
Recombinant Mitochondrial Genomes Reveal Recent Interspecific Hybridization between Invasive Salangid Fishes

A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, Vladivostok 690041, Russia
Life 2022, 12(5), 661; https://doi.org/10.3390/life12050661
Submission received: 12 April 2022
/
Revised: 26 April 2022
/
Accepted: 26 April 2022
/
Published: 29 April 2022
(This article belongs to the Collection Feature Papers in Animal Science)
Abstract
:The interspecific recombination of the mitochondrial (mt) genome, if not an experimental artifact, may result from interbreeding of species with broken reproductive barriers, which, in turn, is a frequent consequence of human activities including species translocations, habitat modifications, and climate change. This issue, however, has not been addressed for Protosalanx chinensis and other commercially important and, simultaneously, invasive salangid fishes that were the product of successful aquaculture in China. To assess the probability of interspecific hybridization, we analyzed the patterns of diversity and recombination in the complete mitochondrial (mt) genomes of these fishes using the GenBank resources. A sliding window analysis revealed a non-uniform distribution of the intraspecific differences in P. chinensis with four highly pronounced peaks of divergence centered at the COI, ND4L-ND4, and ND5 genes, and also at the control region. The corresponding divergent regions in P. chinensis show a high sequence similarity (99–100%) to the related salangid fishes, Neosalanx tangkahkeii and N. anderssoni. This observation suggests that the divergent regions of P. chinensis may represent a recombinant mitochondrial DNA (mtDNA) containing mt genome fragments belonging to different salangid species. Indeed, four, highly significant (pairwise homoplasy index test, P < 0.00001) signals of recombination have been revealed at coordinates closely corresponding to the divergent regions. The recombinant fragments are, however, not fixed, and different mt genomes of P. chinensis are mosaic, containing different numbers of recombinant events. These facts, along with the high similarity or full identity of the recombinant fragments between the donor and the recipient sequences, indicate a recent interspecific hybridization between P. chinensis and two Neosalanx species. Alternative hypotheses, including taxonomical misidentifications, sequence misalignments, DNA contamination, and/or artificial PCR recombinants, are not supported by the data. The recombinant fragments revealed in our study represent diagnostic genetic markers for the identification and distinguishing of hybrids, which can be used to control the invasive dynamics of hybrid salangid fishes.
1. Introduction
1.1. Taxonomy and Distribution
The family Salangidae Bleeker 1859 comprises around five to seven genera (Salanx Cuvier 1816, Leucosoma Gray 1831, Salangichthys Bleeker 1860, Hemisalanx Regan 1908, Protosalanx Regan 1908, Neosalanx Wakiya and Takahashi 1937, and Neosalangichthys Fu, Li, Xia, Lei 2012) and approximately 20 species listed in the Eschmeyer’s Catalog of Fishes [1], the World Register of Marine Species [2], and the FishBase [3]. These fishes (referred to as icefish or noodlefish) are endemic to East Asia and inhabit a wide range of marine, brackish-water, and freshwater habitats in China, Vietnam, Korean Peninsula, Japan, and Russia [4,5,6,7,8,9,10,11]. Salangids are small (reaching a maximum body length of about 21.0 cm), neotenic (adults retain some larval traits) fishes with early maturation, relatively high fecundity, and a life span of about one year [8,12,13]. Although salangids have been studied for more than 100 years (see the references above), the classification and phylogenetic relationships within this family remain poorly resolved, which could be explained in part by their neoteny, high morphological plasticity, and the homoplasious morphological characters used in previous studies [14]. The taxonomic nomenclature of salangid fishes, based on morphological characters and genetic approaches, has been subject to various changes with multiple known synonyms [15,16,17,18,19,20]. Thus, among other taxonomic reorganizations, Zhang et al. [19] showed that Protosalanx hyalocranius (Abbott, 1901) is a synonym of P. chinensis (Basilewsky, 1855), while Neosalanx taihuensis (Chen, 1956) is a synonym of N. tangkahkeii (Wu, 1931). Guo et al. [20] found Hemisalanx to be a junior synonym of Salanx. Fu et al. [18] questioned the generic status of Neosalanx and argued that it should be considered as a junior synonym of Protosalanx.
