
HIV Replication Increases the Mitochondrial DNA Content of Plasma Extracellular Vesicles
 , and
, and
Abstract
:1. Introduction
2. Results
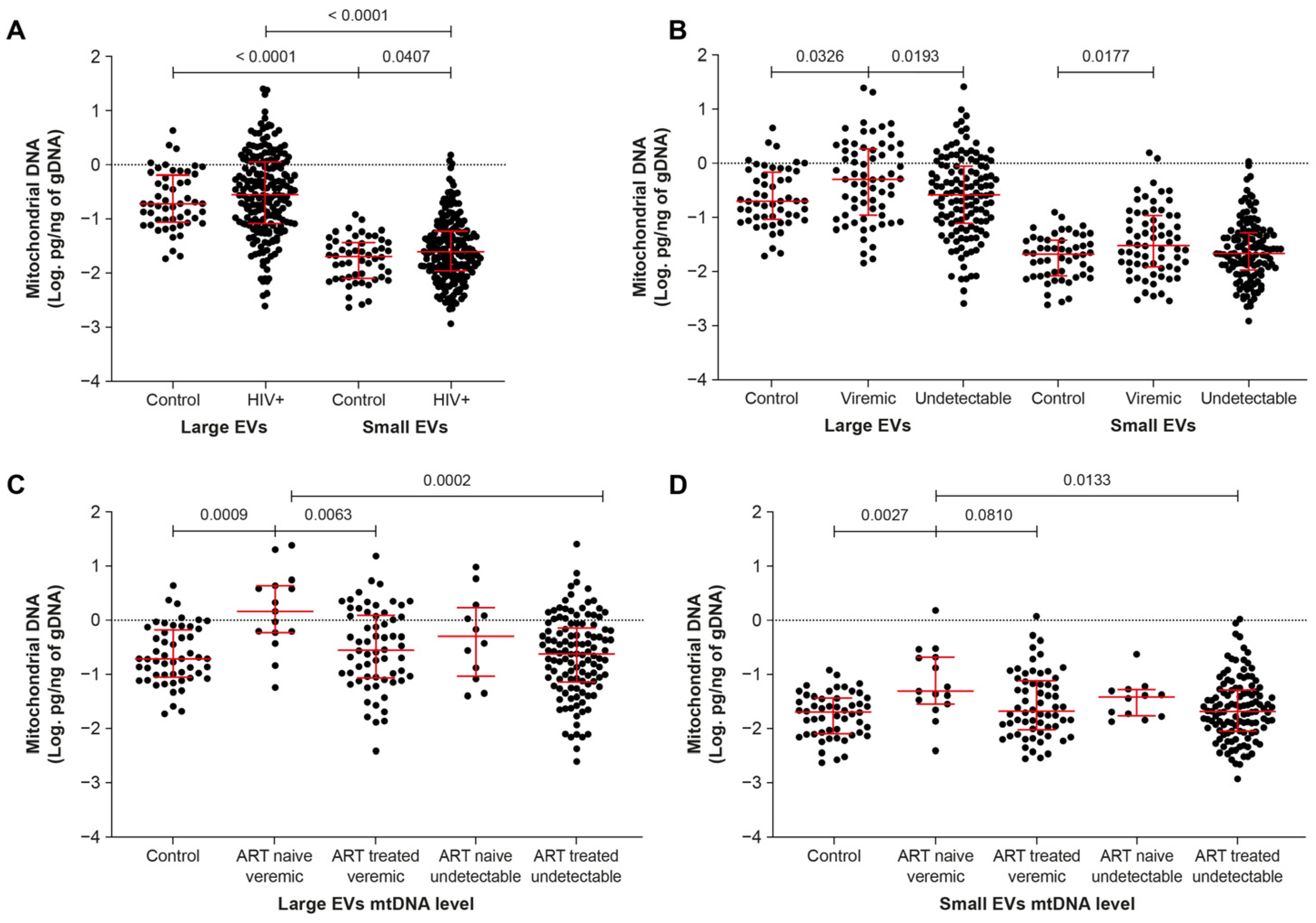
2.1. Mitochondrial DNA Is More Abundant in EVs of ART-Naive Viremic Patients
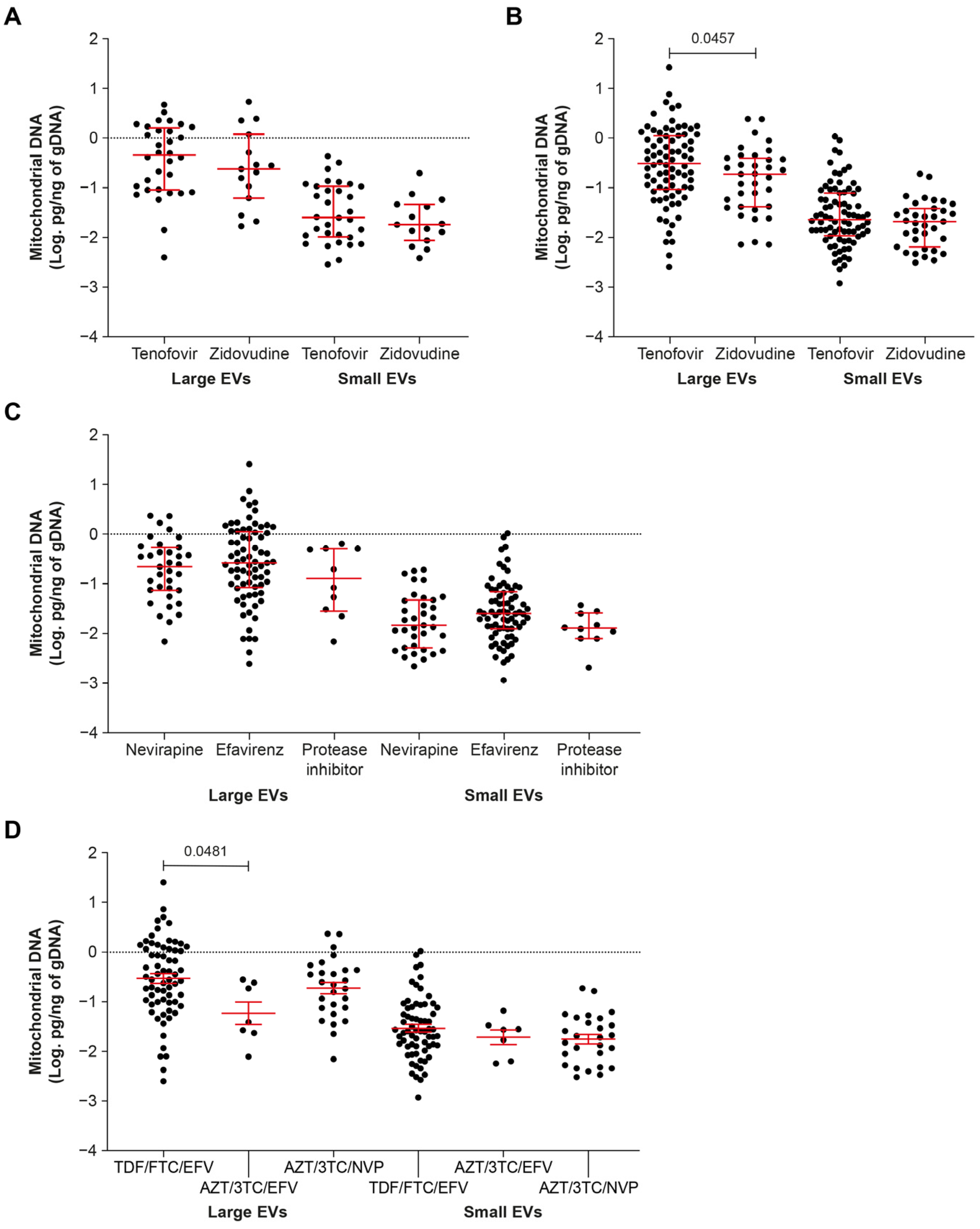
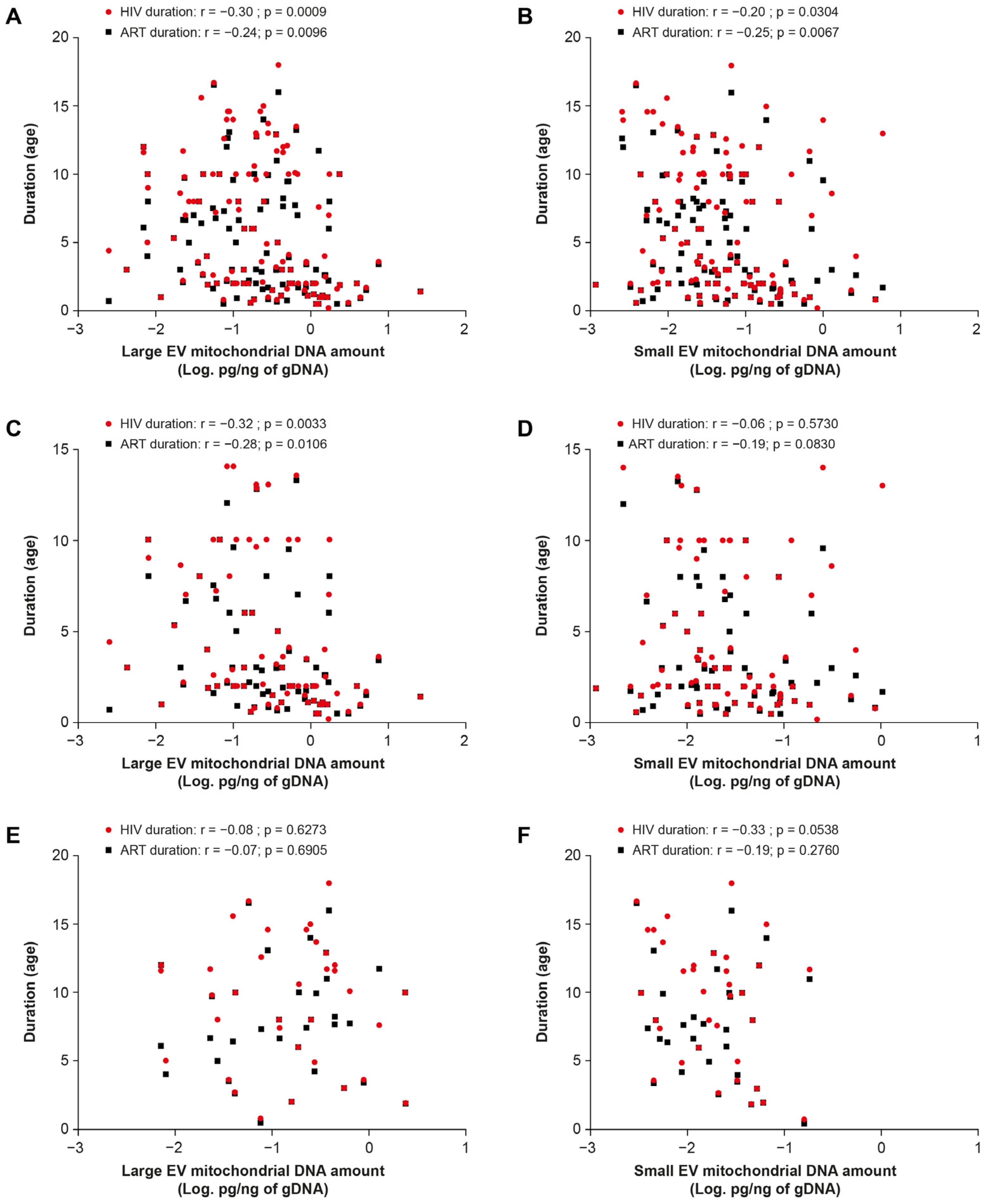
2.2. ART including Tenofovir Increases the mtDNA Content of EVs
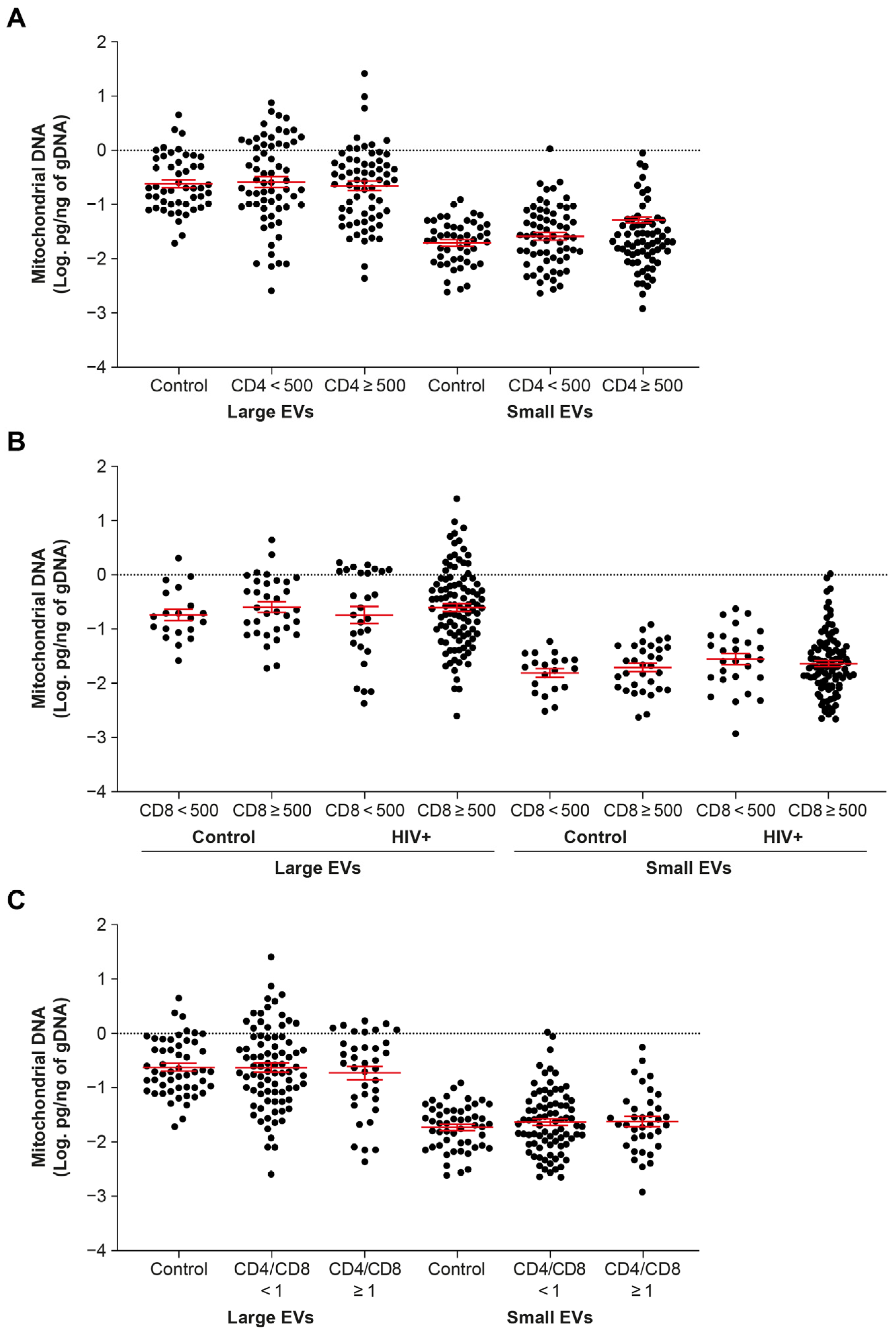
2.3. EV-Borne mtDNA Abundance Appears Indifferent to CD4 and CD8 T Cell Counts
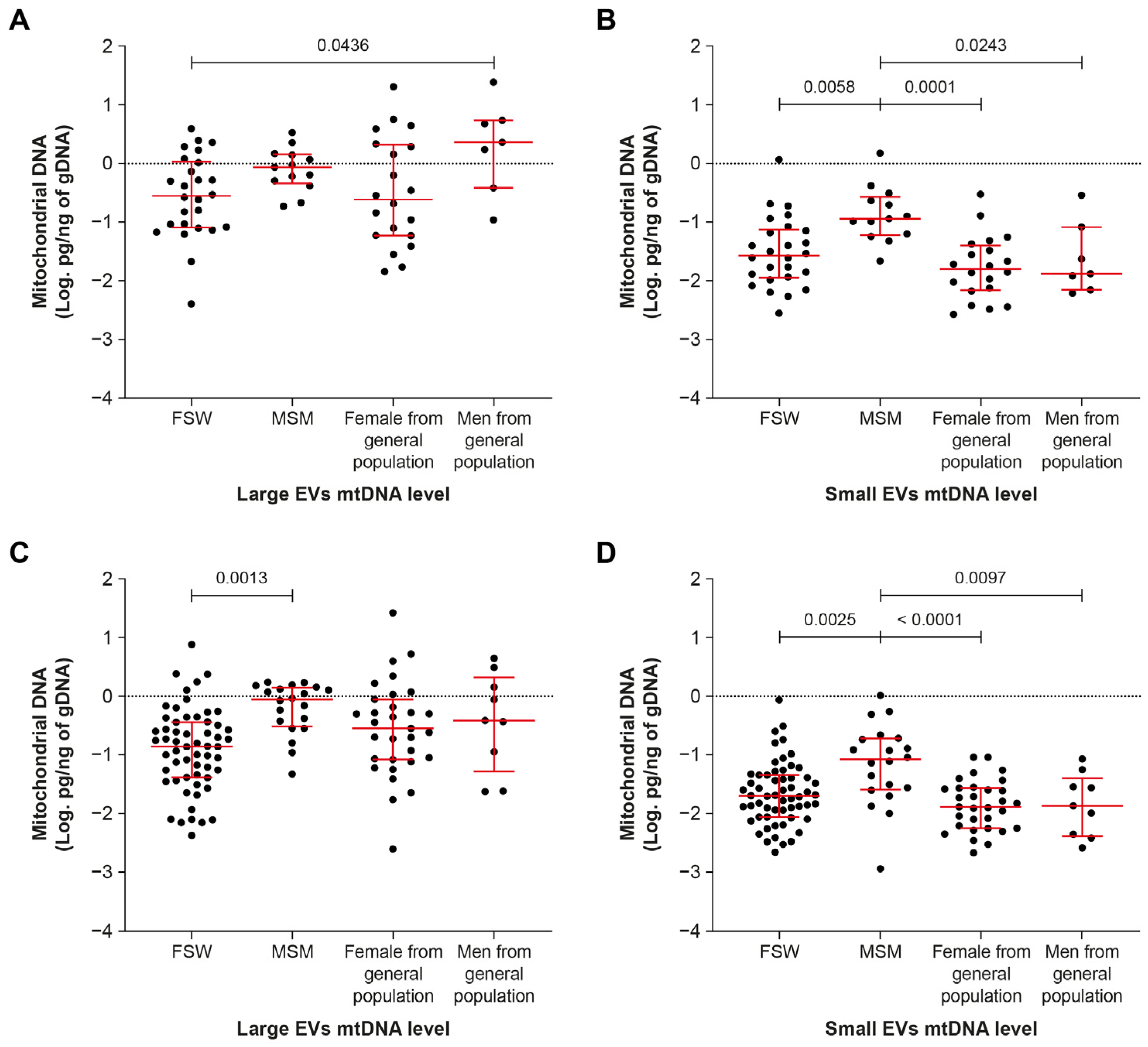
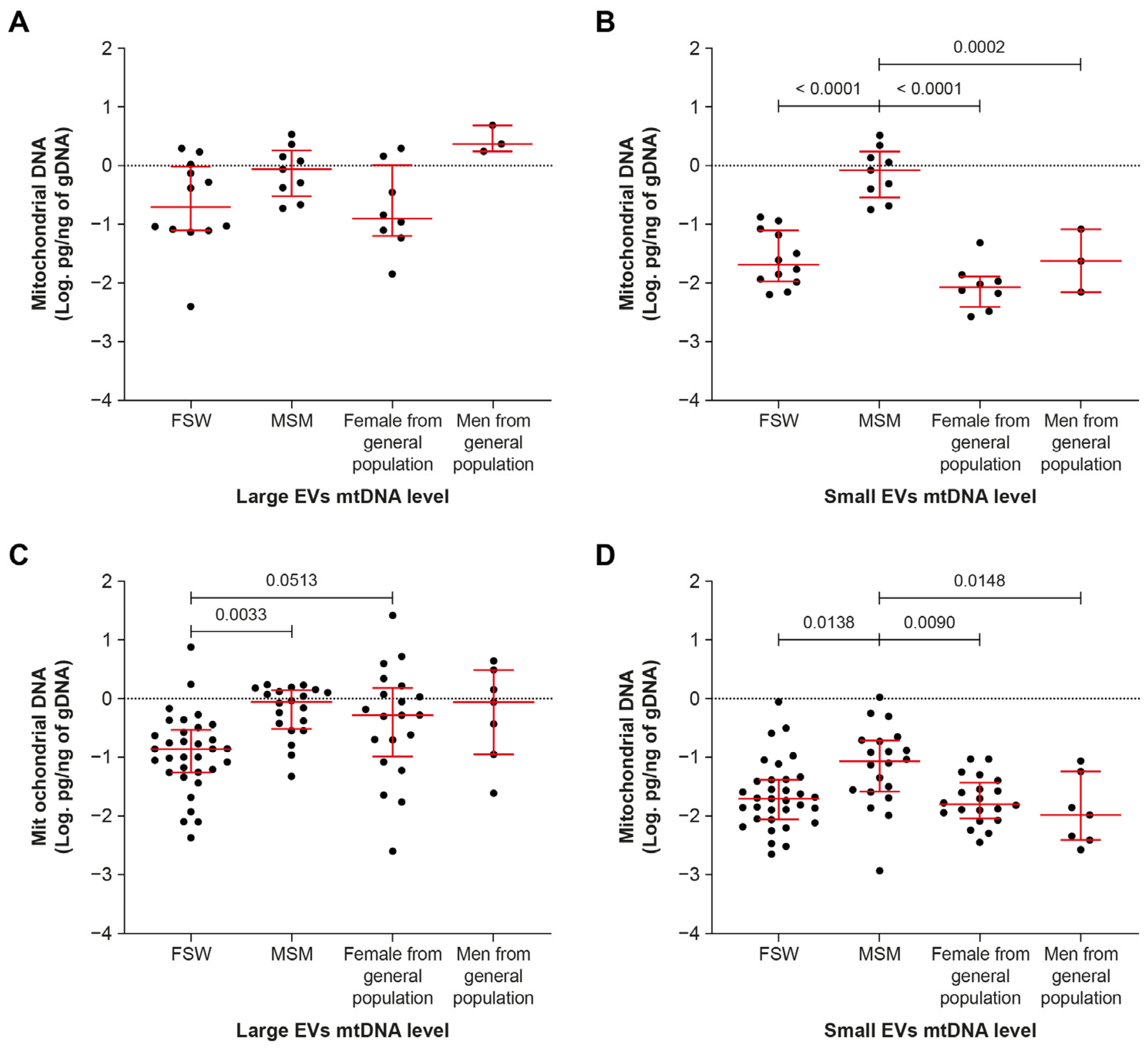
2.4. Men Who Have Sex with Men Have More Small-EV-Borne mtDNA
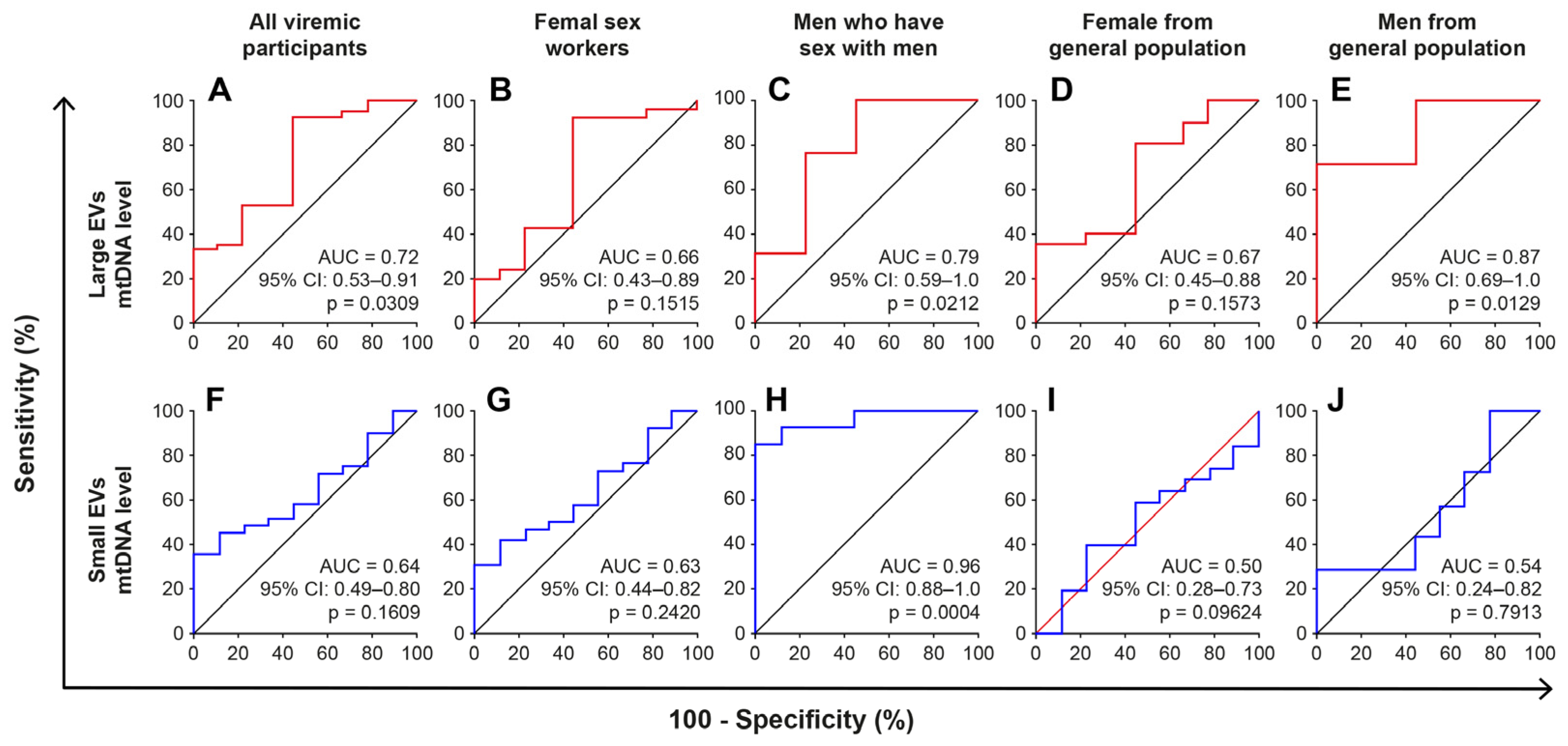
2.5. The mtDNA Content of Large EVs Can Distinguish HIV+ Viremic Patients
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Quantitation of HIV-1 RNA, CD4 and CD8 T Lymphocytes
4.3. Purification of Extracellular Vesicles
4.4. EV Size Measurement
4.5. EV Flow Cytometry Analysis
4.6. Detergent Treatment of Extracellular Vesicles
4.7. DNA Extraction and Mitochondrial DNA Quantification
4.8. Transmission Electron Microscopy
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatischeff, I. Current Search through Liquid Biopsy of Effective Biomarkers for Early Cancer Diagnosis into the Rich Cargoes of Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 5674. [Google Scholar] [CrossRef]
- Nazarenko, I. Extracellular Vesicles: Recent Developments in Technology and Perspectives for Cancer Liquid Biopsy. Recent Results Cancer Res. 2020, 215, 319–344. [Google Scholar] [CrossRef]
- Hubert, A.; Subra, C.; Jenabian, M.A.; Tremblay Labrecque, P.F.; Tremblay, C.; Laffont, B.; Provost, P.; Routy, J.P.; Gilbert, C. Elevated Abundance, Size, and MicroRNA Content of Plasma Extracellular Vesicles in Viremic HIV-1+ Patients: Correlations With Known Markers of Disease Progression. J. Acquir. Immune Defic. Syndr. 2015, 70, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Bazié, W.W.; Boucher, J.; Traoré, I.T.; Kania, D.; Somé, D.Y.; Alary, M.; Gilbert, C. Vesicular MicroRNA as Potential Biomarkers of Viral Rebound. Cells 2022, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Bazié, W.W.; Boucher, J.; Vitry, J.; Goyer, B.; Routy, J.P.; Tremblay, C.; Trottier, S.; Jenabian, M.A.; Provost, P.; Alary, M.; et al. Plasma Extracellular Vesicle Subtypes May be Useful as Potential Biomarkers of Immune Activation in People With HIV. Pathog. Immun. 2021, 6, 1–28. [Google Scholar] [CrossRef]
- Bazié, W.W.; Goyer, B.; Boucher, J.; Zhang, Y.; Planas, D.; Chatterjee, D.; Routy, J.P.; Alary, M.; Ancuta, P.; Gilbert, C. Diurnal Variation of Plasma Extracellular Vesicle Is Disrupted in People Living with HIV. Pathogens 2021, 10, 518. [Google Scholar] [CrossRef]
