
Efficacy of Biological and Chemical Control Agents against Heterobasidion Spore Infections of Norway Spruce and Scots Pine Stumps on Drained Peatland

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Sites
2.2. Stump Inoculation
2.3. Sampling
2.4. Data Preparation
2.5. Statistical Analyses
3. Results
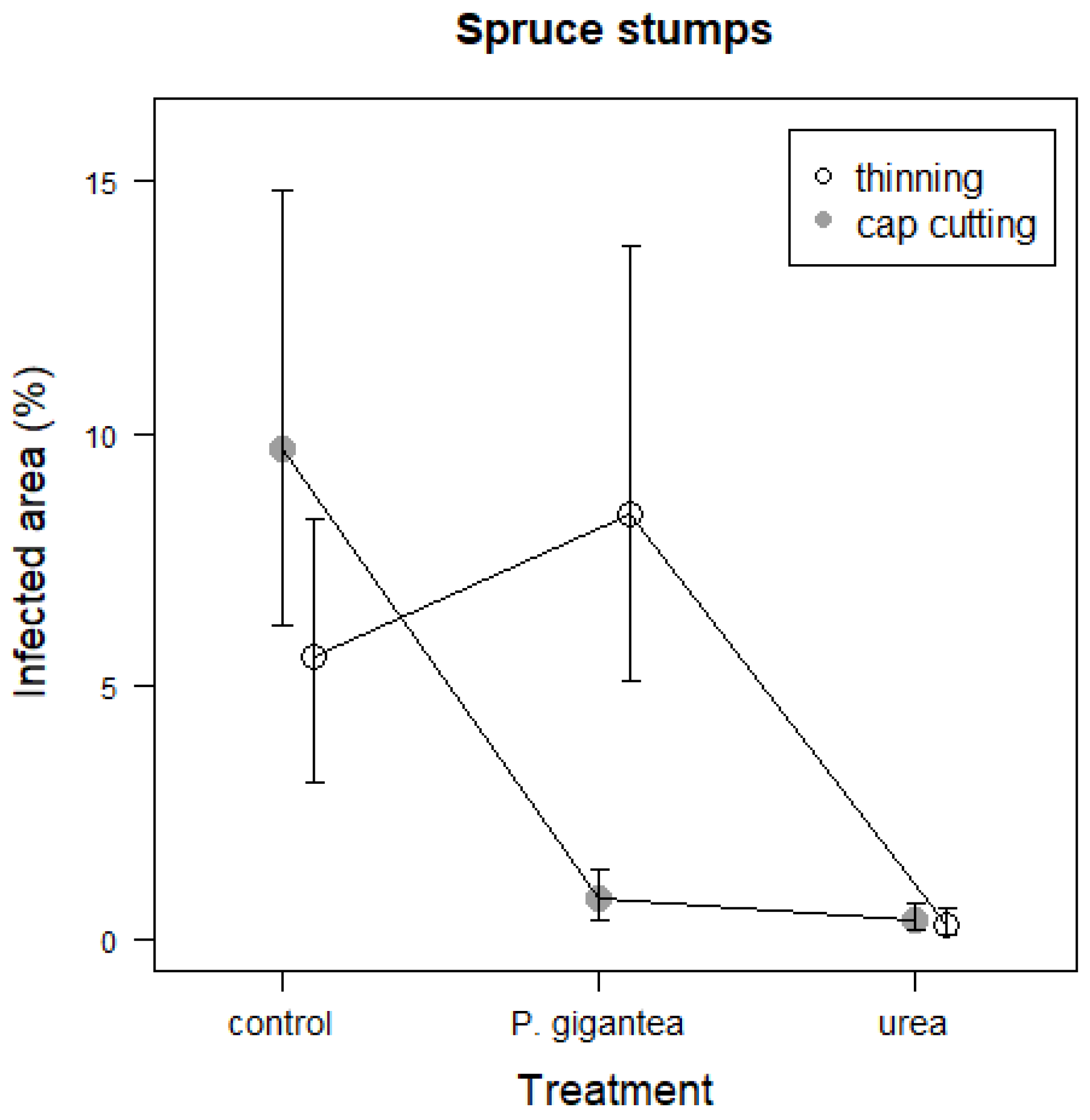
3.1. Artificial and Natural Heterobasidion Infections
3.2. Efficacy of Urea and P. gigantea Treatments
4. Discussion
4.1. Heterobasidion Infections
4.2. Efficacy of P. gigantea
4.3. Efficacy of Urea
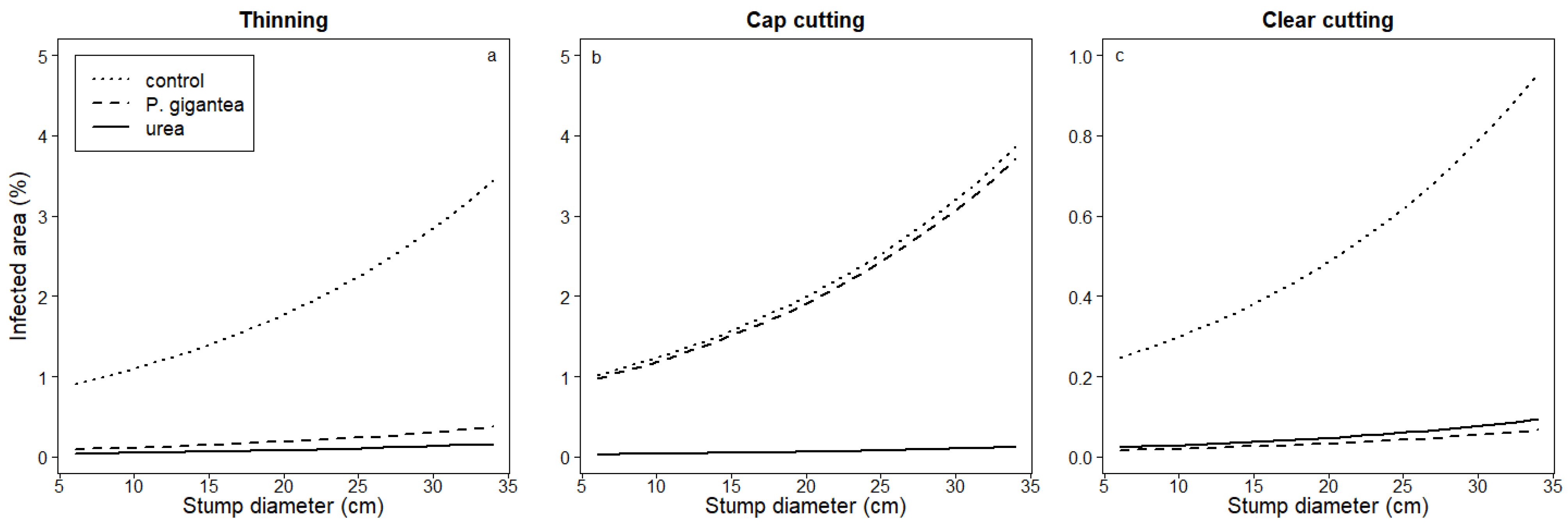
4.4. Effects of Stump and Soil Characteristics on the Control Efficacy
4.4.1. Stump Moisture Content
4.4.2. Stump Diameter
4.4.3. Soil pH
4.4.4. Thickness of the Peat Layer
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asiegbu, F.O.; Adomas, A.; Stenlid, J. Conifer root and butt rot caused by Heterobasidion annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2005, 6, 395–409. [Google Scholar] [CrossRef]
- Garbelotto, M.; Gonthier, P. Biology, epidemiology, and control of Heterobasidion species worldwide. Annu. Rev. Phytopathol. 2013, 51, 39–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redfern, D.B.; Stenlid, J. Spore dispersal and infection. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: London, UK, 1998; pp. 105–124. [Google Scholar]
- Stenlid, J.; Redfern, D.B. Spread within the tree and stand. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: London, UK, 1998; pp. 125–141. [Google Scholar]
- Piri, T.; Valkonen, S. Incidence and spread of Heterobasidionroot rot in uneven-aged Norway spruce stands. Can. J. For. Res. 2013, 43, 872–877. [Google Scholar] [CrossRef]
- Piri, T.; Vainio, E.J.; Nuorteva, H.; Hantula, J. High seedling mortality of Scots pine caused by Heterobasidion annosum s.s. Forests 2021, 12, 1289. [Google Scholar] [CrossRef]
- Kallio, T. Aerial distribution of the root-rot fungus Fomes annosus (Fr.) Cooke in Finland. Acta For. Fenn. 1970, 107, 1–55. [Google Scholar] [CrossRef] [Green Version]
- Bendz-Hellgren, M.; Lipponen, K.; Solheim, H.; Thomsen, I.M. The Nordic Countries. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: London, UK, 1998; pp. 333–345. [Google Scholar]
