
High Expression of PDK4 Could Play a Potentially Protective Role by Attenuating Oxidative Stress after Subarachnoid Hemorrhage

 ,
,
Abstract
:1. Introduction
2. Method and Materials
2.1. Rat Model
2.2. Primary Neuron Culture
2.3. Patient Population
- (1)
- patient age between 16 and 70 years;
- (2)
- signed consent from the subject or next of kin; and
- (3)
- the patient had no contraindications to lumbar puncture.
- (1)
- failure to meet the inclusion criteria or unfit for the experiment as determined by the responsible doctor;
- (2)
- presence of severe cardiac insufficiency, renal dysfunction, diabetes or other systemic diseases; and
- (3)
- a history of traumatic brain injury, severe cerebral edema or hydrocephaly.
- (1)
- patient age between 16 and 70 years;
- (2)
- no current or pre-existing brain injuries, neurological diseases, or bleeding disorders, and subjects who needed surgery for knee arthritis; and
- (3)
- signed consent from the subject or next of kin.
2.4. Sample Collection
2.5. Experimental Design
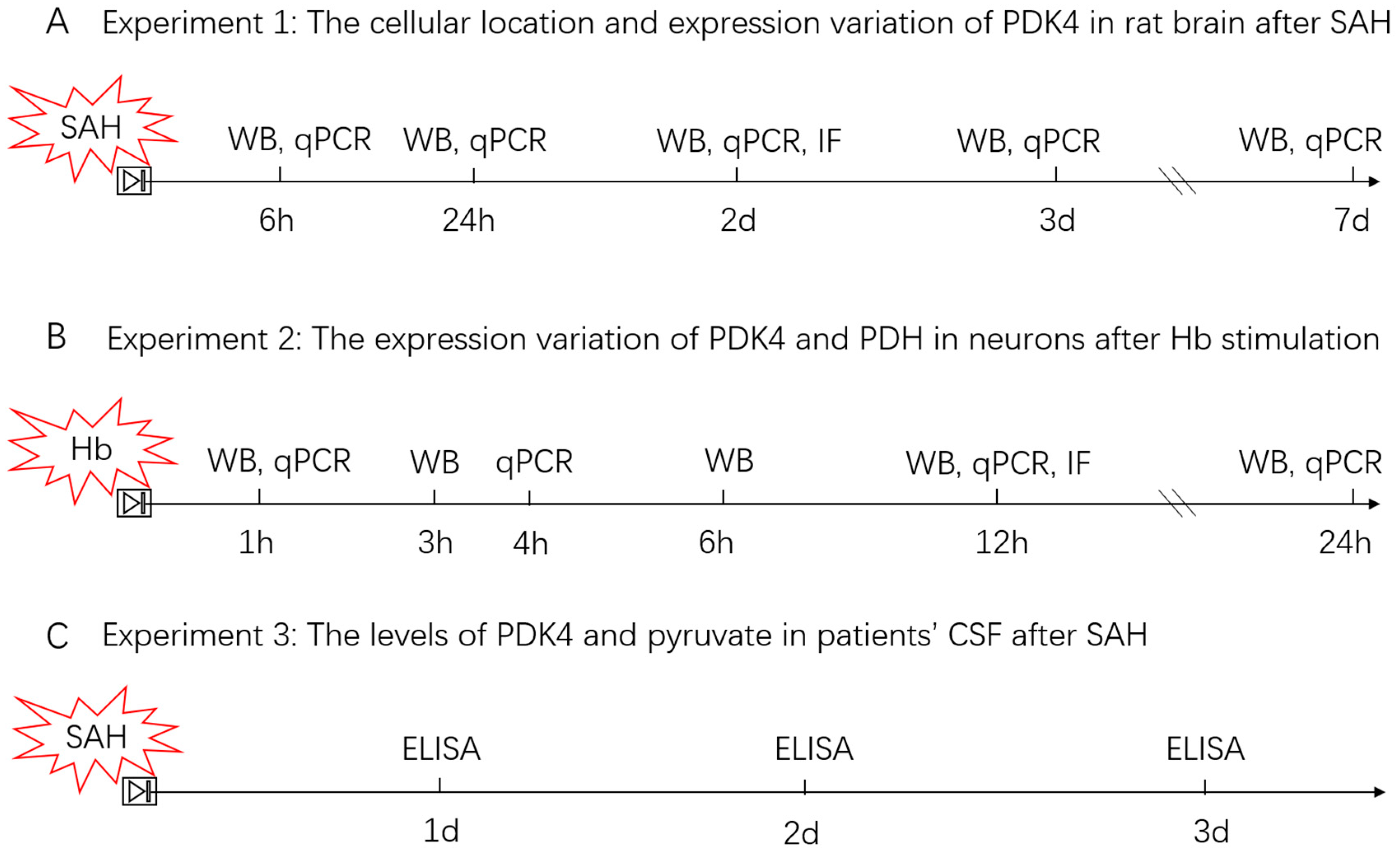
2.5.1. Experiment 1
2.5.2. Experiment 2
2.5.3. Experiment 3
2.6. Western Blot
2.7. Real-Time PCR
2.8. Immunofluorescence Staining
2.9. Enzyme Linked Immunosorbent Assay
2.10. Pyruvate Assay
2.11. Terminal Deoxynucleotidyl Transferase–Mediated dUTP Nick End Labeling
2.12. Modified Garcia Scale
2.13. Statistical Analysis
3. Results
3.1. Mortality and Exclusion
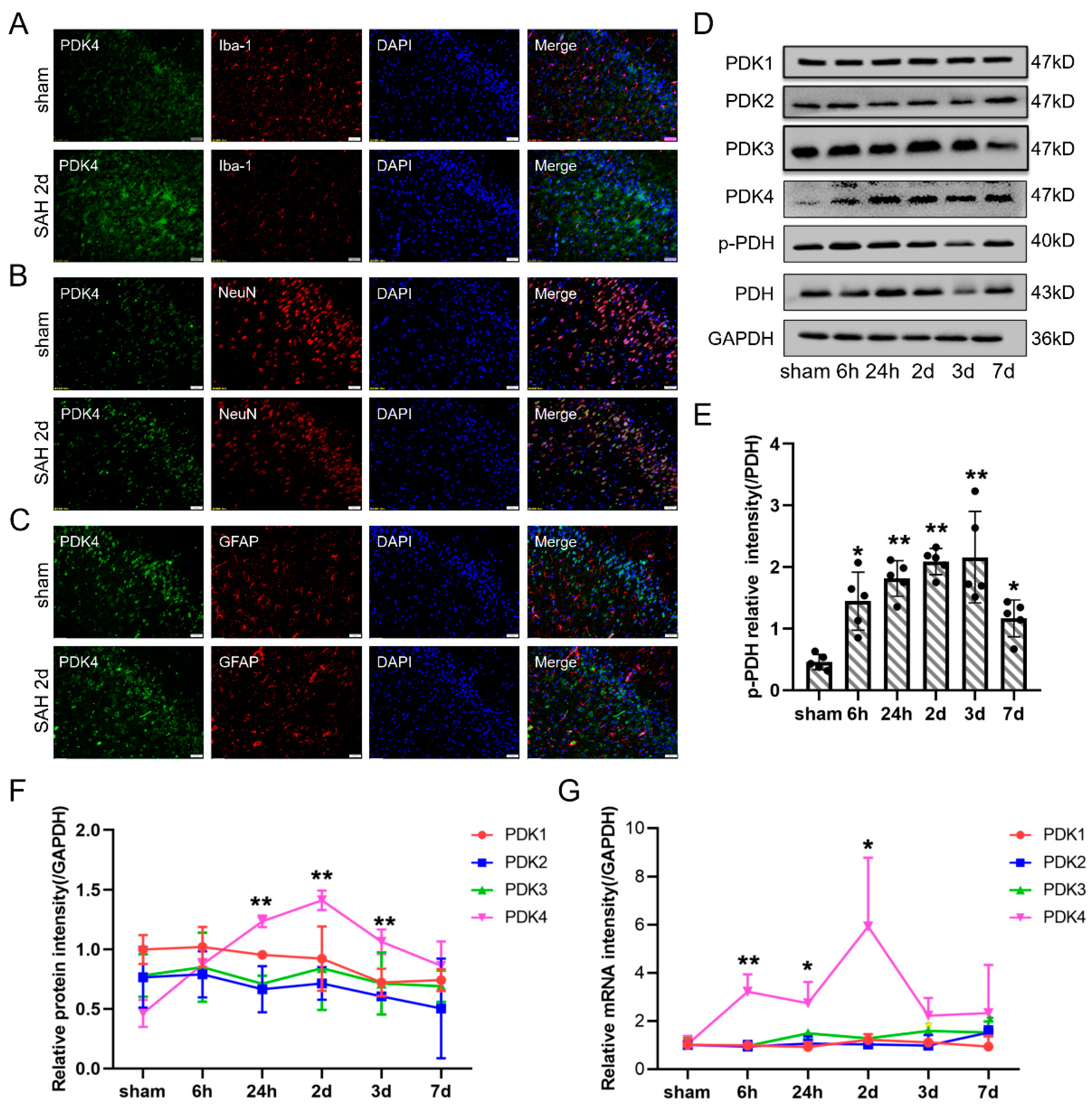
3.2. The Expression of PDK4 but Not PDH Was Elevated in Neurons after SAH in Rats
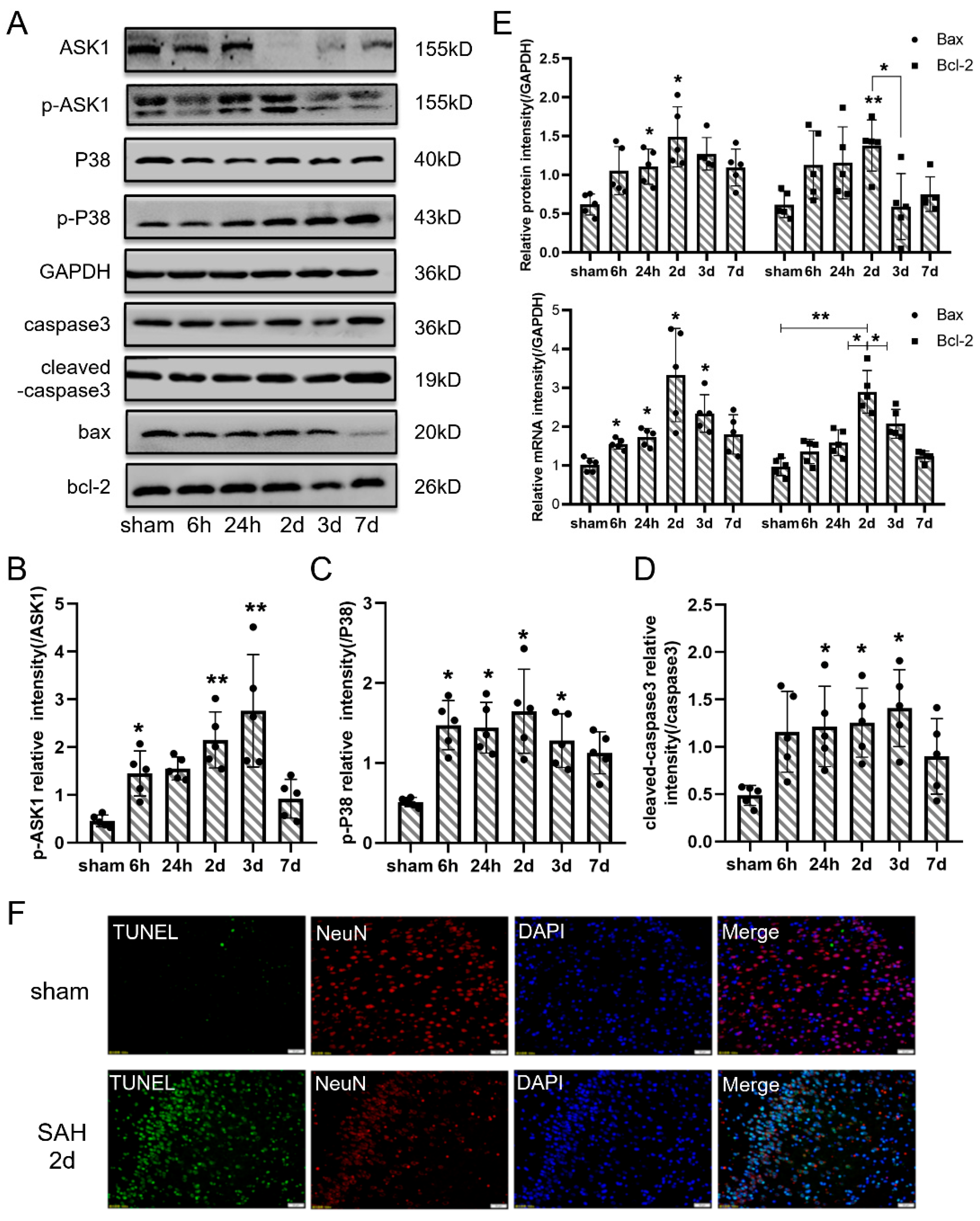
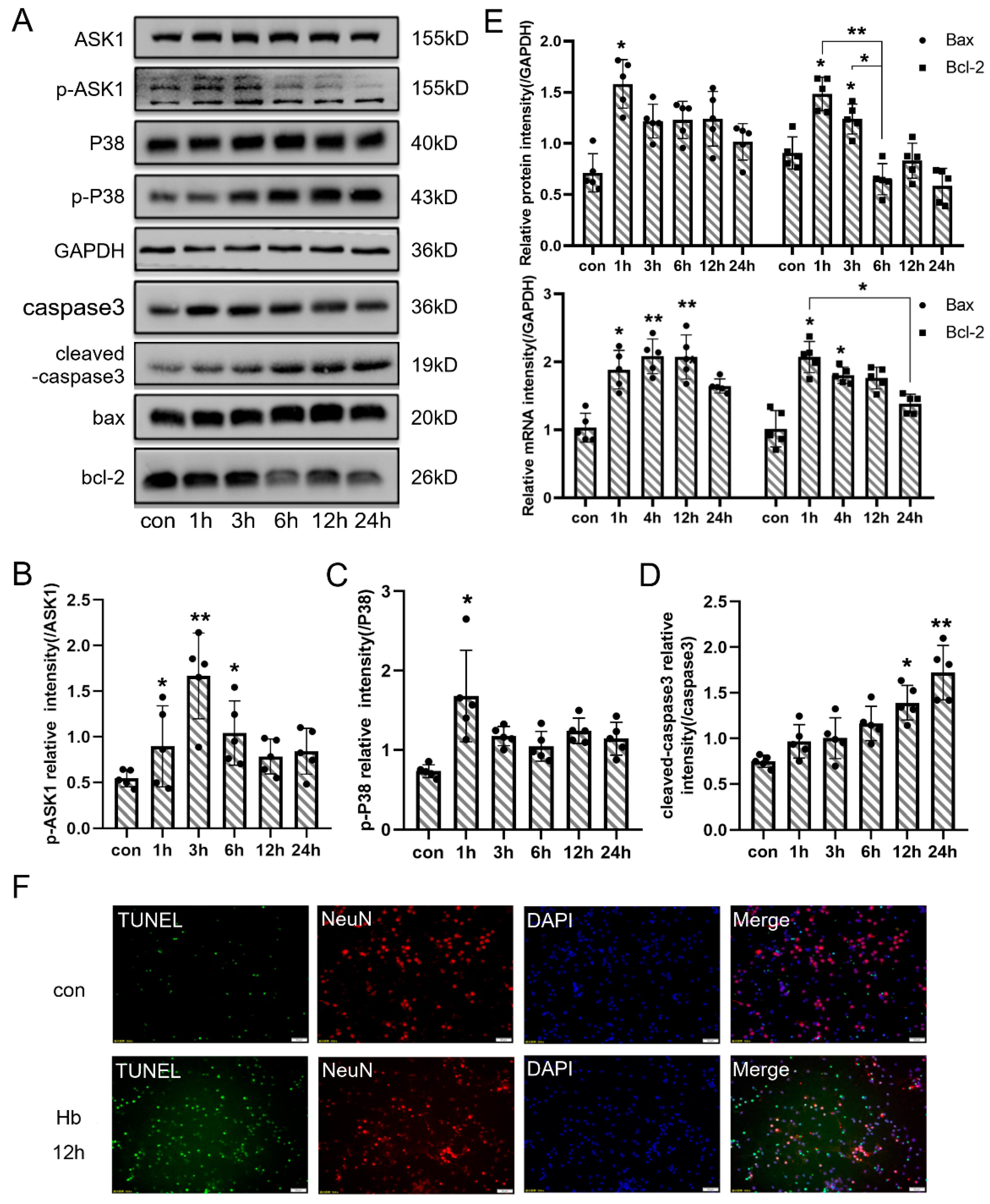
3.3. Neuronal Apoptosis Participated in the Pathology of EBI In Vivo
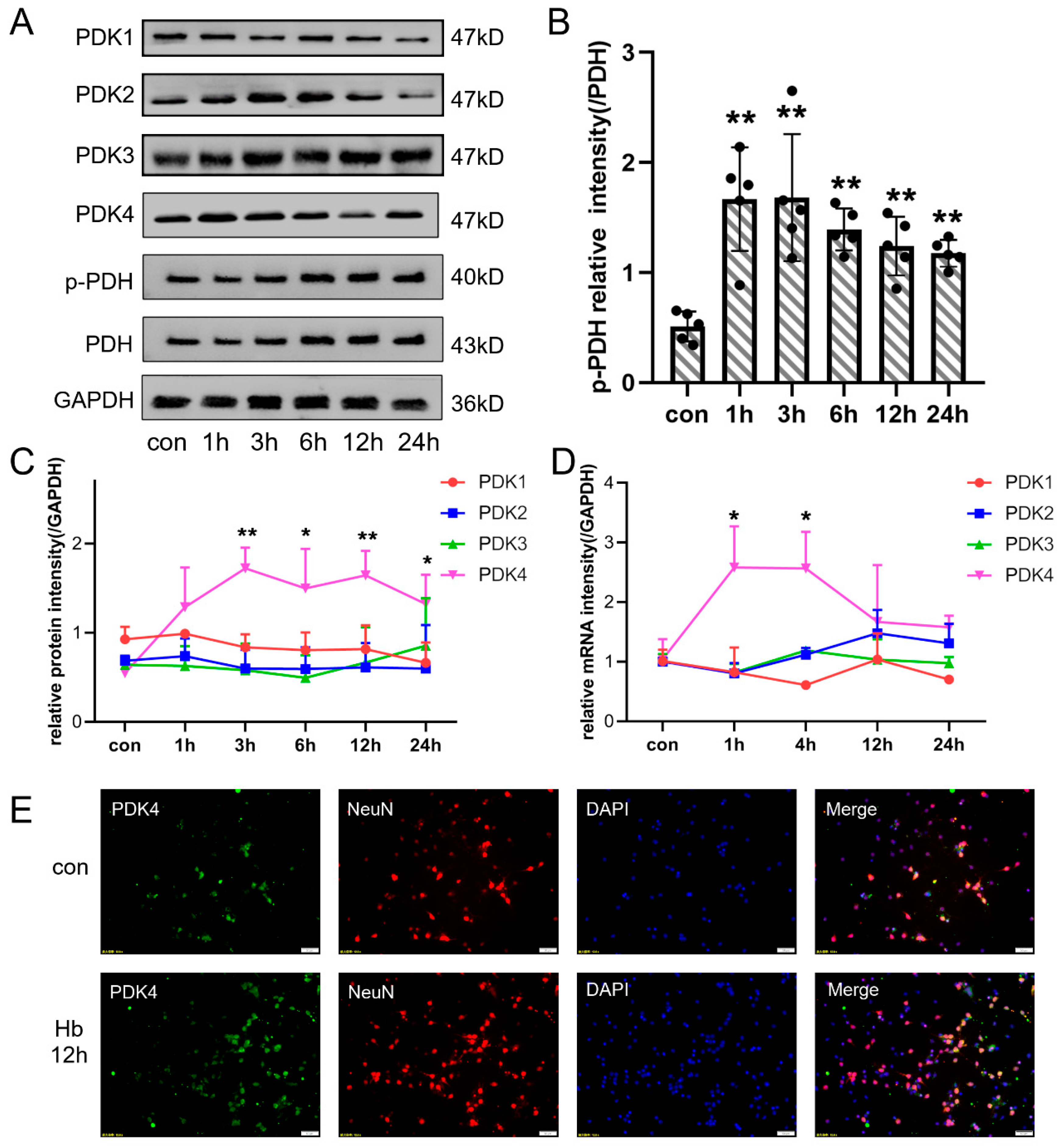
3.4. Hb Exposure Also Induced the Overexpression of PDK4 in Neurons In Vitro
3.5. Hb Exposure Activated the Pathway of Neuronal Apoptosis In Vitro
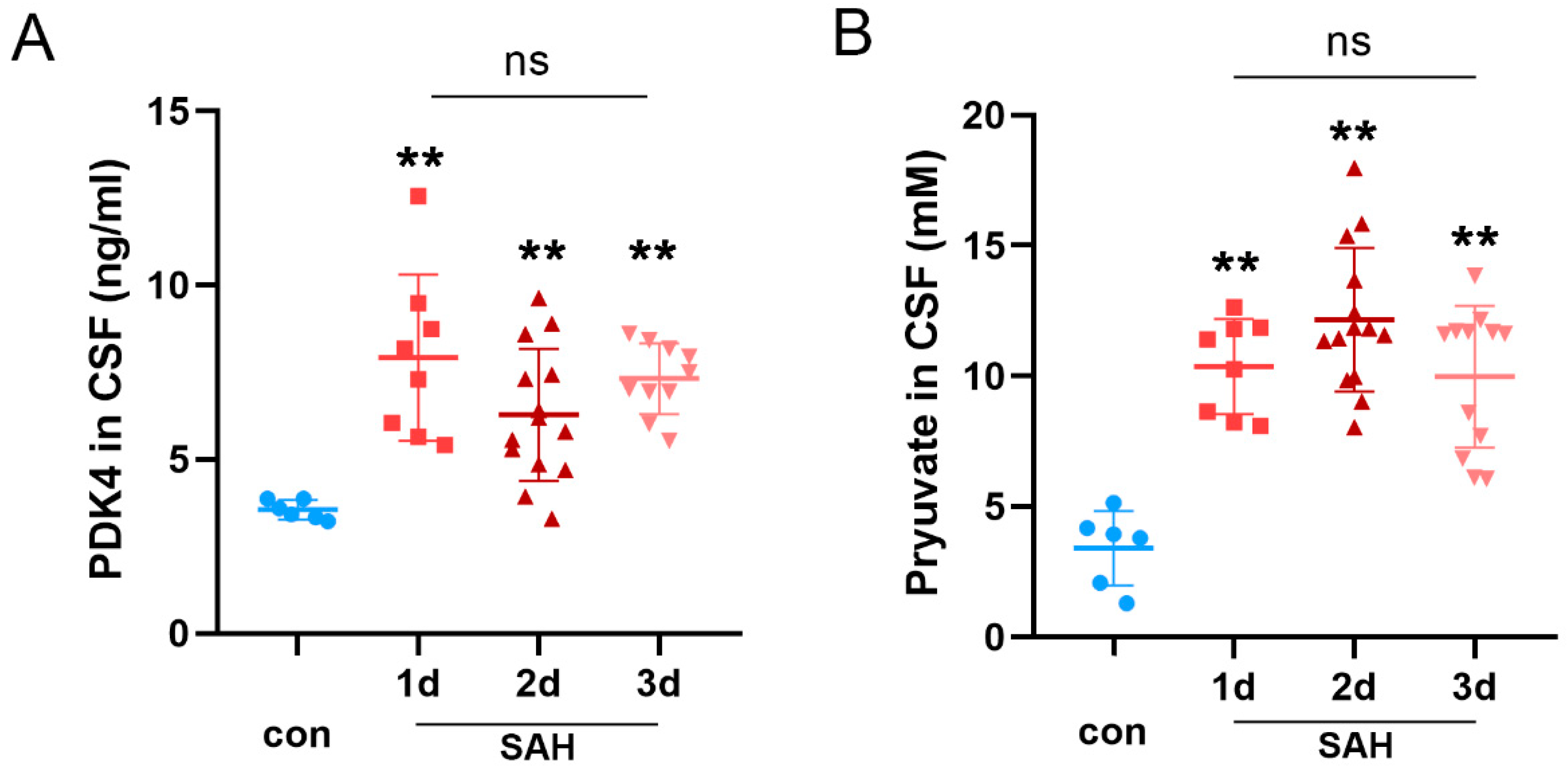
3.6. The Expression of PDK4 Was Elevated in the CSF of SAH Patients
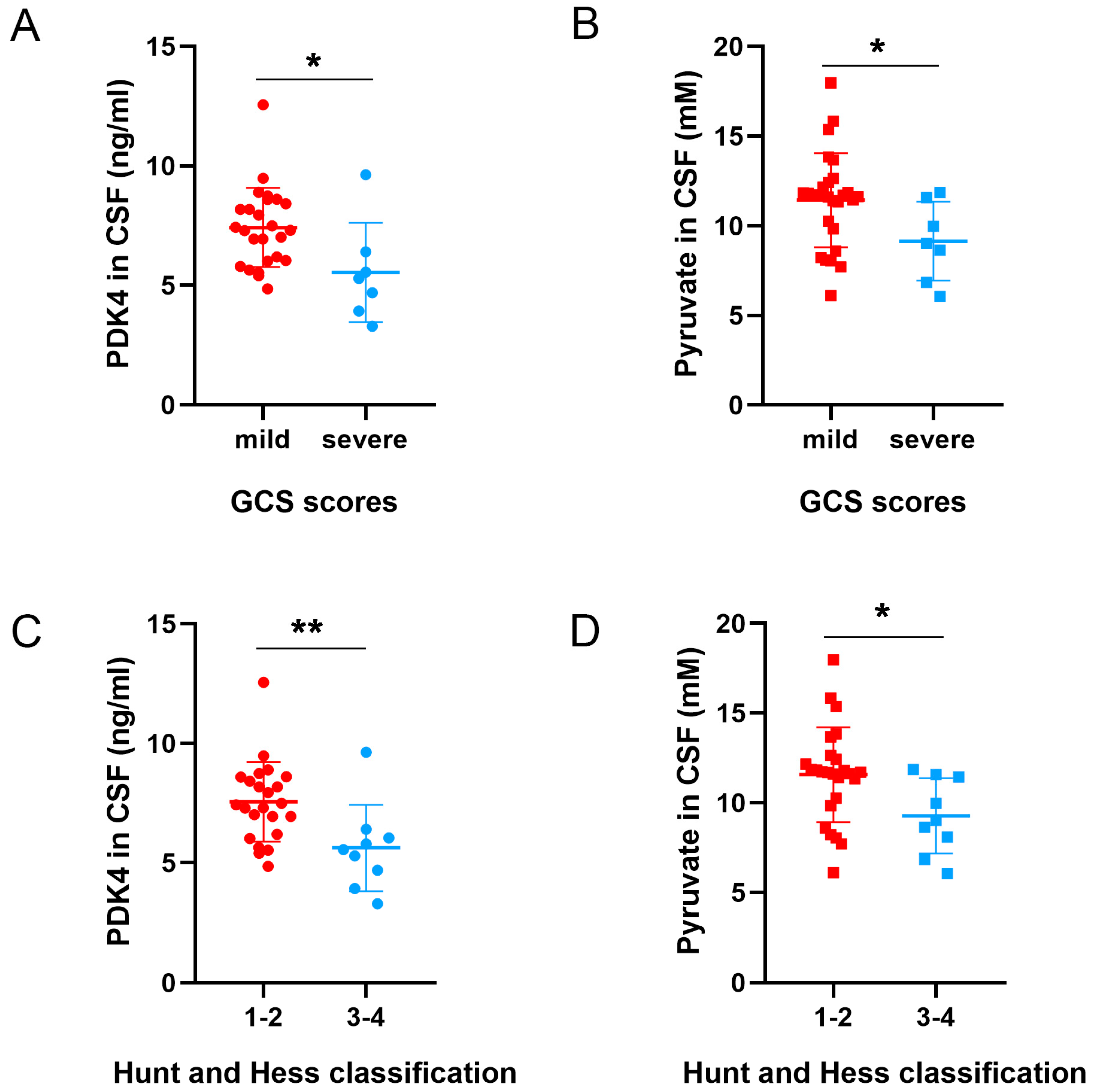