- Madison, M.N.; Okeoma, C.M. Exosomes: Implications in HIV-1 Pathogenesis. Viruses 2015, 7, 4093–4118. [Google Scholar] [CrossRef] [Green Version]
- Ellwanger, J.H.; Veit, T.D.; Chies, J.A.B. Exosomes in HIV infection: A review and critical look. Infect. Genet. Evol. 2017, 53, 146–154. [Google Scholar] [CrossRef]
- Ganta, K.K.; Chaubey, B. Mitochondrial dysfunctions in HIV infection and antiviral drug treatment. Expert Opin. Drug Metab. Toxicol. 2019, 15, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; Kelly, B.; O’Neill, L.A.J. Mitochondria are the powerhouses of immunity. Nat. Immunol. 2017, 18, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Koshiba, T.; Bashiruddin, N.; Kawabata, S. Mitochondria and antiviral innate immunity. Int. J. Biochem. Mol. Biol. 2011, 2, 257–262. [Google Scholar] [PubMed]
- Tiku, V.; Tan, M.W.; Dikic, I. Mitochondrial Functions in Infection and Immunity. Trends Cell Biol. 2020, 30, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annesley, S.J.; Fisher, P.R. Mitochondria in Health and Disease. Cells 2019, 8, 680. [Google Scholar] [CrossRef] [Green Version]
- Schank, M.; Zhao, J.; Moorman, J.P.; Yao, Z.Q. The impact of HIV-and ART-induced mitochondrial dysfunction in cellular senescence and aging. Cells 2021, 10, 174. [Google Scholar] [CrossRef]
- Arts, E.J.; Hazuda, D.J. HIV-1 antiretroviral drug therapy. Cold Spring Harb. Perspect. Med. 2012, 2, a007161. [Google Scholar] [CrossRef] [Green Version]
- Pau, A.K.; George, J.M. Antiretroviral therapy: Current drugs. Infect. Dis. Clin. N. Am. 2014, 28, 371–402. [Google Scholar] [CrossRef] [Green Version]
- Apostolova, N.; Blas-García, A.; Esplugues, J.V. Mitochondrial interference by anti-HIV drugs: Mechanisms beyond Pol-γ inhibition. Trends Pharmacol. Sci. 2011, 32, 715–725. [Google Scholar] [CrossRef]
- Kakuda, T.N. Pharmacology of nucleoside and nucleotide reverse transcriptase inhibitor-induced mitochondrial toxicity. Clin. Ther. 2000, 22, 685–708. [Google Scholar] [CrossRef]
- Côté, H.C.; Brumme, Z.L.; Craib, K.J.; Alexander, C.S.; Wynhoven, B.; Ting, L.; Wong, H.; Harris, M.; Harrigan, P.R.; O’Shaughnessy, M.V.; et al. Changes in mitochondrial DNA as a marker of nucleoside toxicity in HIV-infected patients. N. Engl. J. Med. 2002, 346, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Hammond, E.; Nolan, D.; James, I.; Metcalf, C.; Mallal, S. Reduction of mitochondrial DNA content and respiratory chain activity occurs in adipocytes within 6-12 months of commencing nucleoside reverse transcriptase inhibitor therapy. Aids 2004, 18, 815–817. [Google Scholar] [CrossRef] [PubMed]
