
Establishment of Mucoepidermoid Carcinoma Cell Lines from Surgical and Recurrence Biopsy Specimens
 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
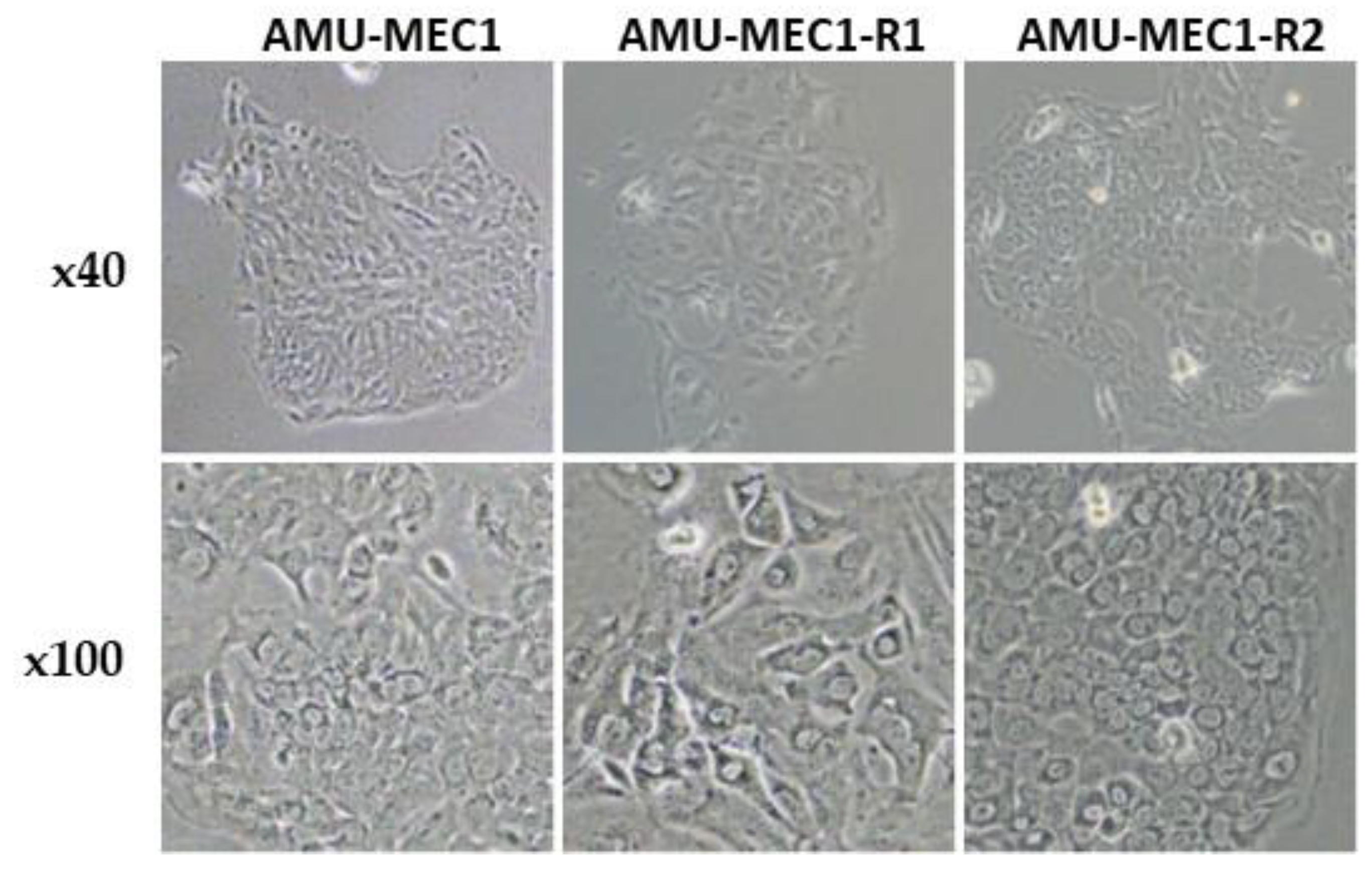
2.1. Morphological Features of the Established MEC Cell Lines
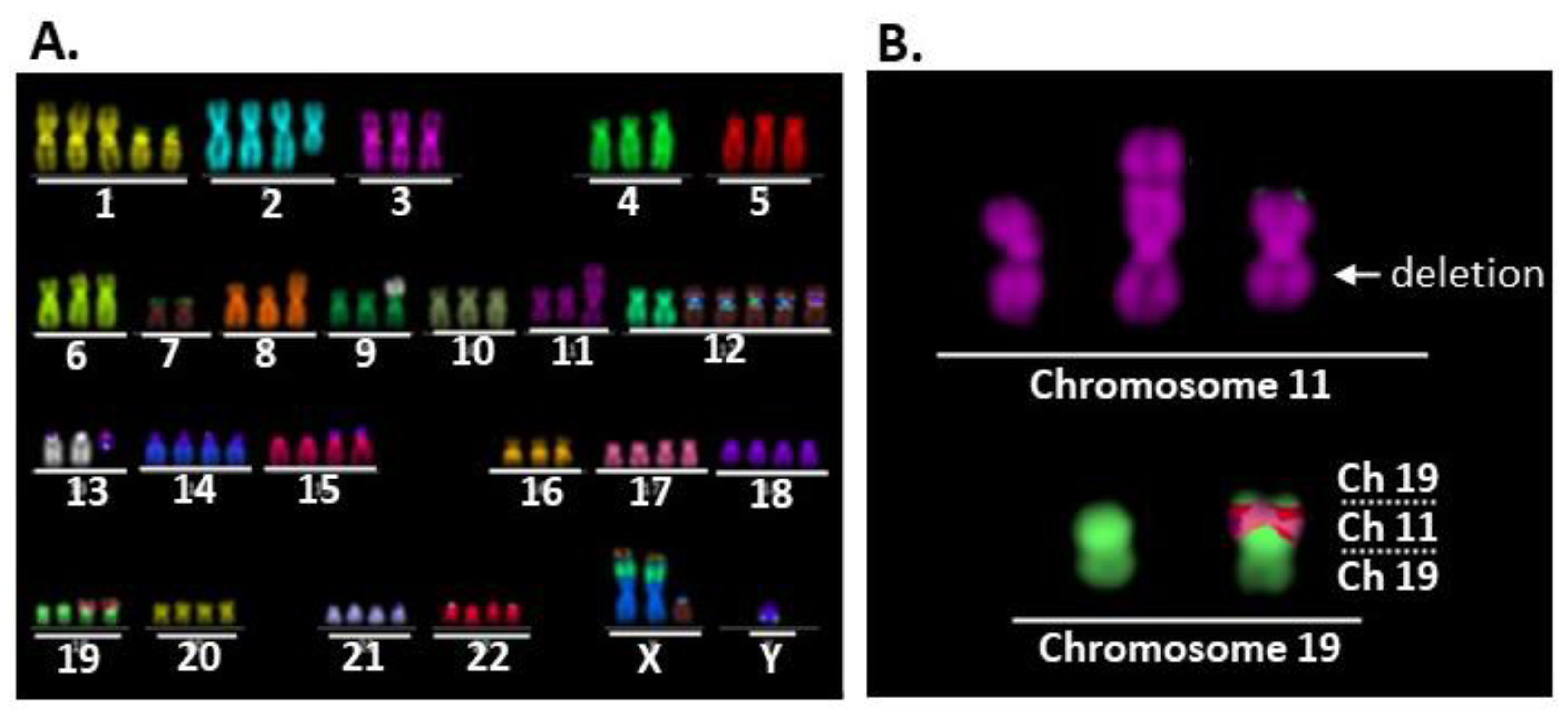
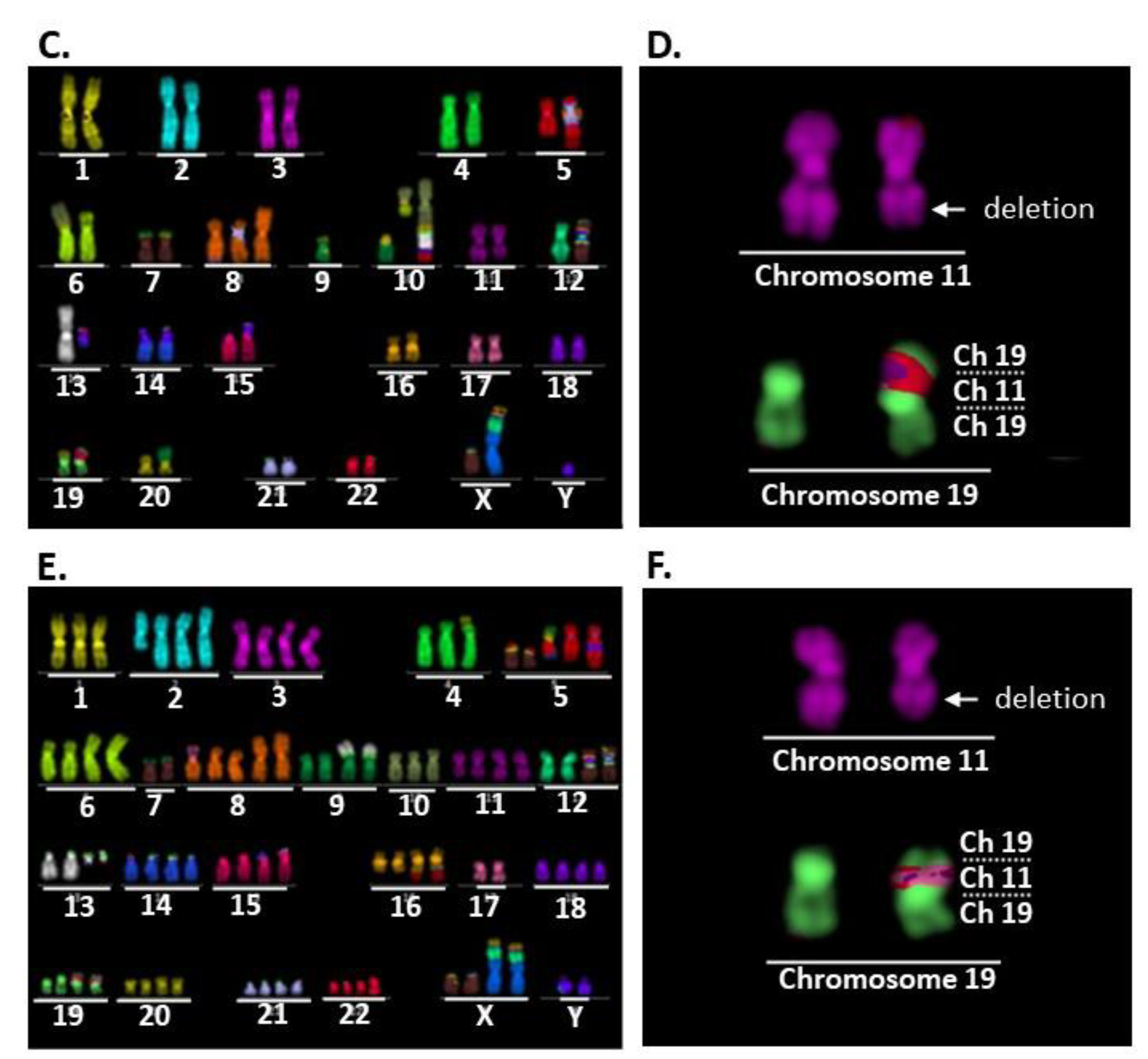
2.2. Comparison of Karyotypes among the Three Cell Lines
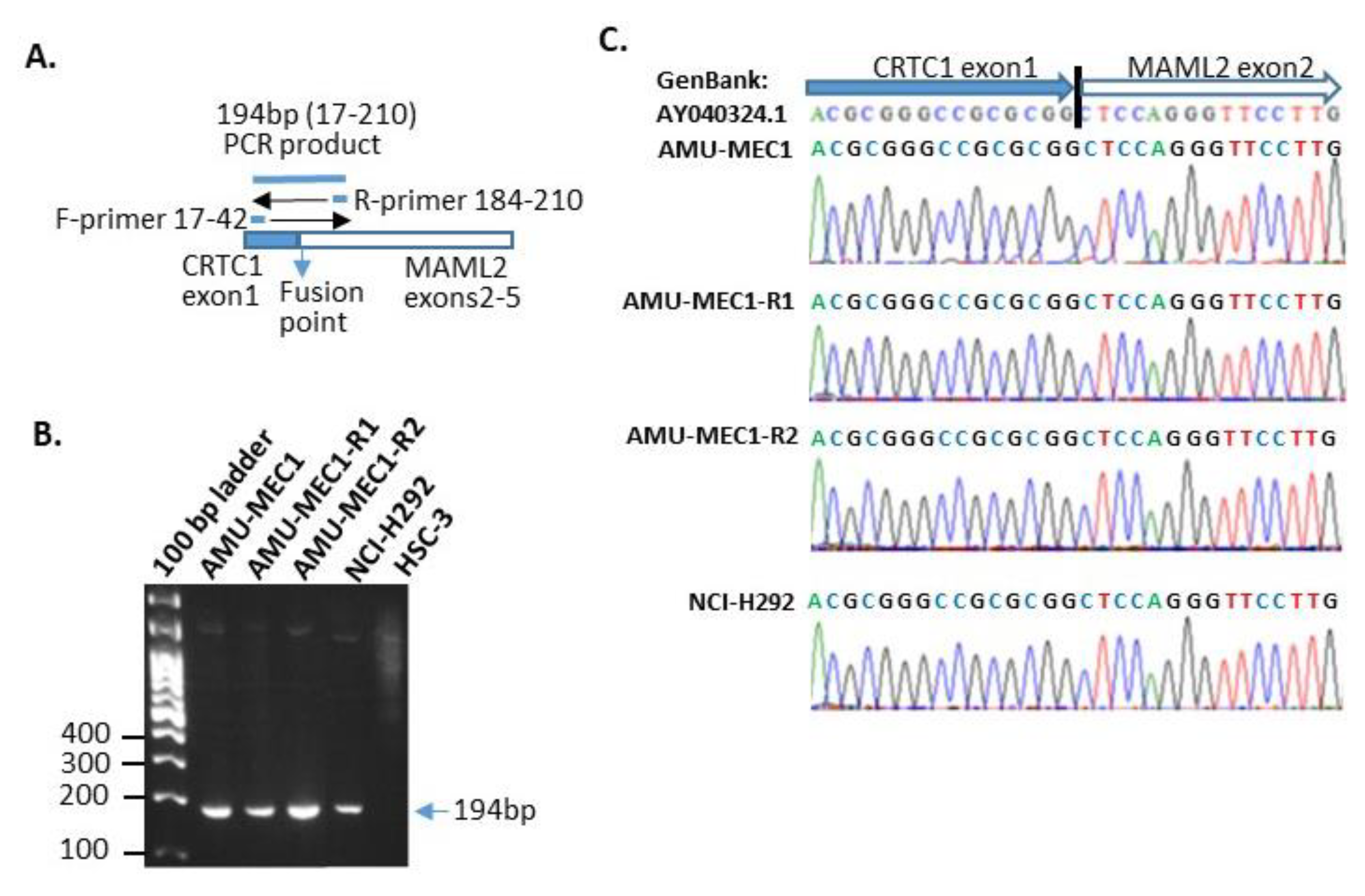
2.3. der(19)ins(19;11)(p13;?) Resulted in CRTC1-MAML2 Chimeric Gene Generation
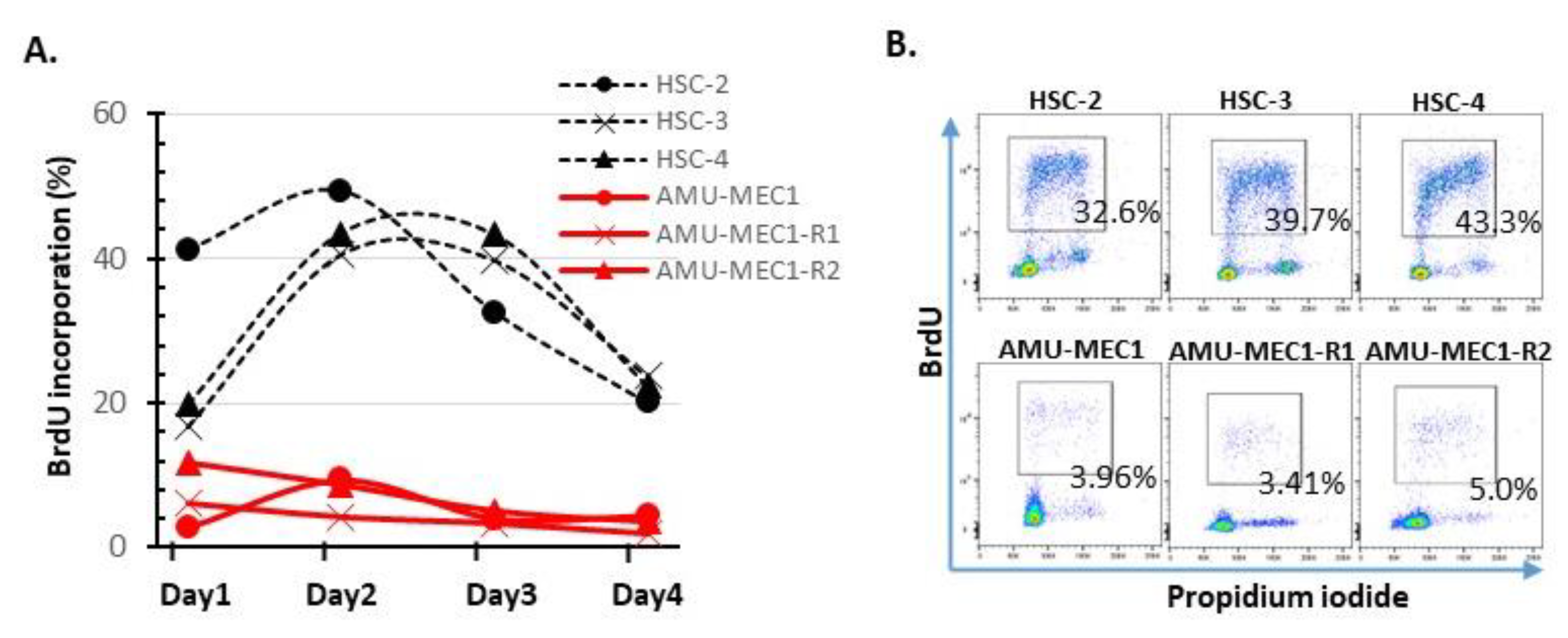
2.4. Growth Activity Analysis
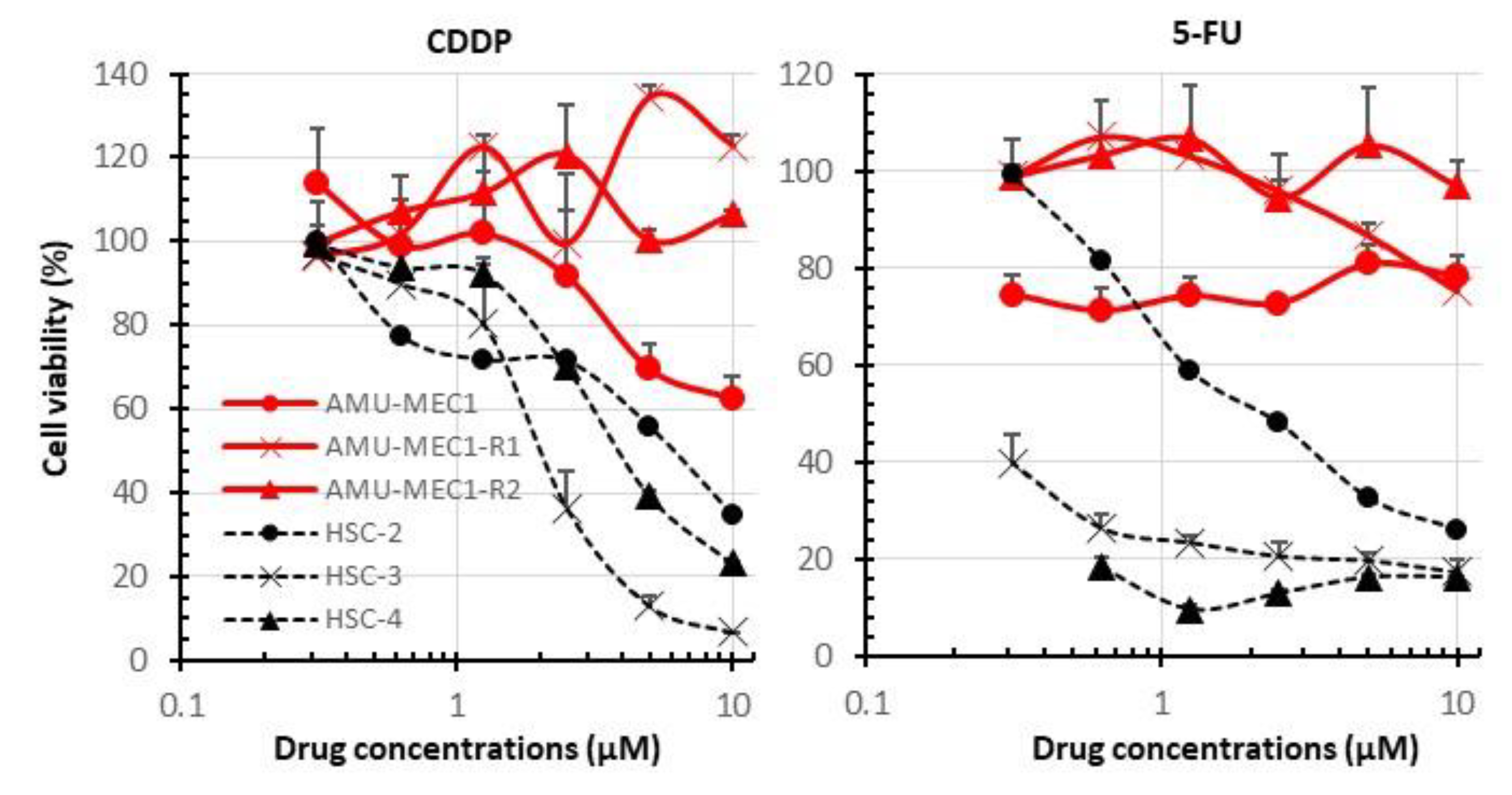
2.5. Comparison of Susceptibility of MEC and OSCC Cell Lines to CDDP and 5-FU
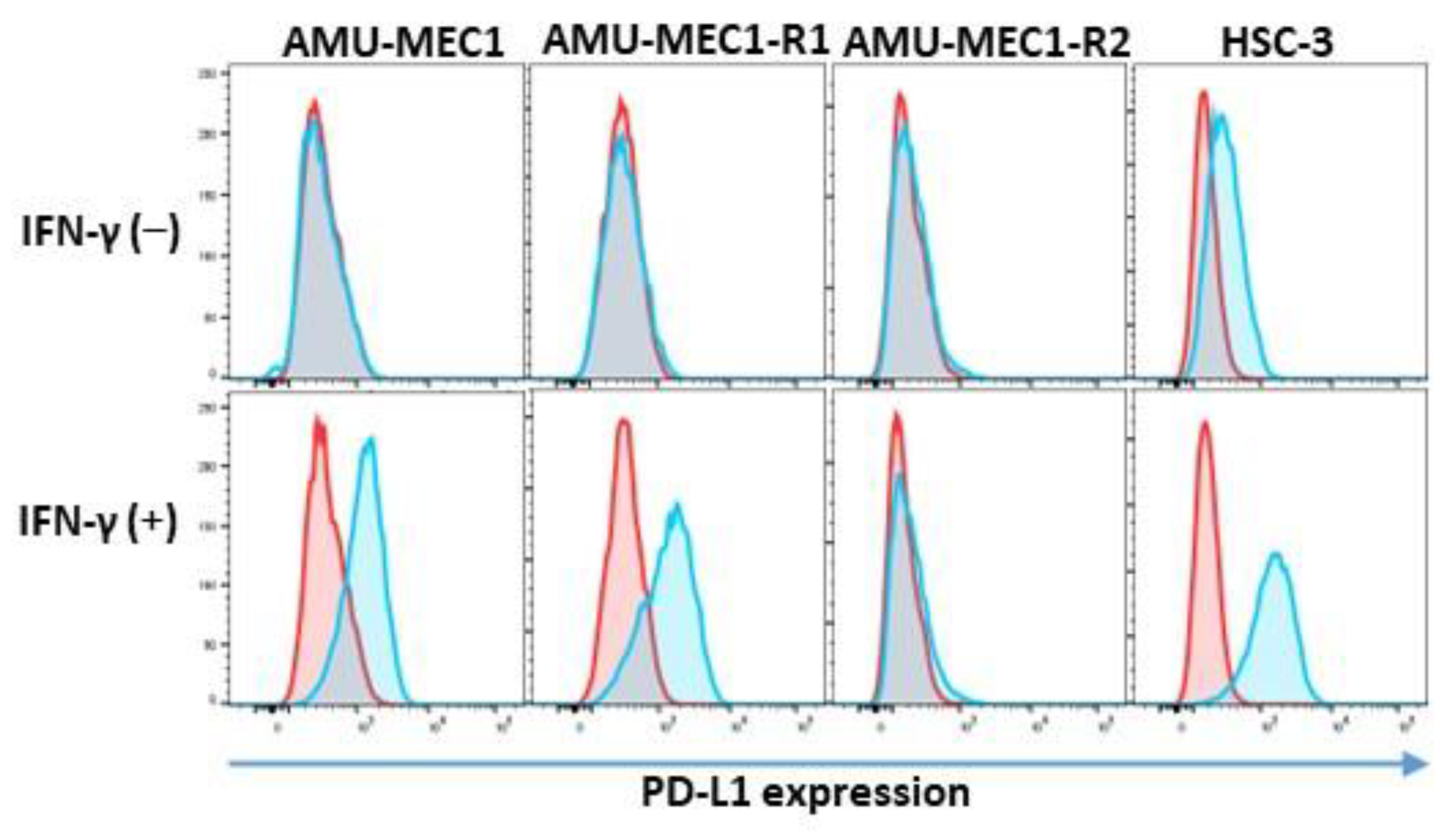
2.6. PD-L1 Expression in the MEC Cell Lines
3. Discussion
4. Materials and Methods
4.1. Patient
4.2. Tumor Specimens and Establishment of MEC Cell Lines
4.3. Measurement of Drug Susceptibility Using Water-Soluble Tetrazolium-1 (WST-1) Assay
4.4. Analysis of Bromodeoxyuridine (BrdU) Incorporation and Detection of PD-L1 by Flow Cytometry
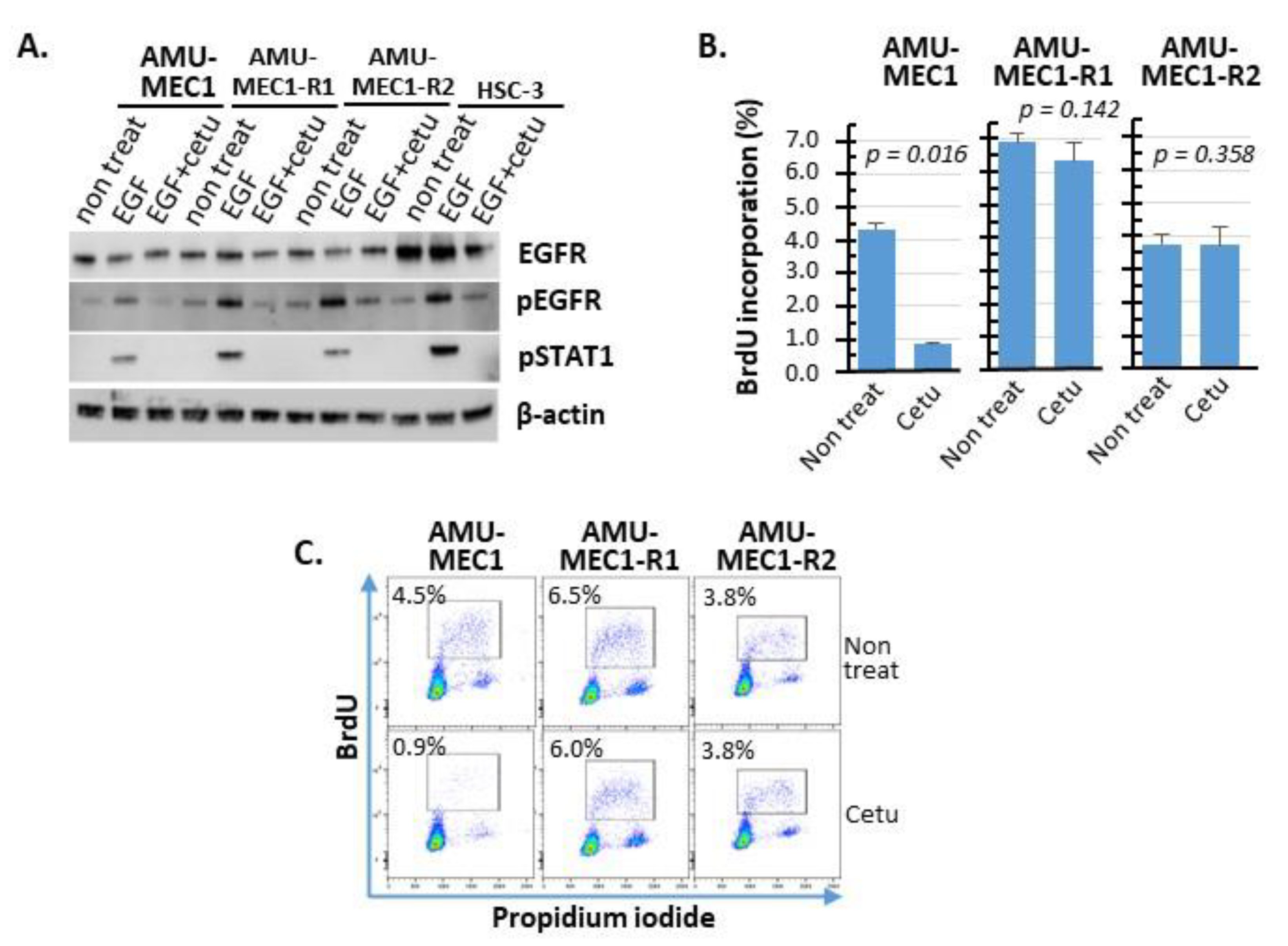
4.5. Western Blot Analysis of EGFR Expression and Phosphorylation
4.6. Karyotype Analysis Using Multi-Color FISH
4.7. Detection and Nucleotide Sequencing of CRTC1-MAML2 Chimeric Gene
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Naggar, A.K.; Chan, J.K.C.; Grandis, J.R.; Takata, T.; Slootweg, P.J.; Brandwein-Gensler, M.; Bell, D.; Inagaki, H.; Katabi, N.; Leivo, I.; et al. Mucoepidermoid carcinoma. In World Health Organization Classification of Head and Neck Tumours; IARC Press: Lyon, France, 2017; pp. 163–164. [Google Scholar]
