Chronic High-Dose Neonicotinoid Exposure Decreases Overwinter Survival of Apis mellifera L.

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
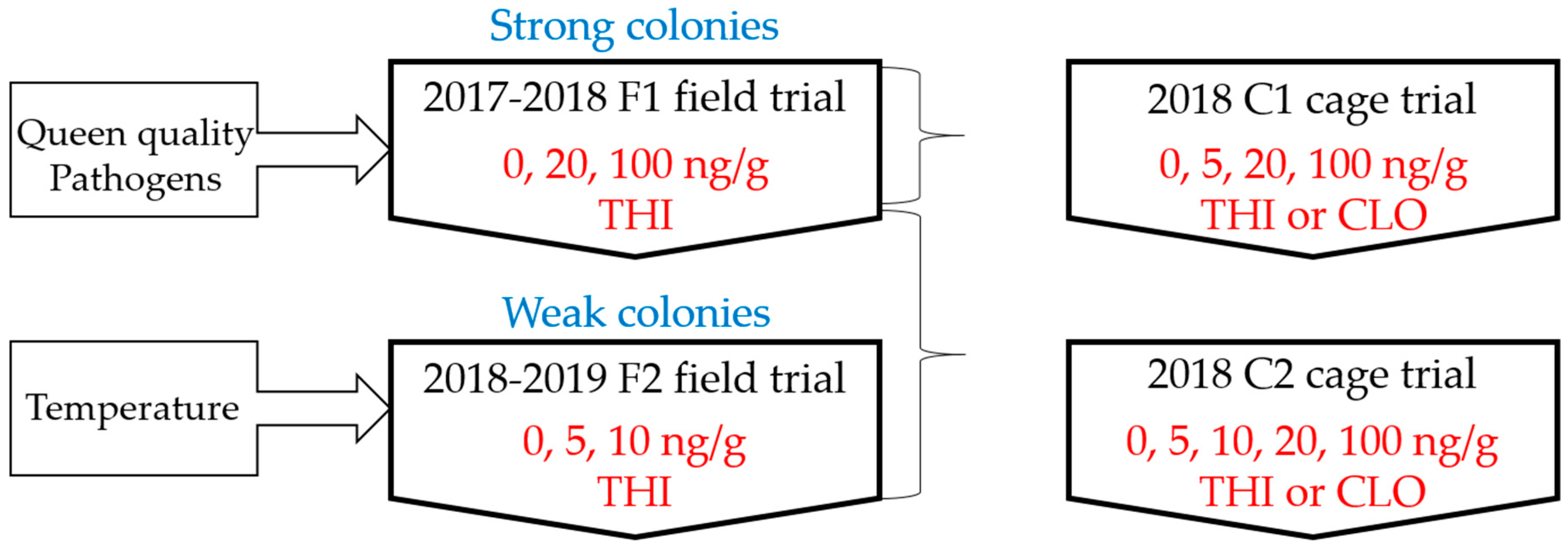
2.1. Experimental Design
2.2. Field Trials
2.2.1. Pathogen Monitoring in F1
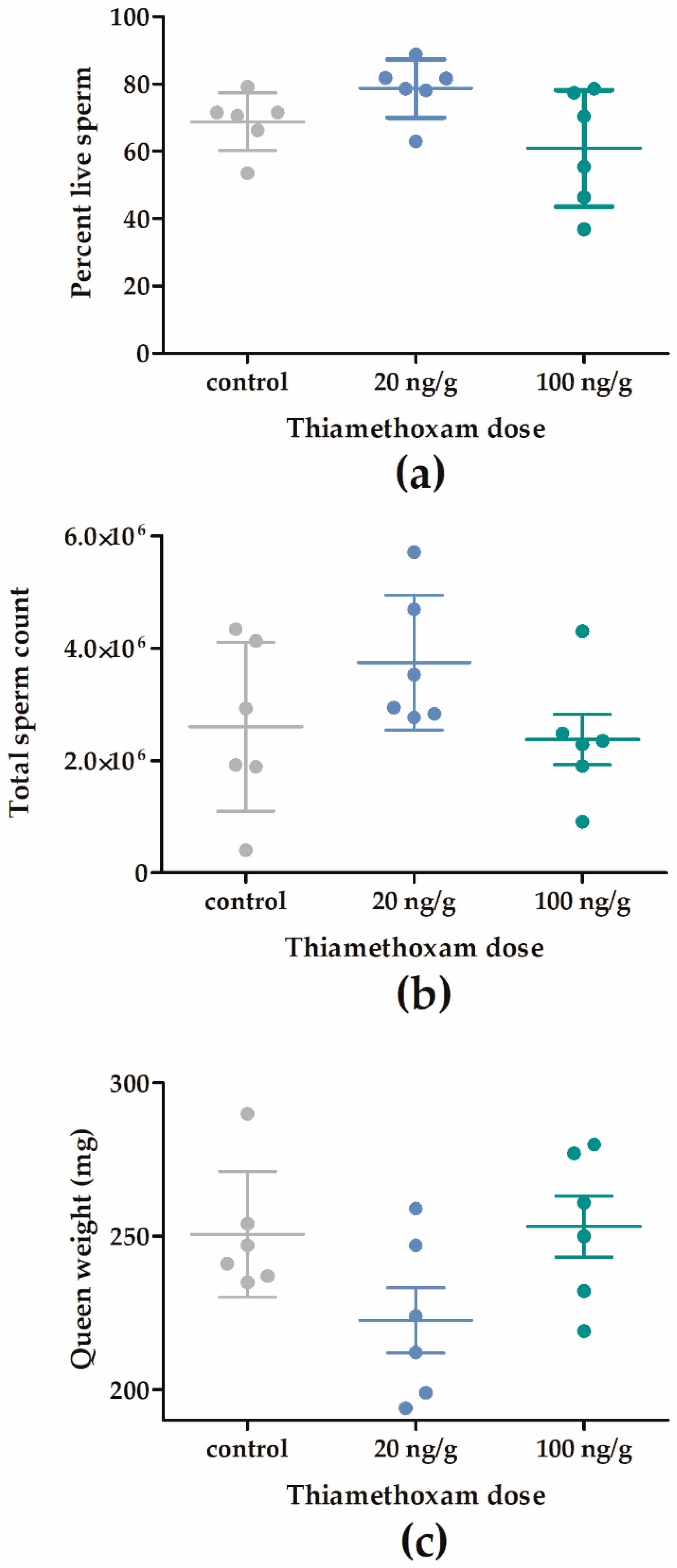
