y.A.; Abdul Rehman, S.A.; Weidlich, S.; Knebel, A.; Kulathu, Y. A single MIU motif of MINDY-1 recognizes K48-linked polyubiquitin chains. EMBO Rep. 2017, 18, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Hermanns, T.; Pichlo, C.; Woiwode, I.; Klopffleisch, K.; Witting, K.F.; Ovaa, H.; Baumann, U.; Hofmann, K. A family of unconventional deubiquitinases with modular chain specificity determinants. Nat. Commun. 2018, 9, 799. [Google Scholar] [CrossRef]
- Blandino, G.; Di Agostino, S. New therapeutic strategies to treat human cancers expressing mutant p53 proteins. J. Exp. Clin. Cancer Res. 2018, 37, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Prives, C. Relevance of the p53-MDM2 axis to aging. Cell Death Differ. 2018, 25, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Luo, K.; Zhang, L.; Cheville, J.C.; Lou, Z. USP10 regulates p53 localization and stability by deubiquitinating p53. Cell 2010, 140, 384–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Ghosh, M.K. Cell death and deubiquitinases: Perspectives in cancer. Biomed. Res. Int. 2014, 2014, 435197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Q.; Jin, J.; Hu, H.; Li, H.S.; Romano, S.; Xiao, Y.; Nakaya, M.; Zhou, X.; Cheng, X.; Yang, P.; et al. USP15 stabilizes MDM2 to mediate cancer-cell survival and inhibit antitumor T cell responses. Nat. Immunol. 2014, 15, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; Challagundla, K.B.; Dai, M.S. Positive regulation of p53 stability and activity by the deubiquitinating enzyme Otubain 1. EMBO J. 2012, 31, 576–592. [Google Scholar] [CrossRef]
- Carneiro, A.P.; Reis, C.F.; Morari, E.C.; Maia, Y.C.; Nascimento, R.; Bonatto, J.M.; de Souza, M.A.; Goulart, L.R.; Ward, L.S. A putative OTU domain-containing protein 1 deubiquitinating enzyme is differentially expressed in thyroid cancer and identifies less-aggressive tumours. Br. J. Cancer 2014, 111, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Lu, Z.; Lu, X.; Chen, L.; Cao, J.; Zhang, S.; Ling, Y.; Zhou, X. OTUD5 regulates p53 stability by deubiquitinating p53. PLoS One 2013, 8, e77682. [Google Scholar] [CrossRef]
- Zhang, X.; Berger, F.G.; Yang, J.; Lu, X. USP4 inhibits p53 through deubiquitinating and stabilizing ARF-BP1. EMBO J. 2011, 30, 2177–2189. [Google Scholar] [CrossRef]
- Qin, J.; Zhou, Z.; Chen, W.; Wang, C.; Zhang, H.; Ge, G.; Shao, M.; You, D.; Fan, Z.; Xia, H.; et al. BAP1 promotes breast cancer cell proliferation and metastasis by deubiquitinating KLF5. Nat. Commun. 2015, 6, 8471. [Google Scholar] [CrossRef] [Green Version]
- Pereg, Y.; Liu, B.Y.; O’Rourke, K.M.; Sagolla, M.; Dey, A.; Komuves, L.; French, D.M.; Dixit, V.M. Ubiquitin hydrolase Dub3 promotes oncogenic transformation by stabilizing Cdc25A. Nat. Cell Biol. 2010, 12, 400–406. [Google Scholar] [CrossRef] [PubMed]
- McClurg, U.L.; Robson, C.N. Deubiquitinating enzymes as oncotargets. Oncotarget 2015, 6, 9657–9668. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, P.; Wei, Y.; Piao, H.L.; Wang, W.; Maddika, S.; Wang, M.; Chen, D.; Sun, Y.; Hung, M.C.; et al. Deubiquitylation and stabilization of PTEN by USP13. Nat. Cell Biol. 2013, 15, 1486–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, K.D. Regulation of ubiquitin-dependent processes by deubiquitinating enzymes. FASEB J. 1997, 11, 1245–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Lv, Y.; Li, H.; Gao, H.; Song, S.; Zhang, Y.; Xing, G.; Kong, X.; Wang, L.; Li, Y.; et al. Deubiquitylase OTUD3 regulates PTEN stability and suppresses tumorigenesis. Nat. Cell Biol. 2015, 17, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argiris, A.; Karamouzis, M.V.; Raben, D.; Ferris, R.L. Head and neck cancer. Lancet 2008, 371, 1695–1709. [Google Scholar] [CrossRef]
