In Vitro Effects of Pesticides on European Foulbrood in Honeybee Larvae

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
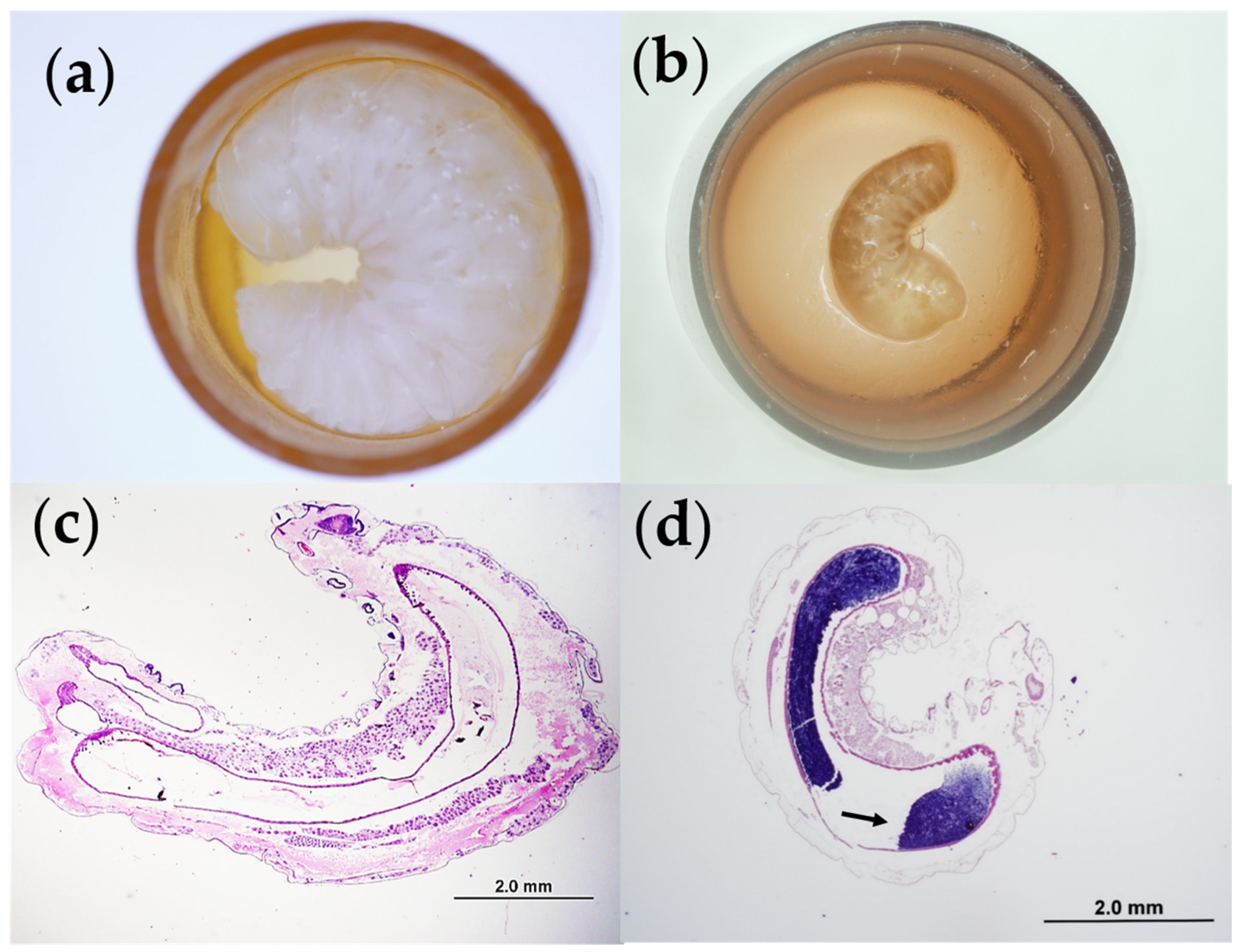
2.1. Isolation of an Atypical Variant of M. Plutonius
2.2. Experimental Animals
2.3. In Vitro Larval Rearing
2.4. Larval Pesticide Exposure
2.5. Larval Infection with M. Plutonius
2.6. Statistical Analysis
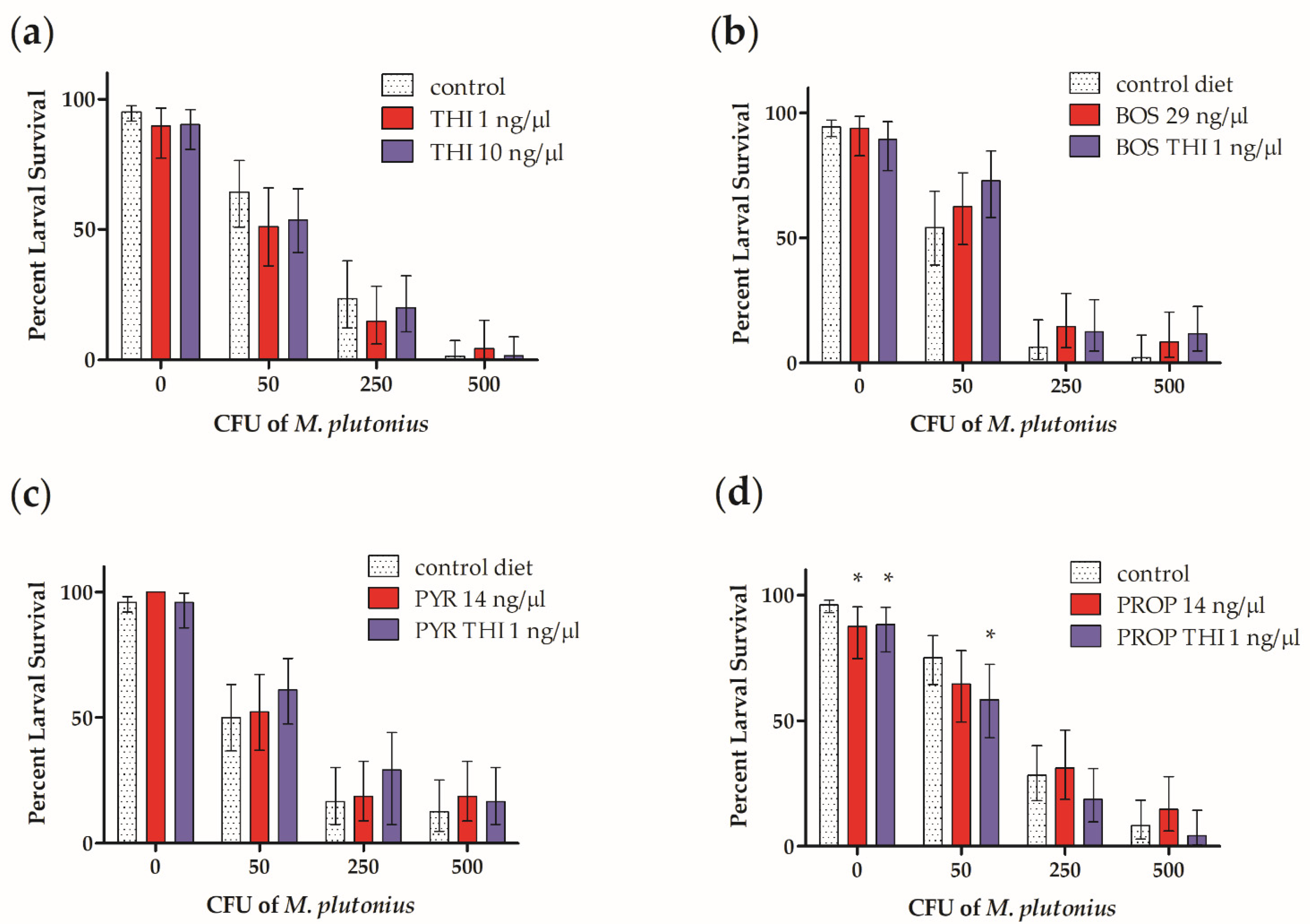
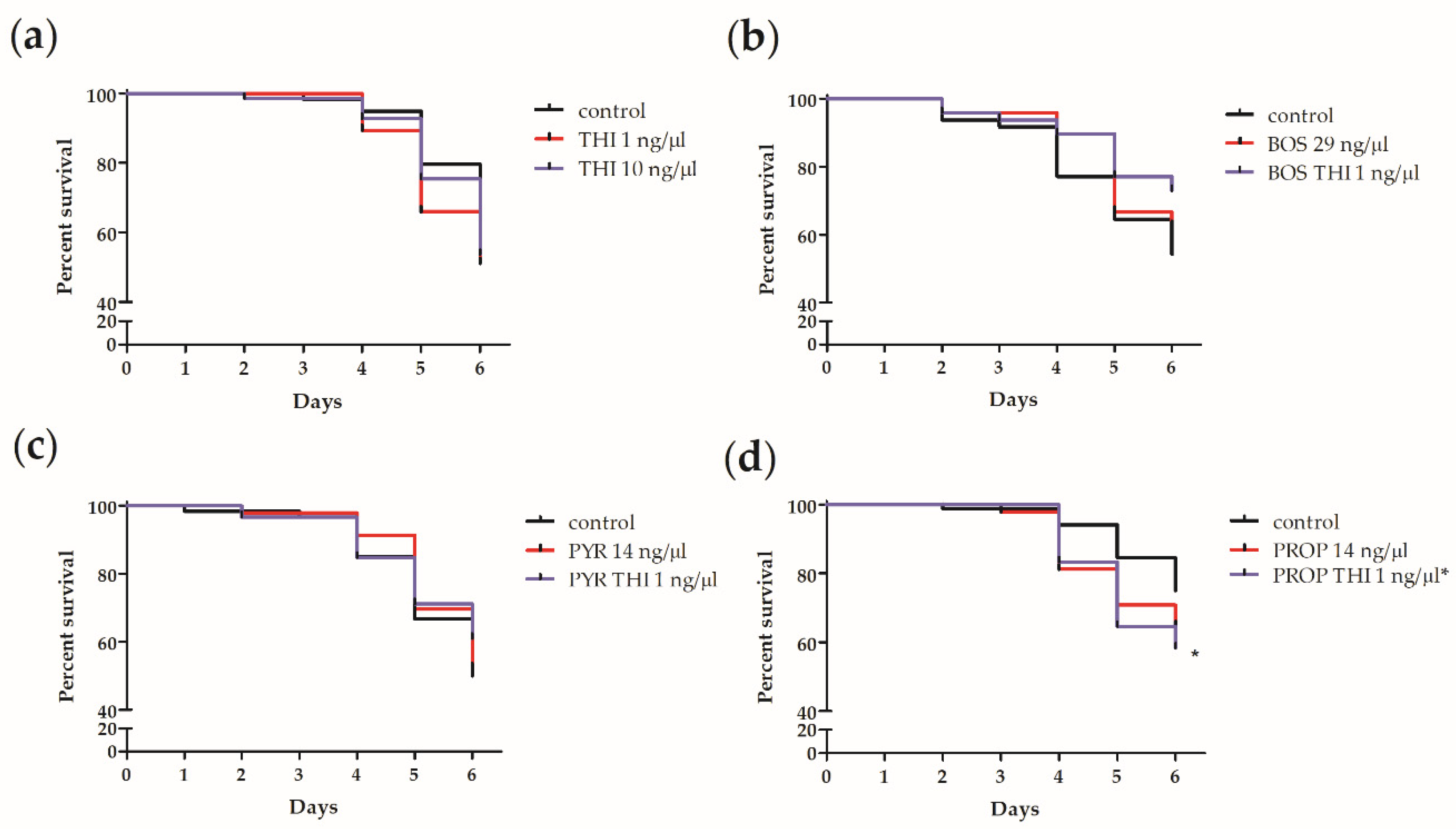
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forsgren, E. European foulbrood in honey bees. J. Invertebr. Pathol. 2010, 103, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- López, J.H.; Krainer, S.; Engert, A.; Schuehly, W.; Riessberger-Gallé, U.; Crailsheim, K. Sublethal pesticide doses negatively affect survival and the cellular responses in American foulbrood-infected honeybee larvae. Sci. Rep. 2017, 7, 40853. [Google Scholar] [CrossRef] [PubMed]
- Gätschenberger, H.; Azzami, K.; Tautz, J.; Beier, H. Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PLoS ONE 2013, 8, e66415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, R.; Miyoshi-Akiyama, T.; Okumura, K.; Morinaga, Y.; Wu, M.; Sugimura, Y.; Yoshiyama, M.; Okura, M.; Kirikae, T.; Takamatsu, D. Development of duplex PCR assay for detection and differentiation of typical and atypical Melissococcus plutonius strains. J. Vet. Med. Sci. 2014, 76, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Djukic, M.; Erler, S.; Leimbach, A.; Grossar, D.; Charrière, J.-D.; Gauthier, L.; Hartken, D.; Dietrich, S.; Nacke, H.; Daniel, R.; et al. Comparative genomics and description of putative virulence factors of Melissococcus plutonius, the causative agent of European foulbrood disease in honey bees. Genes 2018, 9, 419. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Okumura, K.; Harada, M.; Okamoto, M.; Okura, M.; Takamatsu, D. Different impacts of pMP19 on the virulence of Melissococcus plutonius strains with different genetic backgrounds. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Arai, R.; Tominaga, K.; Wu, M.; Okura, M.; Ito, K.; Okamura, N.; Onishi, H.; Osaki, M.; Sugimura, Y.; Yoshiyama, M.; et al. Diversity of Melissococcus plutonius from honeybee larvae in Japan and experimental reproduction of European foulbrood with cultured atypical isolates. PLoS ONE 2012, 7, e33708. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, D.; Sato, M.; Yoshiyama, M. Infection of Melissococcus plutonius clonal complex 12 strain in European honeybee larvae is essentially confined to the digestive tract. J. Vet. Med. Sci. 2016, 78, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Yamazaki, Y.; Shiraishi, A.; Kobayashi, S.; Harada, M.; Yoshiyama, M.; Osaki, M.; Okura, M.; Takamatsu, D. Virulence differences among Melissococcus plutonius strains with different genetic backgrounds in Apis mellifera larvae under an improved experimental condition. Sci. Rep. 2016, 6, 33329. [Google Scholar] [CrossRef] [Green Version]
- McKee, B.A.; Goodman, R.D.; Hornitzky, M.A. The transmission of European foulbrood (Melissococcus plutonius) to artificially reared honey bee larvae (Apis mellifera). J. Apic. Res. 2004, 43, 93–100. [Google Scholar] [CrossRef]
- Lewkowski, O.; Erler, S. Virulence of Melissococcus plutonius and secondary invaders associated with European foulbrood disease of the honey bee. Microbiologyopen 2018, 8, e00649. [Google Scholar] [CrossRef] [PubMed]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traynor, K.S.; Pettis, J.S.; Tarpy, D.R.; Mullin, C.A.; Frazier, J.L.; Frazier, M.; vanEngelsdorp, D. In-hive pesticide exposome: Assessing risks to migratory honey bees from in-hive pesticide contamination in the eastern United States. Sci. Rep. 2016, 6, 33207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdanov, S. Contaminants of bee products. Apidologie 2006, 37, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Burden, R.S.; Cooke, D.T.; Carter, G.A. Inhibitors of sterol biosynthesis and growth in plants and fungi. Phytochemistry 1989, 28, 1791–1804. [Google Scholar] [CrossRef]
- Wood, S.C.; Medici de Mattos, I.; Kozii, I.V.; Klein, C.D.; Dvylyuk, I.; Folkes, C.D.; de Carvalho Macedo Silva, R.; Moshynskyy, I.; Epp, T.; Simko, E. Effects of chronic dietary thiamethoxam and prothioconazole exposure on Apis mellifera worker adults and brood. Pest Manag. Sci. 2019, 76, 85–94. [Google Scholar] [CrossRef]
- Böhme, F.; Bischoff, G.; Zebitz, C.P.W.; Rosenkranz, P.; Wallner, K. From field to food—Will pesticide-contaminated pollen diet lead to a contamination of royal jelly? Apidologie 2018, 49, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Bayo, F.; Goka, K. Pesticide residues and bees—A risk assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [Green Version]
- Codling, G.; Al Naggar, Y.; Giesy, J.P.; Robertson, A.J. Concentrations of neonicotinoid insecticides in honey, pollen and honey bees (Apis mellifera L.) in central Saskatchewan, Canada. Chemosphere 2016, 144, 2321–2328. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Mulhauser, B.; Mulot, M.; Mutabazi, A.; Glauser, G.; Aebi, A. A worldwide survey of neonicotinoids in honey. Science 2017, 358, 109–111. [Google Scholar] [CrossRef] [Green Version]