- Dalakas, M.C.; Semino-Mora, C.; Leon-Monzon, M. Mitochondrial alterations with mitochondrial DNA depletion in the nerves of AIDS patients with peripheral neuropathy induced by 2′3′-dideoxycytidine (ddC). Lab. Investing. 2001, 81, 1537–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernas, B.; Rego-Pérez, I.; Tabernilla, A.; Balboa, V.; Relaño, S.; Grandal, M.; Crespo, M.; Mena, Á.; Castro-Iglesias, Á.; Blanco, F.J.; et al. Plasma mitochondrial DNA levels are inversely associated with HIV-RNA levels and directly with CD4 counts: Potential role as a biomarker of HIV replication. J. Antimicrob. Chemother. 2017, 72, 3159–3162. [Google Scholar] [CrossRef] [PubMed]
- Maagaard, A.; Holberg-Petersen, M.; Løvgården, G.; Holm, M.; Pettersen, F.O.; Kvale, D. Distinct mechanisms for mitochondrial DNA loss in T and B lymphocytes from HIV-infected patients exposed to nucleoside reverse-transcriptase inhibitors and those naive to antiretroviral treatment. J. Infect. Dis. 2008, 198, 1474–1481. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Goto, M.; Hosoya, N.; Odawara, T.; Kitamura, Y.; Nakamura, T.; Iwamoto, A. Depletion of mitochondrial DNA in HIV-1-infected patients and its amelioration by antiretroviral therapy. J. Med. Virol. 2003, 70, 497–505. [Google Scholar] [CrossRef]
- Venhoff, N.; Setzer, B.; Melkaoui, K.; Walker, U.A. Mitochondrial toxicity of tenofovir, emtricitabine and abacavir alone and in combination with additional nucleoside reverse transcriptase inhibitors. Antivir. Ther. 2007, 12, 1075–1085. [Google Scholar] [CrossRef]
- Blas-García, A.; Polo, M.; Alegre, F.; Funes, H.A.; Martínez, E.; Apostolova, N.; Esplugues, J.V. Lack of mitochondrial toxicity of darunavir, raltegravir and rilpivirine in neurons and hepatocytes: A comparison with efavirenz. J. Antimicrob. Chemother. 2014, 69, 2995–3000. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Treviño, P.; Velásquez, M.; García, N. Mechanisms of mitochondrial DNA escape and its relationship with different metabolic diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165761. [Google Scholar] [CrossRef]
- Hayakawa, K.; Esposito, E.; Wang, X.; Terasaki, Y.; Liu, Y.; Xing, C.; Ji, X.; Lo, E.H. Transfer of mitochondria from astrocytes to neurons after stroke. Nature 2016, 535, 551–555. [Google Scholar] [CrossRef]
- Davis, C.H.; Kim, K.Y.; Bushong, E.A.; Mills, E.A.; Boassa, D.; Shih, T.; Kinebuchi, M.; Phan, S.; Zhou, Y.; Bihlmeyer, N.A.; et al. Transcellular degradation of axonal mitochondria. Proc. Natl. Acad. Sci. USA 2014, 111, 9633–9638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcoux, G.; Magron, A.; Sut, C.; Laroche, A.; Laradi, S.; Hamzeh-Cognasse, H.; Allaeys, I.; Cabon, O.; Julien, A.S.; Garraud, O.; et al. Platelet-derived extracellular vesicles convey mitochondrial DAMPs in platelet concentrates and their levels are associated with adverse reactions. Transfusion 2019, 59, 2403–2414. [Google Scholar] [CrossRef] [PubMed]