2.2.2. Queen Quality in F1
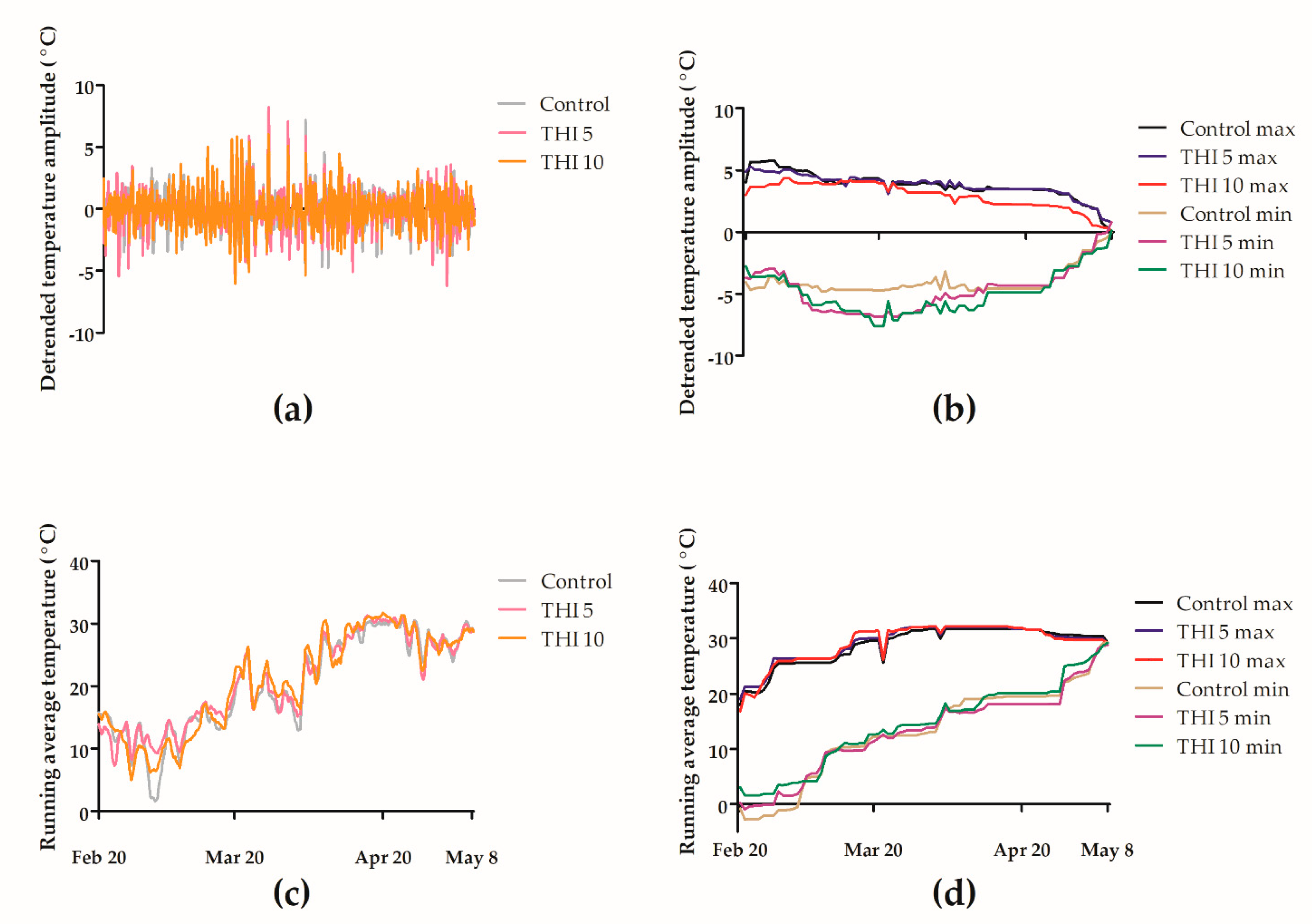
2.2.3. Temperature Monitoring in F2
2.2.4. Analysis of Data
2.3. Laboratory Cage Trials
2.4. Analysis of Data
3. Results
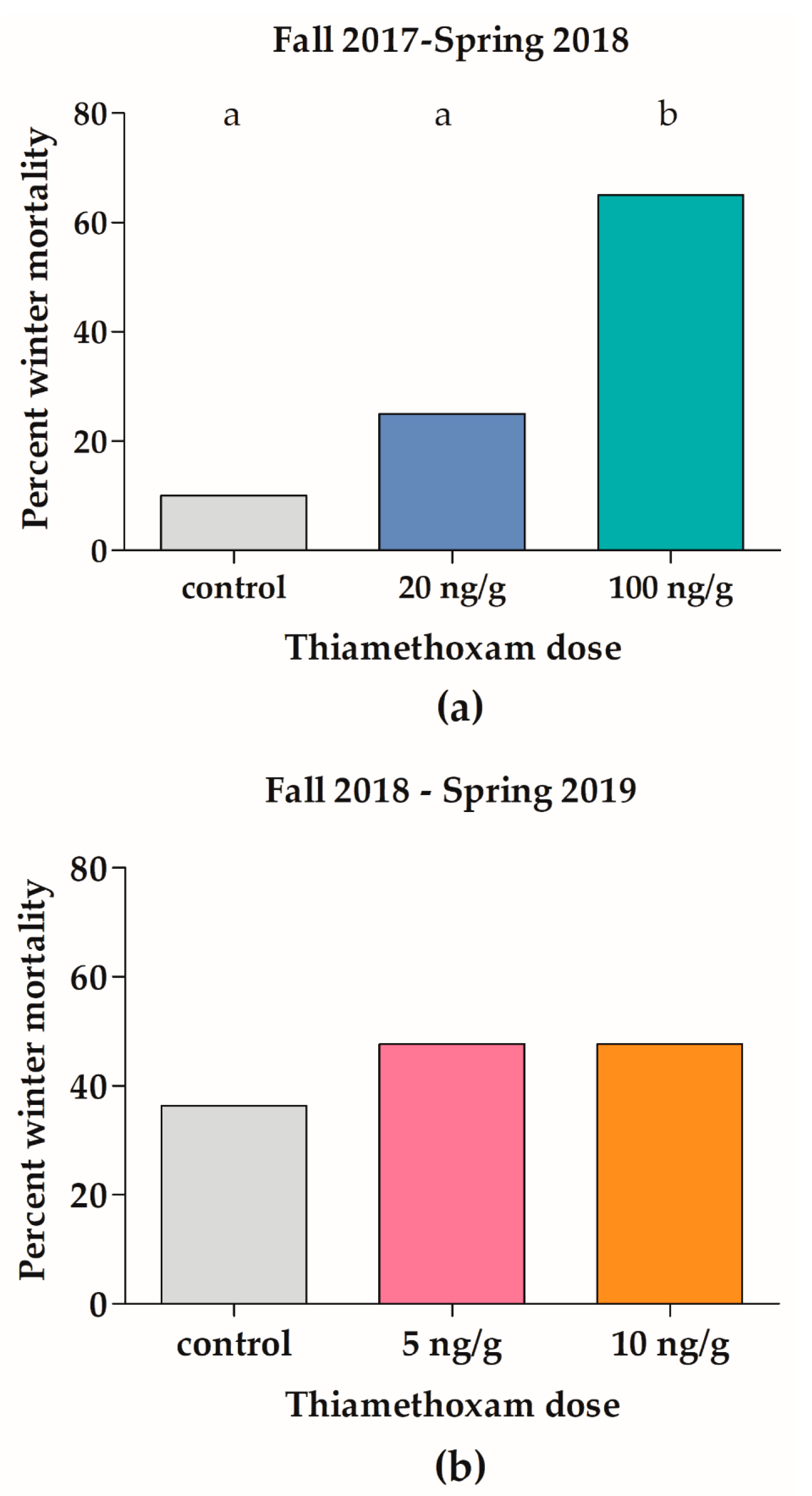
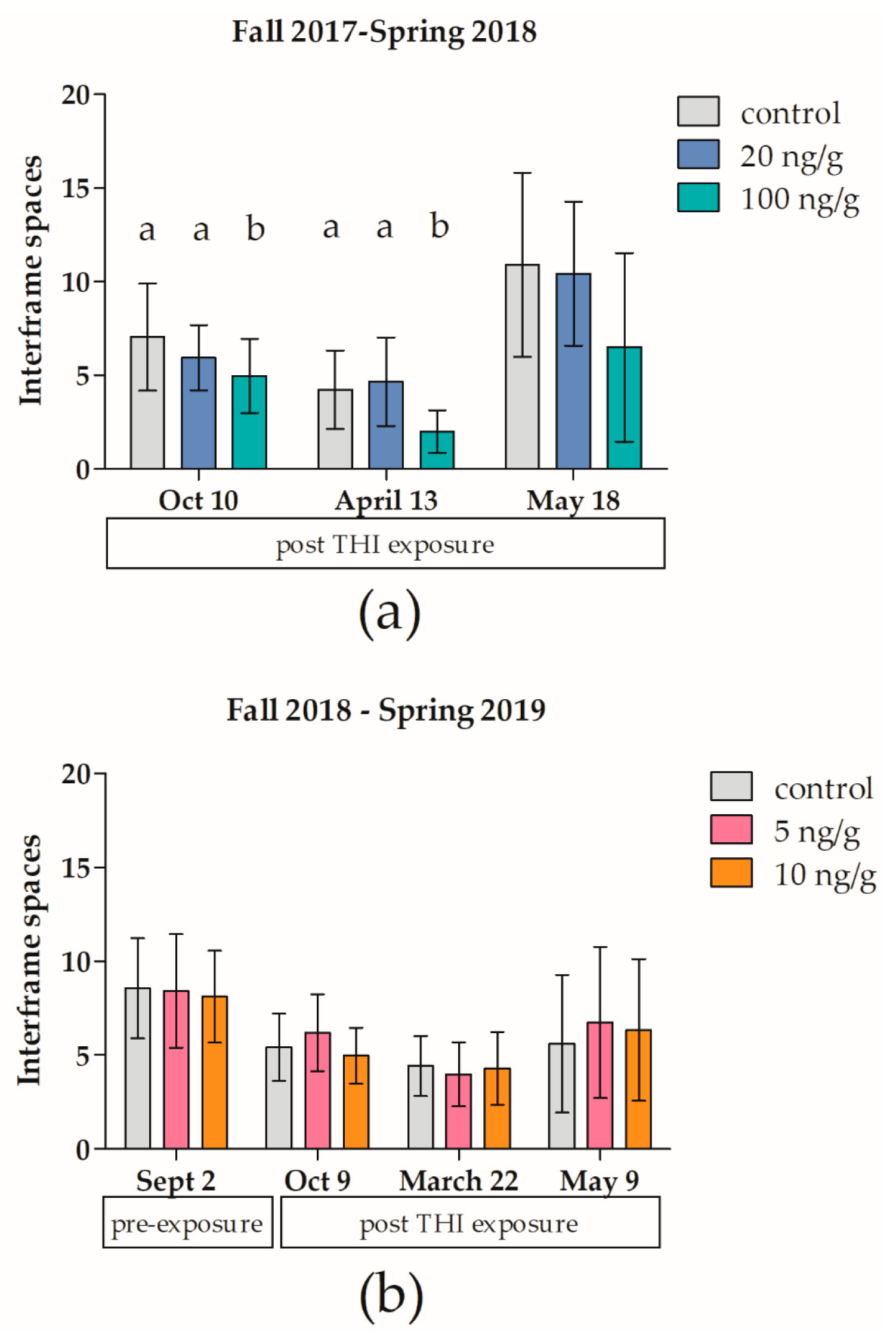
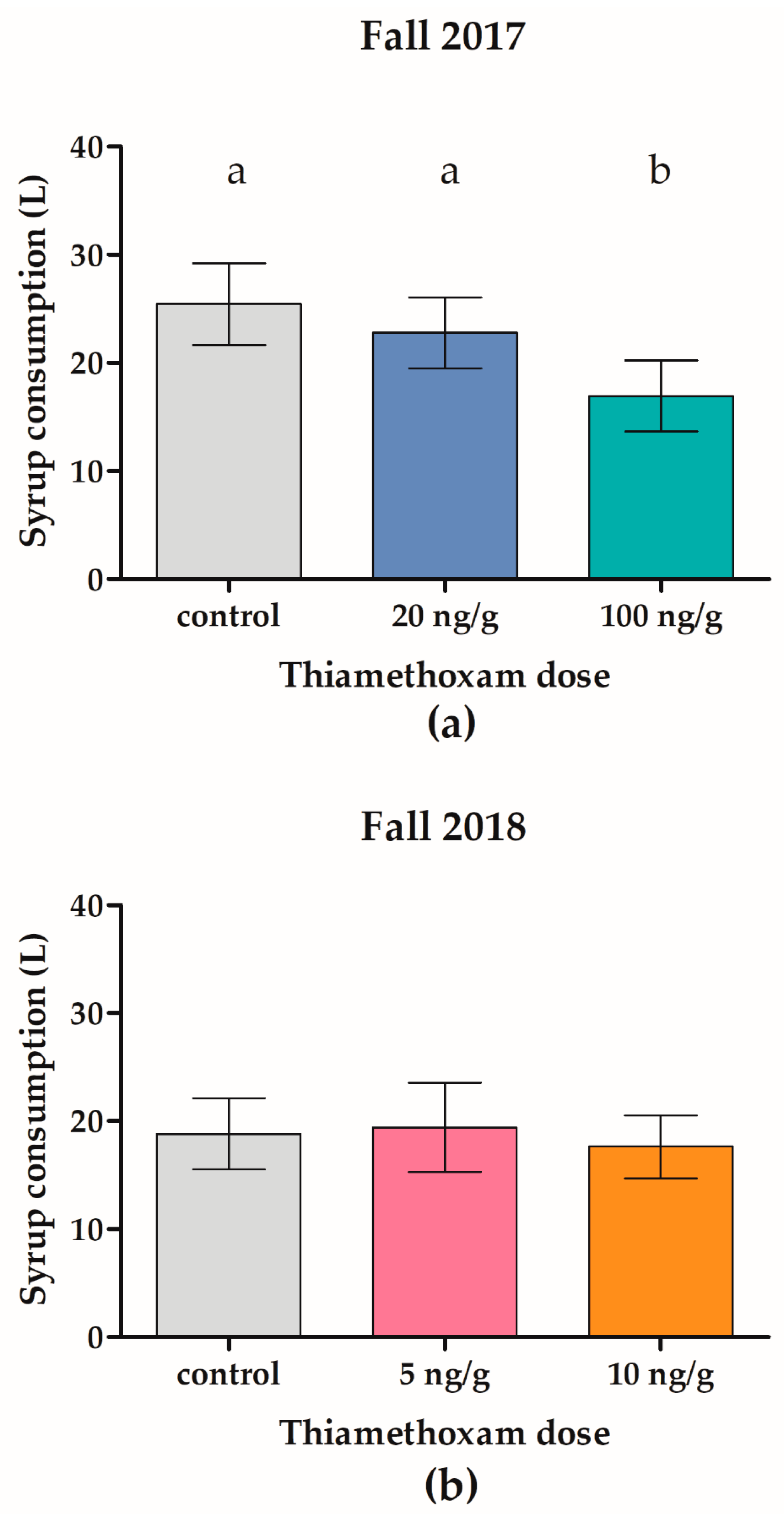