1.2. Genetics
The currently available genetic data were obtained using the mitochondrial and nuclear DNA markers [21,22,23,24,25,26,27,28,29]. Complete mt genomes were sequenced for P. hyalocranius [30], N. anderssoni [31], N. tangkahkeii [32], and other species (see Table S1 for the accession numbers and a full citation list). The complete nuclear genome was sequenced for P. chinensis [33,34,35]. Genetic diversity, population structure, and demographic history of salangid fishes were considered in a series of population genetic studies [28,36,37,38,39,40,41,42]. The problem of fish mislabeling was addressed using the CytB gene [43].
1.3. Species Transplantation and Introduction
Wild populations of salangids have markedly declined in recent years due to overexploitation, hydrotechnical constructions, and water pollution [44,45,46,47], which stimulated the development of fish aquaculture (review by Kang et al. [48]). Among salangids, the clearhead icefish P. chinensis and the short-snout icefish N. tangkahkeii are commercially important aquaculture species. In China, they have been successfully transplanted from Lake Taihu (eastern China) to hundreds of lakes and reservoirs across the country [48,49,50,51]. Special artificial breeding techniques have been developed, with fry released into lakes and reservoirs to improve aquaculture yield [52]. The introduction of icefishes has brought significant economic benefits but, simultaneously, jeopardized biodiversity in native assemblages [48,53,54]. Both P. chinensis and N. tangkahkeii are reported to be successful invasive species [47,48,53,55,56,57].
In Russia, P. chinensis was first recorded from Lake Khanka in 2006 and from the Amur River in 2008 [58], which came as a result of its introduction from Chinese water bodies [13,59,60,61,62]. P. chinensis is considered a potentially aggressive invasive species preying mainly on larvae or juveniles of indigenous fishes [63]. The same has been observed and raised serious concerns in China [47,48,64]. Indeed, P. chinensis is a successful invader: in around three decades [48] the species has substantially expanded its range from Lake Taihu northward up to the coastal waters of the Sea of Okhotsk (the Amur River estuary, Russia) (https://todaykhv.ru/news/in-areas-of-the-province/30507/ (accessed on 8 April 2022)). The distance between Lake Taihu and the Amur River estuary is almost 3000 km.
Transplantations and introductions of non-endemic species frequently cause interspecific hybridization and genetic introgression between introduced and native species with significant ecological and evolutionary consequences for native populations (for review, see [65,66,67,68,69]). A less frequent phenomenon is the interspecific hybridization between invasive species in non-native areas [70,71,72,73,74,75,76,77] with limited or no hybridization in their native ranges. In this case, the hybrid between the two invasive species shows a potential to drive the emergence of new genetic diversity, thus, reducing the effect of inbreeding depression and increasing the opportunities to rapidly adapt to new environmental conditions, which further contributes to invasion success (e.g., [77,78]).
Interspecific hybridizations create the opportunity of mtDNA paternal leakage because interspecific genetic differences (involving both mitochondrial and nuclear DNA) can be large enough to escape the mechanisms destructing paternal mitochondria and leading to heteroplasmy [79,80,81], with possible subsequent recombination (review by, e.g., [82,83,84,85]). Interspecific mtDNA recombination has been reported for a broad range of organisms including hybridizing yeasts [86], brown algae [87], conifers [88], reef building corals [89], salmonid [90,91,92] and cyprinid [93,94] fishes, and primates [95]. Thus, an analysis of mtDNA recombination can be informative for detecting interspecific hybridization, especially for those species which, as with salangids (Section 1.1), are poorly distinguished based on morphological criteria.
As a result of the intentional introduction, the possible hybridization of icefishes (along with some other negative environmental impacts) was suggested [48,53], though not verified by genetic approaches so far. The concern is not ungrounded, because, even without a gene flow, invasive hybridization will likely cause detrimental consequences for the interbreeding species [67,96]. Using sufficiently long mt genome fragments (8141 bp), we previously described a detailed architecture of recombinant events due to anthropogenic hybridization between salmonid fishes such as Siberian taimen Hucho taimen and two lenok subspecies, Brachymystax lenok and B. lenok tsinlingensis [92]. In the present work, we analyzed the patterns of nucleotide diversity in complete mt genomes of icefishes, P. chinensis and N. tangkahkeii, as well as other closely related salangid species, and detected clear signals of mtDNA recombination. We argue that the revealed mt recombinants are not experimental artifacts but reflect the interspecific hybridization between the icefishes studied. The data suggest that even a relatively high genetic divergence (up to 8.5% between N. tangkahkeii and P. chinensis) could not have provided any successful reproductive isolation between the species. The results can be useful for the development of responsible aquaculture and biodiversity conservation management practices to minimize the probability of anthropogenic hybridization that can facilitate the invasion process and threaten the resilience of native populations.