- Pezzuto, F.; Buonaguro, L.; Caponigro, F.; Ionna, F.; Starita, N.; Annunziata, C.; Buonaguro, F.M.; Tornesello, M.L. Update on Head and Neck Cancer: Current Knowledge on Epidemiology, Risk Factors, Molecular Features and Novel Therapies. Oncology 2015, 89, 125–136. [Google Scholar] [CrossRef]
- Morgan, E.L.; Chen, Z.; Van Waes, C. Regulation of NFkappaB Signalling by Ubiquitination: A Potential Therapeutic Target in Head and Neck Squamous Cell Carcinoma? Cancers 2020, 12, 2877. [Google Scholar] [CrossRef]
- Shinriki, S.; Jono, H.; Maeshiro, M.; Nakamura, T.; Guo, J.; Li, J.D.; Ueda, M.; Yoshida, R.; Shinohara, M.; Nakayama, H.; et al. Loss of CYLD promotes cell invasion via ALK5 stabilization in oral squamous cell carcinoma. J. Pathol. 2018, 244, 367–379. [Google Scholar] [CrossRef]
- Liu, S.; de Boeck, M.; van Dam, H.; Ten Dijke, P. Regulation of the TGF-beta pathway by deubiquitinases in cancer. Int. J. Biochem. Cell Biol. 2016, 76, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Chu, M.; Lin, M.; He, Y.; Wang, Z. USP7 stabilizes EZH2 and enhances cancer malignant progression. Am. J. Cancer Res. 2020, 10, 299–313. [Google Scholar] [PubMed]
- Sacco, J.J.; Coulson, J.M.; Clague, M.J.; Urbe, S. Emerging roles of deubiquitinases in cancer-associated pathways. IUBMB Life 2010, 62, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Gatti, V.; Bernassola, F.; Talora, C.; Melino, G.; Peschiaroli, A. The Impact of the Ubiquitin System in the Pathogenesis of Squamous Cell Carcinomas. Cancers 2020, 12, 1595. [Google Scholar] [CrossRef]
- Prieto-Garcia, C.; Hartmann, O.; Reissland, M.; Braun, F.; Fischer, T.; Walz, S.; Schulein-Volk, C.; Eilers, U.; Ade, C.P.; Calzado, M.A.; et al. Maintaining protein stability of ∆Np63 via USP28 is required by squamous cancer cells. EMBO Mol. Med. 2020, 12, e11101. [Google Scholar] [CrossRef]
- Prieto-Garcia, C.; Tomaskovic, I.; Shah, V.J.; Dikic, I.; Diefenbacher, M. USP28: Oncogene or Tumor Suppressor? A Unifying Paradigm for Squamous Cell Carcinoma. Cells 2021, 10, 2652. [Google Scholar] [CrossRef]
- Jing, C.; Duan, Y.; Zhou, M.; Yue, K.; Zhuo, S.; Li, X.; Liu, D.; Ye, B.; Lai, Q.; Li, L.; et al. Blockade of deubiquitinating enzyme PSMD14 overcomes chemoresistance in head and neck squamous cell carcinoma by antagonizing E2F1/Akt/SOX2-mediated stemness. Theranostics 2021, 11, 2655–2669. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, S.; Wang, J.; Cheng, Z. Identification of PSMD7 as a prognostic factor correlated with immune infiltration in head and neck squamous cell carcinoma. Biosci. Rep. 2021, 41, BSR20203829. [Google Scholar] [CrossRef]
- Goncharov, T.; Niessen, K.; de Almagro, M.C.; Izrael-Tomasevic, A.; Fedorova, A.V.; Varfolomeev, E.; Arnott, D.; Deshayes, K.; Kirkpatrick, D.S.; Vucic, D. OTUB1 modulates c-IAP1 stability to regulate signalling pathways. EMBO J. 2013, 32, 1103–1114. [Google Scholar] [CrossRef]
- Ge, W.L.; Xu, J.F.; Hu, J. Regulation of Oral Squamous Cell Carcinoma Proliferation Through Crosstalk Between SMAD7 and CYLD. Cell Physiol. Biochem. 2016, 38, 1209–1217. [Google Scholar] [CrossRef]
