
Implication of Cellular Senescence in Osteoarthritis: A Study on Equine Synovial Fluid Mesenchymal Stromal Cells
 ,
,
Abstract
:1. Introduction
2. Results
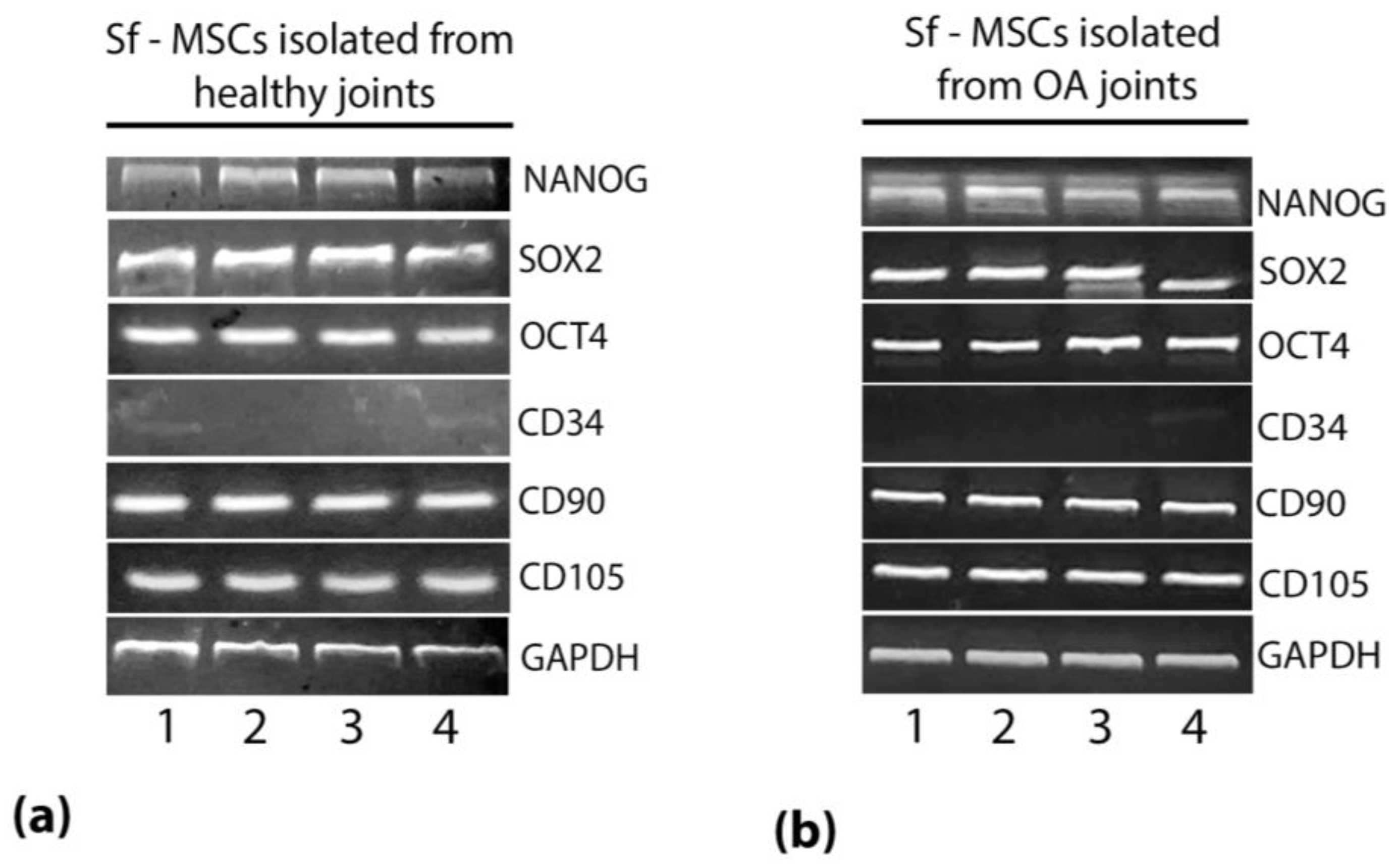
2.1. sf-MSCs Express Mesenchymal and Self-Renewal Markers
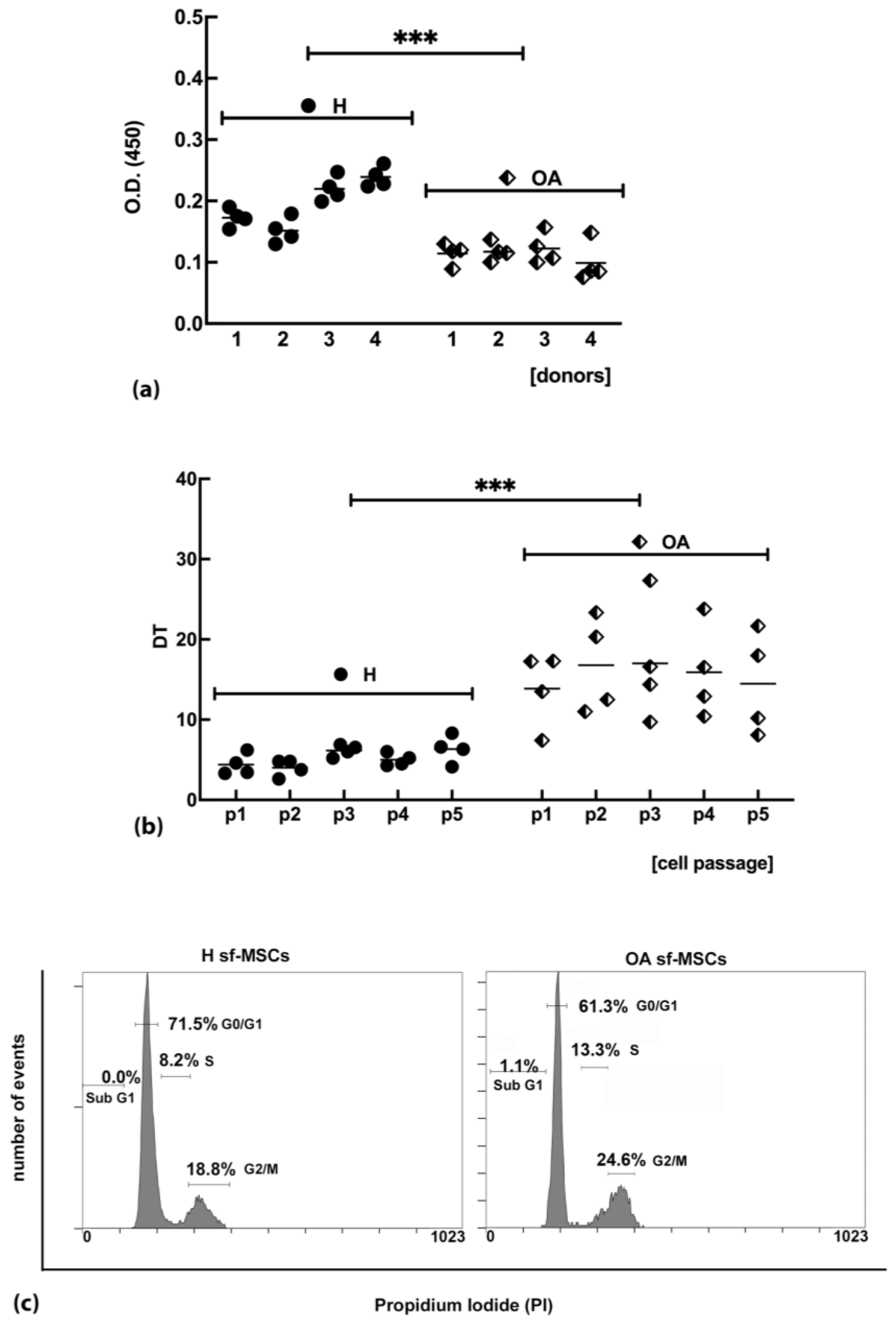
2.2. sf-MSCs Isolated from OA Joints Are Characterized by a Significant Reduction in the Ability to Proliferate Compared to sf-MSCs Isolated from Healthy Joints
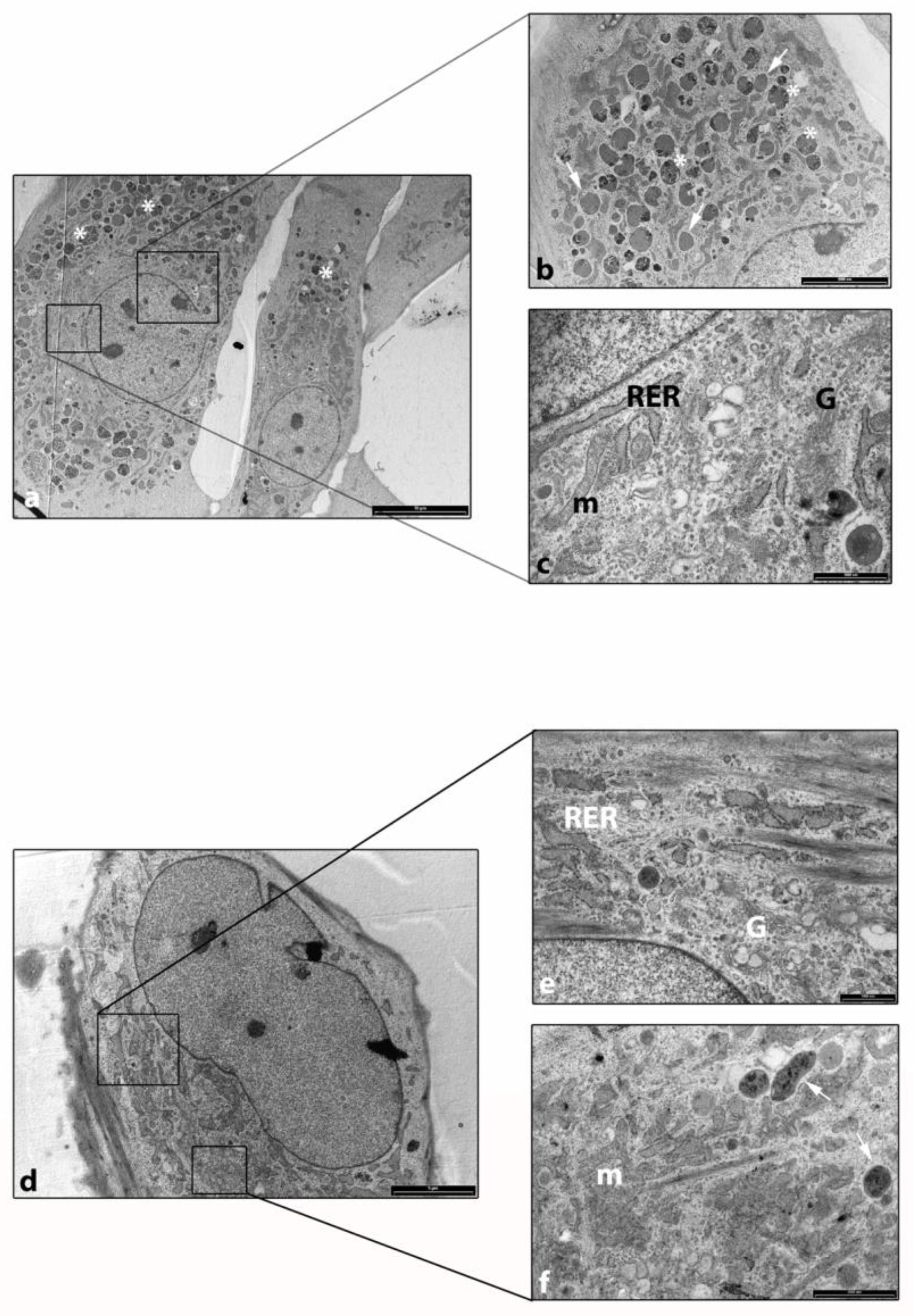
2.3. TEM Analysis Showed a High Activity in Cellular Digestion in H sf-MSCs Compared to OA sf-MSCs
2.4. OA sf-MSCs Showed Higher Intracellular ROS Levels Compared to H sf-MSCs
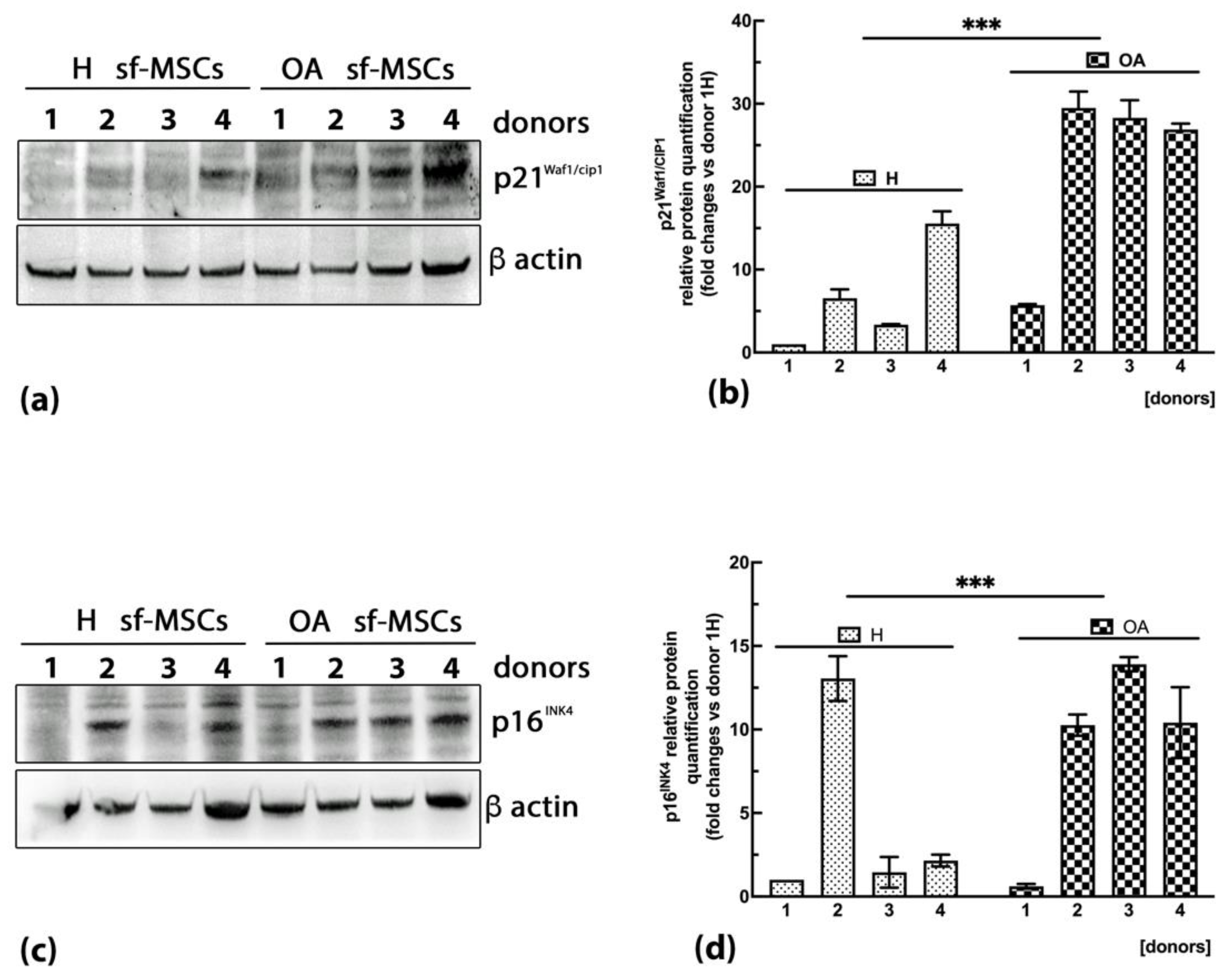
2.5. High Expression of the Senescent Markers Cyclin-Dependent Kinase Inhibitors p21WAF1/Cip1 and p16INK4A Is Detected in OA sf-MSCs
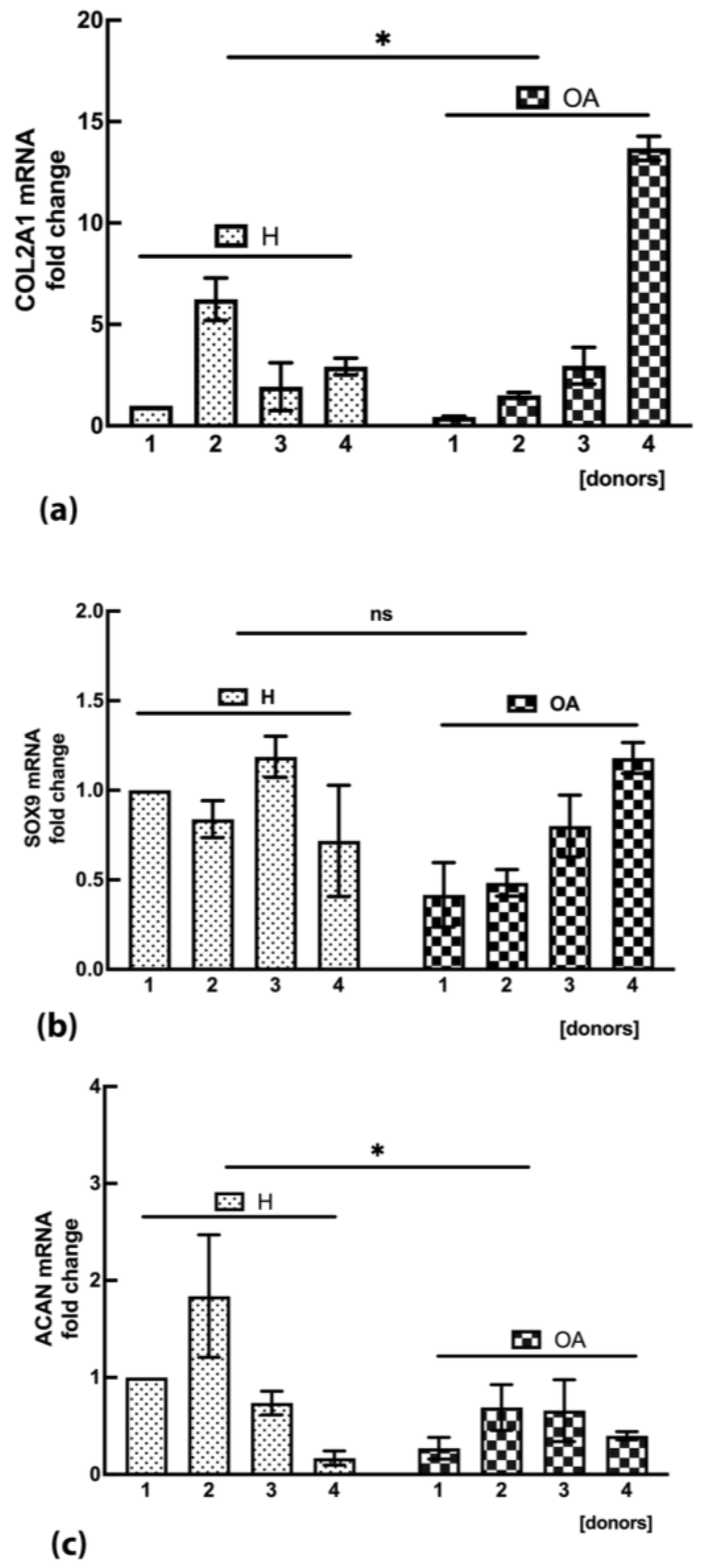
2.6. OA sf-MSCs Demonstrated a Variable Expression of COL2A1 mRNA
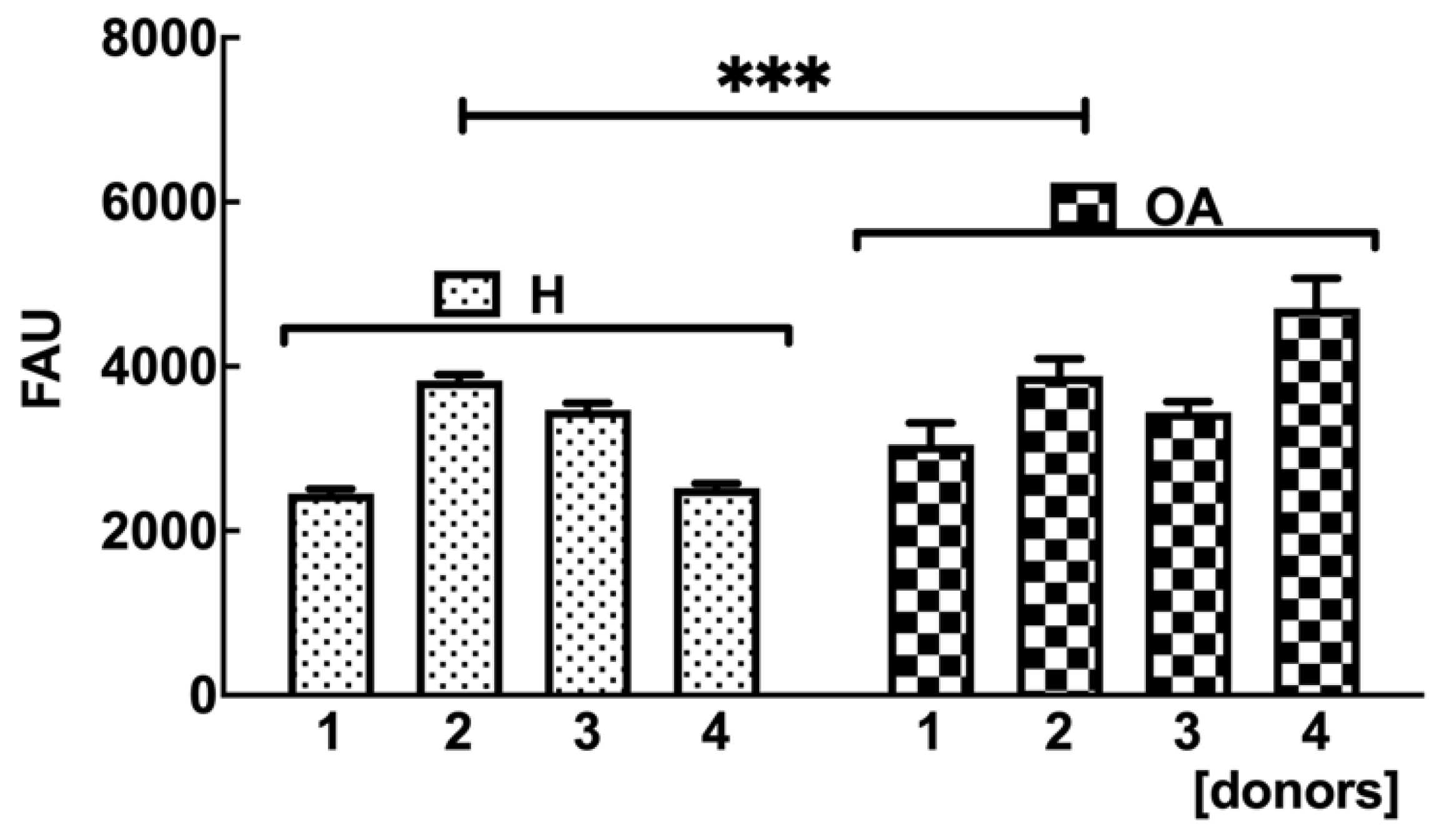
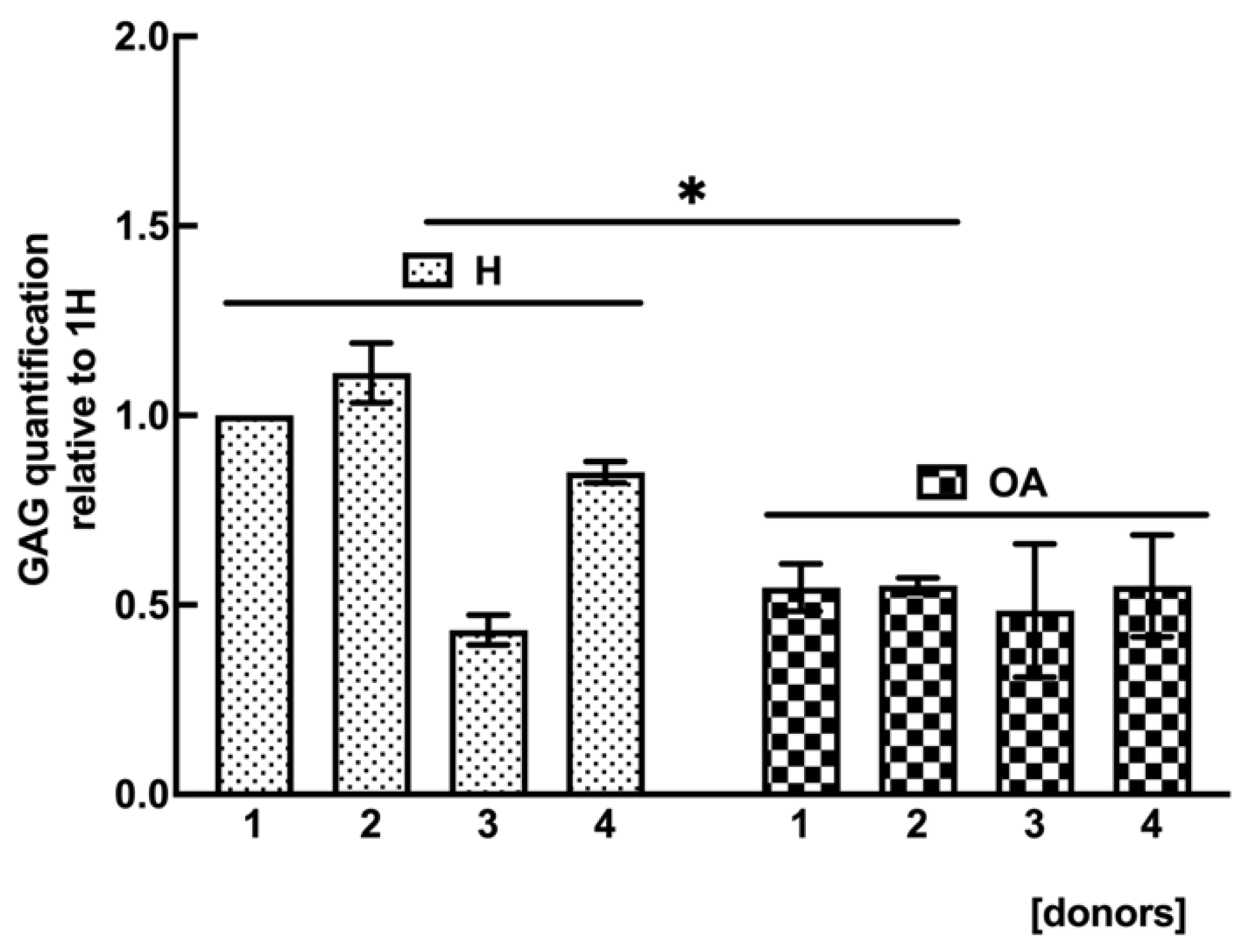
2.7. OA sf-MSCs Showed a Reduced GAG Synthesis
3. Discussion
4. Materials and Methods
4.1. Isolation of Equine sf-MSCs
4.2. mRNA Expression of Mesenchymal and Self-Renewal Markers by RT-PCR
4.3. BrdU Proliferation Assay
4.4. Doubling Time Assay
4.5. Cell Cycle Analysis
4.6. Reactive Oxygen Species (ROS) Detection Assay
4.7. Western Blot Analysis of Senescent Markers
4.8. Transmission Electron Microscopy (TEM)
4.9. Chondrogenic Differentiation
4.10. Real-Time PCR
4.11. GAG Quantification
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oh, J.; Lee, Y.D.; Wagers, A.J. Stem cell aging: Mechanisms, regulators and therapeutic opportunities. Nat. Med. 2014, 20, 870–880. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Mazzotti, E.; Teti, G.; Falconi, M.; Chiarini, F.; Barboni, B.; Mazzotti, A.; Muttini, A. Age-Related Alterations Affecting the Chondrogenic Differentiation of Synovial Fluid Mesenchymal Stromal Cells in an Equine Model. Cells 2019, 8, 1116. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Farr, J.N. Cellular senescence in age-related disorders. Transl. Res. 2020, 226, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Teti, G.; Chiarini, F.; Mazzotti, E.; Ruggeri, A.; Carano, F.; Falconi, M. Cellular senescence in vascular wall mesenchymal stromal cells, a possible contribution to the development of aortic aneurysm. Mech. Ageing Dev. 2021, 197, 111515. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, A.; O’Loghlen, A. Cellular Senescence and Ageing: Mechanisms and Interventions. Front. Aging 2022, 3, 866718. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Gorgoulis, V.; Adams, P.D.; Alimonti, A.; Bennett, D.C.; Bischof, O.; Bishop, C.; Campisi, J.; Collado, M.; Evangelou, K.; Ferbeyre, G.; et al. Cellular Senescence: Defining a Path Forward. Cell 2019, 179, 813–827. [Google Scholar] [CrossRef]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Kulkarni, P.; Martson, A.; Vidya, R.; Chitnavis, S.; Harsulkar, A. Pathophysiological landscape of osteoarthritis. Adv. Clin. Chem. 2021, 100, 37–90. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Coras, R.; Torres, A.; Lane, N.E.; Guma, M. Synovial inflammation in osteoarthritis progression. Nat. Rev. Rheumatol. 2022, 18, 258–275. [Google Scholar] [CrossRef]
- Jeon, O.H.; David, N.; Campisi, J.; Elisseeff, J.H. Senescent cells and osteoarthritis: A painful connection. J. Clin. Investig. 2018, 128, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, Y.; Sekiya, I.; Yagishita, K.; Muneta, T. Comparison of human stem cells derived from various mesenchymal tissues: Superiority of synovium as a cell source. Arthritis Rheum. 2005, 52, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, Y.; Muneta, T.; Tsuji, K.; Koga, H.; Sekiya, I. Mesenchymal stem cells in synovial fluid increase after meniscus injury. Clin. Orthop. Relat. Res. 2014, 472, 1357–1364, Erratum in: Clin. Orthop. Relat. Res. 2014, 472, 1049. [Google Scholar] [CrossRef] [PubMed]