- Sama, S.; Komiya, T.; Guddati, A.K. Advances in the Treatment of Mucoepidermoid Carcinoma. World J. Oncol. 2022, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Eveson, J.W. Salivary tumours. Periodontol. 2000 2011, 57, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kaye, F.J. Emerging Biology of Malignant Salivary Gland Tumors Offers New Insights into the Classification and Treatment of Mucoepidermoid Cancer. Clin. Cancer Res. 2006, 12, 3878–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafique, K.; Zhang, P.J.; Montone, K.T.; Song, S.; Livolsi, V.A.; Baloch, Z. Pathologic grading of mucoepidermoid carcinomas of the salivary gland and its effect on clinicopathologic follow-up: An institutional experience. Hum. Pathol. 2020, 98, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Seethala, R.R.; Dacic, S.; Cieply, K.; Kelly, L.M.; Nikiforova, M.N. A Reappraisal of the MECT1/MAML2 Translocation in Salivary Mucoepidermoid Carcinomas. Am. J. Surg. Pathol. 2010, 34, 1106–1121. [Google Scholar] [CrossRef]
- Birkeland, A.C.; Foltin, S.K.; Michmerhuizen, N.L.; Hoesli, R.C.; Rosko, A.J.; Byrd, S.; Yanik, M.; Nor, J.E.; Bradford, C.R.; Prince, M.E.; et al. Correlation of Crtc1/3-Maml2 fusion status, grade and survival in mucoepidermoid carcinoma. Oral Oncol. 