- Zhang, M.J.; Chen, D.S.; Li, H.; Liu, W.W.; Han, G.Y.; Han, Y.F. Clinical significance of USP7 and EZH2 in predicting prognosis of laryngeal squamous cell carcinoma and their possible functional mechanism. Int. J. Clin. Exp. Pathol. 2019, 12, 2184–2194. [Google Scholar] [PubMed]
- Xu, L.; Li, Y.Y.; Zhang, Y.C.; Wu, Y.X.; Guo, D.D.; Long, D.; Liu, Z.H. A Novel Ferroptosis-Related Gene Signature to Predict Prognosis in Patients with Head and Neck Squamous Cell Carcinoma. Dis. Markers 2021, 2021, 5759927. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Wang, L.; Zhang, L.; Pan, X.; Zhao, W. Ubiquitin-specific protease 4 promotes TNF-alpha-induced apoptosis by deubiquitination of RIP1 in head and neck squamous cell carcinoma. FEBS Lett. 2013, 587, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jingjing, W.; Wenzheng, G.; Donghua, W.; Guangyu, H.; Aiping, Z.; Wenjuan, W. Deubiquitination and stabilization of programmed cell death ligand 1 by ubiquitin-specific peptidase 9, X-linked in oral squamous cell carcinoma. Cancer Med. 2018, 7, 4004–4011. [Google Scholar] [CrossRef] [Green Version]
- Nanayakkara, D.M.; Nguyen, M.N.; Wood, S.A. Deubiquitylating enzyme, USP9X, regulates proliferation of cells of head and neck cancer lines. Cell Prolif. 2016, 49, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Dou, Y.; Lin, J.; Shu, H.; Jiang, N. Role of ubiquitin-specific peptidase 22 in carcinogenesis of human pharyngeal squamous cell carcinoma. Mol. Med. Rep. 2014, 10, 2973–2978. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Kumar, M.; Yang, L.; Molkentine, D.P.; Valdecanas, D.; Yu, S.; Meyn, R.E.; Heymach, J.V.; Skinner, H.D. BAP1 Is a Novel Target in HPV-Negative Head and Neck Cancer. Clin. Cancer Res. 2018, 24, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Glinsky, G.V.; Berezovska, O.; Glinskii, A.B. Microarray analysis identifies a death-from-cancer signature predicting therapy failure in patients with multiple types of cancer. J. Clin. Investig. 2005, 115, 1503–1521. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Varthi, M.; Sykes, S.M.; Phillips, C.; Warzecha, C.; Zhu, W.; Wyce, A.; Thorne, A.W.; Berger, S.L.; McMahon, S.B. The putative cancer stem cell marker USP22 is a subunit of the human SAGA complex required for activated transcription and cell-cycle progression. Mol. Cell 2008, 29, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Ling, S.; Li, J.; Shan, Q.; Dai, H.; Lu, D.; Wen, X.; Song, P.; Xie, H.; Zhou, L.; Liu, J.; et al. USP22 mediates the multidrug resistance of hepatocellular carcinoma via the SIRT1/AKT/MRP1 signaling pathway. Mol. Oncol. 2017, 11, 682–695. [Google Scholar] [CrossRef]
- Zhuang, Y.J.; Liao, Z.W.; Yu, H.W.; Song, X.L.; Liu, Y.; Shi, X.Y.; Lin, X.D.; Zhou, T.C. ShRNA-mediated silencing of the ubiquitin-specific protease 22 gene restrained cell progression and affected the Akt pathway in nasopharyngeal carcinoma. Cancer Biol. Ther. 2015, 16, 88–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liu, N.; Zhao, Y.; Zhu, X.; Wang, C.; Liu, Q.; Gao, C.; Zhao, X.; Li, J. Oncogenic USP22 supports gastric cancer growth and metastasis by activating c-Myc/NAMPT/SIRT1-dependent FOXO1 and YAP signaling. Aging (Albany NY) 2019, 11, 9643–9660. [Google Scholar] [CrossRef] [PubMed]