3.1. Effects of Thiamethoxam on Overwintering Colonies of Apis mellifera in Field Trial 1 (F1)
3.2. Effects of Thiamethoxam on Overwintering Colonies of Apis mellifera in Field Trial 2 (F2)
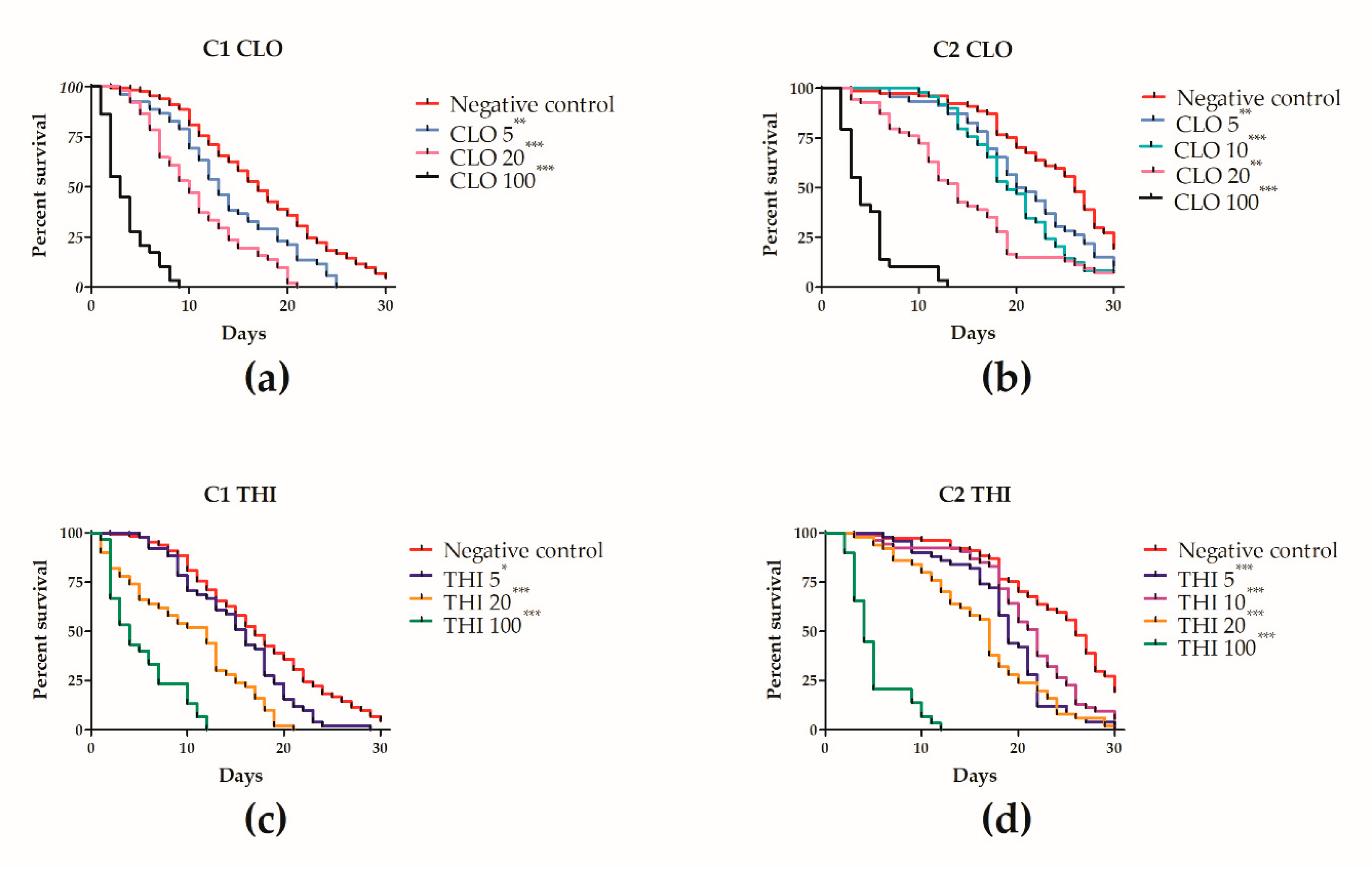
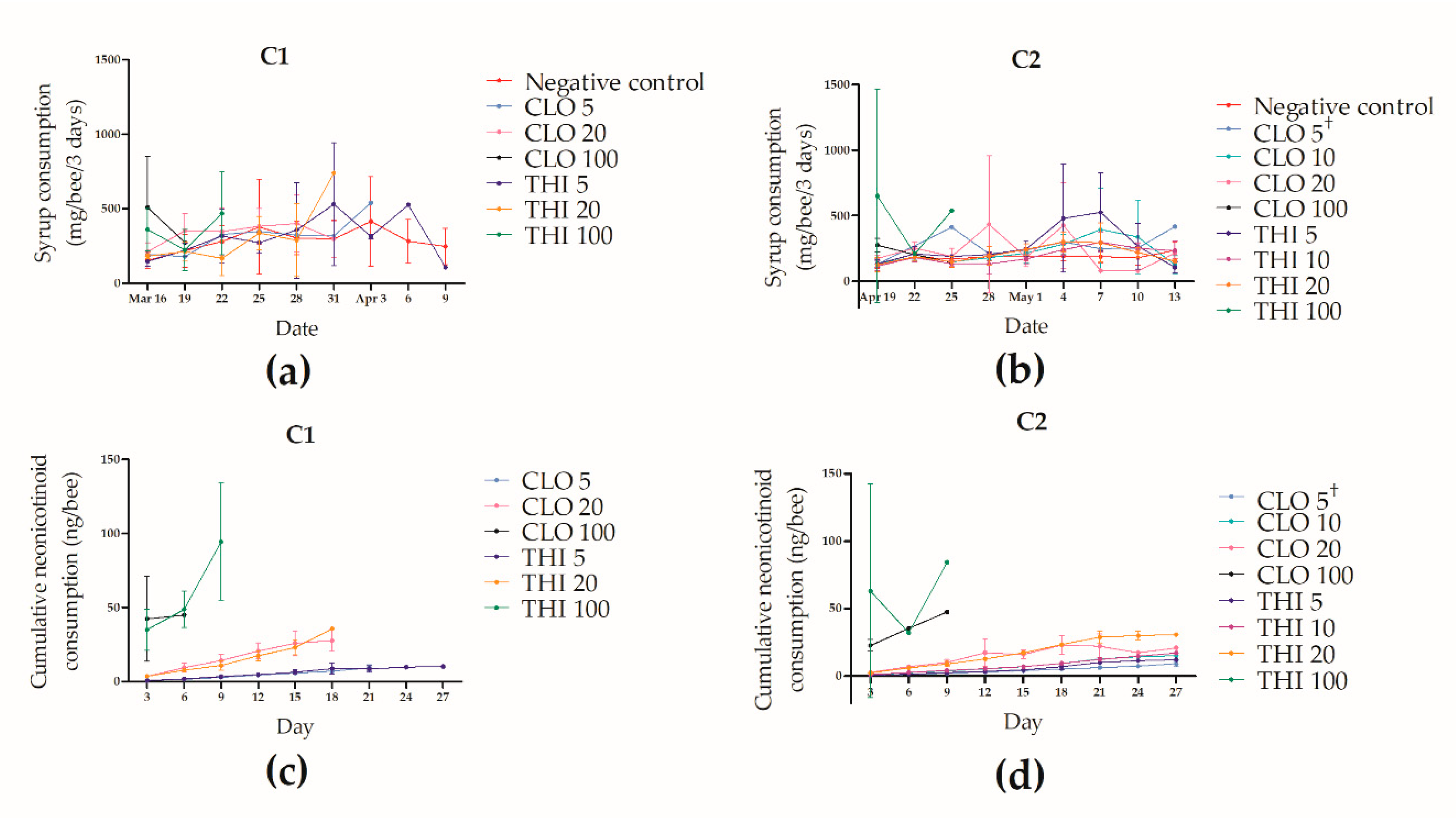
3.3. Effects of Chronic Thiamethoxam or Clothianidin Exposure of Winter Apis mellifera Adult Workers during Laboratory Cage Trial 1 (C1)
3.4. Effects of Chronic Thiamethoxam or Clothianidin Exposure of Winter Apis mellifera Adult Workers during Laboratory Cage Trial 2 (C2)
4. Discussion
4.1. Effects of Thiamethoxam on Overwintering Colonies of Apis mellifera in Field Trial 1 (F1)
4.2. Effects of Thiamethoxam on Overwintering Colonies of Apis mellifera in Field Trial 2 (F2)
4.3. Effects of Chronic Thiamethoxam or Clothianidin Exposure of Winter Apis mellifera Adult Workers in Laboratory Cage Trial 1 (C1)
4.4. Effects of Chronic Thiamethoxam or Clothiandin Exposure of Winter Apis mellifera Adult Workers in Laboratory Cage Trial 2 (C2)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- 2018–2019 Canadian Association of Professional Apiculturists Statement on Colony Losses. Available online: http://www.capabees.com/shared/2018-2019-CAPA-Statement-on-Colony-Losses.pdf (accessed on 21 September 2019).