- Morito, T.; Muneta, T.; Hara, K.; Ju, Y.J.; Mochizuki, T.; Makino, H.; Umezawa, A.; Sekiya, I. Synovial fluid-derived mesenchymal stem cells increase after intra-articular ligament injury in humans. Rheumatology 2008, 47, 1137–1143. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; He, H.; Xiang, W.; He, C. Human adipose and synovial mesenchymal stem cells improve osteoarthritis in rats by reducing chondrocyte reactive oxygen species and inhibiting inflammatory response. J. Clin. Lab. Anal. 2022, 36, e24353. [Google Scholar] [CrossRef]
- Neybecker, P.; Henrionnet, C.; Pape, E.; Mainard, D.; Galois, L.; Loeuille, D.; Gillet, P.; Pinzano, A. In vitro and in vivo potentialities for cartilage repair from human advanced knee osteoarthritis synovial fluid-derived mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 329. [Google Scholar] [CrossRef]
- Malaise, O.; Tachikart, Y.; Constantinides, M.; Mumme, M.; Ferreira-Lopez, R.; Noack, S.; Krettek, C.; Noël, D.; Wang, J.; Jorgensen, C.; et al. Mesenchymal stem cell senescence alleviates their intrinsic and seno-suppressive paracrine properties contributing to osteoarthritis development. Aging 2019, 11, 9128–9146. [Google Scholar] [CrossRef]
- Coryell, P.R.; Diekman, B.O.; Loeser, R.F. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat. Rev. Rheumatol. 2021, 17, 47–57. [Google Scholar] [CrossRef]
- Gonzalez, R.; Maki, C.B.; Pacchiarotti, J.; Csontos, S.; Pham, J.K.; Slepko, N.; Patel, A.; Silva, F. Pluripotent marker expression and differentiation of human second trimester Mesenchymal Stem Cells. Biochem. Biophys. Res. Commun. 2007, 362, 491–497. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Brondello, J.M.; Pers, Y.M. Taking in consideration the bystander effects of articular senescence. Ann. Transl. Med. 2019, 7, S386. [Google Scholar] [CrossRef]
- Astrike-Davis, E.M.; Coryell, P.; Loeser, R.F. Targeting cellular senescence as a novel treatment for osteoarthritis. Curr. Opin. Pharmacol. 2022, 64, 102213. [Google Scholar] [CrossRef] [PubMed]
- Ismail, O.M.; Said, U.N.; El-Omar, O.M. Adult Stem Cells for Cartilage Regeneration. Cureus 2022, 14, e32280. [Google Scholar] [CrossRef] [PubMed]