- Lazo, S.; Noren Hooten, N.; Green, J.; Eitan, E.; Mode, N.A.; Liu, Q.R.; Zonderman, A.B.; Ezike, N.; Mattson, M.P.; Ghosh, P.; et al. Mitochondrial DNA in extracellular vesicles declines with age. Aging Cell 2021, 20, e13283. [Google Scholar] [CrossRef] [PubMed]
- Sansone, P.; Savini, C.; Kurelac, I.; Chang, Q.; Amato, L.B.; Strillacci, A.; Stepanova, A.; Iommarini, L.; Mastroleo, C.; Daly, L.; et al. Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy-resistant breast cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9066–E9075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, J.S.; Tait, S.W. Mitochondrial DNA in inflammation and immunity. EMBO Rep. 2020, 21, e49799. [Google Scholar] [CrossRef]
- van der Wijst, M.G.; van Tilburg, A.Y.; Ruiters, M.H.; Rots, M.G. Experimental mitochondria-targeted DNA methylation identifies GpC methylation, not CpG methylation, as potential regulator of mitochondrial gene expression. Sci. Rep. 2017, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Bellizzi, D.; D’Aquila, P.; Scafone, T.; Giordano, M.; Riso, V.; Riccio, A.; Passarino, G. The control region of mitochondrial DNA shows an unusual CpG and non-CpG methylation pattern. DNA Res. 2013, 20, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Bazié, W.W.; Boucher, J.; Traoré, I.T.; Kania, D.; Somé, D.Y.; Alary, M.; Gilbert, C. Plasma Vesicular miR-155 as a Biomarker of Immune Activation in Antiretroviral Treated People Living with HIV. Front. Immunol. 2022, 13, 916599. [Google Scholar] [CrossRef]
- Marcatti, M.; Saada, J.; Okereke, I.; Wade, C.E.; Bossmann, S.H.; Motamedi, M.; Szczesny, B. Quantification of Circulating Cell Free Mitochondrial DNA in Extracellular Vesicles with PicoGreen™ in Liquid Biopsies: Fast Assessment of Disease/Trauma Severity. Cells 2021, 10, 819. [Google Scholar] [CrossRef]
- Garcia-Martinez, I.; Santoro, N.; Chen, Y.; Hoque, R.; Ouyang, X.; Caprio, S.; Shlomchik, M.J.; Coffman, R.L.; Candia, A.; Mehal, W.Z. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J. Clin. Investing. 2016, 126, 859–864. [Google Scholar] [CrossRef]