3.7. Different Levels of PDK4 and Pyruvate Were Associated with the Neurological Function Scores of SAH Patients
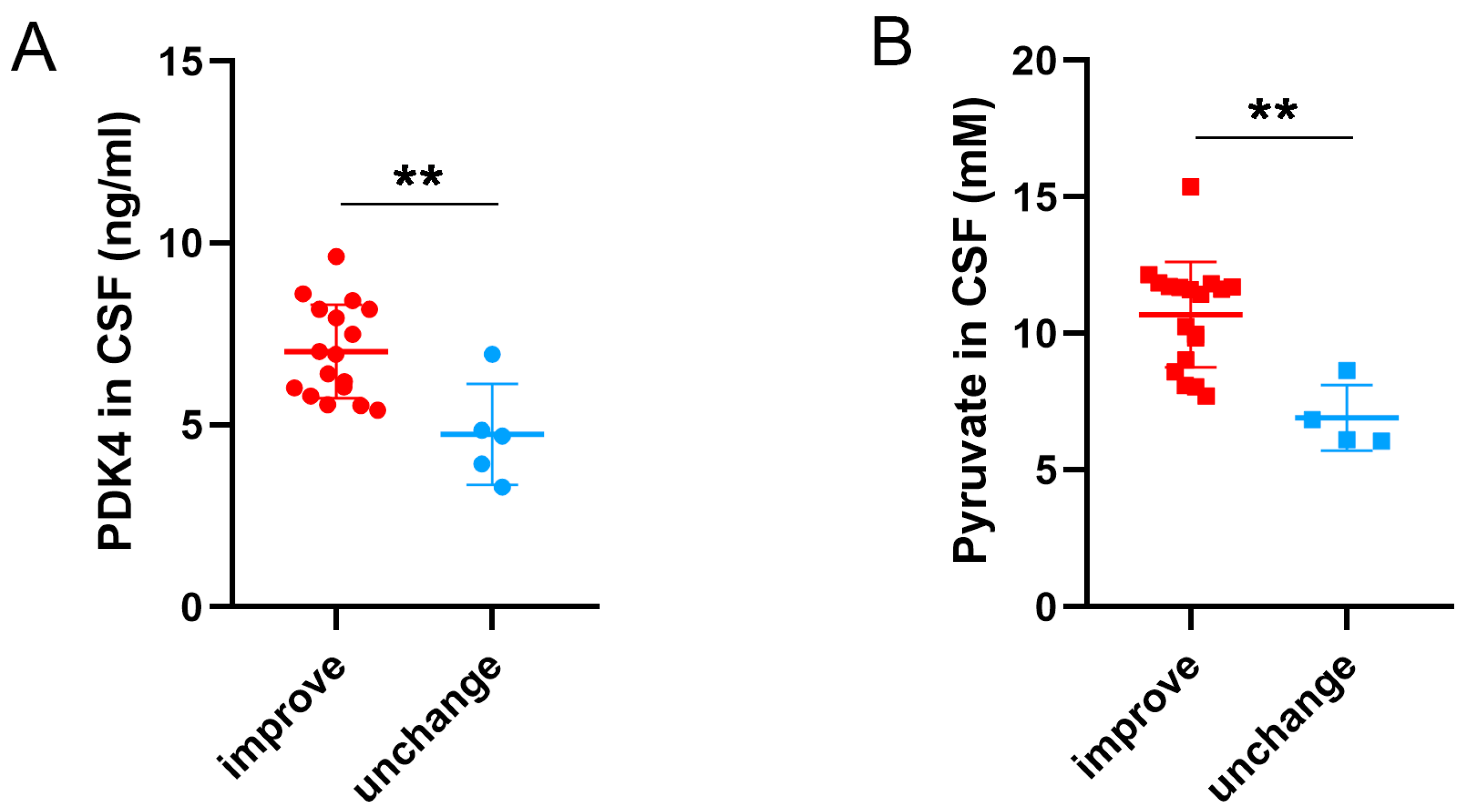
3.8. Different Levels of PDK4 and Pyruvate Were Related to Long-Term Outcomes of SAH Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Popov, K.; Kedishvili, N.; Zhao, Y.; Gudi, R.; Harris, R. Molecular cloning of the p45 subunit of pyruvate dehydrogenase kinase. J. Biol. Chem. 1994, 269, 29720–29724. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H.Y.; Song, S.Y.; Go, S.H.; Sohn, H.S.; Baik, S.; Soh, M.; Kim, K.; Kim, D.; Kim, H.; et al. Synergistic Oxygen Generation and Reactive Oxygen Species Scavenging by Manganese Ferrite/Ceria Co-decorated Nanoparticles for Rheumatoid Arthritis Treatment. ACS Nano 2019, 13, 3206–3217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; St Leger, R.; Fang, W. Pyruvate Accumulation Is the First Line of Cell Defense against Heat Stress in a Fungus. mBio 2017, 8, e01284–e01317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, R.L. Delayed neurological deterioration after subarachnoid haemorrhage. Nat. Rev. Neurol. 2014, 10, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Westermaier, T.; Jauss, A.; Eriskat, J.; Kunze, E.; Roosen, K. The temporal profile of cerebral blood flow and tissue metabolites indicates sustained metabolic depression after experimental subarachnoid hemorrhage. Neurosurgery 2011, 68, 223–230. [Google Scholar] [CrossRef]
- Tao, T.; Liu, G.J.; Shi, X.; Zhou, Y.; Lu, Y.; Gao, Y.Y.; Zhang, X.; Wang, H.; Wu, L.; Chen, C.; et al. DHEA Attenuates Microglial Activation via Induction of JMJD3 in Experimental Subarachnoid Haemorrhage. J. Neuroinflamm. 2019, 16, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolbright, B.L.; Rajendran, G.; Harris, R.A.; Taylor, J.A., 3rd. Metabolic Flexibility in Cancer: Targeting the Pyruvate Dehydrogenase Kinase:Pyruvate Dehydrogenase Axis. Mol. Cancer Ther. 2019, 18, 1673–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide Ion: Generation and Chemical Implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devasagayam, T.; Tilak, J.; Boloor, K.; Sane, K.; Ghaskadbi, S.; Lele, R. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physicians India 2004, 52, 4. [Google Scholar]