2. Materials and Methods
2.1. Mitochondrial Genomes
Complete mt genome sequences of salangid fishes were accessed from the Genetic Sequence Data Bank [97] (the National Center for Biotechnology Information; https://www.ncbi.nlm.nih.gov/; GenBank Flat File Release 244.0; see Table S1 for accession numbers). The outgroup species, including representatives of the genera Plecoglossus and Retropinna, were selected based on the previous molecular evidence of their close relationship to the family Salangidae [17,19,31] and the screening of nucleotide sequences available in GenBank.
2.2. DNA Sequence Analysis
The nucleotide sequences were aligned using the MUSCLE [98] and MAFFT v. 7 [99] software. The DnaSP v. 6 [100], PROSEQ v. 2.9 [101], and MEGA v. 7 [102] programs were used for intra- and interspecific analysis of polymorphism and divergence; MEGA v. 7 [102] was also used for basic phylogenetic analyses. Phylogenetic reconstructions were inferred from the analysis of the complete mt genomes by the maximum-likelihood methods available in IQ-TREE v. 2 [103,104,105]. The TIM2+F+I+G4 model showed the lowest Akaike Information Criterion (AIC; [106]) value (189,144.4265) and the Bayesian information criterion (BIC; [107]) score (189,529.5437); this model was selected for further phylogenetic reconstructions. The ultrafast maximum likelihood bootstrap analysis [108] consisted of 10,000 replicates. The alignments were analyzed for evidence of recombination using the pairwise homoplasy index (PHI) test [109] and various recombination detection methods provided in the RDP4 software [110,111,112,113,114,115,116,117,118]. The sliding window method (see, e.g., [119]) was used to examine the spatial distribution of polymorphism and divergence across the mt genomes studied. In this method, a window of specified length moves over the nucleotide sequence with a fixed size of step and the corresponding estimates are computed over the data in the window. The obtained estimates are assigned to the nucleotide position at the midpoint of the window. The output of the sliding window analysis can be presented graphically; the values of variation are plotted against the nucleotide position.
3. Results
3.1. Nucleotide Diversity and Divergence
A phylogram of the P. chinensis mt genomes along with those of other salangid fishes is displayed in Figure 1. The tree shows the P. chinensis specimens forming a single clade with two significantly different (100% bootstrap support) groups of sequences including (1) MW291629, KP306787, and HM106494, and (2) MH330683 and KJ499917 (Figure 1). The π value is quite high (0.0127 ± 0.0007). Despite the fact that it seems still below the range of divergence between species (e.g., [120]), it is, nevertheless, significantly higher than the level of intraspecific diversity detected for many other freshwater and marine fish species including, e.g., the taimen Hucho taimen (0.0010 ± 0.0002; [92]), the European whitefish Coregonus lavaretus (0.0025 ± 0.0003; [121]), the lake whitefish C. clupeaformis (0.0008 ± 0.0001; [121]), the houting C. oxyrinchus (0.0029 ± 0.0003; [121]), the Atlantic herring Clupea harengus (0.0061 ± 0.0008; [122]), the spotted wolffish Anarhichas minor (0.0005 ± 0.0001; [123]), and the northern wolffish Anarhichas denticulatus (0.0006 ± 0.0001; [123]).
3.2. Sliding Window Analysis
A sliding window analysis has revealed a non-uniform distribution of the nucleotide diversity values along the P. chinensis genomes characterized by four pronounced peaks centered at the (1) COI, (2) ND4L-ND4 (hereinafter referred to as ND4 for simplicity), and (3) ND5 genes, and also at the (4) control region (CR) (Figure 2a). Comparisons including different pairs of the P. chinensis genomes have revealed distinct patterns with one to four strong peaks (Figure 2b–d,f–h), or no peaks at all (Figure 2e). The π values in the valley regions are low (varying from 0 to 0.0117), but follow much higher values in the high divergence (HD) regions: 0.0417 (first peak, COI; midpoint in the alignment 5631), 0.1217 (second peak, ND4; midpoint 11,557), 0.1100 (third peak, ND5; midpoint 13,096), and 0.1083 (fourth peak, CR; midpoint 15,763). The divergence values detected at the HD regions are significantly higher than the values usually observed on the intraspecific level (see above) and approach the intergeneric values, e.g., for scorpion fishes [120,124]. A more detailed interpretation for the distribution of divergent regions is considered below (Section 3.3).