- Main, A.R.; Headley, J.V.; Peru, K.M.; Michel, N.L.; Cessna, A.J.; Morrissey, C.A. Widespread use and frequent detection of neonicotinoid insecticides in wetlands of Canada’s prairie pothole region. PLoS ONE 2014, 9, e92821. [Google Scholar] [CrossRef]
- Sanchez-Bayo, F.; Goka, K. Pesticide residues and bees—A risk assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codling, G.; Al Naggar, Y.; Giesy, J.P.; Robertson, A.J. Concentrations of neonicotinoid insecticides in honey, pollen and honey bees (Apis mellifera L.) in central Saskatchewan, Canada. Chemosphere 2016, 144, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.; Overmyer, J.; Feken, M.; Ruddle, N.; Vaughan, S.; Scorgie, E.; Bocksch, S.; Hill, M. Thiamethoxam: Long-term effects following honey bee colony-level exposure and implications for risk assessment. Sci. Total Environ. 2019, 654, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Alkassab, A.T.; Kirchner, W.H. Impacts of chronic sublethal exposure to clothianidin on winter honeybees. Ecotoxicology 2016, 25, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Baines, D.; Wilton, E.; Pawluk, A.; de Gorter, M.; Chomistek, N. Neonicotinoids act like endocrine disrupting chemicals in newly-emerged bees and winter bees. Sci. Rep. 2017, 7, 10979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilling, E.; Campbell, P.; Coulson, M.; Ruddle, N.; Tornier, I. A four-year field program investigating long-term effects of repeated exposure of honey bee colonies to flowering crops treated with thiamethoxam. PLoS ONE 2013, 8, e77193. [Google Scholar] [CrossRef]
- Cutler, G.C.; Scott-Dupree, C.D.; Sultan, M.; McFarlane, A.D.; Brewer, L.A. large-scale field study examining effects of exposure to clothianidin seed-treated canola on honey bee colony health, development, and overwintering success. PeerJ 2014, 2, e652. [Google Scholar] [CrossRef]
- Hartfelder, K.; Engels, W. Social insect polymorphism: Hormonal regulation of plasticity in development and reproduction in the honeybee. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 40, pp. 45–77. ISBN 0070-2153. [Google Scholar]
- Fluri, P.; Wille, H.; Gerig, L.; Lüscher, M. Juvenile hormone, vitellogenin and haemocyte composition in winter worker honeybees (Apis mellifera). Experientia 1977, 33, 1240–1241. [Google Scholar] [CrossRef]
- Fluri, P.; Lüscher, M.; Wille, H.; Gerig, L. Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees. J. Insect Physiol. 1982, 28, 61–68. [Google Scholar] [CrossRef]
- Engels, W.; Fahrerthorst, H. Age-and caste-dependent changes in the haemolymph protein patterns of Apis mellifica. Willhelm Roux Archiv. 1974, 174, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.-Y.; Robinson, G.E. Seasonal changes in juvenile hormone titers and rates of biosynthesis in honey bees. J. Comp. Physiol. B 1995, 165, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Crailsheim, K. Dependence of protein metabolism on age and season in the honeybee (Apis mellifica carnica Pollm). J. Insect. Physiol. 1986, 32, 629–634. [Google Scholar] [CrossRef]
- Crailsheim, K. The protein balance of the honey bee worker. Apidologie 1990, 21, 417–429. [Google Scholar] [CrossRef]
- Haszonits, O.; Crailsheim, K. Uptake of L-leucine into isolated enterocytes of the honeybee (Apis mellifera L.) depending on season. J. Insect Physiol. 1990, 36, 835–842. [Google Scholar] [CrossRef]
- Smirle, M.J.; Winston, M.L. Intercolony variation in pesticide detoxification by the honey bee (Hymenoptera: Apidae). J. Econ. Entomol. 1987, 80, 5–8. [Google Scholar] [CrossRef]
- Meled, M.; Thrasyvoulou, A.; Belzunces, L.P. Seasonal variations in susceptibility of Apis mellifera to the synergistic action of prochloraz and deltamethrin. Environ. Toxicol. Chem. 1998, 17, 2517–2520. [Google Scholar] [CrossRef]
- Overmyer, J.; Feken, M.; Ruddle, N.; Bocksch, S.; Hill, M.; Thompson, H. Thiamethoxam honey bee colony feeding study: Linking effects at the level of the individual to those at the colony level. Environ. Toxicol. Chem. 2018, 37, 816–828. [Google Scholar] [CrossRef]
- Wood, S.C.; Kozii, I.V.; Koziy, R.V.; Epp, T.; Simko, E. Comparative chronic toxicity of three neonicotinoids on New Zealand packaged honey bees. PLoS ONE 2018, 13, e0190517. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.V.; Moon, R.D.; Burkness, E.C.; Hutchison, W.D.; Spivak, M. Practical sampling plans for Varroa destructor (Acari: Varroidae) in Apis mellifera (Hymenoptera: Apidae) colonies and apiaries. J. Econ. Entomol. 2010, 103, 1039–1050. [Google Scholar] [CrossRef]
- De Jong, D.; Roma, D.D.A.; Goncalves, L.S. A comparative analysis of shaking solutions for the detection of Varroa jacobsoni on adult honeybees. Apidologie 1982, 13, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Human, H.; Brodschneider, R.; Dietemann, V.; Dively, G.; Ellis, J.D.; Forsgren, E.; Fries, I.; Hatjina, F.; Hu, F.-L.; Jaffé, R. Miscellaneous standard methods for Apis mellifera research. J. Apic. Res. 2013, 52, 1–53. [Google Scholar] [CrossRef] [Green Version]
- Moritz, R.F. The effect of different diluents on insemination success in the honeybee using mixed semen. J. Apic. Res. 1984, 23, 164–167. [Google Scholar] [CrossRef]
- Williams, G.R.; Troxler, A.; Retschnig, G.; Roth, K.; Yañez, O.; Shutler, D.; Neumann, P.; Gauthier, L. Neonicotinoid pesticides severely affect honey bee queens. Sci. Rep. 2015, 5, 14621. [Google Scholar] [CrossRef] [Green Version]
- Meikle, W.G.; Weiss, M.; Stilwell, A.R. Monitoring colony phenology using within-day variability in continuous weight and temperature of honey bee hives. Apidologie 2016, 47, 1–14. [Google Scholar] [CrossRef] [Green Version]
- OECD Test No. 245: Honey Bee (Apis mellifera L.), Chronic Oral Toxicity Test (10-Day Feeding). Available online: https://www.oecd-ilibrary.org/content/publication/9789264284081-en (accessed on 15 December 2019).