- Yagi, K.; Lidington, D.; Wan, H.; Fares, J.C.; Meissner, A.; Sumiyoshi, M.; Ai, J.; Foltz, W.D.; Nedospasov, S.A.; Offermanns, S.; et al. Therapeutically Targeting Tumor Necrosis Factor-alpha/Sphingosine-1-Phosphate Signaling Corrects Myogenic Reactivity in Subarachnoid Hemorrhage. Stroke 2015, 46, 2260–2270. [Google Scholar] [CrossRef] [Green Version]
- Cahill, J.; Zhang, J.H. Subarachnoid hemorrhage: Is it time for a new direction? Stroke 2009, 40, S86–S87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nechushtan, A.; Smith, C.; Hsu, Y.; Youle, R. Conformation of the Bax C-terminus regulates subcellular location and cell death. EMBO J. 1999, 18, 2330–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, F.; Wang, J.H.; Fan, W.J.; Meng, Y.T.; Li, M.M.; Li, T.T.; Cui, B.; Wang, H.; Zhao, Y.; An, F.; et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene 2018, 37, 1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Group NCRRGW. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Kumar, T.P.; Guddanti, T.; Yan, Y.; Blackburn, S.L.; McBride, D.W. Neurobehavioral Deficits After Subarachnoid Hemorrhage in Mice: Sensitivity Analysis and Development of a New Composite Score. J. Am. Heart Assoc. 2019, 8, e011699. [Google Scholar] [CrossRef] [Green Version]
- Sehba, F.A.; Hou, J.; Pluta, R.M.; Zhang, J.H. The importance of early brain injury after subarachnoid hemorrhage. Prog. Neurobiol. 2012, 97, 14–37. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Wang, Z.; Ding, Y.; Wang, X.; Lu, S.; Wang, C.; Hea, C.; Piao, M.; Chi, G.; Luo, Y.; et al. RIP1 and RIP3 contribute to shikonin-induced glycolysis suppression in glioma cells via increase of intracellular hydrogen peroxide. Cancer Lett. 2018, 42, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Jennett, B.; Bond, M. Assessment of outcome after severe brain damage. Lancet 1975, 1, 480–484. [Google Scholar] [CrossRef]