The basic local alignment search tool (BLAST) procedure [125] has revealed a surprisingly high similarity (99–100%) between P. chinensis (KJ499917) and N. tangkahkeii (KP170510) for the HD regions 1 (COI), 2 (ND4), and 4 (CR); between P. chinensis (MH330683) and N. tangkahkeii (KP170510) for the HD region 2 (ND4); and between P. chinensis (HM106494) and N. anderssoni (HM106492) for the HD region 3 (ND5). Table 1 provides values of pairwise distances (Dxy ± SE) between P. chinensis and these closely related salangid species, and shows a number of outlier values obtained for the full mt genomes and the HD regions separately.
On the intraspecific level, the pairwise distances between the full mt genomes of P. chinensis vary within a fairly broad range (0.3–1.9%), which is less pronounced (0.1–0.4%) for the mt genomes with the HD regions deleted (Table 1, last column). When the HD regions are considered separately, the pairwise distances vary in a broader range: from zero to 11.6% (ND4 and ND5 regions), between 0.7–6.4% (COI region), and up to 12.9% (CR region) (Table 1). On the interspecific level, the pairwise distance fluctuations are even more pronounced. For instance, the full mt genome divergence between P. chinensis and N. anderssoni amounts to 5.5% with a range from 0.9% (ND5 region) to 14.1% (CR region). For the P. chinensis–N. tangkahkeii pairwise comparisons, the average full mt divergence amounts to 8.5%. However, the Dxy value between these species is 85.0-fold lower (0.1%) for the ND4 HD region. Moreover, for the COI and CR HD regions, there are no differences between P. chinensis (KJ499917) and N. tangkahkeii (KP170510) (Table 1). The pairwise distances inferred from the mt genomes with the HD regions deleted (Table 1, last column) show much lower fluctuations, ranging from 4.4 to 4.5% and from 8.6 to 8.7% for the P. chinensis–N. anderssoni and P. chinensis–N. tangkahkeii comparisons, respectively.
Thus, the HD regions of the P. chinensis genomes demonstrate an unexpectedly high similarity (ND4 and ND5) or even full identity (COI and CR) to N. tangkahkeii and N. anderssoni, and can be explained by a recombination of mtDNA (Section 3.3). The four HD regions (COI, ND4, ND5, and CR) show sharply discordant phylogenetic signals between P. chinensis and the Neosalanx species. As a consequence, the position of P. chinensis is sharply different, depending on the fragments used for tree reconstruction (Figure 3). The trees inferred from the HD regions separately show P. chinensis as identical (or very similar) to N. tangkahkeii or to N. anderssoni, respectively (Figure 3a–d). In the tree without the HD regions (Figure 3e), the P. chinensis mt genomes (KJ499917, MH330683, HM106494, KP306787, and MW291629) are within a single cluster showing a value of π (0.0033 ± 0.0003) similar to that of the mt genomes of other fishes (Section 2.1). The other genera included in this analysis (Salangichthys, Plecoglossus, and Retropinna) do not show any visible discordance in the level of divergence between the HD regions and the rest of the mtDNA (Figure 3a–e). The presence or absence of the N. tangkahkeii and N. anderssoni fragments in the P. chinensis mt genomes erroneously increases or decreases the pairwise distances depending on their combination in particular comparison (Table 1). The same interpretation is suitable to explain the fluctuations in pairwise distances (Table 1).
3.3. Recombination
Based on the above-presented results, we suggest that the HD regions (Figure 2) and the respective phylogenetic inconsistencies (Figure 3) can be explained by recombination of the P. chinensis mt genome. Indeed, the PHI-test [109] has revealed very good evidence of recombination (p < 0.00001) in the alignment including the P. chinensis, N. tangkahkeii, and N. anderssoni mt genome sequences. We, therefore, have analyzed the mtDNA alignments for evidence of recombination using various recombination detection methods implemented in the RDP4 program ([110]; Table 2). Six methods have detected four recombination events in the P. chinensis KJ499917, MH330683, and HM106494 mt genomes with high statistical support (Table 2, Figure 4). The breakpoint positions (Table 2) match closely the coordinates of the HD regions (see Figure 2). The recombinant fragments are not fixed, and the recombinant mt genomes are mosaic, containing different numbers of recombinant events: three for KJ499917, one for MH330683, and one for HM106494 (Figure 4). No recombination events were detected for the P. chinensis KP306787 and MW291629 mt genomes.