- Botías, C.; David, A.; Horwood, J.; Abdul-Sada, A.; Nicholls, E.; Hill, E.; Goulson, D. Neonicotinoid residues in wildflowers, a potential route of chronic exposure for bees. Environ. Sci. Technol. 2015, 49, 12731–12740. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.C.; Kozii, I.V.; Koziy, R.V.; Epp, T.; Simko, E. Comparative chronic toxicity of three neonicotinoids on New Zealand packaged honey bees. PLoS ONE 2018, 13, e0190517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.C.; Kozii, I.V.; Medici de Mattos, I.; de Carvalho Macedo Silva, R.; Klein, C.D.; Dvylyuk, I.; Moshynskyy, I.; Epp, T.; Simko, E. Chronic high-dose neonicotinoid exposure decreases overwinter survival of Apis mellifera L. Insects 2020, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, H.; Overmyer, J.; Feken, M.; Ruddle, N.; Vaughan, S.; Scorgie, E.; Bocksch, S.; Hill, M. Thiamethoxam: Long-term effects following honey bee colony-level exposure and implications for risk assessment. Sci. Total Environ. 2019, 654, 60–71. [Google Scholar] [CrossRef]
- Overmyer, J.; Feken, M.; Ruddle, N.; Bocksch, S.; Hill, M.; Thompson, H. Thiamethoxam honey bee colony feeding study: Linking effects at the level of the individual to those at the colony level. Environ. Toxicol. Chem. 2018, 37, 816–828. [Google Scholar] [CrossRef]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- Sgolastra, F.; Medrzycki, P.; Bortolotti, L.; Renzi, M.T.; Tosi, S.; Bogo, G.; Teper, D.; Porrini, C.; Molowny-Horas, R.; Bosch, J. Synergistic mortality between a neonicotinoid insecticide and an ergosterol-biosynthesis-inhibiting fungicide in three bee species. Pest. Manage Sci. 2017, 73, 1236–1243. [Google Scholar] [CrossRef]
- Thompson, H.M.; Fryday, S.L.; Harkin, S.; Milner, S. Potential impacts of synergism in honeybees (Apis mellifera) of exposure to neonicotinoids and sprayed fungicides in crops. Apidologie 2014, 45, 545–553. [Google Scholar] [CrossRef]