- Anerillas, C.; Abdelmohsen, K.; Gorospe, M. Regulation of senescence traits by MAPKs. Geroscience 2020, 42, 397–408. [Google Scholar] [CrossRef]
- Vlahopoulos, S.A. Aberrant control of NF-kappaB in cancer permits transcriptional and phenotypic plasticity, to curtail dependence on host tissue: Molecular mode. Cancer Biol. Med. 2017, 14, 254–270. [Google Scholar]
- Alnemri, E.; Livingston, D.; Nicholson, D.; Salvesen, G.; Thornberry, N.; Wong, W.; Yuan, Y. Human ICE/CED-3 protease nomenclature. Cell 1996, 87, 171. [Google Scholar] [CrossRef] [Green Version]
- McFate, T.; Mohyeldin, A.; Lu, H.; Thakar, J.; Henriques, J.; Halim, N.D.; Wu, H.; Schell, M.J.; Tsang, T.M.; Teahan, O.; et al. Pyruvate dehydrogenase complex activity controls metabolic and malignant phenotype in cancer cells. J. Biol. Chem. 2008, 283, 22700–22708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [Green Version]
- Bock, F.; Tait, S. Mitochondria as multifaceted regulators of cell death. Nature reviews. Mol. Cell Biol. 2020, 21, 85–100. [Google Scholar]
- Zhang, X.S.; Lu, Y.; Li, W.; Tao, T.; Peng, L.; Wang, W.H.; Gao, S.; Liu, C.; Zhuang, Z.; Xia, D.; et al. Astaxanthin ameliorates oxidative stress and neuronal apoptosis via SIRT1/NRF2/Prx2/ASK1/p38 after traumatic brain injury in mice. Br. J. Pharmacol. 2021, 178, 1114–1132. [Google Scholar] [CrossRef] [PubMed]
- Hattori, K.; Naguro, I.; Runchel, C.; Ichijo, H. The roles of ASK family proteins in stress responses and diseases. Cell Commun. Signal 2009, 7, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermaier, T.; Jauss, A.; Eriskat, J.; Kunze, E.; Roosen, K. Time-course of cerebral perfusion and tissue oxygenation in the first 6 h after experimental subarachnoid hemorrhage in rats. J. Cereb. Blood Flow Metab. 