The sequences involved in the recombination and the breakpoints were identified using the RDP4 suite [110] which incorporates the algorithms RDP (R), GENECONV (G), BOOTSCAN (B), MAXCHI (M), CHIMAERA (C), SISCAN (S), and 3SEQ (Q) (for references, see the Material and Methods section). For each putative recombination breakpoint, a Bonferroni correction P-value was calculated. For each recombination event involving the COI, ND4, and ND5 gene regions, and also CR, the analysis was based on the alignment including three full mitochondrial genomes, as indicated in the column “Recombination event” (the length of the recombinant fragment is in parentheses). For the ND4 gene region, two separate analyses were carried out, including the Protosalanx chinensis genomes KJ499917 (Recombinant 1) and MH330683 (Recombinant 2) as recombinant sequences with the same major and minor parents; conservative P-values are presented. The recombination events are arranged in the order as they occurred along the alignment. The breakpoint positions match closely the coordinates of the HD regions (see Figure 2). The COI recombinant fragment (P. chinensis, KJ499917) starts 73 bp downstream of the COI start codon and ends 1338 bp upstream of the COI stop codon. The ND4 recombinant fragment (P. chinensis, KJ499917 and MH330683) starts 78 bp downstream of the ND4L start codon and covers the full ND4 gene, tRNA-His, and 47 bp of the tRNA-Ser. The ND5 recombinant fragment (P. chinensis, HM106494) starts 976 bp downstream of the ND5 start codon and ends 423 bp upstream of the ND5 stop codon. The CR recombinant fragment (P. chinensis; KJ499917) starts 6 bp downstream of the beginning of the tRNA-Pro, covers the rest part of the tRNA-Pro and includes 295 bp of the CR.
For the COI and CR recombinant fragments, there is 100% identity between the recombinant (P. chinensis, KJ499917) and the minor (N. tangkahkeii, KP170510) parent sequences (Figure 5a,e). However, for the ND4 recombinant fragment, there are two mismatches at sites 10,174 and 10,185 (highlighted in bold in the alignment; Figure 5b,c) between the recombinant (P. chinensis, KJ499917, MH330683) and the minor (N. tangkahkeii, KP170510) parent sequences. Similarly, for the ND5 recombinant fragment there are four mismatches at sites 12,954, 12,991, 12,997, and 13,030 between the recombinant (P. chinensis, HM106494) and the minor parent (N. anderssoni, HM106492) sequences (Figure 5d). See also Texts S1–S3 for the alignment of the variable sites in the recombinant and parent mt genome sequences.
Taking into account the patterns of recombination revealed by the RDP4 program (Table 2, Figure 4), we can now confidently interpret the distribution of the HD regions in pairwise comparisons of the P. chinensis genomes obtained by the sliding window analysis (Figure 2) in the following way. The plot (a), inferred from the total data for the five P. chinensis mt genomes, shows four HD regions corresponding to the four recombination events detected for the three P. chinensis mt genomes: three for KJ499917 (COI, ND4, and CR), one for MH330683 (ND4), and one for HM106494 (ND5). The plots (b,c,h) showing one, two, and three HD regions, respectively, reflect the COI, ND4, and CR recombination events between the P. chinensis and N. tangkahkeii mt genomes. The plot (f) with a single HD region reflects the ND5 recombination event involving P. chinensis and N. anderssoni. The plots (d,g) with four and two HD regions, respectively, reflect the COI, ND4, ND5, and CR recombination events involving P. chinensis and both N. tangkahkeii and N. anderssoni. The sliding window plot (e) shows the variability distribution for the two sequences (KP306787 and MW291629) lacking the recombinant fragments and, correspondingly, lacking any HD regions.
4. Discussion
Recombination of the mt genome can reflect interspecific hybridization; however, as an alternative hypothesis, it can be an artifact produced by polymerase chain reaction (PCR) or other errors. Below, we consider the most relevant alternative hypothesis which can explain the results observed in the present work.