- Atanassov, B.S.; Dent, S.Y. USP22 regulates cell proliferation by deubiquitinating the transcriptional regulator FBP1. EMBO Rep. 2011, 12, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, D.; Liu, L.; Wang, L.; Yan, M. miR-140 inhibits osteosarcoma progression by impairing USP22-mediated LSD1 stabilization and promoting p21 expression. Mol. Ther. Nucleic Acids 2021, 24, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, J.; Chen, Q.; Jin, S.; Mi, S.; Shao, W.; Kudo, Y.; Zeng, S.; Qi, G. Expression of USP22 and the chromosomal passenger complex is an indicator of malignant progression in oral squamous cell carcinoma. Oncol. Lett. 2019, 17, 2040–2046. [Google Scholar] [CrossRef] [PubMed]
- Tchakarska, G.; Sola, B. The double dealing of cyclin D1. Cell Cycle 2020, 19, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.L.; Panasyuk, G.; Gwalter, J.; Nemazanyy, I.; Fenton, T.; Filonenko, V.; Gout, I. Regulation of ribosomal protein S6 kinases by ubiquitination. Biochem. Biophys Res. Commun. 2008, 369, 382–387. [Google Scholar] [CrossRef]
- Schwickart, M.; Huang, X.; Lill, J.R.; Liu, J.; Ferrando, R.; French, D.M.; Maecker, H.; O’Rourke, K.; Bazan, F.; Eastham-Anderson, J.; et al. Deubiquitinase USP9X stabilizes MCL1 and promotes tumour cell survival. Nature 2010, 463, 103–107. [Google Scholar] [CrossRef]
- Panasyuk, G.; Nemazanyy, I.; Filonenko, V.; Gout, I. Ribosomal protein S6 kinase 1 interacts with and is ubiquitinated by ubiquitin ligase ROC1. Biochem. Biophys Res. Commun. 2008, 369, 339–343. [Google Scholar] [CrossRef]
- Khan, O.M.; Carvalho, J.; Spencer-Dene, B.; Mitter, R.; Frith, D.; Snijders, A.P.; Wood, S.A.; Behrens, A. The deubiquitinase USP9X regulates FBW7 stability and suppresses colorectal cancer. J. Clin. Investig. 2018, 128, 1326–1337. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.; Chen, Y.; Ye, D.; Lei, H.; Xu, H.; Yang, L.; Wu, Y.; Gu, W. Ubiquitin-specific protease 14 regulates cell proliferation and apoptosis in oral squamous cell carcinoma. Int. J. Biochem. Cell Biol. 2016, 79, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, B.; Suresh Kumar, K.G. The multifaceted roles of USP7: New therapeutic opportunities. Cell Biochem. Biophys. 2011, 60, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M.; Uchimaru, K. Targeting EZH2 in cancer therapy. Curr. Opin. Oncol. 2017, 29, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.T.; Cai, M.Y.; Wang, X.G.; Kong, L.L.; Mai, S.J.; Liu, Y.H.; Zhang, H.B.; Liao, Y.J.; Zheng, F.; Zhu, W.; et al. EZH2 supports nasopharyngeal carcinoma cell aggressiveness by forming a co-repressor complex with HDAC1/HDAC2 and Snail to inhibit E-cadherin. Oncogene 2012, 31, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, W.J.; Liu, Z.; Loda, M.; Freeman, M.R. The ubiquitin-specific protease USP2a enhances tumor progression by targeting cyclin A1 in bladder cancer. Cell Cycle 2012, 11, 1123–1130. [Google Scholar] [CrossRef] [Green Version]
- da Silva, S.D.; Cunha, I.W.; Nishimoto, I.N.; Soares, F.A.; Carraro, D.M.; Kowalski, L.P.; Graner, E. Clinicopathological significance of ubiquitin-specific protease 2a (USP2a), fatty acid synthase (FASN), and ErbB2 expression in oral squamous cell carcinomas. Oral. Oncol. 2009, 45, e134–e139. [Google Scholar] [CrossRef]