2017, 68, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Tonon, G.; Modi, S.; Wu, L.; Kubo, A.; Coxon, A.B.; Komiya, T.; O’Neil, K.; Stover, K.; Elnaggar, A.K.; Griffin, J.D.; et al. t(11;19)(q21;p13) translocation in mucoepidermoid carcinoma creates a novel fusion product that disrupts a Notch signaling pathway. Nat. Genet. 2003, 33, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Tirado, Y.; Williams, M.D.; Hanna, E.; Kaye, F.J.; Batsakis, J.G.; El-Naggar, A.K. CRTC1/MAML2 fusion transcript in high grade mucoepidermoid carcinomas of salivary and thyroid glands and Warthin’s tumors: Implications for histogenesis and biologic behavior. Genes, Chromosom. Cancer 2007, 46, 708–715. [Google Scholar] [CrossRef]
- Okabe, M.; Miyabe, S.; Nagatsuka, H.; Terada, A.; Hanai, N.; Yokoi, M.; Shimozato, K.; Eimoto, T.; Nakamura, S.; Nagai, N.; et al. MECT1-MAML2 Fusion Transcript Defines a Favorable Subset of Mucoepidermoid Carcinoma. Clin. Cancer Res. 2006, 12, 3902–3907. [Google Scholar] [CrossRef] [Green Version]
- Behboudi, A.; Enlund, F.; Winnes, M.; Andrén, Y.; Nordkvist, A.; Leivo, I.; Flaberg, E.; Szekely, L.; Mäkitie, A.; Grenman, R.; et al. Molecular classification of mucoepidermoid carcinomas—Prognostic significance of theMECT1–MAML2 fusion oncogene. Genes Chromosom. Cancer 2006, 45, 470–481. [Google Scholar] [CrossRef]
