Enhancement of Antiviral CD8+ T-Cell Responses and Complete Remission of Metastatic Melanoma in an HIV-1-Infected Subject Treated with Pembrolizumab

 ,
,  ,
, 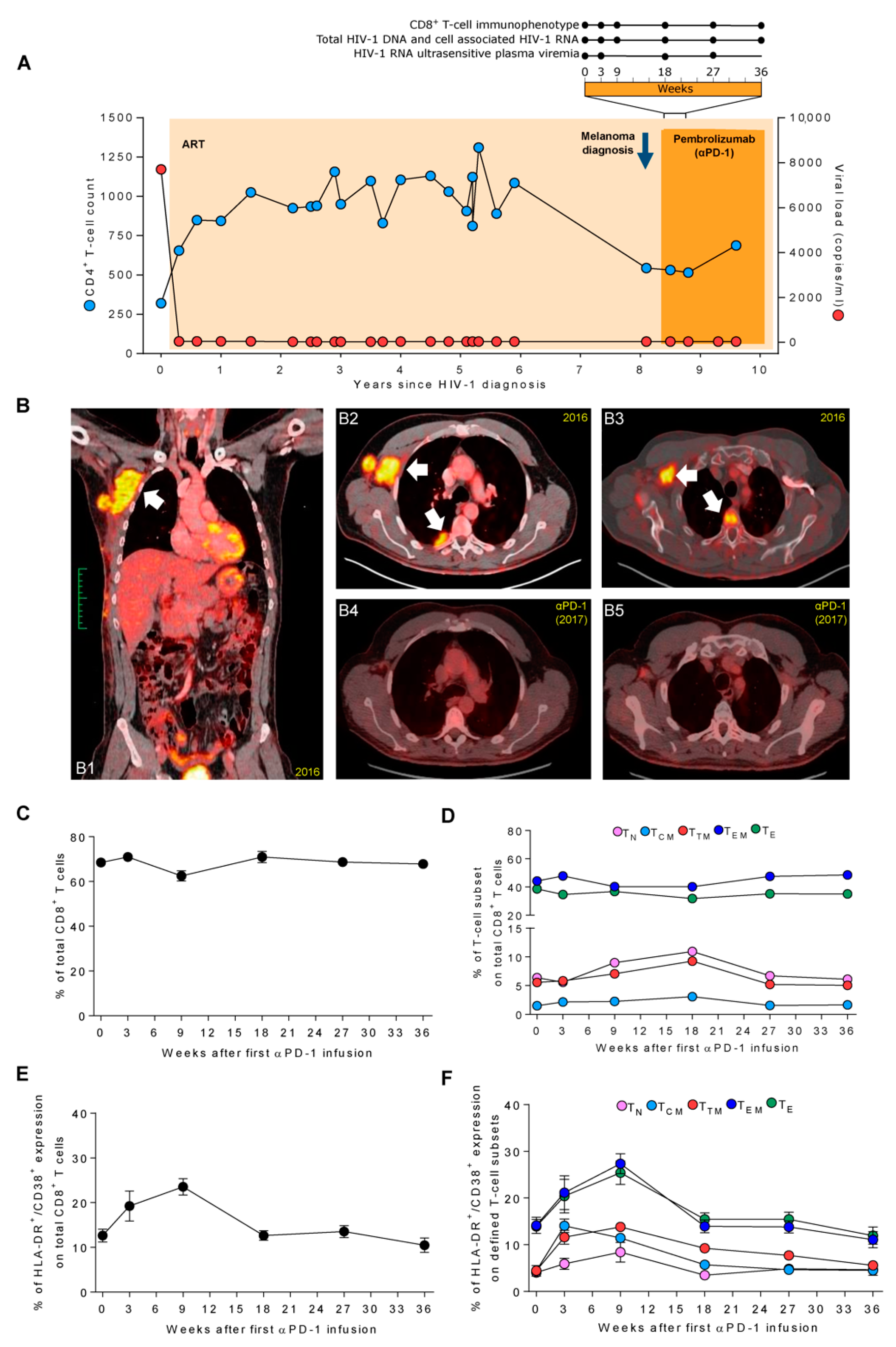{kind=link}
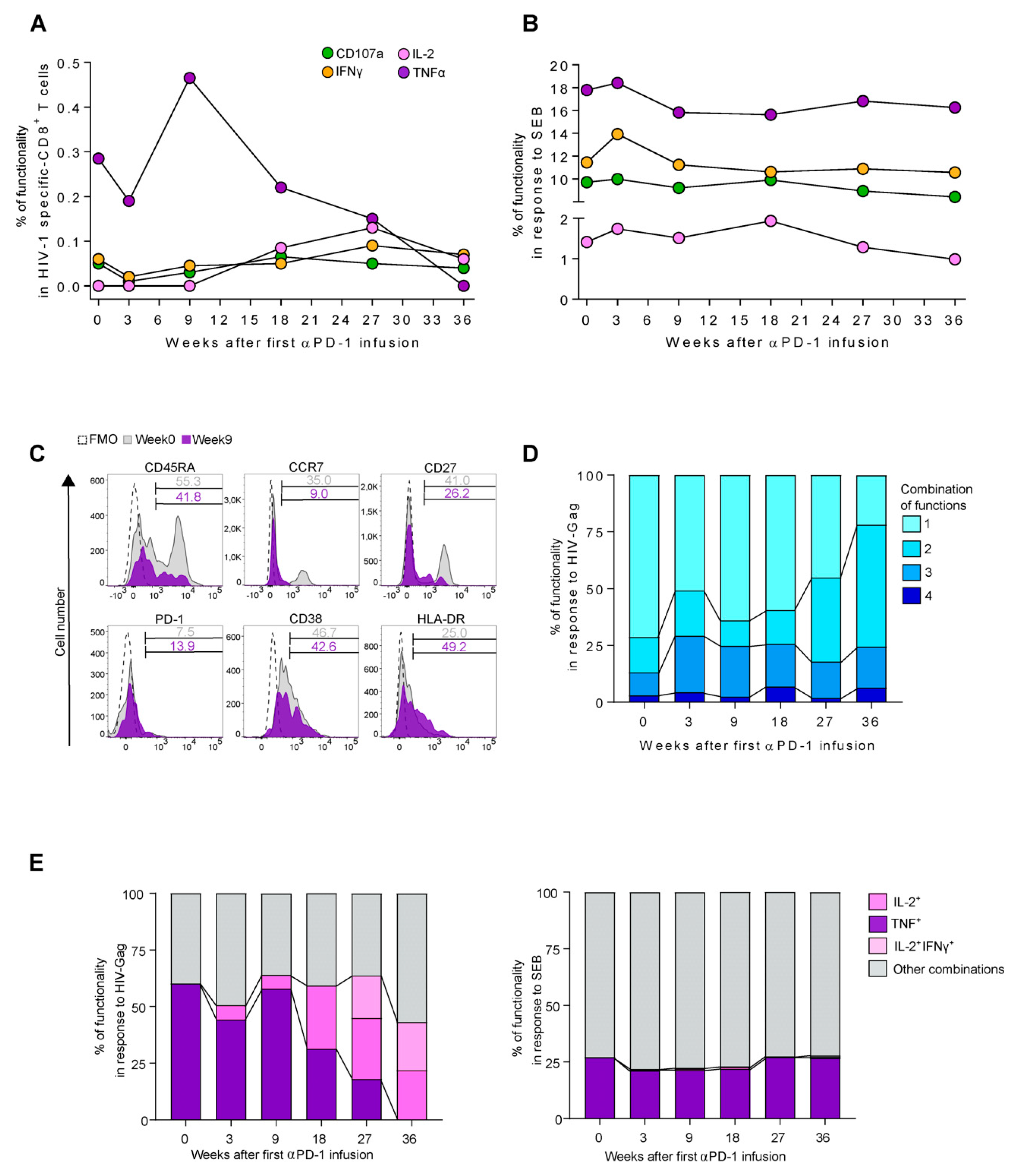{kind=link}
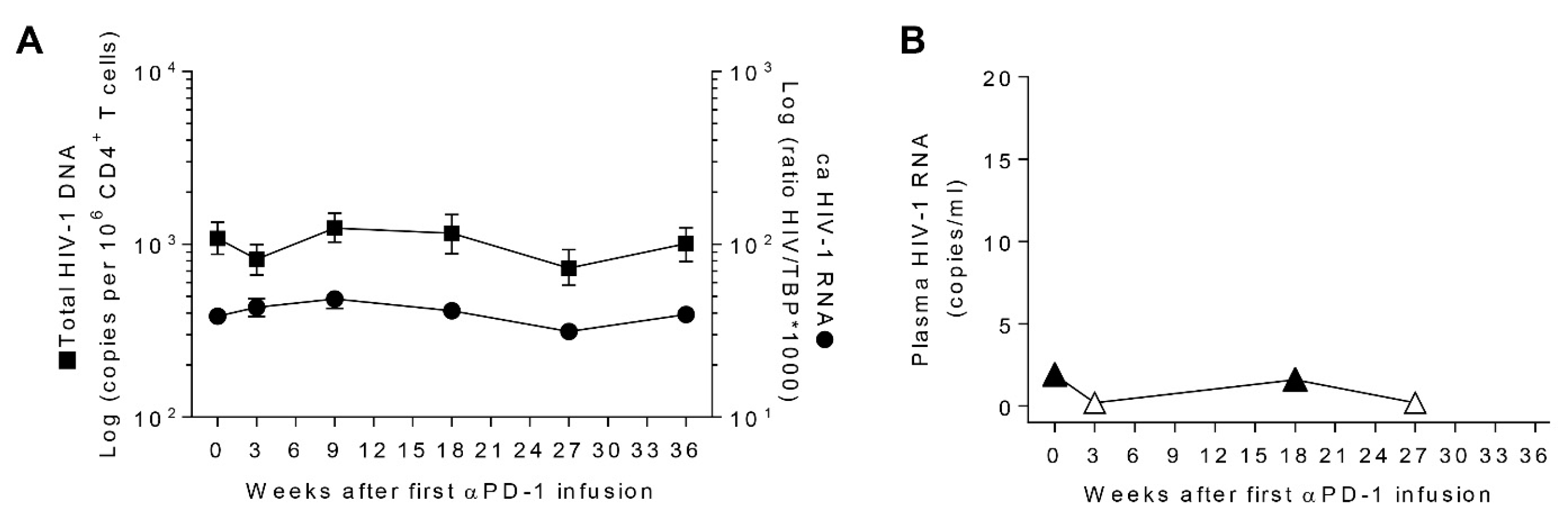{kind=link}
Abstract
:1. Introduction
Case Report
2. Experimental Section
2.1. Immune Phenotype of Total CD8+ and HIV-1 Specific-CD8+ T Cells
2.2. Total HIV-1 DNA, Cell-Associated HIV-1 RNA, and Ultrasensitive Viral Load
3. Results
3.1. Pembrolizumab Administration Induces a Marked and Transient Increase of HLA-DR+/CD38+ Expression in TEM and TE CD8+ T Cells
3.2. Enhancement of TNF Production in HIV-1 Specific-CD8+ T-Cell Responses after Pembrolizumab Initiation
3.3. Functional Switch from TNF to IL-2 Profile in HIV-1 Specific-CD8+ T-Cell Responses after Pembrolizumab Follow Up
3.4. Transient Decays of Total HIV-1 DNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ohaegbulam, K.C.; Assal, A.; Lazar-Molnar, E.; Yao, Y.; Zang, X. Human cancer immunotherapy with antibodies to the PD-1 and PD-L1 pathway. Trends. Mol. Med. 2015, 21, 24–33. [Google Scholar] [CrossRef]
- Hamid, O.; Robert, C.; Daud, A.; Hodi, F.S.; Hwu, W.J.; Kefford, R.; Wolchok, J.D.; Hersey, P.; Joseph, R.; Weber, J.S.; et al. Five-year survival outcomes for patients with advanced melanoma treated with pembrolizumab in KEYNOTE-001. Ann. Oncol. 2019, 30, 582–588. [Google Scholar] [CrossRef]
- Trautmann, L.; Janbazian, L.; Chomont, N.; Said, E.A.; Gimmig, S.; Bessette, B.; Boulassel, M.-R.; Delwart, E.; Sepulveda, H.; Balderas, R.S.; et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat. Med. 2006, 12, 1198. [Google Scholar] [CrossRef]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.J.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350. [Google Scholar] [CrossRef]
