
Impact of Rice and Potato Host Plants Is Higher on the Reproduction than Growth of Corn Strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)

 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Culture
2.2. Host Plants
2.3. Measurement of Larval and Pupal Weight
2.4. Life Table Study
2.5. Life Table Data Analysis
3. Results
3.1. Body Weights of Larvae and Pupae of Fall Armyworm
3.2. Development, Survivorship, and Reproduction of Fall Armyworm
3.3. Population Parameters on Corn, Potato, and Rice Plants
3.4. Life Table Study of Fall Armyworm
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [Green Version]
- Ganiger, P.C.; Yeshwanth, H.M.; Muralimohan, K.; Vinay, N.; Kumar, A.R.V.; Chandrashekara, K. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr. Sci. 2018, 115, 621–623. [Google Scholar] [CrossRef]
- Acharya, R.; Akintola, A.A.; Malekera, M.J.; Kamulegeya, P.; Nyakunga, K.B.; Mutimbu, M.K.; Shrestha, Y.K.; Hemayet, J.S.M.; Hoat, T.X.; Dao, H.T.; et al. Genetic relationship of fall armyworm (Spodoptera frugiperda) populations that invaded Africa and Asia. Insects 2021, 12, 439. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-S.; Seo, B.Y.; Lee, J.; Kim, H.; Song, J.H.; Lee, W. First report of the fall armyworm, Spodoptera frugiperda (Smith, 1797) (Lepidoptera, noctuidae), a new migratory pest in Korea. Korean J. Appl. Entomol. 2020, 59, 73–78. [Google Scholar] [CrossRef]
- Vennila, S.; Wang, Z.; Young, K.; Khurana, J.; Cruz, I.; Chen, J.; Reynaud, B.; Delatte, H.; Baufeld, P.; Rajan; et al. G20 discussion group on ‘fall armyworm Spodoptera frugiperda (J. E. Smith) [Lepidoptera: Noctuidae]. In Proceedings of the International Workshop on Facilitatng International Research Collaboration on Transboundary Plant Pests, Tsukuba, Ibaraki, Japan, 27 November 2019. [Google Scholar]
- Cook, D.C.; Gardiner, P.S.; Spafford, H. What will fall armyworm (Lepidoptera: Noctuidae) cost Western Australian agriculture? J. Econ. Entomol. 2021, 114, 1613–1621. [Google Scholar] [CrossRef]
- Pashley, D.P. Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): A sibling species complex? Ann. Entomol. Soc. Am. 1986, 79, 898–904. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gomez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Hruska, A.J.; Gould, F. Fall armyworm (Lepidoptera: Noctuidae) and diatraea lineolata (Lepidoptera: Pyralidae): Impact of larval population level and temporal occurrence on maize yield in Nicaragua. J. Econ. Entomol. 1997, 90, 611–622. [Google Scholar] [CrossRef]
- Pashley, D.P. Quantitative genetics, development, and physiological adaptation in host strains of fall armyworm. Evolution 1988, 42, 93–102. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Meagher, R.L. Behavior and distribution of the two fall armyworm host strains in Florida. Fla. Entomol. 2004, 87, 440–449. [Google Scholar] [CrossRef]