- Saade, R.E.; Bell, D.; Garcia, J.; Roberts, D.; Weber, R. Role of CRTC1/MAML2 Translocation in the Prognosis and Clinical Outcomes of Mucoepidermoid Carcinoma. JAMA Otolaryngol. Neck Surg. 2016, 142, 234–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, I.D. t(11;19) translocation and CRTC1-MAML2 fusion oncogene in mucoepidermoid carcinoma. Oral Oncol. 2009, 45, 2–9. [Google Scholar] [CrossRef]
- Wu, L.; Liu, J.; Gao, P.; Nakamura, M.; Cao, Y.; Shen, H.; Griffin, J.D. Transforming activity of MECT1-MAML2 fusion oncoprotein is mediated by constitutive CREB activation. EMBO J. 2005, 24, 2391–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Chen, J.; Gu, Y.; Hu, C.; Li, J.-L.; Lin, S.; Shen, H.; Cao, C.; Gao, R.; Ha, P.K.; et al. Aberrantly activated AREG–EGFR signaling is required for the growth and survival of CRTC1–MAML2 fusion-positive mucoepidermoid carcinoma cells. Oncogene 2014, 33, 3869–3877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Ni, W.; Li, J.-L.; Lin, S.; Zhou, X.; Sun, Y.; Li, J.W.; Leon, M.E.; Hurtado, M.D.; Zolotukhin, S.; et al. The CRTC1-MAML2 fusion is the major oncogenic driver in mucoepidermoid carcinoma. J. Clin. Investig. 2021, 6, e139497. [Google Scholar] [CrossRef]
- Brandwein, M.S.; Ivanov, K.; Wallace, D.I.; Hille, J.J.; Wang, B.; Fahmy, A.; Bodian, C.; Urken, M.L.; Gnepp, D.R.; Huvos, A.; et al. Mucoepidermoid Carcinoma: A Clinicopathologic Study of 80 Patients with Special Reference to Histological Grading. Am. J. Surg. Pathol. 2001, 25, 835–845. [Google Scholar] [CrossRef]
- Goode, R.K.; Auclair, P.L.; Eihs, G.L. Mucoepidermoid carcinoma of the major salivary glands: Clinical and histopathologic analysis of 234 cases with evaluation of grading criteria. Cancer 1998, 82, 1217–1224. [Google Scholar] [CrossRef]
