
Direct Enhancement Effect of Hippocampal Cholinergic Neurostimulating Peptide on Cholinergic Activity in the Hippocampus
 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
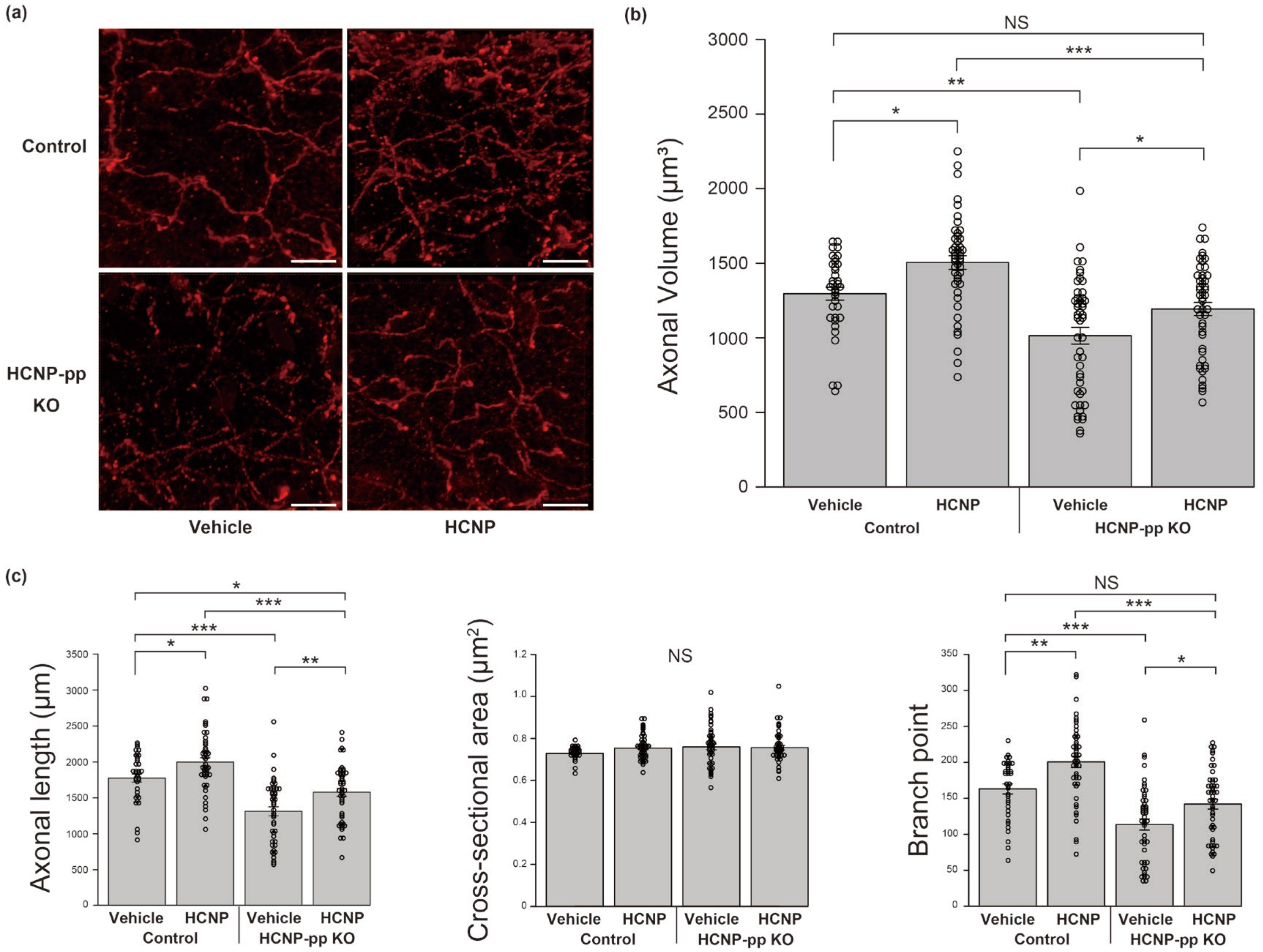
2.1. HCNP Administration Recovers ChAT-Positive Axons in HCNP-pp cKO Mice
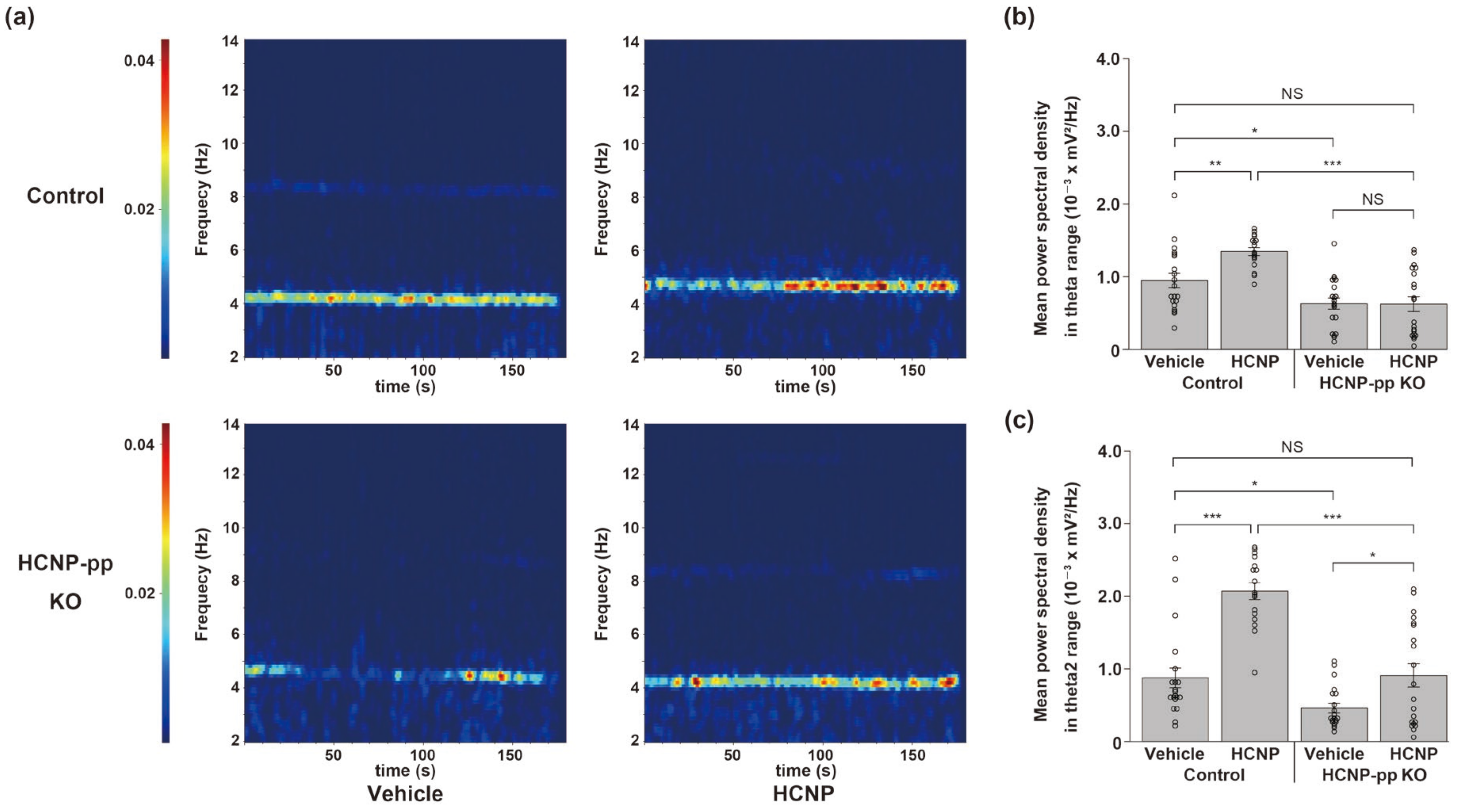
2.2. HCNP Administration Recovers Hippocampal Theta Power
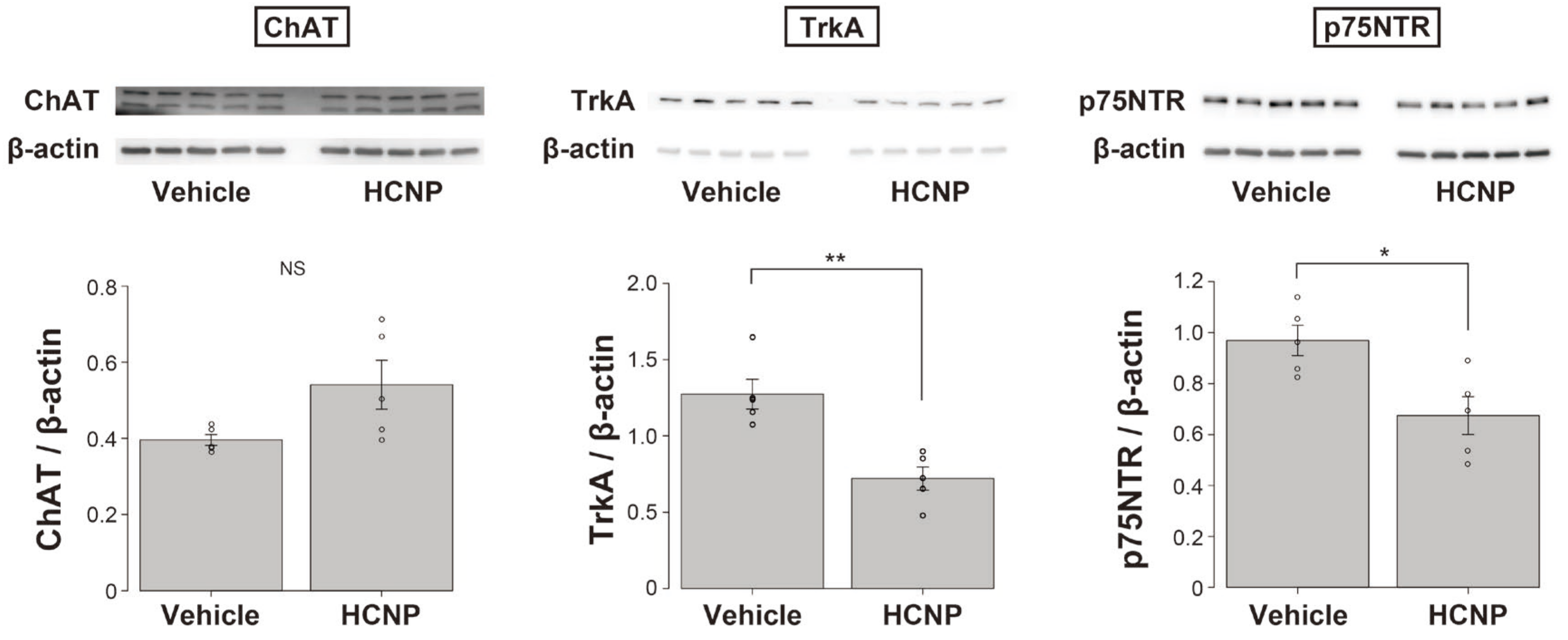
2.3. HCNP Administration Decreases the Abundance of Nerve Growth Factor (NGF) Receptors, TrkA, and p75NTR
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. HCNP Administration
4.3. Quantification of Hippocampal Cholinergic Axons
4.4. Electrophysiological Recordings
4.5. Western Blot Analyses
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benke, T.A.; Lüthi, A.; Isaac, J.T.; Collingridge, G.L. Modulation of AMPA receptor unitary conductance by synaptic activity. Nature 1998, 393, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Sekeres, M.J.; Winocur, G.; Moscovitch, M. The hippocampus and related neocortical structures in memory transformation. Neurosci. Lett. 2018, 680, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Yakel, J.L. Timing-dependent septal cholinergic induction of dynamic hippocampal synaptic plasticity. Neuron 2011, 71, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.L. Rhythms of the hippocampal network. Nat. Rev. Neurosci. 2016, 17, 239–249. [Google Scholar] [CrossRef]
- Benchenane, K.; Peyrache, A.; Khamassi, M.; Tierney, P.L.; Gioanni, Y.; Battaglia, F.P.; Wiener, S.I. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon learning. Neuron 2010, 66, 921–936. [Google Scholar] [CrossRef]
- Peyrache, A.; Khamassi, M.; Benchenane, K.; Wiener, S.I.; Battaglia, F.P. Replay of rule-learning related neural patterns in the prefrontal cortex during sleep. Nat. Neurosci. 2009, 12, 919–926. [Google Scholar] [CrossRef]
- Sirota, A.; Montgomery, S.; Fujisawa, S.; Isomura, Y.; Zugaro, M.; Buzsáki, G. Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron 2008, 60, 683–697. [Google Scholar] [CrossRef]
- Green, J.D.; Arduini, A.A. Hippocampal electrical activity in arousal. J. Neurophysiol. 1954, 17, 533–557. [Google Scholar] [CrossRef]