Species identification of salangid fishes remains a serious challenge (Section 1.1). Consequently, one can argue that the GenBank data for P. chinensis and the close relatives, which we used for the present analyses, are wrong. Indeed, there are some relatively rare examples of incorrect taxonomic identification of fishes detected in GenBank [126,127,128]. However, all five P. chinensis mt genomes form a single cluster; the sequence divergence between them (except recombinant fragments) is low, 0.3% (Section 2.2; Figure 1 and Figure 4e), which closely matches the values of intraspecific diversity reported for many freshwater and marine fishes (Section 2.1). Furthermore, all the P. chinensis recombinant (KJ499917, MH330683, and HM106494) and non-recombinant (KP306787 and MW291629) mt genomes show close affinity to other salangid fishes, both in our analysis (Figure 1) and other publications [31,129]. Thus, the available data suggest that taxonomical misidentification is not responsible for the evidence of recombination obtained in our work.
Even if the taxonomic identification of P. chinensis is not doubted, there is still a risk of artificial recombination generated by PCR errors if more than one template were present in the PCR. In this case, the polymerase may jump from one template to another during the PCR, thus, producing artifactual recombinants [130]. This effect could be suggested for the ND5 recombination involving the P. chinensis (HM106494) and N. anderssoni (HM106492) mt genomes, both investigated by Li et al. [31]. The multi-template PCR containing the P. chinensis DNA and N. anderssoni contamination DNA could generate PCR artifacts. However, if this recombination had occurred through PCR jumping, the exchanged regions found in two different sequences should have been exact copies of one another. An examination of the recombinant P. chinensis (HM106494) sequence has revealed no exact matches with the minor parent sequence (N. anderssoni; HM106492) for the ND5 recombinant event. In particular, there are four mutations distinguishing the recombinant and minor parent sequences (Figure 5d), which makes the “jumping PCR” hypothesis highly unlikely. A similar pattern has been revealed for the ND4 recombinant event (Figure 5b) with two mismatches along the recombinant sequence distinguishing the recombinant (P. chinensis; KJ499917, MH330683) and minor parent (N. tangkahkeii; KP170510) sequences. Thus, the imperfect homologies between the recombinants and minor parents for the ND4 and ND5 regions show that these recombinants are real and probably occurred some time ago, due to a historical hybridization between the icefishes.
The COI and CR recombinant sequences are identical to the minor parent sequences and could be explained by PCR jumping. Both recombinants are detected in the P. chinensis mt genome KJ499917 obtained by Lu et al. [30] (published online, 4 April 2014) from the Heilongjiang River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Harbin, China. The fish sample was collected from Lake Xingkai (Khanka). However, the N. tangkahkeii mt genome KP170510, which represents a minor parent mt genome for the COI and CR recombination events, was sequenced by Zhong et al. [32] (published online 19 February 2015) from the Freshwater Fisheries Research Institute of Jiangsu Province, Nanjing, China. The fish sample was collected from Lake Taihu, Jiangsu Province, China. Thus, the COI and CR recombinants cannot be results of DNA contaminations of the P. chinensis PCR amplifications by the N. tangkahkeii DNA because these mt genomes were sequenced by different researchers at a different time and in different laboratories.
Another argument against PCR jumping is exemplified by two P. chinensis mt genome sequences, KJ400017 [30] and MH330683 [129], which carry out the identical recombination event including the ND4 region (Figure 4a,b and Figure 5b,c). The recombination involves precisely the same region in both mt genomes (Table 2) which, however, were detected by different authors at different laboratories and in different years. The P. chinensis recombinant mt genome sequence KJ400017 was obtained by Lu et al. [30] (see the author’s affiliation and the fish sampling point above), but the P. chinensis recombinant sequence MH330683 was obtained by Liu et al. [129] from the Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi, China. The fish sample was collected from Lake Taihu, Wuxi City, China. It is difficult to imagine how this identical recombination could be obtained simultaneously and erroneously by different authors in different places and at different times.
It might also be suggested that the evidence of recombination obtained in the present study is caused by a problem with the tests of recombination due to, e.g., misalignments, which are a common cause of false positive recombination signals [110]. This explanation could be particularly relevant for the recombination detected within CR, which is the most variable part of the mt genome frequently characterized by multiple insertions, deletions, and the complex arrangement of tandem repeats [131]. However, the recombinant (P. chinensis, KJ499917) and minor parent (N. tangkahkeii, KP170510) sequences are 100% identical in the CR recombinant fragment, which provides fully unambiguous alignment and implies that the CR recombination that we have detected is unlikely to be the product of misalignment.