- Holdenrieder, O.; Greig, B.J.W. Biological methods of control. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: London, UK, 1998; pp. 235–258. [Google Scholar]
- Korhonen, K.; Lipponen, K.; Bendz, M.; Johansson, M.; Ryen, I.; Venn, K.; Seiskari, P.; Niemi, M. Control of Heterobasidion annosum by stump treatment with ’Rotstop’ a new commercial formulation of Phlebiopsis gigantea. In Proceedings of the Eight International Conference on Root and Butt Rots, Sweden and Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Science: Uppsala, Sweden, 1994; pp. 674–685. [Google Scholar]
- Johansson, S.M.; Pratt, J.E.; Asiegbu, F.O. Treatment of Norway spruce and Scots pine stumps with urea against the root and butt rot fungus Heterobasidion annosum—Possible modes of action. For. Ecol. Manag. 2002, 157, 87–100. [Google Scholar] [CrossRef]
- Brandtberg, P.O.; Johansson, M.; Seeger, P. Effects of season and urea treatment on infection of stumps of Picea abies by Heterobasidion annosum in stands on former arable land. Scand. J. For. Res. 1996, 11, 261–268. [Google Scholar] [CrossRef]
- Berglund, M.; Rönnberg, J. Effectiveness of treatment of Norway spruce stumps with Phlebiopsis gigantea at different rates of coverage for the control of Heterobasidion. For. Pathol. 2004, 34, 233–243. [Google Scholar] [CrossRef]
- Nicolotti, G.; Gonthier, P. Stump treatment against Heterobasidion with Phlebiopsis gigantea and some chemicals in Picea abies stands in the western Alps. For. Pathol. 2005, 35, 365–374. [Google Scholar] [CrossRef]
- Oliva, J.; Samils, N.; Johansson, U.; Bendz-Hellgren, M.; Stenlid, J. Urea treatment reduced Heterobasidion annosum s.l. root rot in Picea abies after 15 years. For. Ecol. Manag. 2008, 255, 2876–2882. [Google Scholar] [CrossRef]
- Rönnberg, J.; Cleary, M.R. Presence of Heterobasidion infections in Norway spruce stumps 6 years after treatment with Phlebiopsis gigantea. For. Pathol. 2012, 42, 144–149. [Google Scholar] [CrossRef]
- Gonthier, P. Frequency of stump infections by Heterobasidion annosum s.l. and benefits from urea treatments vary with tree species and season in European Alpine forests. For. Ecol. Manag. 2019, 434, 76–86. [Google Scholar] [CrossRef]