- Hatano, H.; Jain, V.; Hunt, P.W.; Lee, T-H.; Sinclair, E.; Do, T.D.; Hoh, R.; Martin, J.N.; Mccune, J.M.; Hecht, F.; et al. Cell-based measures of viral persistence are associated with immune activation and programmed cell death protein 1 (PD-1)-expressing CD4+ T cells. J. Infect. Dis. 2013, 208, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Fromentin, R.; Bakeman, W.; Lawani, M.B.; Khoury, G.; Hartogensis, W.; DaFonseca, S.; Killian, M.; Epling, L.; Hoh, R.; Sinclair, E.; et al. CD4+ T Cells Expressing PD-1, TIGIT and LAG-3 Contribute to HIV Persistence during ART. PLOS Pathog. 2016, 12, e1005761. [Google Scholar] [CrossRef] [PubMed]
- DaFonseca, S.; Chomont, N.; El Far, M.; Boulassel, R.; Routy, J.; Sékaly, R. Purging the HIV-1 reservoir through the disruption of the PD-1 pathway. J. Int. AIDS Soc. 2010, 13, O15. [Google Scholar] [CrossRef]
- Fromentin, R.; DaFonseca, S.; Costiniuk, C.T.; El-Far, M.; Procopio, F.A.; Hechy, F.M.; Hoh, R.; Deeks, S.G.; Hazuda, D.J.; Lewin, S.R.; et al. PD-1 blockade potentiates HIV latency reversal ex vivo in CD4+ T cells from ART-suppressed individuals. Nat. Commun. 2019, 10, 814. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G. Shock and kill. Nature 2012, 487, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Uldrick, T.S.; Gonçalves, P.H.; Abdul-Hay, M.; Claeys, A.J.; Emu, B.; Ernstoff, M.S.; Fling, S.P.; Fong, L.; Kaiser, J.C.; Lacroix, A.M.; et al. Assessment of the Safety of Pembrolizumab in Patients With HIV and Advanced Cancer—A Phase 1 Study. JAMA Oncol. 2019, 5, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.R.; Kim, C. Safety and Efficacy of Immune Checkpoint Inhibitor Therapy in Patients with HIV Infection and Advanced-Stage Cancer. JAMA Oncol. 2019, 5, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Heppt, M.V.; Schlaak, M.; Eigentler, T.K.; Kähler, K.C.; Kiecker, F.; Loquai, C.; Meier, F.; Tomsitz, D.; Brenner, N.; Niesert, A.C.; et al. Checkpoint blockade for metastatic melanoma and Merkel cell carcinoma in HIV-positive patients. Ann. Oncol. 2017, 28, 3104–3106. [Google Scholar] [CrossRef] [PubMed]
- Davar, D.; Wilson, M.; Pruckner, C.; Kirkwood, J.M. PD-1 Blockade in Advanced Melanoma in Patients with Hepatitis C and/or HIV. Case Rep Oncol. Med. 2015, 2015, 737389. [Google Scholar] [CrossRef] [PubMed]
- Scully, E.P.; Rutishauser, R.L.; Simoneau, C.R.; Delagrèverie, H.; Euler, Z.; Thanh, C.; Li, J.Z.; Hartig, H.; Bakkour, S.; Busch, M.; et al. Inconsistent HIV reservoir dynamics and immune responses following anti-PD-1 therapy in cancer patients with HIV infection. Ann. Oncol. 2018, 29, 2141–2142. [Google Scholar] [CrossRef] [PubMed]
- Le Garff, G.; Samri, A.; Lambert-Niclot, S.; Even, S.; Lavolé, A.; Cadranel, J.; Spano, J.P.; Autran, B.; Marcelin, A.G.; Guihot, A. Transient HIV-specific T cells increase and inflammation in an HIV-infected patient treated with nivolumab. AIDS 2017, 31, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Gay, C.L.; Bosch, R.J.; Ritz, J.; Hataye, J.M.; Aga, E.; Tressler, R.L.; Mason, S.W.; Hwang, C.K.; Grasela, D.M.; Ray, N.; et al. Clinical Trial of the Anti-PD-L1 Antibody BMS-936559 in HIV-1 Infected Participants on Suppressive Antiretroviral Therapy. J. Infect. Dis. 2017, 215, 1725–1733. [Google Scholar] [CrossRef] [PubMed]
- Wightman, F.; Solomon, A.; Kumar, S.S.; Urriola, N.; Gallagher, K.; Hiener, B.; Palmer, S.; Mcneil, C.; Garsia, R.; Lewin, S.R. Effect of ipilimumab on the HIV reservoir in an HIV-infected individual with metastatic melanoma. AIDS 2015, 29, 504. [Google Scholar] [CrossRef] [PubMed]
- Guihot, A.; Marcelin, A.-G.; Massiani, M.-A.; Samri, A.; Soulié, C.; Autran, B.; Spano, J.P. Drastic decrease of the HIV reservoir in a patient treated with nivolumab for lung cancer. Ann. Oncol. 2018, 29, 517–518. [Google Scholar] [CrossRef]
- Breton, G.; Chomont, N.; Takata, H.; Fromentin, R.; Ahlers, J.; Filali-Mouhim, A.; Riou, C.; Boulassel, M.R.; Routy, J.P.; Yassine-Diab, B.; et al. Programmed death-1 is a marker for abnormal distribution of naive/memory T cell subsets in HIV-1 infection. J. Immunol. 2013, 191, 2194–2204. [Google Scholar] [CrossRef]
- Roederer, M.; Nozzi, J.L.; Nason, M.C. SPICE: Exploration and analysis of post-cytometric complex multivariate datasets. Cytom. Part A 2011, 79, 167–174. [Google Scholar] [CrossRef]
- Martínez-Bonet, M.; Puertas, M.C.; Fortuny, C.; Ouchi, D.; Mellado, M.J.; Rojo, P.; Noguera-Julian, A.; Muñoz-Fernández, M.A.; Martinez-Picado, J. Establishment and Replenishment of the Viral Reservoir in Perinatally HIV-1-infected Children Initiating Very Early Antiretroviral Therapy. Clin. Infect. Dis. 2015, 61, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Puertas, M.C.; Salgado, M.; Morón-López, S.; Ouchi, D.; Muñoz-Moreno, J.A.; Moltó, J.; Clotet, B.; Martinez-Picado, J. Effect of lithium on HIV-1 expression and proviral reservoir size in the CD4+ T cells of antiretroviral therapy suppressed patients. AIDS 2014, 28, 2157–2159. [Google Scholar] [CrossRef] [PubMed]
- Amendola, A.; Bloisi, M.; Marsella, P.; Sabatini, R.; Bibbò, A.; Angeletti, C.; Capobianchi, M.R. Standardization and performance evaluation of ‘modified’ and ‘ultrasensitive’ versions of the Abbott RealTime HIV-1 assay, adapted to quantify minimal residual viremia. J. Clin. Virol. 2011, 52, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.N.; Hultin, L.E.; Mitsuyasu, R.T.; Matud, J.L.; Hausner, M.A.; Bockstoce, D.; Chou, C.C.; O’Rourke, S.; Taylor, J.M.; Giorgi, J.V. Circulating HIV-specific CD8+ cytotoxic T cells express CD38 and HLA-DR antigens. J. Immunol. 1993, 150, 3070–3079. [Google Scholar]
- Kestens, L.; Vanham, G.; Gigase, P.; Young, G.; Hannet, I.; Vanlangendonck, F.; Hulstaert, F.; Bach, B. Expression of activation antigens, HLA-DR and CD38, on CD8 lymphocytes during HIV-1 infection. AIDS 1992, 6, 793–798. [Google Scholar] [CrossRef]
- Ruiz, A.; Blanch-Lombarte, O.; Jimenez-Moyano, E.; Ouchi, D.; Mothe, B.; Peña, R.; Galvez, C.; Genescà, M.; Martinez-Picado, J.; Goulder, P. Antigen Production After Latency Reversal and Expression of Inhibitory Receptors in CD8+ T Cells Limit the Killing of HIV-1 Reactivated Cells. Front. Immunol. 2019, 9, 3162. [Google Scholar] [CrossRef]
- Velu, V.; Titanji, K.; Zhu, B.; Husain, S.; Pladevega, A.; Lai, L.; Vanderford, T.H.; Chennareddi, L.; Silvestri, G.; Freeman, G.J.; et al. Enhancing SIV-specific immunity in vivo by PD-1 blockade. Nature 2009, 458, 206. [Google Scholar] [CrossRef]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor β, and TNF-α: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef]
- Lenardo, M.; Chan, F.K-M.; Hornung, F.; McFarland, H.; Siegel, R.; Wang, J.; Zheng, L. Mature T lymphocyte apoptosis—immune regulation in a dynamic and unpredictable antigenic environment. Annu. Rev. Immunol. 1999, 17, 221–253. [Google Scholar] [CrossRef]
- Gaffen, S.; Liu, K. Overview of interleukin-2 function, production and clinical applications. Cytokine 2004, 28, 109–123. [Google Scholar] [CrossRef]
- Aust, S.; Felix, S.; Auer, K.; Bachmayr-Heyda, A.; Kenner, L.; Dekan, S.; Meier, S.M.; Gerner, C.; Grimm, C.; Pils, D. Absence of PD-L1 on tumor cells is associated with reduced MHC I expression and PD-L1 expression increases in recurrent serous ovarian cancer. Sci. Rep. 2017, 7, 42929. [Google Scholar] [CrossRef] [PubMed]
- Uldrick, T.S.; Fling, S.; Scott, V. Pembrolizumab induces HIV latency reversal in HIV+ individuals on ART with cancer. In Proceedings of the Conference on Retroviruses and Opportunistic Infections (CROI), Seattle, WA, USA, 4–7 March 2019; p. 27. Available online: https://www.croiconference.org/abstracts/search-abstracts/ (accessed on 26 November 2019).
- Banga, R.; Procopio, F.A.; Noto, A.; Pollakis, G.; Cavassini, M.; Ohmiti, K.; Corpataux, J.M.; de Leval, L.; Pantaleo, G.; Perreau, M. PD-1+ and follicular helper T cells are responsible for persistent HIV-1 transcription in treated aviremic individuals. Nat. Med. 2016, 22, 754. [Google Scholar] [CrossRef] [PubMed]
- González-Cao, M.; Moran, T.; Dalmau, J.; Garcia-Corbacho, J.; Bernabé, R.; Juan, O.; de Castro, J.; Blanco, R.; Meyerhand, A.; Blanco, J.; et al. Phase II study of durvalumab (MEDI4736) in cancer patients HIV-1-infected. J. Clin. Oncol. 2019, 37, 2501. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanch-Lombarte, O.; Gálvez, C.; Revollo, B.; Jiménez-Moyano, E.; Llibre, J.M.; Manzano, J.L.; Boada, A.; Dalmau, J.; E. Speiser, D.; Clotet, B.; et al. Enhancement of Antiviral CD8+ T-Cell Responses and Complete Remission of Metastatic Melanoma in an HIV-1-Infected Subject Treated with Pembrolizumab. J. Clin. Med. 2019, 8, 2089. https://doi.org/10.3390/jcm8122089
Blanch-Lombarte O, Gálvez C, Revollo B, Jiménez-Moyano E, Llibre JM, Manzano JL, Boada A, Dalmau J, E. Speiser D, Clotet B, et al. Enhancement of Antiviral CD8+ T-Cell Responses and Complete Remission of Metastatic Melanoma in an HIV-1-Infected Subject Treated with Pembrolizumab. Journal of Clinical Medicine. 2019; 8(12):2089. https://doi.org/10.3390/jcm8122089
Chicago/Turabian StyleBlanch-Lombarte, Oscar, Cristina Gálvez, Boris Revollo, Esther Jiménez-Moyano, Josep M. Llibre, José Luís Manzano, Aram Boada, Judith Dalmau, Daniel E. Speiser, Bonaventura Clotet, and et al. 2019. "Enhancement of Antiviral CD8+ T-Cell Responses and Complete Remission of Metastatic Melanoma in an HIV-1-Infected Subject Treated with Pembrolizumab" Journal of Clinical Medicine 8, no. 12: 2089. https://doi.org/10.3390/jcm8122089