The above arguments strongly suggest that the mt genome recombinants detected in our work are not experimental errors or artifacts and, consequently, their formation requires assuming an interspecific hybridization between P. chinensis and the other two related salangid fishes, N. tangkahkeii and N. anderssoni. The necessary prerequisites for the mt genome recombination to occur, including paternal leakage and heteroplasmy, are not investigated for the salangids, but are well documented for interspecific hybrids of fishes (e.g., [94,132]) and many other organisms (reviewed by Ladoukakis and Zouros [84] and Parakatselaki et al. [85]). Mitochondrial fusion has been demonstrated in diverse organisms, from yeast to mammals including humans (review by Westermann [133]). Experiments testing the complementation between different mtDNA molecules after fusion have revealed the presence of an extensive and continuous exchange of genetic contents between mitochondria in mammalian cells [134]. Experimental data also show that mitochondria possess all the necessary molecular mechanisms for mtDNA recombination (review by Chen [135]).
The present data complement a number of examples of the mtDNA recombination reported previously for plants, fungi, protists, and animals (review in [82,84,95,135,136,137,138,139,140,141,142,143]). Estimation of hybridization events based on recombination can be considered a conservative one because, at least in animals, an overwhelming amount of mtDNA is maternally transmitted, and the paternal leakage is restricted to a very low amount [84]. The P. chinensis mtDNA recombination was detected in three out of the five mt genomes studied, thus, suggesting relatively frequent interspecific hybridization in salangid fishes, as it was detected, e.g., in hybrid swarms of other invasive species ([73]; see below).
Different species of salangid fishes frequently occur in sympatry [19,48,144]. Nevertheless, the probability of interbreeding between salangids is apparently low or negligible due to, e.g., different microhabitat preferences (such as salinity tolerances) or a shift of spawning season [9,48]. Indeed, Zhang et al. [19] reported a relatively high genetic divergence (based on the CytB gene) between salangid species and suggested that these fishes have “evolved powerful sorting mechanisms to maintain interspecific isolation” ([19], page 338). However, temporal or spatial reproductive isolation that is likely to occur in native populations (due to, e.g., habitat-based segregation at spawning grounds; [145]) can be lost in the non-native ranges, resulting in an extensive hybridization between invasive species in the absence of environmental cues present in their native range [73,78] (review by Scribner et al. [146]). Due to anthropogenic habitat disturbance, it may be that reinforcement does not produce enough efficient mechanisms to provide reproductive isolation for divergent species that naturally occur in sympatry [147].
Among salangids, P. chinensis and N. tangkahkeii exhibit the widest distribution and the highest ecological plasticity (review by Kang et al. [48]). After being introduced into a new habitat outside the natural distribution range, these species demonstrate flexible life-history traits, especially in their growth and reproductive modes [48,55], which are frequently observed in invasive species (e.g., [148,149,150,151]). The high reproductive plasticity and the lack of native environmental cues that would provide reproductive isolation may have contributed to the hybridization between P. chinensis and the Neosalanx species, as has been shown for other invasive fishes invading non-native ranges [70,71,72,73,74,75,77,78].
Interspecific hybridization between invasive species can be exemplified by one exceptionally problematic case: the introduction of silver carp (Hypophthalmichthys molitrix) and bighead carp (H. nobilis) to the complex system of braided watercourses in the Mississippi River Basin, United States [71,72,73]. These two species currently remain genetically isolated and do not crossbreed within their native ranges in China, but their introduction to a new habitat initiated an unprecedented frequency of interspecific hybridization leading to a hybrid swarm with early-generation hybrids driving the range expansion and contributing to their invasion success, which caused multiple ecological, evolutionary, and economic problems [73,77,78]. The introduction of invasive salangids into the new habitats of Lake Khanka and the Amur River Basin, located at the border between China and Russia [13,58,59,60,61,62,63], might have a similar hybridization potential, as is evidenced by the data obtained in the present work. For instance, three recombinant fragments were detected in the mt genome KJ400017 (Figure 4a) of P. chinensis sampled from Lake Khanka [30].