- Wang, X.J. Pacemaker neurons for the theta rhythm and their synchronization in the septohippocampal reciprocal loop. J. Neurophysiol. 2002, 87, 889–900. [Google Scholar] [CrossRef]
- Dannenberg, H.; Pabst, M.; Braganza, O.; Schoch, S.; Niediek, J.; Bayraktar, M.; Mormann, F.; Beck, H. Synergy of direct and indirect cholinergic septo-hippocampal pathways coordinates firing in hippocampal networks. J. Neurosci. 2015, 35, 8394–8410. [Google Scholar] [CrossRef]
- Mamad, O.; McNamara, H.M.; Reilly, R.B.; Tsanov, M. Medial septum regulates the hippocampal spatial representation. Front. Behav. Neurosci. 2015, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Ojika, K.; Kojima, S.; Ueki, Y.; Fukushima, N.; Hayashi, K.; Yamamoto, M. Purification and structural analysis of hippocampal cholinergic neurostimulating peptide. Brain Res. 1992, 572, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Ojika, K.; Mitake, S.; Tohdoh, N.; Appel, S.H.; Otsuka, Y.; Katada, E.; Matsukawa, N. Hippocampal cholinergic neurostimulating peptides (Hcnp). Prog. Neurobiol. 2000, 60, 37–83. [Google Scholar] [CrossRef] [PubMed]
- Madokoro, Y.; Yoshino, Y.; Kato, D.; Sato, T.; Mizuno, M.; Kanamori, T.; Shimazawa, M.; Hida, H.; Hara, H.; Yoshida, M.; et al. Reduced cholinergic activity in the hippocampus of hippocampal cholinergic neurostimulating peptide precursor protein knockout mice. Int. J. Mol. Sci. 2019, 20, 5367. [Google Scholar] [CrossRef]
- Kondo-Takuma, Y.; Mizuno, M.; Tsuda, Y.; Madokoro, Y.; Suzuki, K.; Sato, T.; Takase, H.; Uchida, Y.; Adachi, K.I.; Hida, H.; et al. Reduction of acetylcholine in the hippocampus of hippocampal cholinergic neurostimulating peptide precursor protein knockout mice. Sci. Rep. 2021, 11, 22072. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Ohi, Y.; Sato, T.; Tsuda, Y.; Madokoro, Y.; Mizuno, M.; Adachi, K.; Uchida, Y.; Haji, A.; Ojika, K.; et al. Reduction of glutamatergic activity through cholinergic dysfunction in the hippocampus of hippocampal cholinergic neurostimulating peptide precursor protein knockout mice. Sci. Rep. 2022, 12, 19161. [Google Scholar] [CrossRef]