2009, 29, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgrove, B.D.; Gilbert, P.M.; Porpiglia, E.; Mourkioti, F.; Lee, S.P.; Corbel, S.Y.; Llewellyn, M.E.; Delp, S.L.; Blau, H.M. Rejuvenation of the muscle stem cell population restores strength to injured aged muscles. Nat. Med. 2014, 20, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierrat, B.; Simonen, M.; Cueto, M.; Mestan, J.; Ferrigno, P.; Heim, J. SH3GLB, a new endophilin-related protein family featuring an SH3 domain. Genomics 2001, 71, 222–234. [Google Scholar] [CrossRef]
- Rao, Y.; Gammon, S.; Zacharias, N.M.; Liu, T.; Salzillo, T.; Xi, Y.; Wang, J.; Bhattacharya, P.; Piwnica-Worms, D. Hyperpolarized [1-(13)C]pyruvate-to-[1-(13)C]lactate conversion is rate-limited by monocarboxylate transporter-1 in the plasma membrane. Proc. Natl. Acad. Sci. USA 2020, 117, 22378–22389. [Google Scholar] [CrossRef]
- Reed, L. Regulation of mammalian pyruvate dehydrogenase complex by a phosphorylation-dephosphorylation cycle. Curr. Top. Cell. Regul. 1981, 18, 95–106. [Google Scholar]
- Kinnaird, A.; Dromparis, P.; Saleme, B.; Gurtu, V.; Watson, K.; Paulin, R.; Zervopoulos, S.; Stenson, T.; Sutendra, G.; Pink, D.B.; et al. Metabolic Modulation of Clear-cell Renal Cell Carcinoma with Dichloroacetate, an Inhibitor of Pyruvate Dehydrogenase Kinase. Eur. Urol. 2016, 69, 734–744. [Google Scholar] [CrossRef]
- Westermaier, T.; Jauss, A.; Eriskat, J.; Kunze, E.; Roosen, K. Acute vasoconstriction: Decrease and recovery of cerebral blood flow after various intensities of experimental subarachnoid hemorrhage in rats. J. Neur. 2009, 110, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, G.; Jennett, B. Assessment of coma and impaired consciousness. A practical scale. Lancet 1974, 2, 81–84. [Google Scholar] [PubMed]
- Kark, R.; Rodriguez-Budelli, M.; Blass, J. Evidence for a primary defect of lipoamide dehydrogenase in Friedreich’s ataxia. Adv. Neurol. 1978, 21, 163–180. [Google Scholar] [PubMed]