- Boudreau, L.H.; Duchez, A.C.; Cloutier, N.; Soulet, D.; Martin, N.; Bollinger, J.; Paré, A.; Rousseau, M.; Naika, G.S.; Lévesque, T.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood 2014, 124, 2173–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, A.; Burch, A.; Dave, K.M.; Sreeram, A.; Reynolds, M.J.; Dobbins, D.X.; Kamte, Y.S.; Zhao, W.; Sabatelle, C.; Joy, G.M.; et al. Microvesicles transfer mitochondria and increase mitochondrial function in brain endothelial cells. J. Control. Release 2021, 338, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Pinti, M.; Nasi, M.; Gibellini, L.; Manzini, S.; Roat, E.; De Biasi, S.; Bertoncelli, L.; Montagna, J.P.; Bisi, L.; et al. Increased plasma levels of extracellular mitochondrial DNA during HIV infection: A new role for mitochondrial damage-associated molecular patterns during inflammation. Mitochondrion 2011, 11, 750–755. [Google Scholar] [CrossRef]
- Thaker, S.K.; Ch’ng, J.; Christofk, H.R. Viral hijacking of cellular metabolism. BMC Biol. 2019, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Elesela, S.; Lukacs, N.W. Role of Mitochondria in Viral Infections. Life 2021, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Ohta, A.; Nishiyama, Y. Mitochondria and viruses. Mitochondrion 2011, 11, 1–12. [Google Scholar] [CrossRef]
- Dinkins, C.; Arko-Mensah, J.; Deretic, V. Autophagy and HIV. Semin. Cell Dev. Biol. 2010, 21, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Chen, L.; Song, Z.; He, H. The fate of damaged mitochondrial DNA in the cell. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119233. [Google Scholar] [CrossRef]
- Szczesny, B.; Marcatti, M.; Ahmad, A.; Montalbano, M.; Brunyánszki, A.; Bibli, S.I.; Papapetropoulos, A.; Szabo, C. Mitochondrial DNA damage and subsequent activation of Z-DNA binding protein 1 links oxidative stress to inflammation in epithelial cells. Sci. Rep. 2018, 8, 914. [Google Scholar] [CrossRef] [Green Version]
- Soubannier, V.; McLelland, G.L.; Zunino, R.; Braschi, E.; Rippstein, P.; Fon, E.A.; McBride, H.M. A vesicular transport pathway shuttles cargo from mitochondria to lysosomes. Curr. Biol. 2012, 22, 135–141. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, M.J.; Koritzinsky, E.H.; Zhou, Z.; Wang, W.; Cao, H.; Yuen, P.S.; Ross, R.A.; Star, R.A.; Liangpunsakul, S.; et al. Mitochondrial DNA-enriched microparticles promote acute-on-chronic alcoholic neutrophilia and hepatotoxicity. JCI Insight 2017, 2, e92634. [Google Scholar] [CrossRef] [PubMed]
- Koczor, C.A.; Lewis, W. Nucleoside reverse transcriptase inhibitor toxicity and mitochondrial DNA. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.; Copeland, W.C.; Day, B.J. Mitochondrial dna depletion, oxidative stress, and mutation: Mechanisms of dysfunction from nucleoside reverse transcriptase inhibitors. Lab. Investig. 2001, 81, 777–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinti, M.; Salomoni, P.; Cossarizza, A. Anti-HIV drugs and the mitochondria. Biochim. Biophys. Acta 2006, 1757, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Lewis, W.; Day, B.J.; Copeland, W.C. Mitochondrial toxicity of NRTI antiviral drugs: An integrated cellular perspective. Nat. Rev. Drug Discov. 2003, 2, 812–822. [Google Scholar] [CrossRef]