- Nascimento, A.G.; Amaral, A.L.P.; Prado, L.A.F.; Kligerman, J.; Silveira, T.R.P. Mucoepidermoid carcinoma of salivary glands: A clinicopathologic study of 46 cases. Head Neck Surg. 1986, 8, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Yih, W.-Y.; Kratochvil, F.J.; Stewart, J.C. Intraoral Minor Salivary Gland Neoplasms: Review of 213 Cases. J. Oral Maxillofac. Surg. 2005, 63, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Haderlein, M.; Lettmaier, S.; Agaimy, A.; Haller, F.; Hecht, M.; Fietkau, R.; Iro, H.; Mantsopoulos, K. Targeted Therapy, Chemotherapy, Immunotherapy and Novel Treatment Options for Different Subtypes of Salivary Gland Cancer. J. Clin. Med. 2022, 11, 720. [Google Scholar] [CrossRef]
- Noguchi, K.; Kanda, S.; Yoshida, K.; Funaoka, Y.; Yamanegi, K.; Yoshikawa, K.; Takaoka, K.; Kishimoto, H.; Nakano, Y. Establishment of a patient-derived mucoepidermoid carcinoma cell line with the CRTC1-MAML2 fusion gene. Mol. Clin. Oncol. 2022, 16, 75. [Google Scholar] [CrossRef] [PubMed]
- Warner, K.A.; Adams, A.; Bernardi, L.; Nor, C.; Finkel, K.A.; Zhang, Z.; McLean, S.A.; Helman, J.; Wolf, G.T.; Divi, V.; et al. Characterization of tumorigenic cell lines from the recurrence and lymph node metastasis of a human salivary mucoepidermoid carcinoma. Oral Oncol. 