- Liu, G.J.; Tao, T.; Wang, H.; Zhou, Y.; Gao, X.; Gao, Y.Y.; Hang, C.; Li, W. Functions of resolvin D1-ALX/FPR2 receptor interaction in the hemoglobin-induced microglial inflammatory response and neuronal injury. J. Neuroinflamm. 2020, 17, 239. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.A.; Turan, N.; Chau, M.; Pradilla, G. Inflammation, vasospasm, and brain injury after subarachnoid hemorrhage. Biomed. Res. Int. 2014, 2014, 384342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zhang, X.S.; Zhou, X.M.; Gao, Y.Y.; Chen, C.L.; Liu, J.P.; Ye, Z.; Zhang, Z.; Wu, L.; Li, W.; et al. Peroxiredoxin 1/2 protects brain against H2O2-induced apoptosis after subarachnoid hemorrhage. FASEB J. 2019, 33, 3051–3062. [Google Scholar] [CrossRef] [Green Version]
- Hop, J.W.; Rinkel, G.J.; Algra, A.; van Gijn, J. Case-fatality rates and functional outcome after subarachnoid hemorrhage: A systematic review. Stroke 1997, 28, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Segales, J.; Perdiguero, E.; Munoz-Canoves, P. Regulation of Muscle Stem Cell Functions: A Focus on the p38 MAPK Signaling Pathway. Front. Cell Dev. Biol. 2016, 4, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuette, A.J.; Barrow, D.L. Epidemiology and long-term mortality in subarachnoid hemorrhage. World Neurosurg. 2013, 80, 264–265. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.J.; Zhang, Q.R.; Gao, X.; Wang, H.; Tao, T.; Gao, Y.Y.; Zhou, Y.; Chen, X.; Li, W.; Hang, C. MiR-146a Ameliorates Hemoglobin-Induced Microglial Inflammatory Response via TLR4/IRAK1/TRAF6 Associated Pathways. Front. Neurosci. 2020, 14, 311. [Google Scholar] [CrossRef] [PubMed]
- Teunissen, C.; Petzold, A.; Bennett, J.; Berven, F.; Brundin, L.; Comabella, M.; Franciotta, D.; Frederiksen, J.L.; Fleming, J.O. A consensus protocol for the standardization of cerebrospinal fluid collection and biobanking. Neurology 2009, 73, 1914–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowker-Kinley, M.; Davis, W.; Wu, P.; Harris, R.; Popov, K. Evidence for existence of tissue-specific regulation of the mammalian pyruvate dehydrogenase complex. Biochem. J. 1998, 329, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Nygaard, G.; Di Paolo, J.A.; Hammaker, D.; Boyle, D.L.; Budas, G.; Notte, G.T.; Mikaelian, I.; Barry, V.; Firestein, G.S. Regulation and function of apoptosis signal-regulating kinase 1 in rheumatoid arthritis. Biochem. Pharmacol. 2018, 151, 282–290. [Google Scholar] [CrossRef]