- Walker, U.A.; Setzer, B.; Venhoff, N. Increased long-term mitochondrial toxicity in combinations of nucleoside analogue reverse-transcriptase inhibitors. Aids 2002, 16, 2165–2173. [Google Scholar] [CrossRef] [Green Version]
- Birkus, G.; Hitchcock, M.J.; Cihlar, T. Assessment of mitochondrial toxicity in human cells treated with tenofovir: Comparison with other nucleoside reverse transcriptase inhibitors. Antimicrob. Agents Chemother. 2002, 46, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Bañó, M.; Morén, C.; Barroso, S.; Juárez, D.L.; Guitart-Mampel, M.; González-Casacuberta, I.; Canto-Santos, J.; Lozano, E.; León, A.; Pedrol, E.; et al. Mitochondrial Toxicogenomics for Antiretroviral Management: HIV Post-exposure Prophylaxis in Uninfected Patients. Front. Genet. 2020, 11, 497. [Google Scholar] [CrossRef]
- Suo, Z.; Johnson, K.A. Selective inhibition of HIV-1 reverse transcriptase by an antiviral inhibitor, (R)-9-(2-Phosphonylmethoxypropyl)adenine. J. Biol. Chem. 1998, 273, 27250–27258. [Google Scholar] [CrossRef] [Green Version]
- Birkus, G.; Hájek, M.; Kramata, P.; Votruba, I.; Holý, A.; Otová, B. Tenofovir diphosphate is a poor substrate and a weak inhibitor of rat DNA polymerases alpha, delta, and epsilon*. Antimicrob. Agents Chemother. 2002, 46, 1610–1613. [Google Scholar] [CrossRef]
- Maagaard, A.; Kvale, D. Long term adverse effects related to nucleoside reverse transcriptase inhibitors: Clinical impact of mitochondrial toxicity. Scand. J. Infect. Dis. 2009, 41, 808–817. [Google Scholar] [CrossRef]
- van der Heijden, W.A.; van de Wijer, L.; Jaeger, M.; Grintjes, K.; Netea, M.G.; Urbanus, R.T.; van Crevel, R.; van den Heuvel, L.P.; Brink, M.; Rodenburg, R.J.; et al. Long-term treated HIV infection is associated with platelet mitochondrial dysfunction. Sci. Rep. 2021, 11, 6246. [Google Scholar] [CrossRef] [PubMed]
- EV-TRACK Consortium; Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods 2017, 14, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, T.; Shibayama, M. Measurement of Particle Size Distribution in Turbid Solutions by Dynamic Light Scattering Microscopy. J. Vis. Exp. 2017, 119, e54885. [Google Scholar] [CrossRef] [Green Version]
- Osteikoetxea, X.; Sódar, B.; Németh, A.; Szabó-Taylor, K.; Pálóczi, K.; Vukman, K.V.; Tamási, V.; Balogh, A.; Kittel, Á.; Pállinger, É.; et al. Differential detergent sensitivity of extracellular vesicle subpopulations. Org. Biomol. Chem. 2015, 13, 9775–9782. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HIV–Control (Never Infected) n = 53 | HIV+ n = 196 | p Value | HIV+ ART > 6 Months, VL Undetectable n = 118 | HIV+ ART-Naïve, VL Undetectable n = 12 | p Value | HIV+ ART > 6 Months, VL Detectable n = 51 | HIV+ ART-Naïve, VL Detectable n = 15 | p Value | |