- Juarez, M.L.; Murua, M.G.; Garcia, M.G.; Ontivero, M.; Vera, M.T.; Vilardi, J.C.; Groot, A.T.; Castagnaro, A.P.; Gastaminza, G.; Willink, E. Host association of Spodoptera frugiperda (Lepidoptera: Noctuidae) corn and rice strains in Argentina, Brazil, and Paraguay. J. Econ. Entomol. 2012, 105, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Yang, J.G.; Wang, Z.Y.; Zhu, J.S.; Jiang, Y.Y.; Xu, Z.C.; Zhu, P.; Wang, Z.H.; Yu, Y.; Men, X.Y.; et al. Spodoptera frugiperda were found damaging potato in Shandong province. Plant Prot. 2019, 45, 84–86. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Ozgokce, M.S.; Guncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Birch, L.C. The intrinsic rate of natural increase of an insectpopulation. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]
- Leslie, P.H. On the use of matrices in certain population mathematics. Biometrika 1945, 33, 183–212. [Google Scholar] [CrossRef]
- Lewis, E.G. On the generation and growth of a population. Indian J. Stat. 1942, 6, 93–96. [Google Scholar]
- Carey, J.R. Applied Demography for Biologists, with Special Emphasis on Insects; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Huang, Y.; Chi, H. The age-stage, two-sex life table with an offspring sex ratio dependent on female age. J. Agric. For. 2011, 60, 337–345. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sex and variable development rate among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Acharya, R.; Yu, Y.S.; Shim, J.K.; Lee, K.Y. Virulence of four entomopathogenic nematodes against the tobacco cutworm Spodoptera litura Fabricius. Biol. Control 2020, 150, 104348. [Google Scholar] [CrossRef]
- Acharya, R.; Hwang, H.S.; Mostafiz, M.M.; Yu, Y.S.; Lee, K.Y. Susceptibility of various developmental stages of the fall armyworm, Spodoptera frugiperda, to entomopathogenic nematodes. Insects 2020, 11, 868. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2022. [Google Scholar]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap, Monographs on Statistics and Applied Probability; Chapman & Hall: New York, NY, USA, 1993. [Google Scholar]
- Huang, Y.B.; Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Wang, W.; He, P.; Zhang, Y.; Liu, T.; Jing, X.; Zhang, S. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in china. Insects 2020, 11, 639. [Google Scholar] [CrossRef]
- Emden, H. Mechanisms of resistance: Antibiosis, antixenosis, tolerance, nutrition. In Encyclopedia of Pest Management; Pimentel, D., Ed.; Marcel Dekke: New York, NY, USA, 2002; pp. 483–600. [Google Scholar]
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches; Smith, C.M., Ed.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Wu, L.H.; Zhou, C.; Long, G.Y.; Yang, X.B.; Wei, Z.Y.; Liao, Y.J.; Yang, H.; Hu, C.X. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J. Integr. Agric. 2021, 20, 755–763. [Google Scholar] [CrossRef]