- Gao, X.; Wu, L.Y. Early Overexpression of Pyruvate Dehydrogenase Kinase 4 Can Affect Oxidate Stress after Subarachnoid Hemorrhage. Res. Sq. 2021, in press. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case No. | Age (Years) | Gender | Hyper-Tension | Aneurysm | Days after Hemorrhage (d) | Initial GCS | Initial Hunt-Hess | Initial GOS | GOS (after 3 Months) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 63 | F | 1 | 1 | 1 | 15 | 3 | 5 | 5 |
| 2 | 72 | F | 1 | 1 | 1 | 15 | 1 | 5 | 5 |
| 3 | 54 | M | 0 | 1 | 1 | 3 | 4 | 1 | 1 |
| 4 | 75 | F | 0 | 1 | 1 | 8 | 4 | 3 | 3 |
| 5 | 65 | F | 1 | 1 | 1 | 15 | 2 | 4 | 5 |
| 6 | 69 | F | 1 | 0 | 1 | 15 | 2 | 5 | 5 |
| 7 | 69 | F | 0 | 1 | 1 | 15 | 3 | 5 | 5 |
| 8 | 48 | M | 0 | 1 | 1 | 14 | 3 | 3 | 4 |
| 9 | 64 | M | 1 | 1 | 2 | 14 | 3 | 4 | 5 |
| 10 | 76 | M | 0 | 1 | 2 | 7 | 4 | 3 | 5 |
| 11 | 40 | F | 1 | 1 | 2 | 15 | 3 | 3 | 5 |
| 12 | 50 | F | 0 | 1 | 2 | 9 | 4 | 3 | 5 |
| 13 | 62 | F | 1 | 1 | 2 | 5 | 4 | 1 | 1 |
| 14 | 49 | F | 0 | 1 | 2 | 12 | 3 | 4 | 5 |
| 15 | 56 | F | 1 | 0 | 2 | 15 | 1 | 4 | 5 |
| 16 | 66 | F | 1 | 1 | 2 | 5 | 3 | 5 | 5 |
| 17 | 63 | M | 0 | 0 | 2 | 14 | 2 | 5 | 5 |
| 18 | 55 | M | 1 | 1 | 2 | 15 | 1 | 5 | 5 |
| 19 | 45 | M | 1 | 1 | 2 | 15 | 3 | 5 | 5 |
| 20 | 63 | M | 1 | 0 | 2 | 15 | 2 | 5 | 5 |
| 21 | 73 | F | 1 | 1 | 2 | 14 | 3 | 3 | 3 |
| 22 | 45 | F | 1 | 1 | 2 | 15 | 2 | 5 | 5 |
| 23 | 47 | F | 0 | 1 | 3 | 15 | 2 | 4 | 4 |
| 24 | 49 | M | 0 | 1 | 3 | 15 | 2 | 4 | 5 |
| 25 | 53 | M | 0 | 0 | 3 | 15 | 2 | 4 | 5 |
| 26 | 54 | F | 0 | 1 | 3 | 15 | 2 | 4 | 5 |
| 27 | 72 | F | 0 | 1 | 3 | 14 | 1 | 4 | 5 |
| 28 | 55 | M | 0 | 1 | 3 | 15 | 2 | 4 | 5 |
| 29 | 63 | F | 1 | 1 | 3 | 15 | 3 | 4 | 5 |
| 30 | 68 | M | 1 | 1 | 3 | 14 | 3 | 4 | 5 |
| 31 | 57 | F | 1 | 1 | 3 | 14 | 2 | 4 | 5 |