- Mikulovic, S.; Restrepo, C.E.; Siwani, S.; Bauer, P.; Pupe, S.; Tort, A.B.L.; Kullander, K.; Leão, R.N. Ventral hippocampal olm cells control type 2 theta oscillations and response to predator odor. Nat. Commun. 2018, 9, 3638. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, G.D.; Luca, A.; Rusu, R.N.; Bild, V.; Beschea Chiriac, S.I.; Solcan, C.; Bild, W.; Ababei, D.C. Alzheimer’s disease pharmacotherapy in relation to cholinergic system involvement. Biomolecules 2019, 10, 40. [Google Scholar] [CrossRef]
- Tuszynski, M.H.; Yang, J.H.; Barba, D.; Hoi-Sang, U.; Bakay, R.A.; Pay, M.M.; Masliah, E.; Conner, J.M.; Kobalka, P.; Roy, S.; et al. nerve growth factor gene therapy: Activation of neuronal responses in Alzheimer disease. JAMA Neurol. 2015, 72, 1139–1147. [Google Scholar] [CrossRef]
- Mufson, E.J.; Counts, S.E.; Perez, S.E.; Ginsberg, S.D. Cholinergic system during the progression of Alzheimer’s disease: Therapeutic implications. Expert Rev. Neurother. 2008, 8, 1703–1718. [Google Scholar] [CrossRef]
- Schliebs, R.; Arendt, T. The cholinergic system in aging and neuronal degeneration. Behav. Brain Res. 2011, 221, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Calissano, P. Nerve growth factor and Alzheimer’s disease: New facts for an old hypothesis. Mol. Neurobiol. 2012, 46, 588–604. [Google Scholar] [CrossRef]
- Triaca, V.; Calissano, P. Impairment of the nerve growth factor pathway driving amyloid accumulation in cholinergic neurons: The incipit of the Alzheimer’s disease story? Neural Regen. Res. 2016, 11, 1553–1556. [Google Scholar] [CrossRef] [PubMed]
- McDade, E.; Bateman, R.J. Stop Alzheimer’s before it starts. Nature 2017, 547, 153–155. [Google Scholar] [CrossRef]
- Ojika, K.; Mitake, S.; Kamiya, T.; Kosuge, N.; Taiji, M. Two different molecules, Ngf and Free-Hcnp, stimulate cholinergic activity in septal nuclei in vitro in a different manner. Brain Res. Dev. Brain Res. 1994, 79, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T.; Dean, R.L., III; Beer, B.; Lippa, A.S. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal forebrain cholinergic circuits and signaling in cognition and cognitive decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef]