- Williams, G.R.; Alaux, C.; Costa, C.; Csaki, T.; Doublet, V.; Eisenhardt, D.; Fries, I.; Kuhn, R.; McMahon, D.P.; Medrzycki, P. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 2013, 52, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Sandrock, C.; Tanadini, M.; Tanadini, L.G.; Fauser-Misslin, A.; Potts, S.G.; Neumann, P. Impact of chronic neonicotinoid exposure on honeybee colony performance and queen supersedure. PLoS ONE 2014, 9, e103592. [Google Scholar] [CrossRef] [Green Version]
- Genersch, E.; Von Der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S. The German bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Bartling, T.M.; Vilcinskas, A.; Lee, K.-Z. Sub-lethal doses of clothianidin inhibit the conditioning and biosensory abilities of the western honeybee Apis mellifera. Insects 2019, 10, 340. [Google Scholar] [CrossRef] [Green Version]
- Colin, T.; Meikle, W.G.; Paten, A.M.; Barron, A.B. Long-term dynamics of honey bee colonies following exposure to chemical stress. Sci. Total. Environ. 2019, 677, 660–670. [Google Scholar] [CrossRef]
- Meikle, W.G.; Weiss, M.; Maes, P.W.; Fitz, W.; Snyder, L.A.; Sheehan, T.; Mott, B.M.; Anderson, K.E. Internal hive temperature as a means of monitoring honey bee colony health in a migratory beekeeping operation before and during winter. Apidologie 2017, 48, 666–680. [Google Scholar] [CrossRef] [Green Version]
- Jacques, A.; Laurent, M.; Ribière-Chabert, M.; Saussac, M.; Bougeard, S.; Budge, G.E.; Hendrikx, P.; Chauzat, M.-P.; EPILOBEE Consortium. A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLoS ONE 2017, 12, e0172591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, L.; Williams, G.R.; Pettis, J.; Fries, I.; Neumann, P. Superorganism resilience: Eusociality and susceptibility of ecosystem service providing insects to stressors. Curr. Opin. Insect Sci. 2015, 12, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.C.; de Mattos, I.M.; Kozii, I.V.; Klein, C.D.; Dvylyuk, I.; Folkes, C.D.; de Carvalho Macedo Silva, R.; Moshynskyy, I.; Epp, T.; Simko, E. Effects of chronic dietary thiamethoxam and prothioconazole exposure on Apis mellifera worker adults and brood. Pest Manag. Sci. 2019, 76, 85–94. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Trial 1 (F1) | Field Trial 2 (F2) | |
|---|---|---|
| Trial dates (September–May) | 2017–2018 | 2018–2019 |
| THI doses (ng/g) | 0, 20, 100 | 0, 5, 10 |
| N (colonies) per dose | 20, 20, 20 | 22, 21, 21 |
| N for queen quality analysis | 6, 6, 6 | - |
| N for temperature analysis | - | 14, 11, 10 |
| Varroa and Nosema monitoring | Yes | No |
| Cage Trial 1 (C1) | Cage Trial 2 (C2) | |||
|---|---|---|---|---|
| Trial Dates | 13 March–11 April 2018 | 16 April–15 May 2018 | ||
| Negative control | 1:1 (w/v) sucrose solution | 1:1 (w/v) sucrose solution | ||
| # negative control cages | 13 | 8 | ||
| Positive control | 1000 ng/g dimethoate | 1000 ng/g dimethoate | ||
| # positive control cages | 3 | 3 | ||
| Mean bees per cage (SD) | 10.1 (0.97) | 9.9 (1.1) | ||
| # diet evaporation cages | 3 | 3 | ||
| Neonicotinoids tested | THI | CLO | THI | CLO |
| Doses (ng/g) | 4.9, 19.5, 97.3 | 4.2, 16.7, 83.2 | 4.9, 9.7, 19.5, 97.3 | 4.2, 8.3, 16.7, 83.2 |
| # cages per dose | 5, 5, 3 | 5, 5, 3 | 5, 5, 5, 3 | 5, 5, 5, 3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, S.C.; Kozii, I.V.; Medici de Mattos, I.; de Carvalho Macedo Silva, R.; Klein, C.D.; Dvylyuk, I.; Moshynskyy, I.; Epp, T.; Simko, E. Chronic High-Dose Neonicotinoid Exposure Decreases Overwinter Survival of Apis mellifera L. Insects 2020, 11, 30. https://doi.org/10.3390/insects11010030
Wood SC, Kozii IV, Medici de Mattos I, de Carvalho Macedo Silva R, Klein CD, Dvylyuk I, Moshynskyy I, Epp T, Simko E. Chronic High-Dose Neonicotinoid Exposure Decreases Overwinter Survival of Apis mellifera L. Insects. 2020; 11(1):30. https://doi.org/10.3390/insects11010030
Chicago/Turabian StyleWood, Sarah C., Ivanna V. Kozii, Igor Medici de Mattos, Roney de Carvalho Macedo Silva, Colby D. Klein, Ihor Dvylyuk, Igor Moshynskyy, Tasha Epp, and Elemir Simko. 2020. "Chronic High-Dose Neonicotinoid Exposure Decreases Overwinter Survival of Apis mellifera L." Insects 11, no. 1: 30. https://doi.org/10.3390/insects11010030