- Yu, H.; Pak, H.; Hammond-Martel, I.; Ghram, M.; Rodrigue, A.; Daou, S.; Barbour, H.; Corbeil, L.; Hebert, J.; Drobetsky, E.; et al. Tumor suppressor and deubiquitinase BAP1 promotes DNA double-strand break repair. Proc. Natl. Acad. Sci. USA 2014, 111, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Molkentine, D.P.; Molkentine, J.M.; Bridges, K.A.; Valdecanas, D.R.; Dhawan, A.; Bahri, R.; Hefner, A.J.; Kumar, M.; Yang, L.; Abdelhakiem, M.; et al. p16 Represses DNA Damage Repair via a Novel Ubiquitin-Dependent Signaling Cascade. Cancer Res. 2022, 82, 916–928. [Google Scholar] [CrossRef]
- Varilla, V.; Atienza, J.; Dasanu, C.A. Immune alterations and immunotherapy prospects in head and neck cancer. Expert Opin. Biol. Ther. 2013, 13, 1241–1256. [Google Scholar] [CrossRef]
- Klatka, J.; Hymos, A.; Szkatula-Lupina, A.; Grywalska, E.; Klatka, B.; Terpilowski, M.; Stepulak, A. T-Lymphocyte Activation Is Correlated With the Presence of Anti-EBV in Patients With Laryngeal Squamous Cell Carcinoma. In Vivo 2019, 33, 2007–2012. [Google Scholar] [CrossRef]
- Whitehurst, C.B.; Ning, S.; Bentz, G.L.; Dufour, F.; Gershburg, E.; Shackelford, J.; Langelier, Y.; Pagano, J.S. The Epstein-Barr virus (EBV) deubiquitinating enzyme BPLF1 reduces EBV ribonucleotide reductase activity. J. Virol. 2009, 83, 4345–4353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyson, O.F.; Pagano, J.S.; Whitehurst, C.B. The Translesion Polymerase Pol eta Is Required for Efficient Epstein-Barr Virus Infectivity and Is Regulated by the Viral Deubiquitinating Enzyme BPLF1. J. Virol. 2017, 91, e00600-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, S.; Murata, T.; Kanda, T.; Isomura, H.; Narita, Y.; Sugimoto, A.; Kawashima, D.; Tsurumi, T. Epstein-Barr virus deubiquitinase downregulates TRAF6-mediated NF-kappaB signaling during productive replication. J. Virol. 2013, 87, 4060–4070. [Google Scholar] [CrossRef] [Green Version]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-kappaB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef]
- Poondla, N.; Chandrasekaran, A.P.; Kim, K.S.; Ramakrishna, S. Deubiquitinating enzymes as cancer biomarkers: New therapeutic opportunities? BMB Rep. 2019, 52, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Singh, A.B. Deubiquitinases and cancer: A snapshot. Crit. Rev. Oncol. Hematol. 2016, 103, 22–26. [Google Scholar] [CrossRef]
- Mofers, A.; Pellegrini, P.; Linder, S.; D’Arcy, P. Proteasome-associated deubiquitinases and cancer. Cancer Metastasis Rev. 2017, 36, 635–653. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.R.; Choi, W.C.; Lee, S.; Hwang, J.; Hwang, E.; Guchhait, K.; Haas, J.; Toth, Z.; Jeon, Y.H.; Oh, T.K.; et al. Bilateral inhibition of HAUSP deubiquitinase by a viral interferon regulatory factor protein. Nat. Struct. Mol. Biol. 2011, 18, 1336–1344. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; D’Arcy, P.; Wang, X.; Ray, A.; Tai, Y.T.; Hu, Y.; Carrasco, R.D.; Richardson, P.; Linder, S.; Chauhan, D.; et al. A novel small molecule inhibitor of deubiquitylating enzyme USP14 and UCHL5 induces apoptosis in multiple myeloma and overcomes bortezomib resistance. Blood 2014, 123, 706–716. [Google Scholar] [CrossRef]
- Woo, R.A.; Jack, M.T.; Xu, Y.; Burma, S.; Chen, D.J.; Lee, P.W. DNA damage-induced apoptosis requires the DNA-dependent protein kinase, and is mediated by the latent population of p53. EMBO J. 2002, 21, 3000–3008. [Google Scholar] [CrossRef]