- Whitehouse, P.J.; Struble, R.G.; Clark, A.W.; Price, D.L. Alzheimer disease: Plaques, tangles, and the basal forebrain. Ann. Neurol. 1982, 12, 494. [Google Scholar] [CrossRef]
- Grothe, M.; Heinsen, H.; Teipel, S.J. Atrophy of the cholinergic basal forebrain over the adult age range and in early stages of Alzheimer’s disease. Biol. Psychiatry 2012, 71, 805–813. [Google Scholar] [CrossRef]
- Grothe, M.; Zaborszky, L.; Atienza, M.; Gil-Neciga, E.; Rodriguez-Romero, R.; Teipel, S.J.; Amunts, K.; Suarez-Gonzalez, A.; Cantero, J.L. Reduction of basal forebrain cholinergic system parallels cognitive impairment in patients at high risk of developing Alzheimer’s disease. Cereb. Cortex 2010, 20, 1685–1695. [Google Scholar] [CrossRef]
- Teipel, S.J.; Meindl, T.; Grinberg, L.; Grothe, M.; Cantero, J.L.; Reiser, M.F.; Möller, H.J.; Heinsen, H.; Hampel, H. The cholinergic system in mild cognitive impairment and Alzheimer’s disease: An in vivo Mri and Dti study. Hum. Brain Mapp. 2011, 32, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, P.J.; Hedreen, J.C.; White, C.L., III; Price, D.L. Basal forebrain neurons in the dementia of Parkinson disease. An. Neurol. 1983, 13, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Perry, E.K.; Curtis, M.; Dick, D.J.; Candy, J.M.; Atack, J.R.; Bloxham, C.A.; Blessed, G.; Fairbairn, A.; Tomlinson, B.E.; Perry, R.H. Cholinergic correlates of cognitive impairment in Parkinson’s disease: Comparisons with Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 1985, 48, 413–421. [Google Scholar] [CrossRef]
- Aarsland, D.; Mosimann, U.P.; McKeith, I.G. Role of cholinesterase inhibitors in Parkinson’s disease and dementia with Lewy bodies. J. Geriatr. Psychiatry Neurol. 2004, 17, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Lippa, C.F.; Smith, T.W.; Perry, E. Dementia with Lewy bodies: Choline acetyltransferase parallels nucleus basalis pathology. J. Neural. Transm. 1999, 106, 525–535. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, L.M.; De Bastiani, M.A.; Rico, E.P.; Schonhofen, P.; Pfaffenseller, B.; Wollenhaupt-Aguiar, B.; Grun, L.; Barbé-Tuana, F.; Zimmer, E.R.; Castro, M.A.A.; et al. Cholinergic differentiation of human neuroblastoma Sh-Sy5y cell line and its potential use as an in vitro model for Alzheimer’s disease studies. Mol. Neurobiol. 2019, 56, 7355–7367. [Google Scholar] [CrossRef]