2013, 49, 1059–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenman, R.; Pekkola-Heino, K.; Joensuu, H.; Aitasalo, K.; Klemi, P.; Lakkala, T. UT-MUC-1, a New Mucoepidermoid Carcinoma Cell Line, and Its Radiosensitivity. JAMA Otolaryngol. Neck Surg. 1992, 118, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Reid, V.; Yu, Z.; Schuman, T.; Li, S.; Singh, P.; Fong, Y.; Wong, R.J. Herpes oncolytic therapy of salivary gland carcinomas. Int. J. Cancer 2008, 122, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Banks-Schlegel, S.P.; Gazdar, A.F.; Harris, C.C. Intermediate filament and cross-linked envelope expression in human lung tumor cell lines. Cancer Res. 1985, 45, 1187–1197. [Google Scholar]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Ni, W.; Chen, Z.; Zhou, X.; Yang, R.; Yu, M.; Lu, J.; Kaye, F.J.; Wu, L. Targeting Notch and EGFR signaling in human mucoepidermoid carcinoma. Signal Transduct. Target. Ther. 2021, 6, 27. [Google Scholar] [CrossRef]
- Pao, W.; Chmielecki, J. Rational, biologically based treatment of EGFR-mutant non-small-cell lung cancer. Nat. Rev. Cancer 2010, 10, 760–774. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, E.; Togashi, Y.; Takeuchi, Y.; Shinya, S.; Tada, Y.; Kataoka, K.; Tane, K.; Sato, E.; Ishii, G.; Goto, K.; et al. Blockade of EGFR improves responsiveness to PD-1 blockade in EGFR -mutated non–small cell lung cancer. Sci. Immunol. 2020, 5, eaav3937. [Google Scholar] [CrossRef]