- Wade, A.; Lin, C.-H.; Kurkul, C.; Regan, E.R.; Johnson, R.M. Combined toxicity of insecticides and fungicides applied to California almond orchards to honey bee larvae and adults. Insects 2019, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Tesovnik, T.; Zorc, M.; Ristanić, M.; Glavinić, U.; Stevanović, J.; Narat, M.; Stanimirović, Z. Exposure of honey bee larvae to thiamethoxam and its interaction with Nosema ceranae infection in adult honey bees. Environ. Pollut. 2020, 256, 113443. [Google Scholar] [CrossRef] [PubMed]
- Glavinic, U.; Tesovnik, T.; Stevanovic, J.; Zorc, M.; Cizelj, I.; Stanimirovic, Z.; Narat, M. Response of adult honey bees treated in larval stage with prochloraz to infection with Nosema ceranae. PeerJ 2019, 7, e6325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neal, S.T.; Anderson, T.D.; Wu-Smart, J.Y. Interactions between pesticides and pathogen susceptibility in honey bees. Curr. Opin. Insect Sci. 2018, 26, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christen, V.; Schirrmann, M.; Frey, J.E.; Fent, K. Global transcriptomic effects of environmentally relevant concentrations of the neonicotinoids clothianidin, imidacloprid, and thiamethoxam in the brain of honey bees (Apis mellifera). Environ. Sci. Technol. 2018, 52, 7534–7544. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [Green Version]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; vanEngelsdorp, D. Crop pollination exposes honey bees to pesticides which alters their susceptibility to the gut pathogen Nosema ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef]
- Cruickshank, A. Scientists Buzzing Over Whether Blueberry Fields Pose A Health Risk to Bees. Available online: https://www.thestar.com/vancouver/2018/10/27/scientists-buzzing-over-whether-blueberry-fields-pose-a-health-risk-to-bees.html (accessed on 27 October 2018).