- Burke, R.M.; Norman, T.A.; Haydar, T.F.; Slack, B.E.; Leeman, S.E.; Blusztajn, J.K.; Mellott, T.J. Bmp9 ameliorates amyloidosis and the cholinergic defect in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2013, 110, 19567–19572. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Hellweg, R.; von Richthofen, S.; Anders, D.; Baethge, C.; Röpke, S.; Hartung, H.D.; Gericke, C.A. The time course of nerve growth factor content in different neuropsychiatric diseases–a unifying hypothesis. J. Neural. Transm. 1998, 105, 871–903. [Google Scholar] [CrossRef]
- Sharma, M.; Waghela, S.; Mhatre, R.; Saraogi, G.K. A recent update on intranasal delivery of high molecular weight proteins, peptides, and hormones. Cur. Pharm. Des. 2021, 27, 4279–4299. [Google Scholar] [CrossRef]
- Morishita, M.; Otsuka, Y.; Matsukawa, N.; Suzuki, H.; Nakazawa, H.; Maki, M.; Katou, H.; Ueda, R.; Ojika, K. Specific binding of 125i-hippocampal cholinergic neurostimulating peptide (Hcnp) to rat brain membranes: Characterization and regional distribution. Brain Res. 2003, 965, 194–202. [Google Scholar] [CrossRef]
- Hara, K.; Harris, R.A. The Anesthetic Mechanism of Urethane: The Effects on Neurotransmitter-Gated Ion Channels. Anesth. Analg. 2002, 94, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Colom, L.V.; García-Hernández, A.; Castañeda, M.T.; Perez-Cordova, M.G.; Garrido-Sanabria, E.R. Septo-Hippocampal Networks in Chronically Epileptic Rats: Potential Antiepileptic Effects of Theta Rhythm Generation. J. Neurophysiol. 2006, 95, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Bland, B.H.; Colom, L.V. Extrinsic and Intrinsic Properties Underlying Oscillation and Synchrony in Limbic Cortex. Prog. Neurobiol. 1993, 41, 157–208. [Google Scholar] [CrossRef]