|---|---|---|---|---|---|---|---|---|---|
| Male: n (%) | 14 (26.4) | 54 (27.5) | 0.8692 | 29 (24.6) | 5 (41.67) | 0.1993 | 14 (27.45) | 6 (40.00) | 0.3525 |
| Female sex workers: n (%) | - | 85 (43.4) | - | 58 (49.1) | 1 (8.3) | 0.0002 | 24 (47.1) | 2 (13.3) | 0.0870 |
| Men who have sex with men: n (%) | - | 33 (16.8) | - | 20 (16.9) | 0 (0.0) | 9 (17.6) | 4 (26.7) | ||
| Age in years, median (IQR) | 25 (22–32) | 37 (31–44) | <0.0001 | 38 (32–44) | 43 (28–52) | 0.2072 | 35 (30–42) | 35 (26–42) | 0.4629 |
| HIV+ duration, median months (IQR) | - | 43 (22–121) | - | 50 (24–120) | 24 (12–72) | 0.1229 | 43 (24–117) | 8 (1–114) | 0.4643 |
| CD4 T cells/µL, median (IQR) | 937 (800–1316) | 473 (340–696) | <0.0001 | 492 (368–750) | 717 (466–875) | 0.1057 | 372 (188–507) | 632 (304–798) | 0.0691 |
| CD8 T cells/µL, median (IQR) | 564 (451–717) | 757 (535–1011) | <0.0001 | 739 (536–1089) | 650 (398–759) | 0.1245 | 856 (544–1016) | 791 (556–1016) | 0.4482 |
| CD4/CD8 ratio, median (IQR) | 1.8 (1.4–2.1) | 0.6 (0.4–1.0) | <0.0001 | 0.7 (0.5–1.0) | 1.2 (0.7–1.5) | 0.0028 | 0.4 (0.2–0.7) | 0.7 (0.5–1.2) | 0.0120 |
| ART, n (%) | - | 169 (86.22) | 118 (100) | 0 | 51 (100) | 0 | |||
| NRTI ART drugs, n (%) | - | ||||||||
| Abacavir | - | 7 (4.1) | 3 (2.5) | - | 4 (7.8) | - | |||
| Tenofovir | - | 112 (66.3) | 80 (67.8) | - | 32 (62.7) | - | |||
| Zidovudine | - | 50 (29.6) | 35 (29.7) | - | 15 (29.4) | - | |||
| NNRTI ART drugs or PI, n (%) | - | ||||||||
| Efavirenz | - | 104 (61.5) | 73 (61.9) | - | 31 (60.8) | - | |||
| Nevirapine | - | 46 (27.2) | 35 (29.7) | - | 11 (21.6) | - | |||
| Protease inhibitors | - | 19 (11.2) | 10 (8.5) | - | 9 (17.6) | - | |||
| Months on ART, median (IQR) | - | 36 (21–88) | 38 (21–92) | - | 24 (21–69) | - | |||
| HIV undetectable (<20 copies/mL) while on ART, n (%) | - | 118 (69.8) | - | - | - | - | |||
| HIV load, median copies/mL (IQR) | - | 2.841 (82–29.240) | - | - | 1.606 (77–22.510) | 13.174 (2.297–70.711) | 0.2451 |
| Female Sex Workers (n = 58) | Men Who Have Sex with Men (n = 20) | Females from General Population (n = 31) | Males from General Population (n = 9) | p Value | |
|---|---|---|---|---|---|
| Age, median (IQR) | 38 (33–44) | 30 (25–38) | 42 (35–46) | 45 (45–46) | 0.0003 |
| CD4 T cell count, median (IQR) | 507 (372–825) | 485 (351–565) | 543 (434–758) | 295 (265–455) | 0.0759 |
| CD8 T cell count, median (IQR) | 748 (569–1075) | 559 (430–653) | 822 (590–1206) | 751 (540–856) | 0.1768 |