- Olmstead, S.; McCallum, R.; Shaw, J. Evaluating the Effect of Feeding Pollen Substitute to Honey Bee colonies Destined for Wild Blueberry Pollination in Colchester County, Nova Scotia. Available online: https://www.perennia.ca/wp-content/uploads/2019/10/ATTTA-FactSheet-Oct-2019.pdf (accessed on 17 February 2020).
- Wardell, G.I. European Foulbrood: Association with Michigan Blueberry Pollination and Control. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 1982. [Google Scholar]
- Guarna, M.M.; Higo, H.; Foster, L.; Pernal, S.F.; Wolf Veiga, P. Bee health and blueberry pollination. HiveLights 2019, 32, 14. [Google Scholar]
- Forsgren, E.; Budge, G.E.; Charrière, J.-D.; Hornitzky, M.A. Standard methods for European foulbrood research. J. Apic. Res. 2013, 52, 1–14. [Google Scholar] [CrossRef]
- Govan, V.A.; Brözel, V.; Allsopp, M.H.; Davison, S. A PCR detection method for rapid identification of Melissococcus pluton in honeybee larvae. Appl. Environ. Microbiol. 1998, 64, 1983–1985. [Google Scholar] [CrossRef] [Green Version]
- Haynes, E.; Helgason, T.; Young, J.P.W.; Thwaites, R.; Budge, G.E. A typing scheme for the honeybee pathogen Melissococcus plutonius allows detection of disease transmission events and a study of the distribution of variants. Environ. Microbiol. Rep. 2013, 5, 525–529. [Google Scholar] [CrossRef]
- Budge, G.E.; Shirley, M.D.; Jones, B.; Quill, E.; Tomkies, V.; Feil, E.J.; Brown, M.A.; Haynes, E.G. Molecular epidemiology and population structure of the honey bee brood pathogen Melissococcus plutonius. ISME J. 2014, 8, 1588. [Google Scholar] [CrossRef] [PubMed]
- Schmehl, D.R.; Tomé, H.V.V.; Mortensen, A.N.; Ferreira Martins, G.; Ellis, J.D. Protocol for the in vitro rearing of honey bee (Apis mellifera L.) workers. J. Apic. Res. 2016, 55, 113–129. [Google Scholar] [CrossRef] [Green Version]
- Dai, P.; Jack, C.J.; Mortensen, A.N.; Bustamante, T.A.; Bloomquist, J.R.; Ellis, J.D. Chronic toxicity of clothianidin, imidacloprid, chlorpyrifos, and dimethoate to Apis mellifera L. larvae reared in vitro. Pest. Manage Sci. 2018, 75, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Environmental Protection Agency BeeREX Version 1.0. Available online: https://www.epa.gov/pesticide-science-and-assessing-pesticide-risks/models-pesticide-risk-assessment (accessed on 17 February 2020).