- Sousa, F.F.; Mendes, S.M.; Santos-Amaya, O.F.; Araujo, O.G.; Oliveira, E.E.; Pereira, E.J.G. Life-history traits of Spodoptera frugiperda populations exposed to low-dose Bt maize. PLoS ONE 2016, 11, e0156608. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.J.; Zhang, L.; Cheng, Y.X.; Jiang, X. Age-stage two-sex life table for laboratory populations of fall armyworm, Spodoptera frugiperda at different temperatures. Plant Prot. 2019, 45, 20–27. [Google Scholar]
- Ozgokce, M.S.; Chi, H.; Atlıhan, R.; Kara, H. Demography and population projection of myzus persicae (sulz.) (Hemiptera: Aphididae) on five pepper (Capsicum annuum L.) cultivars. Phytoparasitica 2018, 46, 153–167. [Google Scholar] [CrossRef]
- Kazemi, M.H.; Talebi-Chaichi, P.; Shakiba, M.R.; Jafarloo, M.M. Biological responses of Russian wheat aphid, Diuraphis noxia (Mordvilko) (Homoptera: Aphididae), to different wheat varieties. J. Agric. Sci. Technol. 2001, 3, 249–255. [Google Scholar]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2020, 28, 602–610. [Google Scholar] [CrossRef]
- Tsai, J.H.; Wang, J. Effects of host plants on biology and life table parameters of Aphis spiraecola (Homoptera: Aphididae) JAMES. Environ. Entomol. 2001, 30, 44–50. [Google Scholar] [CrossRef]
- Silva-Brandao, K.L.; Horikoshi, R.J.; Bernardi, D.; Omoto, C.; Figueria, A.; Brandao, M.M. Transcript expression plasticity as a response to alternative larval host plants in the speciation process of corn and rice strains of Spodoptera frugiperda. BMC Genom. 2017, 18, 792. [Google Scholar] [CrossRef]
- Orsucci, M.; Mone, Y.; Audiot, P.; Gimenez, S.; Nhim, S.; Boudon, J.-P.; Vabre, M.; Rialle, S.; Koual, R.; Kergoat, G.J.; et al. Transcriptional differences between the two host strains of Spodoptera frugiperda (Lepidoptera: Noctuidae). bioRxiv 2020, 2, 263186. [Google Scholar] [CrossRef]
- Dunse, K.M.; Stevens, J.A.; Lay, F.T.; Gaspar, Y.M.; Heath, R.L.; Anderson, M.A. Coexpression of potato type I and II proteinase inhibitors gives cotton plants protection against insect damage in the field. Proc. Natl. Acad. Sci. USA 2010, 107, 15011–15015. [Google Scholar] [CrossRef] [Green Version]
- Duceppe, M.O.; Cloutier, C.; Michaud, D. Wounding, insect chewing and phloem sap feeding differentially alter the leaf proteome of potato, Solanum tuberosum L. Proteome Sci. 2012, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, P.C.; Kimmins, F.M.; Grayer, R.J.; Raveendranath, S. Schaftosides from rice phloem as feeding inhibitors and resistance factors to brown planthoppers, Nilaparvata lugens. Entomol. Exp. Appl. 1996, 80, 246–249. [Google Scholar] [CrossRef]






{kind=link}
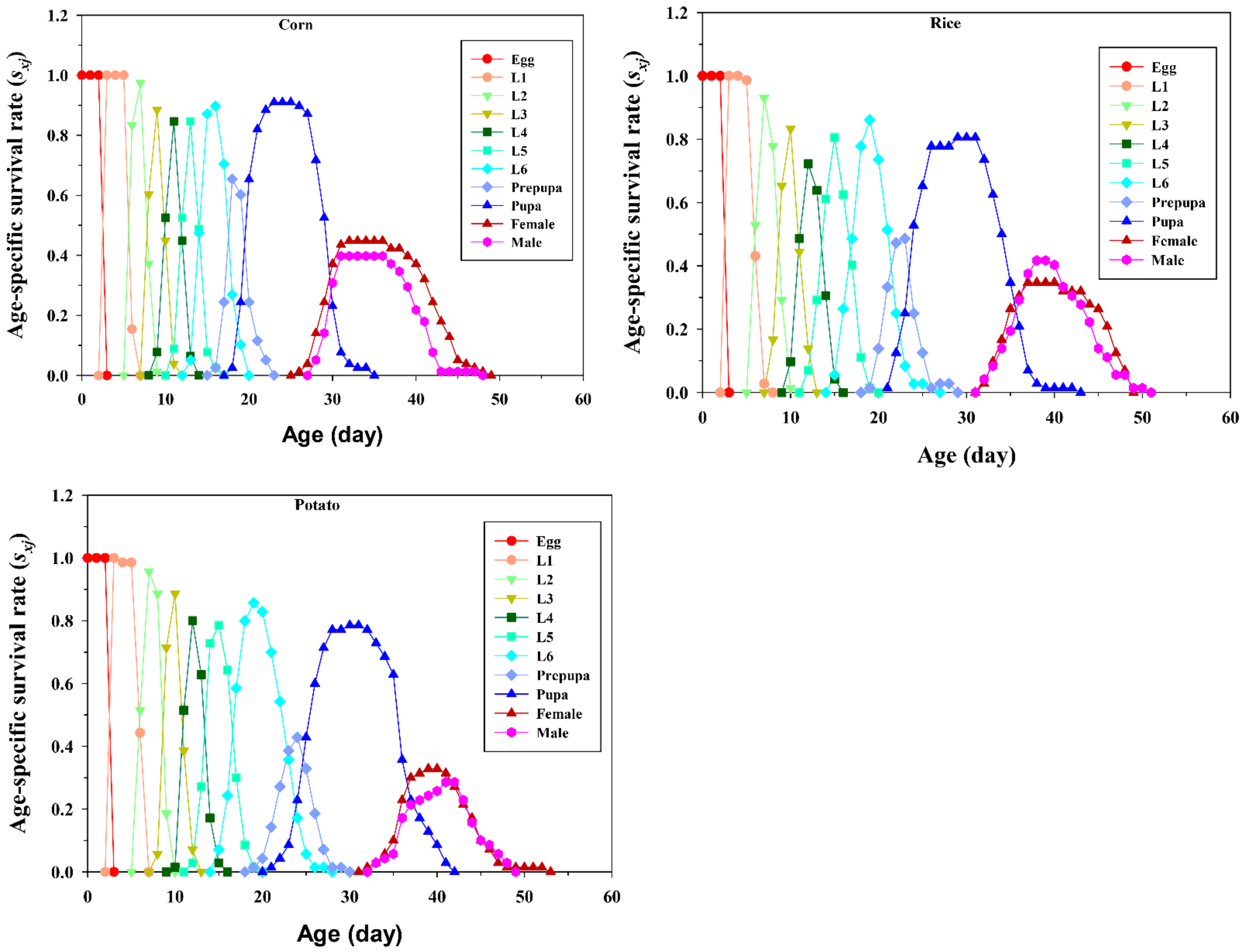{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Corn | Rice | Potato | |||