| Years HIV+, median (IQR) | 6.0 (3.1–10.0) | 1.7 (1.0–4.0) | 7.1 (2.0–12.8) | 2.0 (1.5–7.0) | 0.0108 |
| Years on ART, median (IQR) | 4.6 (2.6–8) | 1.7 (1.1–2.2) | 5.3 (2.0–10.0) | 1.0 (0.9–6.6) | 0.0021 |
| On Tenofovir, n (%) | 33 (56.9) | 20 (100) | 20 (64.5) | 7 (77.8) | 0.0012 |
| Large EV count, log10 EVs/µL plasma, median (IQR) | 3.6 (3.4–3.8) | 3.7 (3.6–3.8) | 3.7 (3.4–4.1) | 4.2 (4.0–4.3) | 0.0006 |
| Small EV count, log10 EVs/µL plasma, median (IQR) | 4.0 (3.9–4.4) | 4.0 (3.8–4.1) | 3.9 (3.8–4.4) | 4.3 (3.8–4.4) | 0.1540 |
| Female Sex Workers (n = 26) | Men Who Have Sex with Men (n = 13) | Females from General Population (n = 20) | Males from General Population (n = 7) | p Value | |
|---|---|---|---|---|---|
| Age, median (IQR) | 35 (33–41) | 25 (25–29) | 40 (33–44) | 35 (25–49) | <0.0001 |
| CD4 T cell count, median (IQR) | 394 (266–550) | 395 (321–514) | 465 (176–703) | 208 (164–563) | 0.4819 |
| CD8 T cell count, median (IQR) | 883 (641–1013) | 770 (438–913) | 729 (543–1196) | 630 (480–1147) | 0.2414 |
| Viral copies/mL, median (IQR) | 2432 (90–38,126) | 5442 (98–19,481) | 3632 (72–34,053) | 1563 (186–35,481) | 0.6147 |
| Years HIV+, median (IQR) | 3.7 (2.0–10.8) | 0.8 (0.5–2.1) | 6.0 (3.3–13.0) | 1.7 (0.8–18.8) | 0.0053 |
| ART-treated, n (%) | 24 (92.3) | 9 (69.2) | 13 (65.0) | 5 (71.4) | 0.0869 |
| Years on ART, median (IQR) | 3.4 (2.0–7.2) | 1.0 (0.7–2.0) | 5.0 (2.4–7.5) | 0.7 (0.7–6.1) | 0.0385 |
| On Tenofovir, n (%) | 12 (50) | 9 (100) | 8 (61) | 3 (60) | 0.0908 |
| Large EV count, log10 EVs/µL plasma, median (IQR) | 3.8 (3.6–4.0) | 3.9 (3.8–4.1) | 4.1 (3.5–4.2) | 4.3 (4.1–4.5) | 0.0375 |
| Small EV count, log10 EVs/µL plasma, median (IQR) | 3.9 (3.8–4.4) | 3.9 (3.8–4.0) | 3.9 (3.8–4.5) | 4.2 (3.8–4.5) | 0.6392 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazié, W.W.; Boucher, J.; Goyer, B.; Kania, D.; Traoré, I.T.; Somé, D.Y.; Alary, M.; Gilbert, C. HIV Replication Increases the Mitochondrial DNA Content of Plasma Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 1924. https://doi.org/10.3390/ijms24031924
Bazié WW, Boucher J, Goyer B, Kania D, Traoré IT, Somé DY, Alary M, Gilbert C. HIV Replication Increases the Mitochondrial DNA Content of Plasma Extracellular Vesicles. International Journal of Molecular Sciences. 2023; 24(3):1924. https://doi.org/10.3390/ijms24031924
Chicago/Turabian StyleBazié, Wilfried Wenceslas, Julien Boucher, Benjamin Goyer, Dramane Kania, Isidore Tiandiogo Traoré, Diane Yirgnur Somé, Michel Alary, and Caroline Gilbert. 2023. "HIV Replication Increases the Mitochondrial DNA Content of Plasma Extracellular Vesicles" International Journal of Molecular Sciences 24, no. 3: 1924. https://doi.org/10.3390/ijms24031924