- Zaluma, A.; Sherwood, P.; Bruna, L.; Skola, U.; Gaitnieks, T.; Rönnberg, J. Control of Heterobasidion in Norway spruce stands: The impact of stump cover on efficacy of urea and Phlebiopsis gigantea and implications for forest management. Forests 2021, 12, 679. [Google Scholar] [CrossRef]
- Korhonen, K. Simulated stump treatment experiments for monitoring the efficacy of Phlebiopsis gigantea against Heterobasidion infection. In Proceedings of the 10th International Conference on Root and Butt Rots of Forest Trees, Quebec City, QC, Canada, 16–22 September 2001; Information Report LAU-X-126; LaFlamme, G., Bérubé, J., Bussières, G., Sainte-Foy, G., Eds.; Canadian Forest Service: Quebec City, QC, Canada, 2003; pp. 206–210. [Google Scholar]
- Thomsen, I.M. Effect of stump treatment on transfer of Heterobasidion annosum root rot in Norway spruce. In Proceedings of the 10th International Conference on Root and Butt Rots of Forest Trees, Quebec City, QC, Canada, 16–22 September 2001; Information Report LAU-X-126; LaFlamme, G., Bérubé, J., Bussières, G., Sainte-Foy, G., Eds.; Canadian Forest Service: Quebec City, QC, Canada, 2003; pp. 160–169. [Google Scholar]
- Drenkhan, T.; Hanso, S.; Hanso, M. Effect of the stump treatment with Phlebiopsis gigantea against Heterobasidionroot rot in Estonia. Balt. For. 2008, 14, 16–25. [Google Scholar]
- Pellicciaro, M.; Lione, G.; Ongaro, S.; Gonthier, P. Comparative efficacy of state-of-art and new biological stump treatments in forests infested by the native and the alien invasive Heterobasidion species present in Europe. Pathogens 2021, 10, 1272. [Google Scholar] [CrossRef]
- La Porta, N.; Grillo, R.; Ambrosi, P.; Korhonen, K. Stump treatment experiments against Heterobasidion in the Italian Alps. In Proceedings of the 10th International Conference on Root and Butt Rots of Forest Trees, Quebec City, QC, Canada, 16–22 September 2001; Information Report LAU-X-126; LaFlamme, G., Bérubé, J., Bussières, G., Sainte-Foy, G., Eds.; Canadian Forest Service: Quebec City, QC, Canada, 2003; pp. 176–180. ISBN 0662333322. Available online: http://hdl.handle.net/10449/16731 (accessed on 17 October 2022).