|---|---|---|---|---|---|---|
| n | (Mean ± SE) | n | (Mean ± SE) | n | (Mean ± SE) | |
| Egg | 3 ± 0.00 a | 3 ± 0.00 a | 3 ± 0.00 a | |||
| First instar | 78 | 3.16 ± 0.04 a | 72 | 3.48 ± 0.07 b | 70 | 3.46 ± 0.06 b |
| Second instar | 77 | 2.24 ± 0.05 a | 69 | 2.66 ± 0.06 b | 76 | 2.68 ± 0.06 b |
| Third instar | 76 | 2.03 ± 0.06 a | 68 | 2.37 ± 0.06 b | 63 | 2.35 ± 0.06 b |
| Fourth instar | 76 | 2.01 ± 0.02 a | 67 | 2.46 ± 0.06 b | 63 | 2.40 ± 0.07 b |
| Fifth instar | 76 | 2.11 ± 0.04 a | 67 | 3.18 ± 0.09 b | 63 | 3.19 ± 0.08 b |
| Sixth instar | 74 | 3.59 ± 0.08 a | 65 | 4.53 ± 0.13 b | 62 | 6.00 ± 0.14 c |
| Prepupa | 74 | 2.04 ± 0.02 a | 60 | 2.28 ± 0.06 b | 59 | 2.27 ± 0.06 b |
| Pupa | 72 | 9.58 ± 0.12 a | 58 | 11.03 ± 0.12 b | 55 | 11.57 ± 0.16 c |
| Preadult | 72 | 29.75 ± 0.19 a | 58 | 35.19 ± 0.27 b | 55 | 36.87 ± 0.32 c |
| Preadult survival rate | 72 | 0.91 ± 0.03 c | 58 | 0.81 ± 0.05 b | 55 | 0.77 ± 0.05 a |
| Adult longevity | ||||||
| Female | 37 | 12.86 ± 0.49 b | 26 | 11.31 ± 0.62 b | 26 | 8.04 ± 0.49 a |
| Male | 34 | 10.17 ± 0.46 c | 32 | 8.77 ± 0.41 b | 28 | 6.18 ± 0.44 a |
| Mean fecundity | 37 | 955.62 ± 58.72 c | 26 | 590.77 ± 46.30 b | 26 | 231.54 ± 28.48 a |
| APOP | 37 | 2.92 ± 0.10 a | 26 | 4.46 ± 0.16 c | 26 | 4.05 ± 0.10 b |
| TPOP | 37 | 32.41 ± 0.31 a | 26 | 38.96 ± 0.37 b | 26 | 40.32 ± 0.43 c |
| Oviposition days | 37 | 7.68 ± 0.388 c | 26 | 5.83 ± 0.28 b | 26 | 3.23 ± 0.24 a |
| Parameter | Corn | Rice | Potato |
|---|---|---|---|
| Cohort size (n) | 78 | 72 | 70 |
| Intrinsic rate of increase (r) (d−1) | 0.17 ± 0.01 b | 0.13 ± 0.01 a | 0.11 ± 0.01 a |
| Finite rate of increase (λ) (d−1) | 1.19 ± 0.01 b | 1.14 ± 0.01 a | 1.11 ± 0.01 a |
| Net reproductive rate (R0) (offspring/individual) | 472.28 ± 62.89 c | 213.33 ± 37.28 b | 86 ± 16.98 a |
| Mean generation time (T) (d) | 35.22 ± 0.30 a | 41.19 ± 0.38 b | 41.89 ± 0.48 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acharya, R.; Malekera, M.J.; Dhungana, S.K.; Sharma, S.R.; Lee, K.-Y. Impact of Rice and Potato Host Plants Is Higher on the Reproduction than Growth of Corn Strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 256. https://doi.org/10.3390/insects13030256
Acharya R, Malekera MJ, Dhungana SK, Sharma SR, Lee K-Y. Impact of Rice and Potato Host Plants Is Higher on the Reproduction than Growth of Corn Strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects. 2022; 13(3):256. https://doi.org/10.3390/insects13030256
Chicago/Turabian StyleAcharya, Rajendra, Matabaro Joseph Malekera, Sanjeev Kumar Dhungana, Sushant Raj Sharma, and Kyeong-Yeoll Lee. 2022. "Impact of Rice and Potato Host Plants Is Higher on the Reproduction than Growth of Corn Strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)" Insects 13, no. 3: 256. https://doi.org/10.3390/insects13030256