- Tomizawa, M.; Casida, J.E. Selective toxicity of neonicotinoids attributable to specificity of insect and mammalian nicotinic receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef]
- Avenot, H.F.; Michailides, T.J. Resistance to boscalid fungicide in Alternaria alternata isolates from pistachio in California. Plant Dis. 2007, 91, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Fritz, R.; Lanen, C.; Colas, V.; Leroux, P. Inhibition of methionine biosynthesis in Botrytis cinerea by the anilinopyrimidine fungicide pyrimethanil. Pestic. Sci. 1997, 49, 40–46. [Google Scholar] [CrossRef]
- Feldman, A.T.; Wolfe, D. Tissue processing and hematoxylin and eosin staining. In Histopathology; Methods in Molecular Biology (Methods and Protocols); Day, C., Ed.; Humana Press: New York, NY, USA, 2014; Volume 1180, pp. 31–43. [Google Scholar]
- Brown, R.C.; Hopps, H.C. Staining of bacteria in tissue sections: A reliable Gram stain method. Am. J. Clin. Pathol. 1973, 60, 234–240. [Google Scholar] [CrossRef]
- Fine, J.D.; Cox-Foster, D.L.; Mullin, C.A. An inert pesticide adjuvant synergizes viral pathogenicity and mortality in honey bee larvae. Sci. Rep. 2017, 7, 40499. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Evans, J.D.; Scharf, M.; Ellis, J.D. Gene expression in honey bee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). J. Insect Physiol. 2012, 58, 1042–1049. [Google Scholar] [CrossRef]
- Dickel, F.; Münch, D.; Amdam, G.V.; Mappes, J.; Freitak, D. Increased survival of honeybees in the laboratory after simultaneous exposure to low doses of pesticides and bacteria. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Inoculation with M. Plutonius D0 | Pesticide Administration D0 to D5 |
|---|---|---|
| Pesticide and M. plutonius | 0.5 µL M. plutonius with 500, 250, or 50 CFU | THI and/or BOS, PYR, or PROP |
| Pesticide only | 0.5 µL PBS | THI and/or BOS, PYR, or PROP |
| Survival control | 0.5 µL PBS | none |
| Infected control | 0.5 µL M. plutonius with 500, 250, or 50 CFU | none |
| Positive control | 0.5 µL PBS | THI and BOS |
| Pesticide | Mode of Action | Diet Concentration (ng/µL) | Total Dose (ng) |
|---|---|---|---|
| Thiamethoxam (THI) | Neonicotinoid insecticide which is a nicotinic acetylcholine receptor agonist [48] | 1, 10 | 160, 1600 |
| Boscalid (BOS) † | Carboxamide fungicide which inhibits cellular respiration [49] | 29 | 4680 |
| Pyrimethanil (PYR) | Anilinopyrimidine fungicide which inhibits protein synthesis [50] | 14 | 2240 |
| Propiconazole (PROP) | Triazole fungicide which inhibits sterol biosynthesis and cytochrome P450 monooxygenase enzymes [15] | 14 | 2240 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wood, S.C.; Chalifour, J.C.; Kozii, I.V.; Medici de Mattos, I.; Klein, C.D.; Zabrodski, M.W.; Moshynskyy, I.; Guarna, M.M.; Wolf Veiga, P.; Epp, T.; et al. In Vitro Effects of Pesticides on European Foulbrood in Honeybee Larvae. Insects 2020, 11, 252. https://doi.org/10.3390/insects11040252
Wood SC, Chalifour JC, Kozii IV, Medici de Mattos I, Klein CD, Zabrodski MW, Moshynskyy I, Guarna MM, Wolf Veiga P, Epp T, et al. In Vitro Effects of Pesticides on European Foulbrood in Honeybee Larvae. Insects. 2020; 11(4):252. https://doi.org/10.3390/insects11040252
Chicago/Turabian StyleWood, Sarah C., Jocelyne C. Chalifour, Ivanna V. Kozii, Igor Medici de Mattos, Colby D. Klein, Michael W. Zabrodski, Igor Moshynskyy, M. Marta Guarna, Patricia Wolf Veiga, Tasha Epp, and et al. 2020. "In Vitro Effects of Pesticides on European Foulbrood in Honeybee Larvae" Insects 11, no. 4: 252. https://doi.org/10.3390/insects11040252