- Sun, H.; Korhonen, K.; Hantula, J.; Kasanen, R. Variation in properties of Phlebiopsis gigantea related to biocontrol against infection by Heterobasidion spp. in Norway spruce stumps. For. Pathol. 2009, 39, 133–144. [Google Scholar] [CrossRef]
- Blomquist, M.; Larsson Herrera, S.; Hofmann, J.; Beram, R.C.; Cleary, M.; Rönnberg, J. Size matters but is big always better? Effectiveness of urea and Phlebiopsis gigantea as treatment against Heterobasidion on Picea abies stumps of variable size. For. Ecol. Manag. 2020, 462, 117998. [Google Scholar] [CrossRef]
- Rönnberg, J.; Sidorov, E.; Petrylaitè, E. Efficacy of different concentrations of Rotstop® and Rotstop®S and imperfect coverage of Rotstop®S against Heterobasidion spp. spore infections on Norway spruce stumps. For. Pathol. 2006, 36, 422–433. [Google Scholar] [CrossRef]
- Tubby, K.V.; Scott, D.; Weber, J.F. Relationship between stump treatment coverage using the biological control product PG Suspension, and control of Heterobasidion annosum on Corsican pine, Pinus nigra spp. laricio. For. Pathol. 2008, 38, 37–46. [Google Scholar] [CrossRef]
- Pratt, J.E.; Redfern, D.B. Infection of Sitka spruce stumps by spores of Heterobasidion annosum: Control by means of urea. Forestry 2001, 74, 73–78. [Google Scholar] [CrossRef]
- Sirota, Z.H. Dry weight loss of wood after the inoculation of Scots pine stumps with Phlebiopsis gigantea. Eur. J. For. Path. 1997, 27, 179–185. [Google Scholar] [CrossRef]
- Żółciak, A.; Sikora, K.; Wrzosek, M.; Damszel, M.; Sierota, Z. Why does Phlebiopsis gigantea not always inhibit root and butt rot in conifers? Forests 2020, 11, 129. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.E.; Johansson, M.; Hüttermann, A. Chemical control of Heterobasidion annosum. In Heterobasidion annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: London, UK, 1998; pp. 259–283. [Google Scholar]
- Korhonen, K.T.; Ahola, A.; Heikkinen, J.; Henttonen, H.M.; Hotanen, J.-P.; Ihalainen, A.; Melin, M.; Pitkänen, J.; Räty, M.; Sirviö, M.; et al. Forests of Finland 2014–2018 and their development 1921–2018. Silva Fenn. 2021, 55, 10662. [Google Scholar] [CrossRef]
- Ala-Ilomäki, J.; Högnäs, T.; Lamminen, S.; Sirén, M. Equipping a conventional wheeled forwarder for peatland operations. Int. J. For. Eng. 2011, 22, 7–13. [Google Scholar] [CrossRef]
- Salmivaara, A.; Launiainen, S.; Perttunen, J.; Nevalainen, P.; Pohjankukka, J.; Ala-Ilomäki, J.; Sirén, M.; Laurén, A.; Tuominen, S.; Uusitalo, J.; et al. Towards dynamic forest trafficability prediction using open spatial data, hydrological modelling and sensor technology. For. Int. J. For. Res. 2020, 93, 662–674. [Google Scholar] [CrossRef]
- Palander, T.; Kärhä, K. Utilization of image, LiDAR and gamma-ray information to Improve environmental sustainability of cut-to-length wood harvesting operations in peatlands: A management systems perspective. ISPRS Int. J. Geo-Inf. 2021, 10, 273. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Bruna, L.; Zaluma, A.; Burnevica, N.; Klavina, D.; Legzdina, L.; Jansons, J.; Piri, T. Development of Heterobasidion spp. fruit bodies on decayed Picea abies. For. Ecol. Manag. 2021, 482, 118835. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Silbauma, L.; Muižnieks, I.; Zaļuma, A.; Kļaviņa, D.; Burņeviča, N.; Grosberga, M.; Lazdiņš, A.; Piri, T. Spread of Heterobasidion genotypes in Norway spruce stands on drained peat soil in Latvia. Can. J. For. Res. 2022, 52, 499–510. [Google Scholar] [CrossRef]
- Silver, T.; Piri, T. Observations of Heterobasidionroot rot in Scots pine stands on peatlands in southern Finland. Suo 2017, 68, 1–12. Available online: http://www.suo.fi/pdf/article10110.pdf (accessed on 17 October 2022). (In Finish, English Summary).
- Laine, J.; Vasander, H.; Hotanen, J.-P.; Nousiainen, H.; Saarinen, M.; Penttilä, T. Suotyypit ja Turvekankaat—Kasvupaikkaopas; Metsäkustannus Oy: Helsinki, Finland, 2018; pp. 1–160. (In Finnish) [Google Scholar]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-157; R Foundation for Statistical Computing: Vienna, Austria, 2022. Available online: https://CRAN.R-project.org/package=nlme (accessed on 28 December 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 28 December 2022).
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c), R Package Version 2.3-1; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 28 December 2022).
- Kasanen, R.; Terhonen, E.; Huuskonen, S.; Sun, H.; Uotila, A. High infection rate of residual conifer stumps by Heterobasidion species in an area with assumed low infection pressure. Scand. J. For. Res. 2011, 26, 404–412. [Google Scholar] [CrossRef]
- Kenigsvalde, K.; Brauners, I.; Korhonen, K.; Zaļuma, A.; Mihailova, A.; Gaitnieks, T. Evaluation of the biological control agent Rotstop in controlling the infection of spruce and pine stumps by Heterobasidion in Latvia. Scand. J. For. Res. 2016, 31, 254–261. [Google Scholar] [CrossRef]
- Berglund, M.; Rönnberg, J.; Holmer, L.; Stenlid, J. Comparison of five strains of Phlebiopsis gigantea and two Trichoderma formulations for treatment against natural Heterobasidion spore infections on Norway spruce stumps. Scand. J. For. Res. 2005, 20, 12–17. [Google Scholar] [CrossRef]
- Möykkynen, T.; Kontiokari, J. Spore deposition of Heterobasidion annosum coll. in Picea abies stands of North Karelia, eastern Finland. For. Pathol. 2001, 31, 107–114. [Google Scholar] [CrossRef]
- Redfern, D.B. Infection of Picea sitchensis and Pinus contorta stumps by basidiospores of Heterobasidion annosum. Eur. J. For. Path. 1982, 12, 11–25. [Google Scholar] [CrossRef]
- Annesi, T.; Curcio, G.; D’Amico, L.; Motta, E. Biological control of Heterobasidion annosum on Pinus pinea by Phlebiopsis gigantea. For. Pathol. 2005, 35, 127–134. [Google Scholar] [CrossRef]
- Oliva, J.; Zhao, A.; Zarei, S.; Sedlák, P.; Stenlid, J. Effect of temperature on the interaction between Phlebiopsis gigantea and the root-rot forest pathogen Heterobasidion spp. For. Ecol. Manag. 2015, 340, 22–30. [Google Scholar] [CrossRef]
- Redfern, D.B.; Gregory, S.C.; Macaskill, G.A. Inoculum concentration and the colonization of Picea sitchensis stumps by basidiospores of Heterobasidion annosum. Scand. J. For. Res. 1997, 12, 41–49. [Google Scholar] [CrossRef]
- Witzell, J.; Berglund, M.; Rönnberg, J. Does temperature regime govern the establishment of Heterobasidion annosum in Scandinavia? Int. J. Biometeorol. 2011, 55, 275–284. [Google Scholar] [CrossRef]
- La Porta, N.; Grillo, R.; Ambrosi, P.; Korhonen, K. Biological control of Heterobasidion annosum in Norway spruce forest by non-pathogenic wood decay fungi. IOBC Bull. 2001, 24, 59–62. [Google Scholar]
- Sun, H.; Korhonen, K.; Hantula, J.; Asiegbu, F.O.; Kasanen, R. Use of breeding approach for improving biocontrol efficacy of Phlebiopsis gigantea against Heterobasidion infection of Norway spruce stumps. FEMS Microbiol. Ecol. 2009, 69, 266–273. [Google Scholar] [CrossRef]
- Pratt, J.E.; Niemi, M.; Sierota, Z.H. Comparison of three products based on Phlebiopsis gigantea for the control of Heterobasidion annosum in Europe. Biocontrol Sci. Technol. 2000, 10, 467–477. [Google Scholar] [CrossRef]
- Finnish Meteorological Institute. Ilmastovuosikatsaus. 2021. Available online: https://www.ilmastokatsaus.fi/2022/02/10/ilmastovuosikatsaus-2021/ (accessed on 3 October 2022).
- Redfern, D.B. The effect of wood moisture on infection of Sitka spruce stumps by basidiospores of Heterobasidion annosum. Eur. J. For. Pathol. 1993, 23, 218–235. [Google Scholar] [CrossRef]
- Bendz-Hellgren, M.; Stenlid, J. Effects of clear-cutting, thinning, and wood moisture content on the susceptibility of Norway spruce stumps to Heterobasidion annosum. Can. J. For. Res. 1998, 28, 759–765. [Google Scholar] [CrossRef]
- Phillips, D.H.; Greig, B.J.W. Some chemicals to prevent stump colonization by Fomes annosus (Fr.) Cooke. Ann. Appl. Biol. 1970, 66, 441–452. [Google Scholar] [CrossRef]
- Lipponen, K. Juurikäävän kantotartunta ja sen torjunta ensiharvennusmetsiköissä. Summary: Stump infection by Heterobasidion annosum and its control in stands at the first thinning stage. Folia For. 1991, 770, 1–12. [Google Scholar]
- Shain, L. The response of sapwood of Norway spruce to infection by Fomes annosus. Phytopathology 1971, 61, 301–307. [Google Scholar] [CrossRef]
- Rishbeth, J. Observations on the biology of Fomes annosus, with particular reference to East Anglian pine plantations. III Natural and experimental infection of pines, and some factors affecting severity of the disease. Ann. Bot. 1951, 15, 221–246. [Google Scholar] [CrossRef]
- Paludan, F. Infektion og Spredning af Fomes annosus i Ung Rødgran. [Infection and Spread of Fomes annosus in Young Norway Spruce]; Det Forstlige Førsøksvæsen: Copenhagen, Danmark, 1966; Volume 30, pp. 19–47. (In Danish) [Google Scholar]
- Solheim, H. Infeksjon av Rotkjuke på Granstubber Til Ulike Årstider og Effekten av Urea Behandling. [Seasonal Infection of Heterobasidion annosum on Stumps of Norway Spruce and Surface Coating with Urea]; Rapport fra Skogforsk; Norsk Institutt for Skogforskning: Oslo, Norway, 1994; Volume 3, pp. 1–10. Available online: http://hdl.handle.net/11250/2560330 (accessed on 17 October 2022). (In Norwegian)
- Gaitnieks, T.; Brauners, I.; Kenigsvalde, K.; Zaļuma, A.; Brūna, L.; Jansons, J.; Burņeviča, N.; Lazdiņš, A.; Vasaitis, R. Infection of pre-commercially cut stumps of Picea abies and Pinus sylvestris by Heterobasidion spp.—A comparative study. Silva Fenn. 2018, 52, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sellin, A. Sapwood–heartwood proportion related to tree diameter, age, and growth rate in Picea abies. Can. J. For. Res. 1994, 24, 1022–1028. [Google Scholar] [CrossRef]
- Wallis, G.W. Survey of Fomes annosus in East Anglian pine plantations. Forestry 1962, 33, 203–214. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Klavina, D.; Muiznieks, I.; Pennanen, T.; Velmala, S.; Vasaitis, R.; Menkis, A. Impact of Heterobasidion root rot on fine root morphology and associated fungi in Picea abies stands on peat soils. Mycorrhiza 2016, 26, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Mattila, U.; Nuutinen, T. Assessing the incidence of butt rot in Norway spruce in southern Finland. Silva Fenn. 2007, 41, 29–43. Available online: www.metla.fi/silvafennica/full/sf41/sf411029.pdf (accessed on 7 July 2022). [CrossRef] [Green Version]
- Arhipova, N.; Gaitnieks, T.; Donis, J.; Stenlid, J.; Vasaitis, R. Butt rot incidence, causal fungi, and related yield loss in Picea abies stands of Latvia. Can. J. For. Res. 2011, 41, 2337–2345. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Stand | Location (Lat/Lon) | Site Type a | Soil pH | Avg. Thickness of the Peat Layer, cm | Tree Species | Avg. Stand Age, Years | Avg. Stump Diameter b cm | Type of Cutting |
|---|---|---|---|---|---|---|---|---|
| Kuru | 61°59′37.109″ | MtkgII/ | 3.8 | 135 | pine | 65 | 21 | clearcutting |
| 23°41′13.743″ | Ptkg II | |||||||
| Multia 1 | 62°26′55.351″ | Ptkg II | 4.0 | 150 | pine | 117 | 18 | thinning |
| 25°0′28.664″ | ||||||||
| Keuruu 1 | 62°29′27.446″ | Ptkg II | 3.8 | 110 | pine | 78 | 21 | clearcutting |
| 24°21′14.783″ | ||||||||
| Keuruu 2 | 62°29′35.835″ | Ptkg II | 3.7 | 90 | pine | 65 | 12 | thinning |
| 24°21′15.578″ | ||||||||
| Suonenjoki 1 | 62°33′40.972″ | MtkgII | 3.8 | 60 | pine | 110 | 25 | cap cutting |
| 27°14′59.115″ | ||||||||
| Suonenjoki 2 | 62°33′46.092″ | Rhtkg I | 3.8 | 50 | spruce | 103 | 24 | cap cutting |
| 27°14′50.895″ | ||||||||
| Suonenjoki 3 | 62°33′35.468″ | Rhtkg I | 3.9 | 50 | spruce | 98 | 21 | thinning |
| 27°15′5.650″ | ||||||||
| Multia 2 | 62°32′14.467″ | Mtkg I/ | 3.9 | 80 | spruce | 74 | 17 | thinning |
| 24°57′55.717″ | Ptkg I | |||||||
| Hämeenlinna | 61°11′5.974″ | Mtkg II | 3.9 | 60 | spruce | 88 | 22 | cap cutting |
| 25°15′46.907″ | ||||||||
| Vesijako | 61°25′11.925″ | Mtkg I | 3.9 | 80 | spruce | 78 | 18 | cap cutting |
| 25°0′23.800″ |
| Experimental Stand | Treatment Date | Duration of the Exp., Day | No. of Het Spores, per cm−2 | No. of Treated/Analyzed Stumps | No. of Stumps inf. by Heterobasidion (%) | Treatment Efficiency, % | |||
|---|---|---|---|---|---|---|---|---|---|
| Urea | P. gigantea | Urea | P. gigantea | Urea | P. gigantea | ||||
| Pine stands | |||||||||
| Kuru | 28.7.2020 | 62/76 a | 2.1 | 25/1 | 29/2 | - | - | - | - |
| Multia 1 | 25.8.2020 | 63 | NA | 21/13 | 25/21 | 1 (7.7) | 6 (28.6) | 99.6 | 89.7 |
| Keuruu 1 | 26.8.2020 | 75 | 3.0 | 25/11 | 25/14 | 0 | 0 | 100 | 100 |
| Keuruu 2 | 26.8.2020 | 83 | 3.0 | 24/17 | 25/18 | 0 | 6 (33.3) | 100 | 22.9 |
| Suonenjoki 1 | 11.8.2021 | 69 | 4.6 | 30/30 | 30/27 | 0 | 26 (96.3) | 100 | −56.6 |
| Spruce stands | |||||||||
| Suonenjoki 2 | 30.7.2021 | 66 | 4.4 | 28/27 | 30/30 | 21 (77.8) | 30 (100) | 87.1 | −29.3 |
| Suonenjoki 3 | 30.7.2021 | 67 | 4.4 | 25/23 | 30/30 | 14 (60.9) | 30 (100) | 92.0 | 5.9 |
| Multia 2 | 23.8.2021 | 65 | 7.2 | 30/20 | 30/25 | 16 (80.0) | 23 (92.0) | 41.9 | −39.4 |
| Hämeenlinna | 20.6.2022 | 78 | 4.7 | 30/28 | 28/27 | 18 (64.3) | 7 (25.9) | 79.5 | 98.2 |
| Vesijako | 21.6.2022 | 76 | 3.0 | 30/28 | 30/27 | 6 (21.4) | 9 (33.3) | 97.2 | 95.9 |
| Explanatory Variables | Spruce Stump Model | Pine Stump Model | ||
|---|---|---|---|---|
| Coeff. ± SE | p | Coeff. ± SE | p | |
| Intercept | 0.666 ± 0.884 | 0.452 | −6.139 ± 0.753 | <0.001 |
| P. gigantea a | −2.489 ± 0.199 | <0.001 | −2.667 ± 0.360 | <0.001 |
| Urea a | −3.106 ± 0.201 | <0.001 | −2.325 ± 0.404 | <0.001 |
| Cap cutting b | - | - | 1.427 ± 0.337 | 0.148 |
| Thinning b | −0.682 ± 0.315 | 0.275 | 1.307 ± 0.299 | 0.143 |
| Stump diameter c | 0.030 ± 0.014 | 0.035 | 0.048 ± 0.020 | 0.014 |
| Moisture in sapwood d | 0.002 ± 0.002 | 0.251 | −0.328 × 10−3 ± 0.004 | 0.934 |
| Moisture in heartwood d | −0.006 ± 0.005 | 0.235 | −0.003 ± 0.008 | 0.741 |
| Thickness of peat e | −0.054 ± 0.009 | 0.102 | - | - |
| P. gigantea: cap cutting f | - | - | 2.625 ± 0.448 | <0.001 |
| Urea: cap cutting f | - | - | −1.106 ± 0.476 | 0.022 |
| P. gigantea: thinning f | 3.022 ± 0.320 | <0.001 | 0.417 ± 0.423 | 0.326 |
| Urea: thinning f | 0.510 ± 0.339 | 0.133 | −0.735 ± 0.473 | 0.122 |
| Experimental Stand | Initial MC% | Final MC% | Initial MC% | Final MC% |
|---|---|---|---|---|
| Sapwood | Sapwood | Heartwood | Heartwood | |
| Pine stands | ||||
| Kuru | 105 (42–144) | 70 (40–169) | 42 (32–59) | 49 (32–74) |
| Multia 1 | 103 (57–149) | 127 (68–174) | 44 (32–84) | 46 (31–121) |
| Keuruu 1 | 104 (42–249) | 125 (84–170) | 43 (35–68) | 47 (33–75) |
| Keuruu 2 | 115 (56–181) | 135 (74–181) | 44 (15–60) | 54 (31–103) |
| Suonenjoki 1 | 133 (58–270) | 132 (71–287) | 48 (33–155) | 44 (31–80) |
| Spruce stands | ||||
| Suonenjoki 2 | 120 (49–203) | 134 (26–233) | 53 (38–111) | 61 (37–125) |
| Suonenjoki 3 | 79 (40–220) | 90 (38–190) | 49 (39–85) | 51 (35–80) |
| Multia 2 | 114 (36–186) | 160 (51–255) | 40 (27–61) | 56 (41–159) |
| Hämeenlinna | 132 (30–200) | 142 (33–278) | 48 (32–197) | 45 (22–142) |
| Vesijako | 89 (32–207) | 109 (40–195) | 42 (33–59) | 44 (33–74) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piri, T.; Saarinen, M.; Hamberg, L.; Hantula, J.; Gaitnieks, T. Efficacy of Biological and Chemical Control Agents against Heterobasidion Spore Infections of Norway Spruce and Scots Pine Stumps on Drained Peatland. J. Fungi 2023, 9, 346. https://doi.org/10.3390/jof9030346
Piri T, Saarinen M, Hamberg L, Hantula J, Gaitnieks T. Efficacy of Biological and Chemical Control Agents against Heterobasidion Spore Infections of Norway Spruce and Scots Pine Stumps on Drained Peatland. Journal of Fungi. 2023; 9(3):346. https://doi.org/10.3390/jof9030346
Chicago/Turabian StylePiri, Tuula, Markku Saarinen, Leena Hamberg, Jarkko Hantula, and Talis Gaitnieks. 2023. "Efficacy of Biological and Chemical Control Agents against Heterobasidion Spore Infections of Norway Spruce and Scots Pine Stumps on Drained Peatland" Journal of Fungi 9, no. 3: 346. https://doi.org/10.3390/jof9030346