- Cervera-Ferri, A.; Guerrero-Martínez, J.; Bataller-Mompeán, M.; Taberner-Cortes, A.; Martínez-Ricós, J.; Ruiz-Torner, A.; Teruel-Martí, V. Theta Synchronization between the Hippocampus and the Nucleus Incertus in Urethane-Anesthetized Rats. Exp. Brain Res. 2011, 211, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Vandecasteele, M.; Varga, V.; Berényi, A.; Papp, E.; Barthó, P.; Venance, L.; Freund, T.F.; Buzsáki, G. Optogenetic Activation of Septal Cholinergic Neurons Suppresses Sharp Wave Ripples and Enhances Theta Oscillations in the Hippocampus. Proc. Natl. Acad. Sci. USA 2014, 111, 13535–13540. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: New York, NY, USA, 2008; p. 376. [Google Scholar]
- Takata, N.; Yoshida, K.; Komaki, Y.; Xu, M.; Sakai, Y.; Hikishima, K.; Mimura, M.; Okano, H.; Tanaka, K.F. Optogenetic activation of Ca1 pyramidal neurons at the dorsal and ventral hippocampus evokes distinct brain-wide responses revealed by mouse Fmri. PLoS ONE 2015, 10, e0121417. [Google Scholar] [CrossRef]
- Aitken, P.; Zheng, Y.; Smith, P.F. The modulation of hippocampal theta rhythm by the vestibular system. J. Neurophysiol. 2018, 119, 548–562. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madokoro, Y.; Kato, D.; Tsuda, Y.; Arakawa, I.; Suzuki, K.; Sato, T.; Mizuno, M.; Uchida, Y.; Ojika, K.; Matsukawa, N. Direct Enhancement Effect of Hippocampal Cholinergic Neurostimulating Peptide on Cholinergic Activity in the Hippocampus. Int. J. Mol. Sci. 2023, 24, 8916. https://doi.org/10.3390/ijms24108916
Madokoro Y, Kato D, Tsuda Y, Arakawa I, Suzuki K, Sato T, Mizuno M, Uchida Y, Ojika K, Matsukawa N. Direct Enhancement Effect of Hippocampal Cholinergic Neurostimulating Peptide on Cholinergic Activity in the Hippocampus. International Journal of Molecular Sciences. 2023; 24(10):8916. https://doi.org/10.3390/ijms24108916
Chicago/Turabian StyleMadokoro, Yuta, Daisuke Kato, Yo Tsuda, Itsumi Arakawa, Kengo Suzuki, Toyohiro Sato, Masayuki Mizuno, Yuto Uchida, Kosei Ojika, and Noriyuki Matsukawa. 2023. "Direct Enhancement Effect of Hippocampal Cholinergic Neurostimulating Peptide on Cholinergic Activity in the Hippocampus" International Journal of Molecular Sciences 24, no. 10: 8916. https://doi.org/10.3390/ijms24108916