- An, J.; Mo, D.; Liu, H.; Veena, M.S.; Srivatsan, E.S.; Massoumi, R.; Rettig, M.B. Inactivation of the CYLD deubiquitinase by HPV E6 mediates hypoxia-induced NF-kappaB activation. Cancer Cell. 2008, 14, 394–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiran, S.; Dar, A.; Singh, S.K.; Lee, K.Y.; Dutta, A. The Deubiquitinase USP46 Is Essential for Proliferation and Tumor Growth of HPV-Transformed Cancers. Mol. Cell 2018, 72, 823–835.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.; Kim, J. The potential role of YAP in head and neck squamous cell carcinoma. Exp. Mol. Med. 2020, 52, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Luo, J.; Xiao, Z.; Zang, Y.; Li, X.; Zhou, Y.; Zhou, J.; Tian, Z.; Zhu, J.; Zhao, X. USP36 facilitates esophageal squamous carcinoma progression via stabilizing YAP. Cell Death Dis. 2022, 13, 1021. [Google Scholar] [CrossRef]
- Frappier, L.; Verrijzer, C.P. Gene expression control by protein deubiquitinases. Curr. Opin. Genet. Dev. 2011, 21, 207–213. [Google Scholar] [CrossRef]
- Bonacci, T.; Suzuki, A.; Grant, G.D.; Stanley, N.; Cook, J.G.; Brown, N.G.; Emanuele, M.J. Cezanne/OTUD7B is a cell cycle-regulated deubiquitinase that antagonizes the degradation of APC/C substrates. EMBO J. 2018, 37, e98701. [Google Scholar] [CrossRef]
- Yeh, C.; Coyaud, E.; Bashkurov, M.; van der Lelij, P.; Cheung, S.W.; Peters, J.M.; Raught, B.; Pelletier, L. The Deubiquitinase USP37 Regulates Chromosome Cohesion and Mitotic Progression. Curr. Biol. 2015, 25, 2290–2299. [Google Scholar] [CrossRef] [Green Version]
- Lecona, E.; Rodriguez-Acebes, S.; Specks, J.; Lopez-Contreras, A.J.; Ruppen, I.; Murga, M.; Munoz, J.; Mendez, J.; Fernandez-Capetillo, O. USP7 is a SUMO deubiquitinase essential for DNA replication. Nat. Struct. Mol. Biol. 2016, 23, 270–277. [Google Scholar] [CrossRef]
- Weber, A.; Heinlein, M.; Dengjel, J.; Alber, C.; Singh, P.K.; Hacker, G. The deubiquitinase Usp27x stabilizes the BH3-only protein Bim and enhances apoptosis. EMBO Rep. 2016, 17, 724–738. [Google Scholar] [CrossRef]
Figure 1.
Schematic of key events in ubiquitination and deubiquitination. Under the conditions where ATP provides energy, ubiquitin binds to the target protein through the cascade catalytic reaction of ubiquitin-activating enzyme (E1), ubiquitin-conjugating enzyme (E2), and ubiquitin ligase (E3). The ubiquitinated target protein is recognized and degraded by 26S proteasome. At present, 2 E1s, about 50 E2s, and more than 600 E3s are known to be encoded by the human genome. Deubiquitinating enzymes (DUBs) maintain ubiquitin system homeostasis by cleaving polyubiquitin chains or completely removing ubiquitin chains from ubiquitinated proteins. Via deubiquitination, free ubiquitins are generated and recycled.
Figure 1.
Schematic of key events in ubiquitination and deubiquitination. Under the conditions where ATP provides energy, ubiquitin binds to the target protein through the cascade catalytic reaction of ubiquitin-activating enzyme (E1), ubiquitin-conjugating enzyme (E2), and ubiquitin ligase (E3). The ubiquitinated target protein is recognized and degraded by 26S proteasome. At present, 2 E1s, about 50 E2s, and more than 600 E3s are known to be encoded by the human genome. Deubiquitinating enzymes (DUBs) maintain ubiquitin system homeostasis by cleaving polyubiquitin chains or completely removing ubiquitin chains from ubiquitinated proteins. Via deubiquitination, free ubiquitins are generated and recycled.

Figure 2.
Involvement of DUBs in HNSCC.