| 32 | 65 | F | 1 | 1 | 3 | 14 | 2 | 3 | 5 |
| 33 | 57 | F | 1 | 1 | 3 | 15 | 1 | 4 | 5 |
| 34 | 52 | M | 1 | 1 | 3 | 15 | 1 | 5 | 5 |
| Target Gene | Forward (5′ to 3′) | Reverse (5′ to 3′) |
|---|---|---|
| PDK1 | GTTCAGTACTTTTTGGATCGGTTC | TCGACTACATCACAGTTTGGATTT |
| PDK2 | TGGACCGCTTCTACCTCAG | TCTTTCACCACATCAGACACG |
| PDK3 | TGACCTAGGTGGTGGAGTCCCA | ACCAAATCCAGCCAAGGGAGCA |
| PDK4 | GAACACCCCTTCCGTCCAGCT | TGTGCCATCGTAGGGACCACA |
| BAX | GACACCTGAGCTGACCTTGG | GAGGAAGTCCAGTGTCCAGC |
| BCL2 | TATGATAACCGGGAGATCGTGATC | GTGCAGATGCCGGTTCAGGTACTC |
| GAPDH | TGTGAAGCTCATTTCCTGGTA | TTACTCCTTGGAGGCCATGT |
| Groups | Mortality Rate | Excluded |
|---|---|---|
| Stage 1 | ||
| Sham | 0 (0/5) | 0 |
| SAH 6 h | 11% (1–9) | 3 |
| SAH 24 h | 22% (2–9) | 2 |
| SAH 2 d | 22% (2–9) | 2 |
| SAH 3 d | 11% (1–9) | 3 |
| SAD 7 d | 22% (2–9) | 2 |
| Stage 2 | ||
| sham for IF | 0 (0/6) | 0 |
| SAH 2 d for IF | 22% (2–9) | 1 |
| Total | ||
| Sham | 0 (0–11) | 0 |
| SAH | 18.5% (10–54) | 13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Gao, Y.-Y.; Wu, L.-Y.; Peng, Z.; Liu, X.-Z.; Chen, X.-X.; Gao, S.; Zhang, H.-S.; Lu, Y.; Hang, C.-H.; et al. High Expression of PDK4 Could Play a Potentially Protective Role by Attenuating Oxidative Stress after Subarachnoid Hemorrhage. J. Clin. Med. 2022, 11, 3974. https://doi.org/10.3390/jcm11143974
Gao X, Gao Y-Y, Wu L-Y, Peng Z, Liu X-Z, Chen X-X, Gao S, Zhang H-S, Lu Y, Hang C-H, et al. High Expression of PDK4 Could Play a Potentially Protective Role by Attenuating Oxidative Stress after Subarachnoid Hemorrhage. Journal of Clinical Medicine. 2022; 11(14):3974. https://doi.org/10.3390/jcm11143974
Chicago/Turabian StyleGao, Xuan, Yong-Yue Gao, Ling-Yun Wu, Zheng Peng, Xun-Zhi Liu, Xiang-Xin Chen, Sen Gao, Hua-Sheng Zhang, Yue Lu, Chun-Hua Hang, and et al. 2022. "High Expression of PDK4 Could Play a Potentially Protective Role by Attenuating Oxidative Stress after Subarachnoid Hemorrhage" Journal of Clinical Medicine 11, no. 14: 3974. https://doi.org/10.3390/jcm11143974