The genetic data obtained in the present study show clear signals of recombination in the mt genomes of P. chinensis containing four mtDNA fragments from two related salangid species, N. tangkahkeii and N. anderssoni. The recombinant fragments are not fixed, and the different mt genomes of P. chinensis are mosaic, containing a different number of recombinant events. These observations, along with the full identity of the COI and CR recombinant fragments between the donor and the recipient sequences, indicate the contemporary interspecific hybridization between P. chinensis and N. tangkahkeii, which can be at least partly explained by human-mediated activities involving the transplantation and introduction of the icefishes. Among the protein-coding mt genes, COI (along with CytB) exhibits the highest evolutionary rates, which are two–five-fold higher for the noncoding CR region [152]. These regions (COI and CR) in case of historical hybridization are expected to rapidly gain new mutations distinguishing them from the parental sequences. The ND4 and ND5 recombination events show two and four mismatches, respectively, between the recombinant and parent sequences (Figure 5b,d), which may indicate a historical hybridization between P. chinensis and the two related salangid fishes, N. tangkahkeii and N. anderssoni. Further evidence from biparentally inherited nuclear DNA is required to critically evaluate the revealed patterns and, in particular, to determine the fraction of the extant P. chinensis genome that was affected by the gene flow from the Neosalanx species, and the time when it happened.
The four recombinant fragments, including the COI, ND4, and ND5 gene regions, and also the CR, are specific for distinguishing hybrids between P. chinensis and the two Neosalanx species. Two of them, COI and CR, that show full identity between P. chinensis and N. tangkahkeii, can be used as diagnostic markers to monitor the salangids’ human-mediated hybridization dynamics and their invasion fronts. Extensive spatial and temporal genetic sampling from both native and invasive ranges, including the Amur River Basin and Lake Khanka, should be carried out to determine the full extent of the current range of pure salangid species and their hybrids, which will help to elaborate the most appropriate aquaculture regimes to minimize the negative effects of the species introduction and transplantation on the local biodiversity and ecosystem resilience.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/life12050661/s1, Table S1: GenBank accession numbers. Texts S1–S3 (format: docx): Alignments of variable sites in recombinant and parental mitochondrial genome sequences. Refs. [19,30,31,32,110,129,153,154,155,156,157,158,159,160,161,162,163,164,165] are mentioned in the Supplementary Materials.
Funding
This research received budgetary support within the framework of the Research Work “World Ocean biodiversity: taxonomy, barcoding, phylogenetics, reproductive and evolutionary biology, biogeography.” (State registry no. 121082600036-9, Ministry of Science and Higher Education of the Russian Federation), A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, Vladivostok, Russia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author greatly appreciates encouragement and help from Elena I. Balakireva and Larisa G. Erofeeva. Special thanks are due to D.P. Martin for his valuable comments concerning recombination breakpoints. E.P. Shvetsov (A.V. Zhirmunsky National Scientific Center of Marine Biology FEB RAS, Vladivostok, Russia) kindly proofread the manuscript. The author is also very grateful to two anonymous reviewers for helpful criticism and suggestions on the previous versions of the manuscript.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. The author alone is responsible for the content and writing of the paper. The author acknowledges no financial interest or benefit from the direct applications of this research. The data analysis and manuscript preparation were conducted at the A.V. Zhirmunsky National Scientific Center of Marine Biology, Vladivostok, Russia.
References
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 16 March 2022).
- WoRMS Editorial Board. World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 16 March 2022).
- Froese, R.; Pauly, D. (Eds.) FishBase. World Wide Web Electronic Publication; Leibniz Institute of Marine Sciences: Kiel, Germany; Available online: http://www.fishbase.org/ (accessed on 16 March 2022).
- Regan, C.T. A synopsis of the fishes of the subfamily Salanginae. Ann. Mag. Nat. Hist. 1908, 2, 444–446. [Google Scholar] [CrossRef] [Green Version]
- Wakjya, Y.; Takahasi, N. Salangidae of Japan. Zool. Mag. Tokyo 1913, 25, 551–555. [Google Scholar]
- Wakiya, Y.; Takahasi, N. Studies on fishes of the family Salangidae. J. Coll. Agric. Tokyo Imp. Univ. 1937, 14, 265–296. [Google Scholar]
- Fang, P.W. Study on the fishes referring to Salangidae of China. Sinensia 1934, 4, 231–268. [Google Scholar]
- Roberts, T.R. Skeletal anatomy and classification of the neotenic Asian Salmoniform superfamily Salangoidea (icefishes or noodlefishes). