
Ethylene Promotes Expression of the Appressorium- and Pathogenicity-Related Genes via GPCR- and MAPK-Dependent Manners in Colletotrichum gloeosporioides



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Pathogen and Host Plant Materials
2.2. Ethylene Treatment and Sample Preparation
2.3. Total RNA Extraction
2.4. Library Preparation and RNA Sequencing
2.5. Transcriptomic Data Analysis
2.6. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
2.7. Fungal Transformation
2.8. Fluorescence-Based Reporter Assay
2.9. Appressorial Formation and Pathogenicity Assay
2.10. Staining Analysis for Reactive Oxygen Species in Plant Leaves
2.11. FDA-PI Dual Fluorescence Staining
2.12. Calcofluor White-EosinY Double Fluorescence Staining
2.13. Statistical Analysis
3. Results
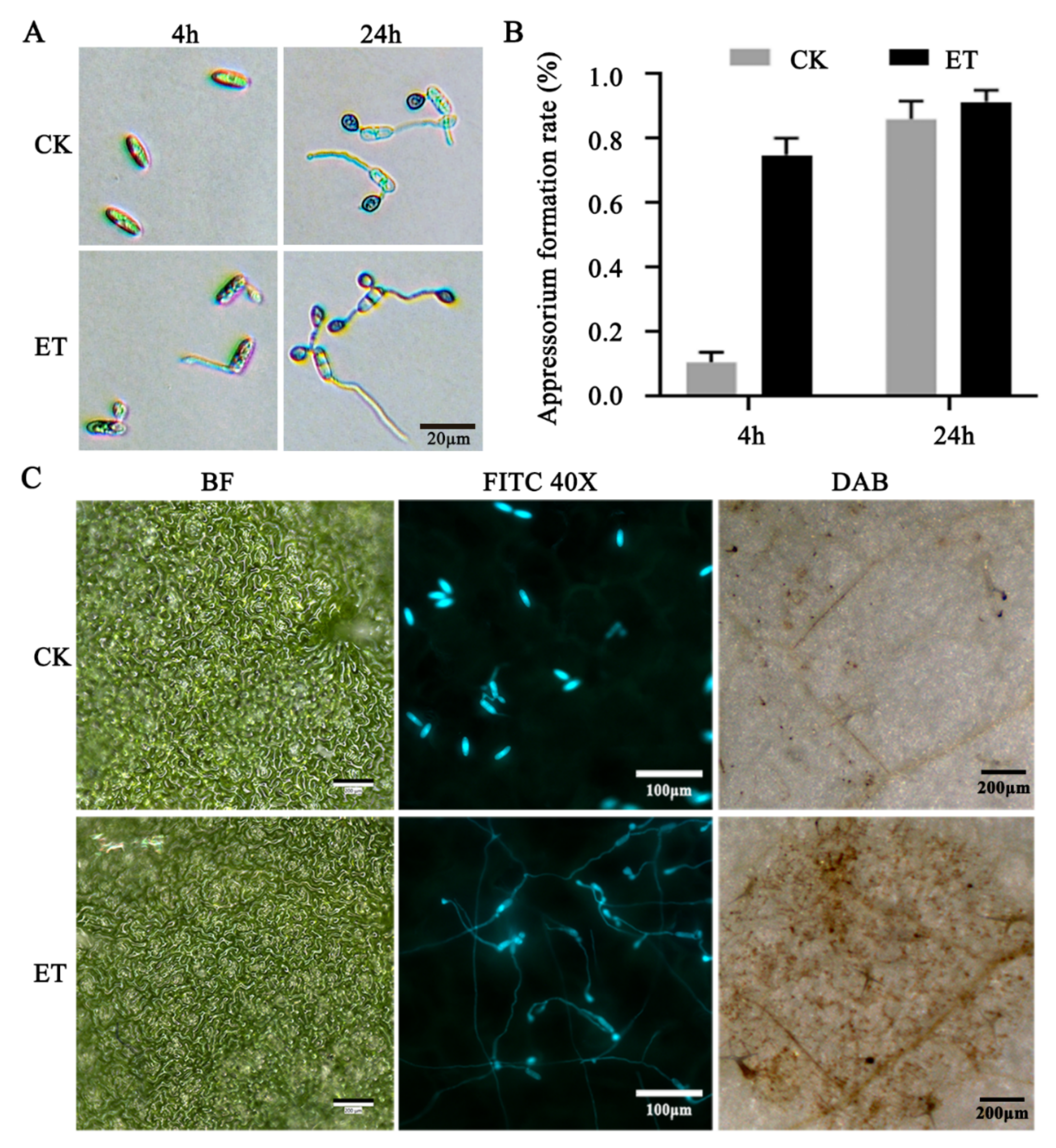
3.1. Ethylene Accelerated Conidial Germination, Appressorium Formation, and Pathogenicity of C. gloeosporioides
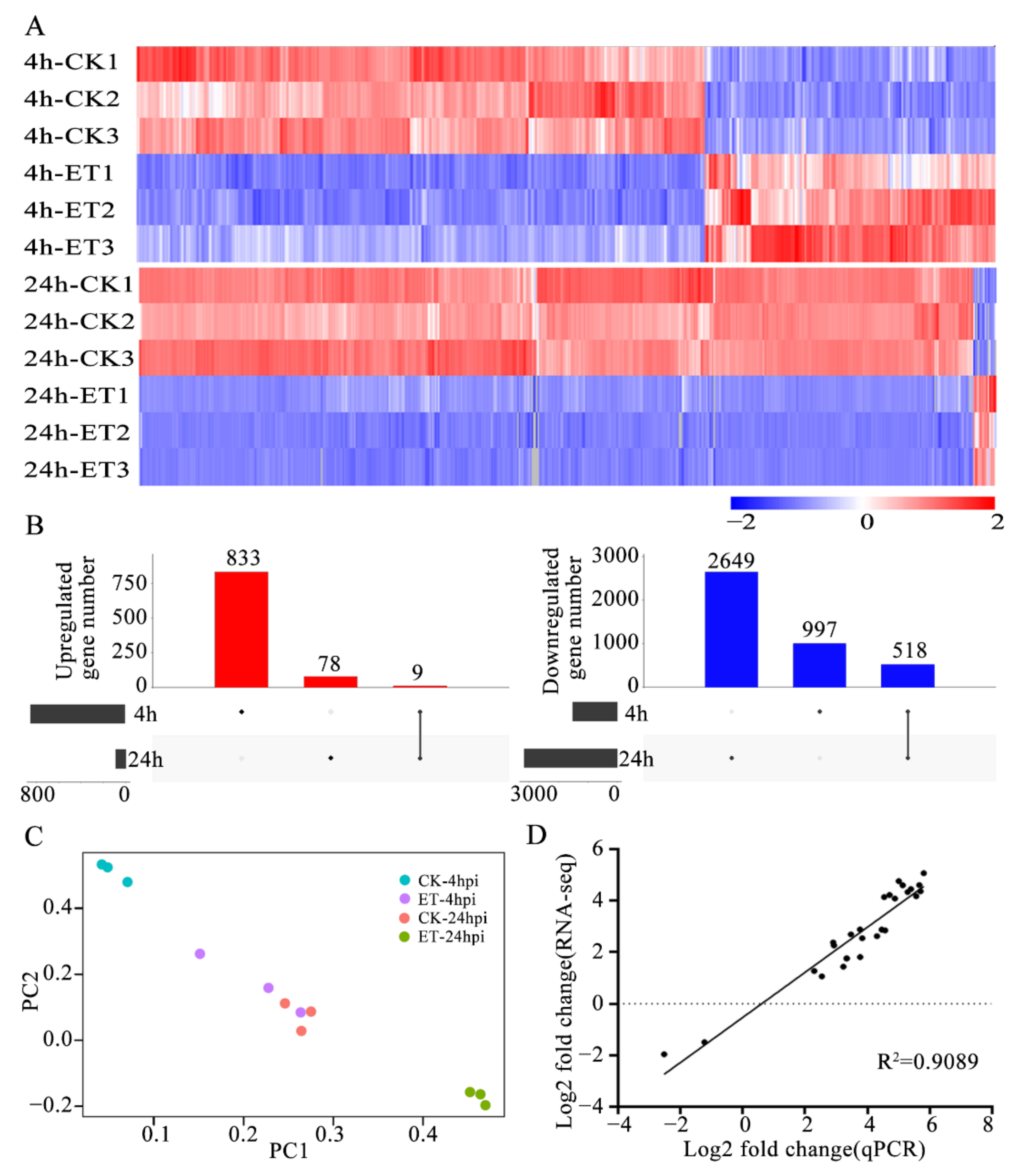
3.2. ET Treatment Altered Transcript Levels of a Large Set of Genes in Germinating Conidia of C. gloeosporioides
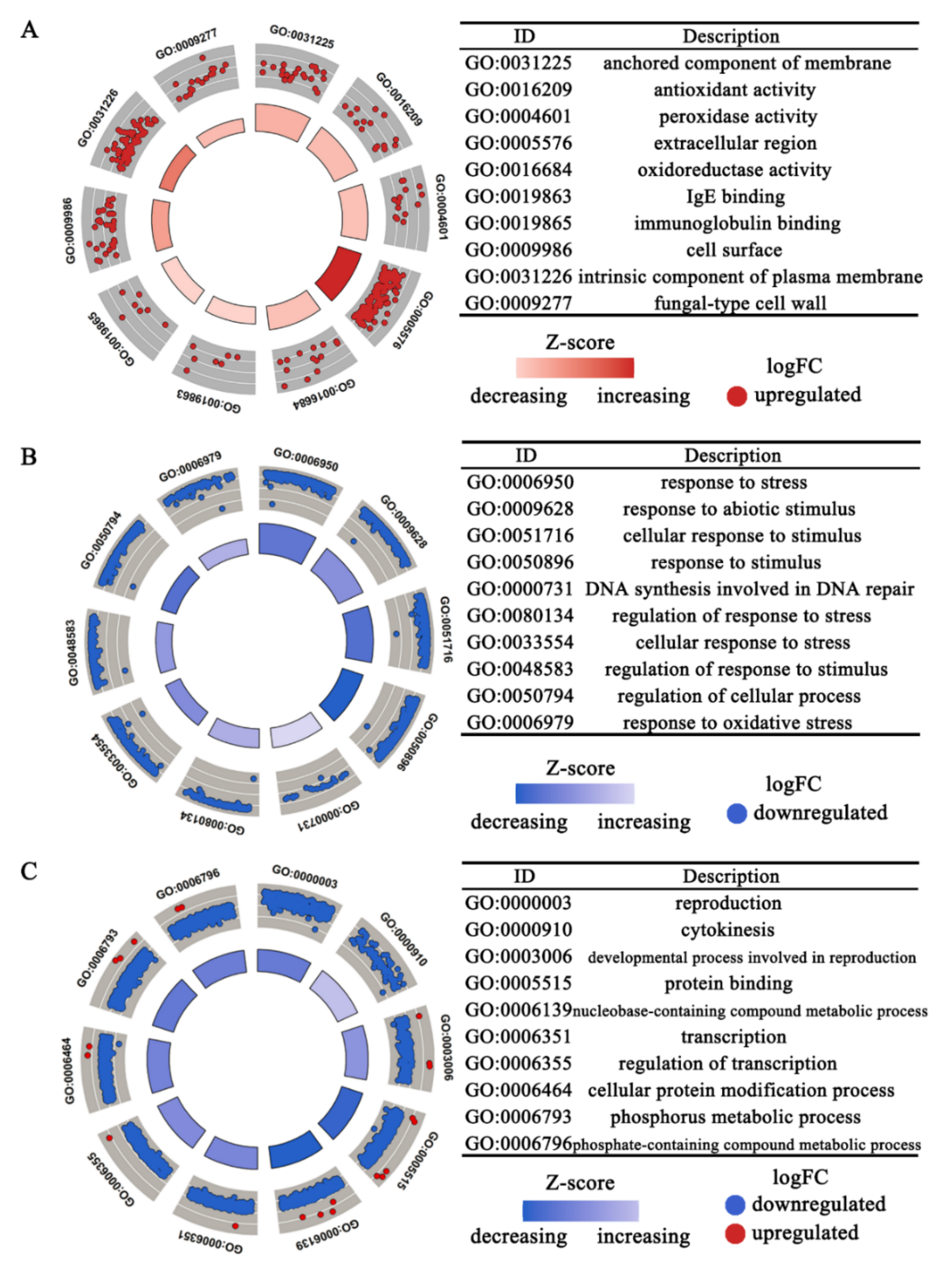
3.3. Functional Enrichment Analysis of Differentially Expressed Genes
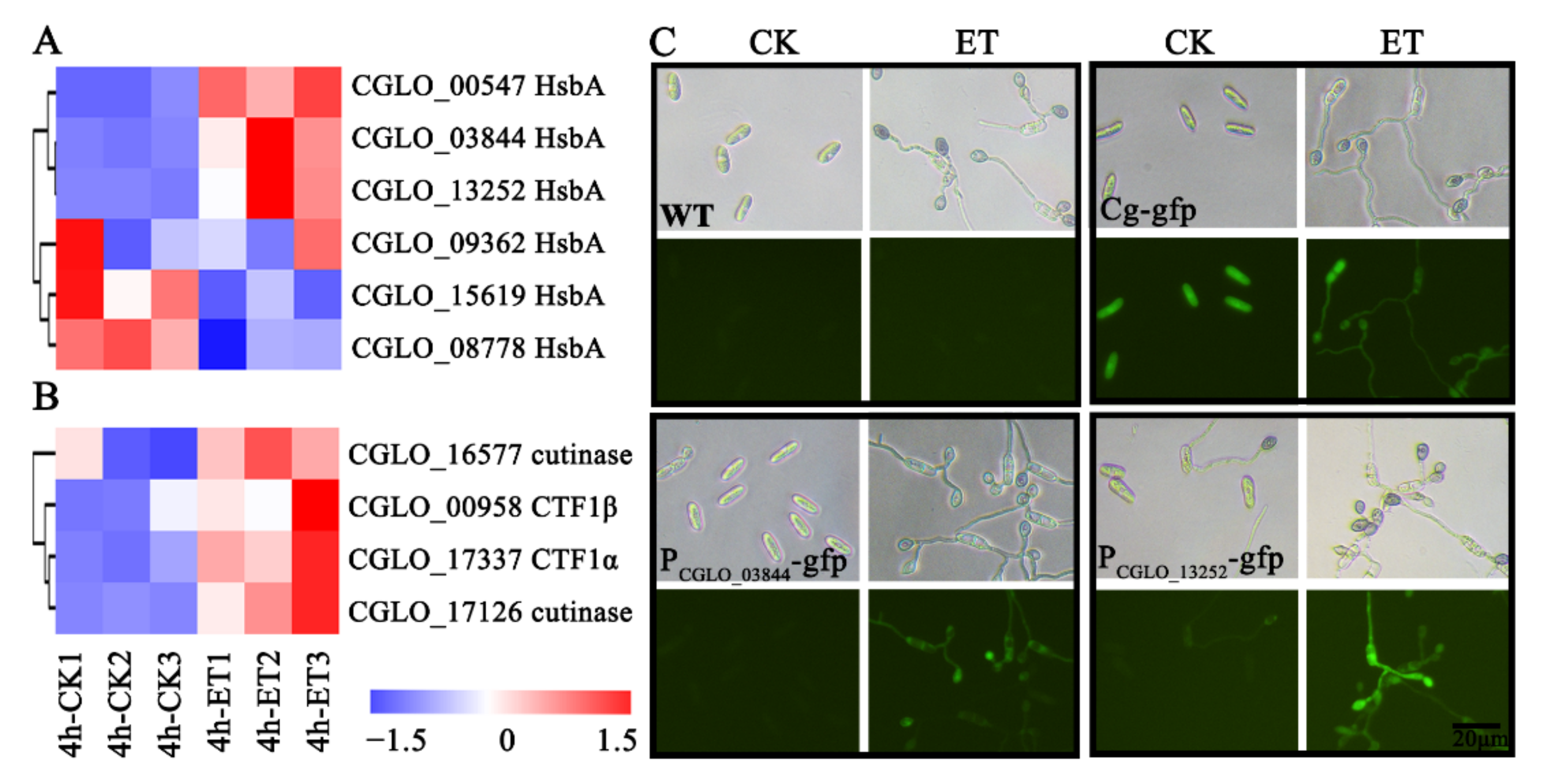
3.4. Hydrophobic Surface Binding Protein A and Cutinase-Related Genes Were Induced by ET Treatment at the Early Germinating Stage
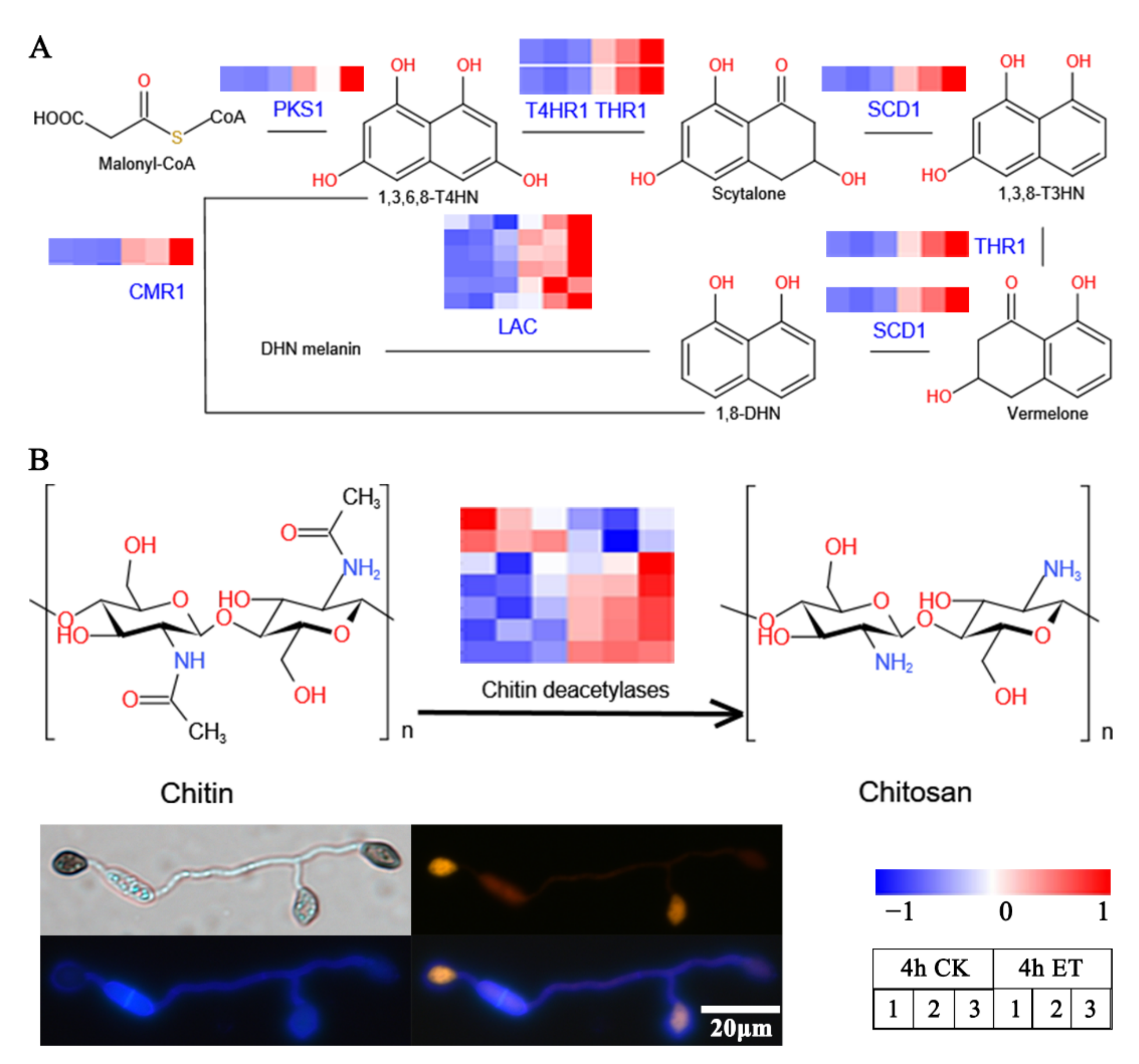
3.5. ET Promoted the Melanin Synthesis and Chitin Deacetylation at the Appressorial Cell Wall
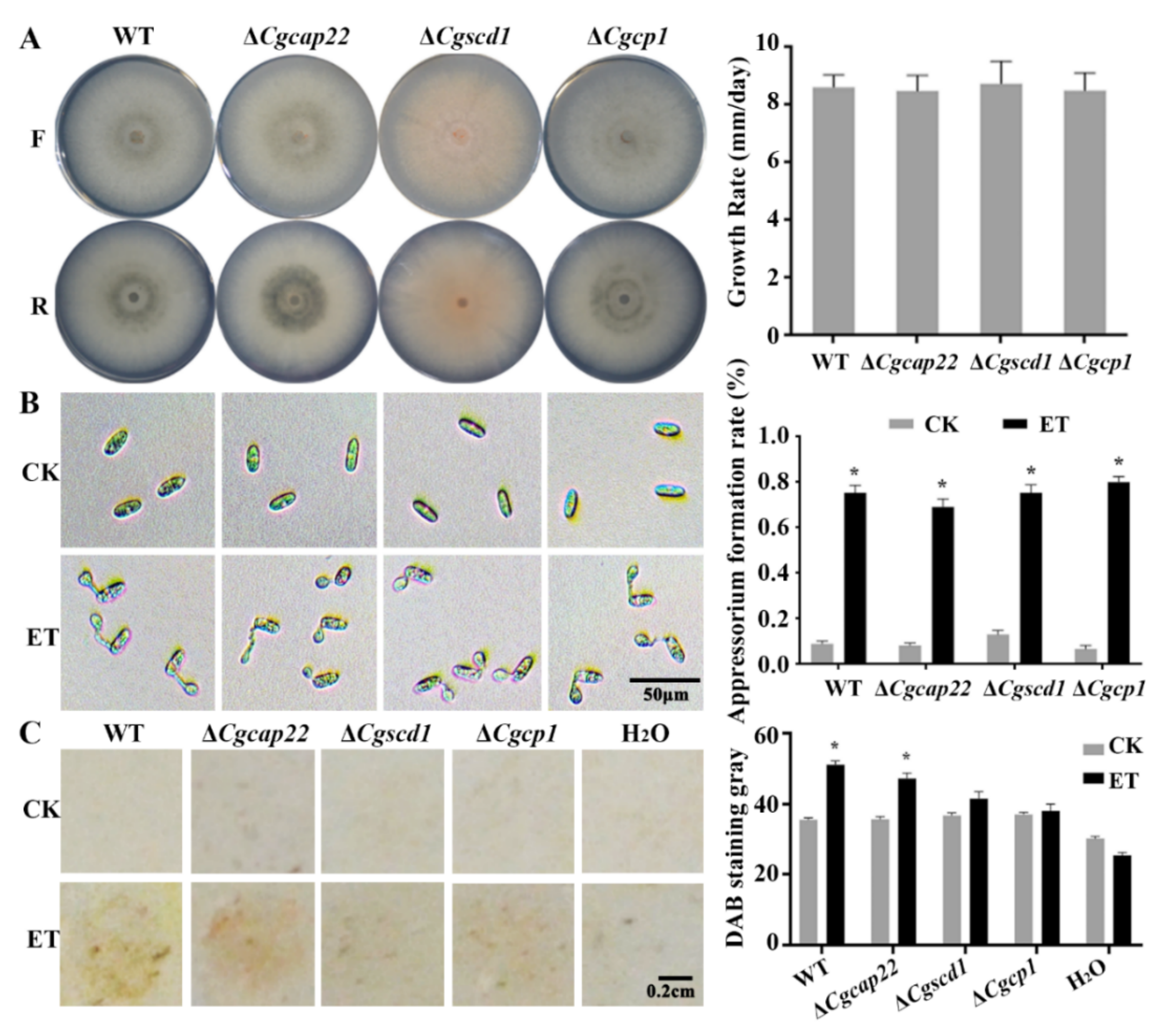
3.6. Pathogenicity Enhancement of C. gloeosporioides in Response to ET Was Partially Dependent on the Upregulation of Melanin Synthase and Effector Genes
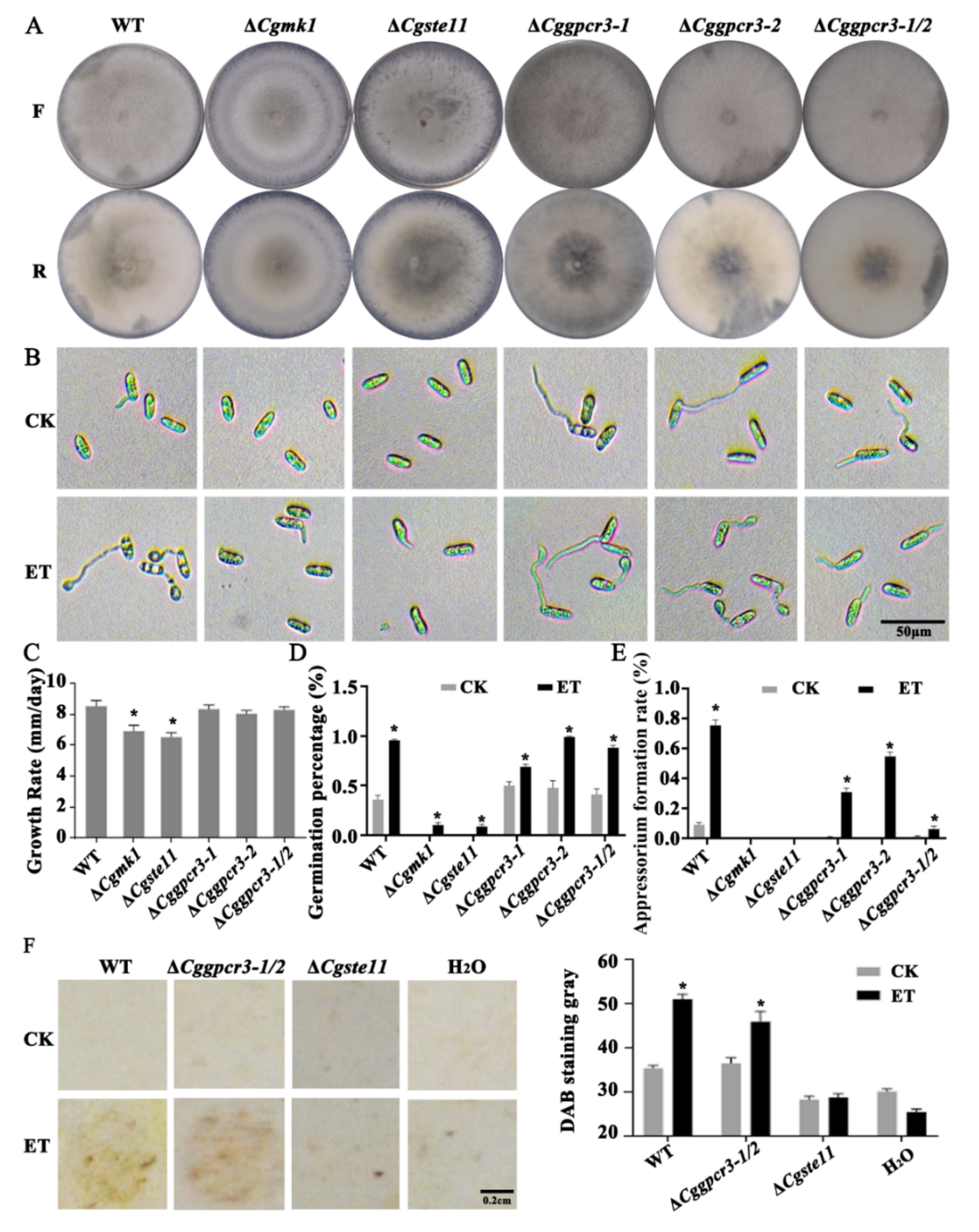
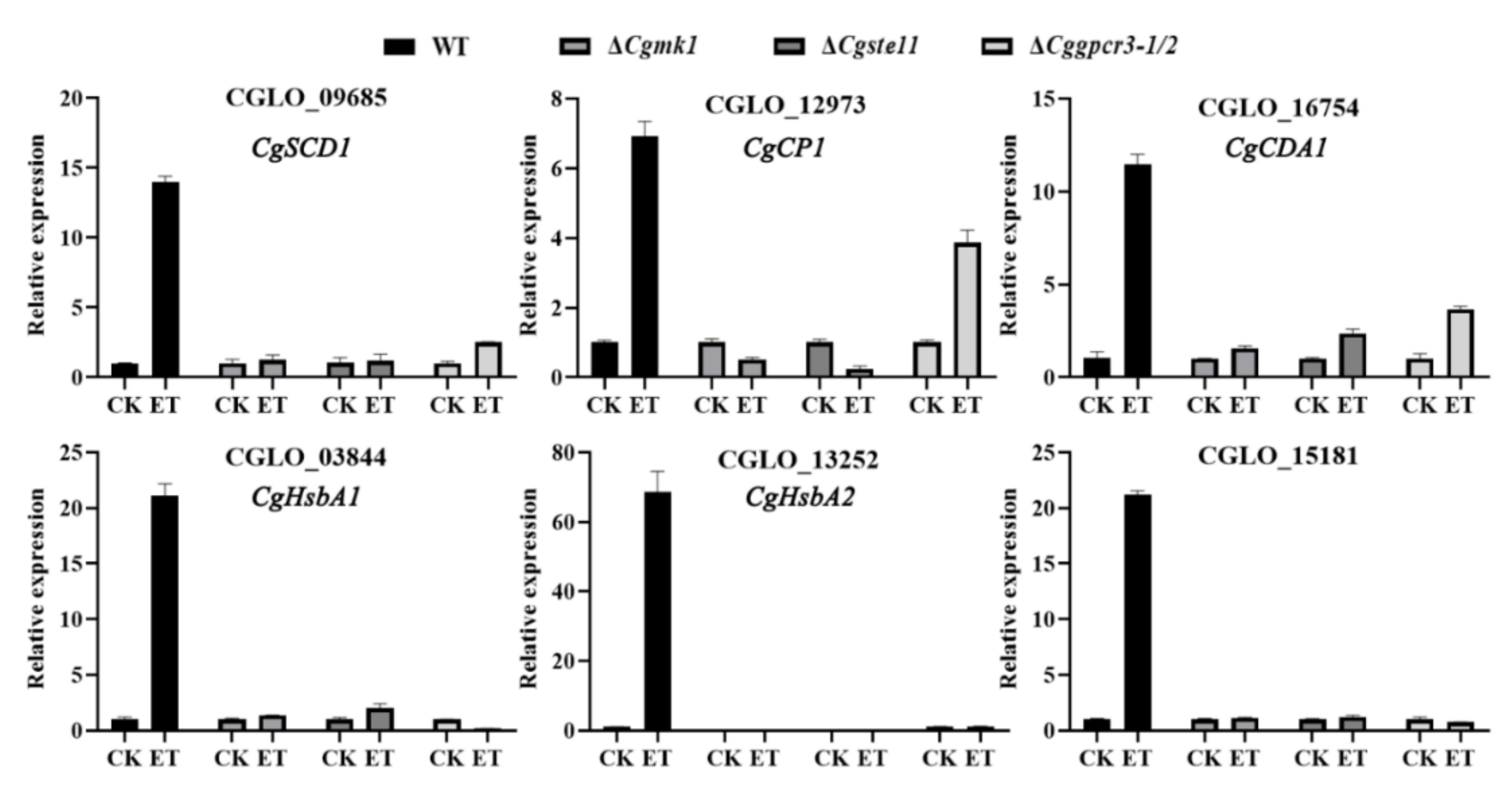
3.7. ET Promoting Appressorium Formation and Virulence Was Dependent on MAPK and GPCR Pathways
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, Z.; Zhong, S.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Pirrello, J.; Chervin, C.; Roustan, J.P.; Bouzayen, M. Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation. Plant Physiol. 2015, 169, 2380–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakeel, S.N.; Wang, X.; Binder, B.M.; Schaller, G.E. Mechanisms of signal transduction by ethylene: Overlapping and non-overlapping signalling roles in a receptor family. AoB Plants 2013, 5, plt010. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Meyerowitz, E.M. Ethylene Responses Are Negatively Regulated by a Receptor Gene Family in Arabidopsis thaliana. Cell 1998, 94, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Phukan, U.J.; Jeena, G.S.; Tripathi, V.; Shukla, R.K. Regulation of Apetala2/Ethylene Response Factors in Plants. Front. Plant Sci. 2017, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhang, Y.; Li, Z.; Liu, M. Role of ethylene response factors (ERFs) in fruit ripening. Food Qual. Saf. 2020, 4, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tang, M.; Liu, M.; Su, D.; Chen, J.; Gao, Y.; Bouzayen, M.; Li, Z. The Molecular Regulation of Ethylene in Fruit Ripening. Small Methods 2020, 4, 1900485. [Google Scholar] [CrossRef]
- Tintor, N.; Ross, A.; Kanehara, K.; Yamada, K.; Fan, L.; Kemmerling, B.; Nürnberger, T.; Tsuda, K.; Saijo, Y. Layered pattern receptor signaling via ethylene and endogenous elicitor peptides during Arabidopsis immunity to bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, 6211–6216. [Google Scholar] [CrossRef] [Green Version]
- Barth, M.; Hankinson, T.R.; Zhuang, H.; Breidt, F. Microbiological Spoilage of Fruits and Vegetables. In Compendium of the Microbiological Spoilage of Foods and Beverages; Sperber, W.H., Doyle, M.P., Eds.; Springer: New York, NY, USA, 2009; pp. 135–183. [Google Scholar]
- Darras, A.I. Postharvest Disease Management. In Handbook of Florists’ Crops Diseases; McGovern, R.J., Elmer, W.H., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 253–279. [Google Scholar]
- Dong, X. SA, JA, ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1998, 1, 316–323. [Google Scholar] [CrossRef]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of Hormone Signaling Networks in Plant Defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 integrates signals from ethylene and jasmonate pathways in plant defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flaishman, M.A.; Kolattukudy, P.E. Timing of fungal invasion using host’s ripening hormone as a signal. Proc. Natl. Acad. Sci. USA 1994, 91, 6579–6583. [Google Scholar] [CrossRef] [Green Version]
- El-Kazzaz, M.K.; Sommer, N.F.; Kader, A.A. Ethylene effects on in vitro and in vivo growth of certain postharvest fruit-infecting fungi. Phytopathology 1983, 73, 998–1001. [Google Scholar] [CrossRef]
- Kępczyńska, E. Ethylene requirement during germination of Botiytis cinerea spores. Physiol. Plant. 1989, 77, 369–372. [Google Scholar] [CrossRef]
- Kępczyńska, E. Involvement of ethylene in spore germination and mycelial growth of Alternaria alternata. Mycol. Res. 1994, 98, 118–120. [Google Scholar] [CrossRef]
- Chagué, V.; Danit, L.V.; Siewers, V.; Schulze-Gronover, C.; Tudzynski, P.; Tudzynski, B.; Sharon, A. Ethylene sensing and gene activation in Botrytis cinerea: A missing link in ethylene regulation of fungus-plant interactions? Mol. Plant Microbe Interact. 2006, 19, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.Q.; Jiang, H.F.; Zhou, Y.Q.; Lei, Y.; Wang, S.Y.; Liao, B.S. Ethylene inhibited aflatoxin biosynthesis is due to oxidative stress alleviation and related to glutathione redox state changes in Aspergillus flavus. Int. J. Food Microbiol. 2009, 130, 17–21. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.R.; Jones, E.B.G.; et al. Colletotrichum–Names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Cannon, P.F.; Bridge, P.D.; Monte, E. Linking the past, present, and future of Colletotrichum systematics. In Colletotrichum: Host Specificity, Pathology, and Host Pathogen Interaction; Prusky, D.F.S., Dickman, M.B., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2000; pp. 1–20. [Google Scholar]
- Sutton, B.C. The genus Glomerella and its anamorph Colletotrichum. In Colletotrichum: Biology, pathology andcontrol; Bailey, J.A., Jeger, M.J., Eds.; CAB International: Wallingford, UK, 1992; pp. 1–26. [Google Scholar]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot Nicholas, J. Autophagic Fungal Cell Death Is Necessary for Infection by the Rice Blast Fungus. Science 2006, 312, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Chen, C.; Yang, J.; Feng, W.; Liu, X.; Zuo, R.; Wang, J.; Yang, L.; Zhong, K.; Gao, C.; et al. Histone acetyltransferase MoHat1 acetylates autophagy-related proteins MoAtg3 and MoAtg9 to orchestrate functional appressorium formation and pathogenicity in Magnaporthe oryzae. Autophagy 2019, 15, 1234–1257. [Google Scholar] [CrossRef]
- Li, L.; Wright, S.J.; Krystofova, S.; Park, G.; Borkovich, K.A. Heterotrimeric G Protein Signaling in Filamentous Fungi. Annu. Rev. Microbiol. 2007, 61, 423–452. [Google Scholar] [CrossRef]
- Brown, N.A.; Schrevens, S.; van Dijck, P.; Goldman, G.H. Fungal G-protein-coupled receptors: Mediators of pathogenesis and targets for disease control. Nat. Microbiol. 2018, 3, 402–414. [Google Scholar] [CrossRef]
- Liu, S.; Dean, R.A. G protein alpha subunit genes control growth, development, and pathogenicity of Magnaporthe grisea. Mol. Plant-Microbe Interact. MPMI 1997, 10, 1075–1086. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Park, G.; Xu, J.R. The G-beta subunit MGB1 is involved in regulating multiple steps of infection-related morphogenesis in Magnaporthe grisea. Mol. Microbiol. 2003, 50, 231–243. [Google Scholar] [CrossRef]
- Liang, S.; Wang, Z.; Liu, P.; Li, D. A Gγ subunit promoter T-DNA insertion mutant—A1-412 of Magnaporthe grisea is defective in appressorium formation, penetration and pathogenicity. Chin. Sci. Bull. 2006, 51, 2214–2218. [Google Scholar] [CrossRef]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Ruan, H.; Akutse, K.S.; Lai, B.; Lin, Y.; Hou, Y.; Zhong, F. Ethylene and Benzaldehyde Emitted from Postharvest Tomatoes Inhibit Botrytis cinerea via Binding to G-Protein Coupled Receptors and Transmitting with cAMP-Signal Pathway of the Fungus. J. Agric. Food Chem. 2019, 67, 13706–13717. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, D.; Tian, C. Mitogen-activated protein kinase cascade CgSte50-Ste11-Ste7-Mk1 regulates infection-related morphogenesis in the poplar anthracnose fungus Colletotrichum gloeosporioides. Microbiol. Res. 2021, 248, 126748. [Google Scholar] [CrossRef] [PubMed]
- Osés-Ruiz, M.; Cruz-Mireles, N.; Martin-Urdiroz, M.; Soanes, D.M.; Eseola, A.B.; Tang, B.; Derbyshire, P.; Nielsen, M.; Cheema, J.; Were, V.; et al. Appressorium-mediated plant infection by Magnaporthe oryzae is regulated by a Pmk1-dependent hierarchical transcriptional network. Nat. Microbiol. 2021, 6, 1383–1397. [Google Scholar] [CrossRef]
- Priyatno, T.P.; Abu Bakar, F.D.; Kamaruddin, N.; Mahadi, N.M.; Abdul Murad, A.M. Inactivation of the catalytic subunit of cAMP-dependent protein kinase A causes delayed appressorium formation and reduced pathogenicity of Colletotrichum gloeosporioides. Sci. World J. 2012, 2012, 545784. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Wang, Y.; Wang, X.; Zhang, X.; Tian, C. The Mitogen-Activated Protein Kinase CgMK1 Governs Appressorium Formation, Melanin Synthesis, and Plant Infection of Colletotrichum gloeosporioides. Front. Microbiol. 2017, 8, 2216. [Google Scholar] [CrossRef]
- Wang, T.; Xiang, S.; Ren, D.D.; Zhu, P.K.; Xu, L. First Report of Colletotrichum gloeosporioides Causing Postharvest Fruit Rot on Citrus aurantifolia in China. Plant Dis. 2019, 103, 2686. [Google Scholar] [CrossRef]
- Wang, T.; Ren, D.; Guo, H.; Chen, X.; Zhu, P.; Nie, H.; Xu, L. CgSCD1 Is Essential for Melanin Biosynthesis and Pathogenicity of Colletotrichum gloeosporioides. Pathogens 2020, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Xu, L.; Zhang, C.; Toyoda, H.; Gan, S.-S. Ethylene produced by Botrytis cinerea can affect early fungal development and can be used as a marker for infection during storage of grapes. Postharvest Biol. Technol. 2012, 66, 23–29. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. Embnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, A.; Lu, P.; Dahl-Roshak, A.M.; Paress, P.S.; Kennedy, S.; Tkacz, J.S.; An, Z. Efficient disruption of a polyketide synthase gene (pks1) required for melanin synthesis through Agrobacterium-mediated transformation of Glarea lozoyensis. Mol. Genet Genom. 2003, 268, 645–655. [Google Scholar] [CrossRef]
- Fister, A.S.; O’Neil, S.T.; Shi, Z.; Zhang, Y.; Tyler, B.M.; Guiltinan, M.J.; Maximova, S.N. Two Theobroma cacao genotypes with contrasting pathogen tolerance show aberrant transcriptional and ROS responses after salicylic acid treatment. J. Exp. Bot. 2015, 66, 6245–6258. [Google Scholar] [CrossRef] [Green Version]