- Vinod, E.; Parameswaran, R.; Amirtham, S.M.; Rebekah, G.; Kachroo, U. Comparative analysis of human bone marrow mesenchymal stem cells, articular cartilage derived chondroprogenitors and chondrocytes to determine cell superiority for cartilage regeneration. Acta Histochem. 2021, 123, 151713. [Google Scholar] [CrossRef]
- Armiento, A.R.; Ladner, Y.D.; Della Bella, E.; Stoddart, M.J. Isolation and In Vitro Chondrogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stromal Cells. Methods Mol. Biol. 2023, 2598, 65–73. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, X.; Cao, X.; Qin, A.; Zhao, J. Current applications of adipose-derived mesenchymal stem cells in bone repair and regeneration: A review of cell experiments, animal models, and clinical trials. Front. Bioeng. Biotechnol. 2022, 10, 942128. [Google Scholar] [CrossRef]
- Kulus, M.; Sibiak, R.; Stefańska, K.; Zdun, M.; Wieczorkiewicz, M.; Piotrowska-Kempisty, H.; Jaśkowski, J.M.; Bukowska, D.; Ratajczak, K.; Zabel, M.; et al. Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues-Origins, Characteristics, Signaling Pathways, and Clinical Trials. Cells 2021, 10, 3278. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, S.A.; Song, J.S.; Shetty, A.A.; Kim, B.H.; Kim, S.J. Cartilage Regeneration Using Human Umbilical Cord Blood Derived Mesenchymal Stem Cells: A Systematic Review and Meta-Analysis. Medicina 2022, 58, 1801. [Google Scholar] [CrossRef]
- Song, Y.; Jorgensen, C. Mesenchymal Stromal Cells in Osteoarthritis: Evidence for Structural Benefit and Cartilage Repair. Biomedicines 2022, 10, 1278. [Google Scholar] [CrossRef]
- Poole, A.R.; Rosenberg, L.C.; Reiner, A.; Ionescu, M.; Bogoch, E.; Roughley, P.J. Contents and distributions of the proteoglycans decorin and biglycan in normal and osteoarthritic human articular cartilage. J. Orthop. Res. 1996, 14, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Olivier, A.; Nurton, J.P.; Guthrie, A.J. An epizoological study of wastage in thoroughbred racehorses in Gauteng, South Africa. J. S. Afr. Vet. Assoc. 