- Kondo, Y.; Suzuki, S.; Ono, S.; Goto, M.; Miyabe, S.; Ogawa, T.; Tsuchida, H.; Ito, H.; Takahara, T.; Satou, A.; et al. In Situ PD-L1 Expression in Oral Squamous Cell Carcinoma Is Induced by Heterogeneous Mechanisms among Patients. Int. J. Mol. Sci. 2022, 23, 4077. [Google Scholar] [CrossRef]
- Marabelle, A.; Fakih, M.; Lopez, J.; Shah, M.; Shapira-Frommer, R.; Nakagawa, K.; Chung, H.C.; Kindler, H.L.; Lopez-Martin, J.A.; Miller, W.H., Jr.; et al. Association of tumour mutational burden with outcomes in patients with advanced solid tumours treated with pembrolizumab: Prospective biomarker analysis of the multicohort, open-label, phase 2 KEYNOTE-158 study. Lancet Oncol. 2020, 21, 1353–1365. [Google Scholar] [CrossRef]
- Farhat, R.; Asna, N.; Avraham, Y.; Khater, A.; Asakla, M.; Safia, A.; Szvalb, S.; Elkhatib, N.; Merchavy, S. Pembrolizumab as a first line therapy in a patient with extensive mucoepidermoid salivary gland carcinoma. A complete clinical, radiological and pathological response. A very specific case. Discov. Oncol. 2022, 13, 37. [Google Scholar] [CrossRef]
- Tagawa, H.; Karnan, S.; Kasugai, Y.; Tuzuki, S.; Suzuki, R.; Hosokawa, Y.; Seto, M. MASL1, a candidate oncogene found in amplification at 8p23.1, is translocated in immunoblastic B cell lymphoma cell line OCI-LY8. Oncogene 2004, 23, 2576–2581. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cibulskis, K.; Lawrence, M.S.; Carter, S.L.; Sivachenko, A.; Jaffe, D.B.; Sougnez, C.; Gabriel, S.B.; Meyerson, M.L.; Lander, E.S.; Getz, G. Sensitive detection of somatic point mutations in impure and heterogeneous cancer samples. Nat. Biotechnol. 