{kind=link}
{kind=link}
Table 1.
DUB families and their members.
| DUB Family | Type | Member | |
|---|---|---|---|
| USPs | ubiquitin-specific proteases | cysteine protease | USP1-USP8, USP9X, USP9Y, USP10-USP16, USP18-USP22, USP24-USP26, USP27X, USP28-USP54, CYLD, DUB3 |
| UCHs | ubiquitin carboxy-terminal hydrolases | cysteine protease | BAP1, UCHL1, UCHL3, UCHL5 |
| JAMMs | JAB1/MPN/MOV34 proteases | zinc-dependent metalloprotease | BRCC3, COPS5, PSMD, MPND, MYSM1, STAMBP, STAMBPL1, PRPF8, COPS6, EIF3F, PSMD7, EIF3H |
| OTUs | otubain/ovarian tumor-domain containing proteins | cysteine protease | OTUB1, OTUB2, OTUD1, OTUD3, OTUD4, OTUD5, OTUD6A, OTUD6B, OTUD7A, OTUD7B, OTULIN, OTULINL, TNFAIP3, ZRANB1, YOD1, VCPIP1 |
| MJDs (Josephins) | Machado–Joseph disease domain superfamily | cysteine protease | ATXN3, ATXN3L, JOSD1, JOSD2, JOSD3 |
| MCPIPs | monocyte chemotactic protein- induced protein | cysteine protease | MCPIP1, MCPIP2, MCPIP3, MCPIP4, MCPIP5, MCPIP6, MCPIP7 |
| MINDYs | motif interacting with ubiquitin- containing novel DUB family | cysteine protease | FAM63A/MINDY1, FAM63B/MINDY2, FAM188A/MINDY3, FAM188B/MINDY4 |
| ZUFSP/SUP1 | Zn-finger and UFSP domain protein | cysteine protease | ZUP1 |
Table 2.
DUB function in HNSCC.
| DUB | Abnormal Regulation | The Roles in HNSCC | Substrates | References |
|---|---|---|---|---|
| CYLD | mutation | removes K63-polyubiquitin and M1 linear-ubiquitin chains and inhibits NFκB signaling | RIP1, TRAF2, TRAF6, TAK1, NEMO | [49] |
| Low expression | promotes TGF-β signaling and cell invasion | ALK5 | [50] | |
| Low expression | removes K63-polyubiquitin, thereby inhibiting TGF-β signaling | SMAD7 | [51,60] | |
| USP4 | overexpression | removes K63-polyubiquitin and promotes TNF-α induced apoptosis | RIP1 | [63] |
| USP7 | overexpression | promotes cell growth, cell migration, and invasion | EZH2 | [52,61] |
| USP9X | overexpression | deubiquitinates and stabilizes PD-L1 to promote cell proliferation | PD-L1 | [64] |
| Low expression | mTOR pathway | [65] | ||
| USP22 | overexpression | associates with lymph node metastasis and histological grade | [66] | |
| USP28 | overexpression | inhibits p53 on the promoter of pro-apoptotic genes | Np63 | [53,54,55,56] |
| BAP1 | overexpression | deubiquitinates H2A at the DSB site suppresses transcription, and promotes DNA repair | H2Aub(K119) | [67] |
| PSMD14 | overexpression | stabilizes E2F1, gives stemness to cells by SOX2 expression and Akt signal activation | E2F1 | [57] |
| PSMD7 | overexpression | a prognostic factor correlated with immune infiltration | [58] | |
| OTUB1 | overexpression | a risk factor | [49,59,62] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jin, S.; Kudo, Y.; Horiguchi, T. The Role of Deubiquitinating Enzyme in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2023, 24, 552. https://doi.org/10.3390/ijms24010552
AMA Style
Jin S, Kudo Y, Horiguchi T. The Role of Deubiquitinating Enzyme in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2023; 24(1):552. https://doi.org/10.3390/ijms24010552
Chicago/Turabian StyleJin, Shengjian, Yasusei Kudo, and Taigo Horiguchi. 2023. "The Role of Deubiquitinating Enzyme in Head and Neck Squamous Cell Carcinoma" International Journal of Molecular Sciences 24, no. 1: 552. https://doi.org/10.3390/ijms24010552
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.