- Choquer, M.; Rascle, C.; Gonçalves, I.R.; de Vallée, A.; Ribot, C.; Loisel, E.; Smilevski, P.; Ferria, J.; Savadogo, M.; Souibgui, E.; et al. The infection cushion of Botrytis cinerea: A fungal ‘weapon’ of plant-biomass destruction. Environ. Microbiol. 2021, 23, 2293–2314. [Google Scholar] [CrossRef]
- Ohtaki, S.; Maeda, H.; Takahashi, T.; Yamagata, Y.; Hasegawa, F.; Gomi, K.; Nakajima, T.; Abe, K. Novel hydrophobic surface binding protein, HsbA, produced by Aspergillus oryzae. Appl. Environ. Microbiol. 2006, 72, 2407–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.; Ribera, J. Microbial production of melanin and its various applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, G.; Kenmochi, Y.; Takano, Y.; Sweigard, J.; Farrall, L.; Furusawa, I.; Horino, O.; Kubo, Y. Novel fungal transcriptional activators, Cmr1p of Colletotrichum lagenarium and pig1p of Magnaporthe grisea, contain Cys2His2 zinc finger and Zn(II)2Cys6 binuclear cluster DNA-binding motifs and regulate transcription of melanin biosynthesis genes in a developmentally specific manner. Mol. Microbiol. 2000, 38, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.S.; Kolattukudy, P.E. Isolation and characterization of genes expressed uniquely during appressorium formation by Colletotrichum gloeosporioides conidia induced by the host surface wax. Mol. Gen. Genet 1995, 247, 282–294. [Google Scholar] [CrossRef]
- Wang, W.; An, B.; Feng, L.; He, C.; Luo, H. A Colletotrichum gloeosporioides cerato-platanin protein, CgCP1, contributes to conidiation and plays roles in the interaction with rubber tree. Can. J. Microbiol. 2018, 64, 826–834. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef] [Green Version]
- Ju, C.; Chang, C. Mechanistic Insights in Ethylene Perception and Signal Transduction. Plant Physiol. 2015, 169, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Musgrave, A.; Walters, J. Ethylene-stimulated growth and auxin ansport in ranunculus sceleratus petioles. New Phytol. 1973, 72, 783–789. [Google Scholar] [CrossRef]
- Métraux, J.P.; Kende, H. The role of ethylene in the growth response of submerged deep water rice. Plant Physiol. 1983, 72, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Dagdas, Y.F.; Yoshino, K.; Dagdas, G.; Ryder, L.S.; Bielska, E.; Steinberg, G.; Talbot, N.J. Septin-mediated plant cell invasion by the rice blast fungus, Magnaporthe oryzae. Science 2012, 336, 1590–1595. [Google Scholar] [CrossRef]
- Jacobson, E. Pathogenic Roles for Fungal Melanins. Clin. Microbiol. Rev. 2000, 13, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Lohrer, M.; Hempel, M.; Mathea, S.; Schliebner, I.; Menzel, M.; Kiesow, A.; Schaffrath, U.; Deising, H.B.; Horbach, R. Melanin is not required for turgor generation but enhances cell-wall rigidity in appressoria of the corn pathogen Colletotrichum graminicola. Mol. Plant Microbe Interact. 2014, 27, 315–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinekamp, T.; Thywissen, A.; Macheleidt, J.; Keller, S.; Valiante, V.; Brakhage, A. Aspergillus fumigatus melanins: Interference with the host endocytosis pathway and impact on virulence. Front. Microbiol. 2013, 3, 440. [Google Scholar] [CrossRef] [Green Version]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P.; Georgeault, S.; Mallet, R.; Chabasse, D.; Symoens, F.; Bouchara, J.-P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Frías, M.; Gonzalez, C.; Brito, N. BcSpl1, a cerato-platanin family protein, contributes to Botrytis cinerea virulence and elicits the hypersensitive response in the host. New Phytol. 2011, 192, 483–495. [Google Scholar] [CrossRef]
- Jeong, J.; Mitchell, T.; Dean, R. The Magnaporthe grisea snodprot1 homolog, MSPI, is required for virulence. FEMS Microbiol. Lett. 2007, 273, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Zhao, X.; Lü, Y.; Jin, C. Chitin deacetylases Cod4 and Cod7 are involved in polar growth of Aspergillus fumigatus. Microbiologyopen 2020, 9, e00943. [Google Scholar] [CrossRef] [Green Version]
- Kuroki, M.; Okauchi, K.; Yoshida, S.; Ohno, Y.; Murata, S.; Nakajima, Y.; Nozaka, A.; Tanaka, N.; Nakajima, M.; Taguchi, H.; et al. Chitin-deacetylase activity induces appressorium differentiation in the rice blast fungus Magnaporthe oryzae. Sci. Rep. 2017, 7, 9697. [Google Scholar] [CrossRef] [Green Version]
- Deising, H.; Siegrist, J. Chitin deacetylase activity of the rust Uromyces viciae-fabae is controlled by fungal morphogenesis. FEMS Microbiol. Lett. 2006, 127, 207–211. [Google Scholar] [CrossRef]
- Baker, L.G.; Specht, C.A.; Donlin, M.J.; Lodge, J.K. Chitosan, the deacetylated form of chitin, is necessary for cell wall integrity in Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 855–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoghegan, I.A.; Gurr, S.J. Chitosan Mediates Germling Adhesion in Magnaporthe oryzae and Is Required for Surface Sensing and Germling Morphogenesis. PLoS Pathog. 2016, 12, e1005703. [Google Scholar] [CrossRef] [PubMed]
- Perez-Dulzaides, R.; Camacho, E.; Cordero, R.J.B.; Casadevall, A. Cell-wall dyes interfere with Cryptococcus neoformans melanin deposition. Microbiology 2018, 164, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhang, B.S.; Zhao, J.H.; Huang, J.F.; Jia, P.S.; Wang, S.; Zhang, J.; Zhou, J.M.; Guo, H.S. Deacetylation of chitin oligomers increases virulence in soil-borne fungal pathogens. Nat. Plants 2019, 5, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cruz, J.M.; Polonio, Á.; Zanni, R.; Romero, D.; Gálvez, J.; Fernández-Ortuño, D.; Pérez-García, A. Chitin Deacetylase, a Novel Target for the Design of Agricultural Fungicides. J. Fungi 2021, 7, 1009. [Google Scholar] [CrossRef]
- Li, D.; Sirakova, T.; Rogers, L.; Ettinger, W.F.; Kolattukudy, P.E. Regulation of constitutively expressed and induced cutinase genes by different zinc finger transcription factors in Fusarium solani f. sp. pisi (Nectria haematococca). J. Biol. Chem. 2002, 277, 7905–7912. [Google Scholar] [CrossRef] [Green Version]
- Rocha, A.L.; Di Pietro, A.; Ruiz-Roldan, C.; Roncero, M.I. Ctf1, a transcriptional activator of cutinase and lipase genes in Fusarium oxysporum is dispensable for virulence. Mol. Plant Pathol. 2008, 9, 293–304. [Google Scholar] [CrossRef]
- Saunders, D.G.; Aves, S.J.; Talbot, N.J. Cell cycle-mediated regulation of plant infection by the rice blast fungus. Plant Cell 2010, 22, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.D.; Johnson, A.M.; Dean, R.A. Chemical signals responsible for appressorium formation in the rice blast fungus Magnaporthe grisea. Physiol. Mol. Plant Pathol. 1996, 48, 335–346. [Google Scholar] [CrossRef]
- Van der Linde, K.; Göhre, V. How Do Smut Fungi Use Plant Signals to Spatiotemporally Orientate on and In Planta? J. Fungi 2021, 7, 107. [Google Scholar] [CrossRef]
- Choi, W.; Dean, R.A. The adenylate cyclase gene MAC1 of Magnaporthe grisea controls appressorium formation and other aspects of growth and development. Plant Cell 1997, 9, 1973–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.M.; Kolattukudy, P.E. Early expression of the calmodulin gene, which precedes appressorium formation in Magnaporthe grisea, is inhibited by self-inhibitors and requires surface attachment. J. Bacteriol. 1999, 181, 3571–3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Kawano, T.; Li, D.; Kolattukudy, P.E. A mitogen-activated protein kinase kinase required for induction of cytokinesis and appressorium formation by host signals in the conidia of Colletotrichum gloeosporioides. Plant Cell 2000, 12, 1331–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papon, N.; Binder, B.M. An Evolutionary Perspective on Ethylene Sensing in Microorganisms. Trends Microbiol. 2019, 27, 193–196. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Genotype Description | Strain Source |
|---|---|---|
| WT | Wild-type C. gloeosporioides (EX2016-02) | [41] |
| Cg-gfp | Fluorescent strain of C. gloeosporioides | In this study |
| ΔCgcap22 | Appressorium structural protein null mutant | In this study |
| ΔCgscd1 | Scytalone dehydratase null mutant | [42] |
| ΔCgcp1 | Cerato-platanin protein null mutant | In this study |
| ΔCgmk1 | MAPK null mutant | In this study |
| ΔCgste11 | MEK kinase null mutant | In this study |
| ΔCggpcr3-1 | G-protein-coupled receptor 3-1 null mutant | In this study |
| ΔCggpcr3-2 | G-protein-coupled receptor 3-2 null mutant | In this study |
| ΔCggpcr3-1/2 | G-protein-coupled receptors 3-1 and 3-2 double deletion mutant | In this study |
| PCGLO_03844-gfp | Transgenic strain expressing GFP driven by CGLO_03844 promoter | In this study |
| PCGLO_13252-gfp | Transgenic strain expressing GFP driven by CGLO_13252 promoter | In this study |
| Id | log2FC | Description | Length (aa) | Domain |
|---|---|---|---|---|
| CGLO_15589 | 5.0674 | DJ-1/PfpI family protein | 248 | GATase1_PfpI_2 (34–229) |
| CGLO_07422 | 4.8147 | extracellular-serine-rich protein | 198 | Cupredoxin (53–154) |
| CGLO_11351 | 4.7586 | hypothetical protein | 208 | SodA (5–193) |
| CGLO_03249 | 4.6034 | hypothetical protein | 354 | M35_deuterolysin_like (168–343) |
| CGLO_07611 | 4.6008 | hypothetical protein | 218 | LPMO_auxiliary-like (21–138) |
| CGLO_14431 | 4.5783 | hypothetical protein | 237 | nucleoside_deaminase (65–151) |
| CGLO_03844 | 4.4491 | cell wall protein | 223 | HsbA (29–150) |
| CGLO_02789 | 4.339 | catalase | 504 | catalase_fungal (44–492) |
| CGLO_05804 | 4.2188 | WSC-domain-containing protein | 446 | WSC (43–122,152–227,255–332) |
| CGLO_00547 | 4.1784 | hypothetical protein | 167 | HsbA (4–123) |
| CGLO_13252 | 4.1765 | hypothetical protein | 319 | HsbA (27–146) |
| CGLO_11882 | 4.018 | cas1-appressorium-specific protein | 241 | DUF3129 (19–193) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, D.; Wang, T.; Zhou, G.; Ren, W.; Duan, X.; Gao, L.; Chen, J.; Xu, L.; Zhu, P. Ethylene Promotes Expression of the Appressorium- and Pathogenicity-Related Genes via GPCR- and MAPK-Dependent Manners in Colletotrichum gloeosporioides. J. Fungi 2022, 8, 570. https://doi.org/10.3390/jof8060570
Ren D, Wang T, Zhou G, Ren W, Duan X, Gao L, Chen J, Xu L, Zhu P. Ethylene Promotes Expression of the Appressorium- and Pathogenicity-Related Genes via GPCR- and MAPK-Dependent Manners in Colletotrichum gloeosporioides. Journal of Fungi. 2022; 8(6):570. https://doi.org/10.3390/jof8060570
Chicago/Turabian StyleRen, Dandan, Tan Wang, Ganghan Zhou, Weiheng Ren, Xiaomin Duan, Lin Gao, Jiaxu Chen, Ling Xu, and Pinkuan Zhu. 2022. "Ethylene Promotes Expression of the Appressorium- and Pathogenicity-Related Genes via GPCR- and MAPK-Dependent Manners in Colletotrichum gloeosporioides" Journal of Fungi 8, no. 6: 570. https://doi.org/10.3390/jof8060570