2013, 31, 213–219. [Google Scholar] [CrossRef]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Wilm, A.; Aw, P.P.K.; Bertrand, D.; Yeo, G.H.T.; Ong, S.H.; Wong, C.H.; Khor, C.C.; Petric, R.; Hibberd, M.L.; Nagarajan, N. LoFreq: A sequence-quality aware, ultra-sensitive variant caller for uncovering cell-population heterogeneity from high-throughput sequencing datasets. Nucleic Acids Res. 2012, 40, 11189–11201. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamanaka, S.; Suzuki, S.; Ito, H.; Sivasundaram, K.; Hanamura, I.; Okubo, I.; Yoshikawa, K.; Ono, S.; Takahara, T.; Satou, A.; et al. Establishment of Mucoepidermoid Carcinoma Cell Lines from Surgical and Recurrence Biopsy Specimens. Int. J. Mol. Sci. 2023, 24, 1722. https://doi.org/10.3390/ijms24021722
Yamanaka S, Suzuki S, Ito H, Sivasundaram K, Hanamura I, Okubo I, Yoshikawa K, Ono S, Takahara T, Satou A, et al. Establishment of Mucoepidermoid Carcinoma Cell Lines from Surgical and Recurrence Biopsy Specimens. International Journal of Molecular Sciences. 2023; 24(2):1722. https://doi.org/10.3390/ijms24021722
Chicago/Turabian StyleYamanaka, Shunpei, Susumu Suzuki, Hideaki Ito, Karnan Sivasundaram, Ichiro Hanamura, Ikuko Okubo, Kazuhiro Yoshikawa, Shoya Ono, Taishi Takahara, Akira Satou, and et al. 2023. "Establishment of Mucoepidermoid Carcinoma Cell Lines from Surgical and Recurrence Biopsy Specimens" International Journal of Molecular Sciences 24, no. 2: 1722. https://doi.org/10.3390/ijms24021722