1997, 68, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Hawkins, D.L. Training failure among yearling horses. Am. J. Vet. Res. 2001, 62, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.A.; Crawford, A.; English, A.; Henshaw, K.; Mundy, J.; Corscadden, D.; Chapman, T.; Emery, P.; Hatton, P.; McGonagle, D. Synovial fluid mesenchymal stem cells in health and early osteoarthritis: Detection and functional evaluation at the single-cell level. Arthritis Rheum. 2008, 58, 1731–1740. [Google Scholar] [CrossRef]
- de Sousa, E.B.; Casado, P.L.; Moura Neto, V.; Duarte, M.E.; Aguiar, D.P. Synovial fluid and synovial membrane mesenchymal stem cells: Latest discoveries and therapeutic perspectives. Stem Cell Res. Ther. 2014, 5, 112. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Sun, Z.; Chen, X.; Han, B.; Vangsness, C.T., Jr. Synovial fluid mesenchymal Stem Cells for knee arthritis and cartilage defects: A review of the literature. J. Knee Surg. 2021, 34, 1476–1485. [Google Scholar] [CrossRef]
- Zhang, L.; Pitcher, L.E.; Yousefzadeh, M.J.; Niedernhofer, L.J.; Robbins, P.D.; Zhu, Y. Cellular senescence: A key therapeutic target in aging and diseases. J. Clin. Investig. 2022, 132, e158450. [Google Scholar] [CrossRef]
- Varela-Eirín, M.; Carpintero-Fernández, P.; Guitián-Caamaño, A.; Varela-Vázquez, A.; García-Yuste, A.; Sánchez-Temprano, A.; Bravo-López, S.B.; Yañez-Cabanas, J.; Fonseca, E.; Largo, R.; et al. Extracellular vesicles enriched in connexin 43 promote a senescent phenotype in bone and synovial cells contributing to osteoarthritis progression. Cell Death Dis. 2022, 13, 681. [Google Scholar] [CrossRef]
- Chandler, H.; Peters, G. Stressing the cell cycle in senescence and aging. Curr. Opin. Cell Biol. 2013, 25, 765–771. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Chang, N.C. Autophagy and Stem Cells: Self-Eating for Self-Renewal. Front. Cell Dev. Biol. 2020, 8, 138. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Alzahrani, A.M.; Hanieh, H.N.; Kumar, S.A.; Ben Ammar, R.; Rengarajan, T.; Alhoot, M.A. Autophagy and senescence: A new insight in selected human diseases. J. Cell Physiol. 2019, 234, 21485–21492. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, S.; Das, S.K.; Emdad, L.; Fisher, P.B. Autophagy and senescence: Insights from normal and cancer stem cells. Adv. Cancer Res. 2021, 150, 147–208. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.T.; Lee, K.B.; Kim, S.Y.; Choi, H.R.; Park, S.C. Autophagy impairment induces premature senescence in primary human fibroblasts. PLoS ONE 2011, 6, e23367. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, W.; Tan, H.Y.; Lu, Y.; Li, Z.; Qu, Y.; Wang, N.; Wang, D. Role of Autophagy in the Maintenance of Stemness in Adult Stem Cells: A Disease-Relevant Mechanism of Action. Front. Cell Dev. Biol. 2021, 9, 715200. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Wu, S.B.; Wu, Y.T.; Wei, Y.H. Oxidative stress response elicited by mitochondrial dysfunction: Implication in the pathophysiology of aging. Exp. Biol. Med. 2013, 238, 450–460. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- De Gaetano, A.; Gibellini, L.; Zanini, G.; Nasi, M.; Cossarizza, A.; Pinti, M. Mitophagy and Oxidative Stress: The Role of Aging. Antioxidants 2021, 10, 794. [Google Scholar] [CrossRef]
- Liu, L.; Luo, P.; Yang, M.; Wang, J.; Hou, W.; Xu, P. The role of oxidative stress in the development of knee osteoarthritis: A comprehensive research review. Front. Mol. Biosci. 2022, 9, 1001212. [Google Scholar] [CrossRef]
- Greenwald, R.A. Oxygen radicals, inflammation, and arthritis: Pathophysiological considerations and implications for treatment. Semin. Arthritis Rheum. 1991, 20, 219–240. [Google Scholar] [CrossRef]
- Mendes, A.F.; Caramona, M.M.; Carvalho, A.P.; Lopes, M.C. Differential roles of hydrogen peroxide and superoxide in mediating IL-1-induced NF-kappa B activation and iNOS expression in bovine articular chondrocytes. J. Cell Biochem. 2003, 88, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Aggarwal, A.; Aggarwal, A.; Bhattacharyya, S.; Kumar, V.; Sharma, V.; Sahni, D. Senescent chondrogenic progenitor cells derived from articular cartilage of knee osteoarthritis patients contributes to senescence-associated secretory phenotype via release of IL-6 and IL-8. Acta Histochem. 2022, 124, 151867. [Google Scholar] [CrossRef] [PubMed]
- Diekman, B.O.; Sessions, G.A.; Collins, J.A.; Knecht, A.K.; Strum, S.L.; Mitin, N.K.; Carlson, C.S.; Loeser, R.F.; Sharpless, N.E. Expression of p16INK4a is a biomarker of chondrocyte aging but does not cause osteoarthritis. Aging Cell 2018, 17, e12771. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V.; Dvir-Ginzberg, M. SOX9 and the many facets of its regulation in the chondrocyte lineage. Connect. Tissue Res. 2017, 58, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Rim, Y.A.; Ju, J.H. The Role of Fibrosis in Osteoarthritis Progression. Life 2020, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Bolia, I.K.; Mertz, K.; Faye, E.; Sheppard, J.; Telang, S.; Bogdanov, J.; Hasan, L.K.; Haratian, A.; Evseenko, D.; Weber, A.E.; et al. Cross-Communication Between Knee Osteoarthritis and Fibrosis: Molecular Pathways and Key Molecules. Open Access J. Sports Med. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Rainaldi, G.; Pinto, B.; Piras, A.; Vatteroni, L.; Simi, S.; Citti, L. Reduction of proliferative heterogeneity of CHEF18 Chinese hamster cell line during the progression toward tumorigenicity. In Vitro Cell. Dev. Biol. 1991, 27, 949–952. [Google Scholar] [CrossRef]
- Lonetti, A.; Cappellini, A.; Bertaina, A.; Locatelli, F.; Pession, A.; Buontempo, F.; Evangelisti, C.; Orsini, E.; Zambonin, L.; Neri, L.M.; et al. Improving Nelarabine Efficacy in T Cell Acute Lymphoblastic Leukemia by Targeting Aberrant Pi3k/Akt/Mtor Signaling Pathway. J. Hematol. Oncol. 2016, 9, 114. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H Group | G0/G1(%) | S Phase (%) | G2/M (%) |
| 1H | 82.5 ± 6.4 | 4.4 ± 2.4 | 12.8 ± 5.7 |
| 2H | 65.5 ± 1.7 * | 3.7 ± 1.4 | 15.5 ± 1.3 |
| 3H | 75.5 ± 5.7 | 7.8 ± 0.4 * | 15.2 ± 5.0 |
| 4H | 59.7 ± 6.4 * | 13.9 ± 6.3 * | 24.3 ± 9.3 * |
| OA Group | G0/G1(%) | S Phase (%) | G2/M (%) |
| 1 OA | 63.7 ± 3.4 * | 10.4 ± 4.0 * | 25.1 ± 0.7 * |
| 2 OA | 62.6 ± 1.6 * | 17,8 ± 3.4 * | 17.1 ± 2.3 * |
| 3 OA | 52.9 ± 9.7 * | 13.5 ± 3.4 * | 32.6 ± 13.2 * |
| 4 OA | 56.1 ± 1.4 * | 24.0 ± 2.9 * | 16.9 ± 0.7 * |
| Group H | Breed | Age (Years) | Gender |
| 1 H | Thoroughbred | 3 | M |
| 2 H | Standardbred | 6 | F |
| 3 H | Standardbred | 10 | F |
| 4 H | Standardbred | 14 | F |
| Group OA | Breed | Age (Years) | Gender |
| 1 OA | Thoroughbred | 3 | M |
| 2 OA | Thoroughbred | 6 | F |
| 3 OA | Standardbred | 8 | M |
| 4 OA | Pleasure riding horse | 10 | F |
| Name | Forward | Reverse | bp |
|---|---|---|---|
| CD90 | 5′-ATGAGAATACCACCGCCACA-3′ | 5′-AGTTTGTCTCGGAGCACAGA-3′ | 262 |
| CD105 | 5′-TCAGGTCCCCAACACTAACC-3′ | 5′-AGTCTTGTTCGTGCTGAGGA-3′ | 148 |
| CD34 | 5′-CCTTGCCCAGTCTGAGGTTA-3′ | 5′-GTCTTGCGGGAATAGTGCTG-3′ | 172 |
| NANOG | 5′-TCTCTCCTCTGCCTTCCTCC -3′ | 5′-TCTGCTGGAGGCTGAGGTAT-3′ | 225 |
| OCT4 | 5′-GGTACGAGTGTGGTTCTGCA-3′ | 5′-ACCGAGGAGTACAGCGTAGT-3′ | 192 |
| SOX2 | 5′-GCCCTGCAGTACAACTCCAT-3′ | 5′-GACTTGACCACCGAACCCAT-3′ | 128 |
| GAPDH | 5′-TGCCCCAATGTTTGTGATGG-3′ | 5′-CACTGTGGTCATGAGTCCCT-3′ | 154 |
| SOX9 | 5′-GAACAGCCCGTCTACACACA-3′ | 5′-GCCACTGATTCGCAACAAGG-3′ | 235 |
| COL2A1 | 5′-CTGGCAAGCAAGGAGACAGA-3′ | 5′-CCATTAGCGCCATCTTTGCC-3′ | 292 |
| ACAN | 5′-TCATCTAGAGCCCACTGCCT-3′ | 5′-AGTCCACCGAGGTCCTCTAC-3′ | 234 |
| GAPDH | 5′-TGCCCCAATGTTTGTGATGG-3′ | 5′-CACTGTGGTCATGAGTCCCT-3′ | 154 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teti, G.; Mazzotti, E.; Gatta, V.; Chiarini, F.; Alfieri, M.L.; Falconi, M. Implication of Cellular Senescence in Osteoarthritis: A Study on Equine Synovial Fluid Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2023, 24, 3109. https://doi.org/10.3390/ijms24043109
Teti G, Mazzotti E, Gatta V, Chiarini F, Alfieri ML, Falconi M. Implication of Cellular Senescence in Osteoarthritis: A Study on Equine Synovial Fluid Mesenchymal Stromal Cells. International Journal of Molecular Sciences. 2023; 24(4):3109. https://doi.org/10.3390/ijms24043109
Chicago/Turabian StyleTeti, Gabriella, Eleonora Mazzotti, Valentina Gatta, Francesca Chiarini, Maria Laura Alfieri, and Mirella Falconi. 2023. "Implication of Cellular Senescence in Osteoarthritis: A Study on Equine Synovial Fluid Mesenchymal Stromal Cells" International Journal of Molecular Sciences 24, no. 4: 3109. https://doi.org/10.3390/ijms24043109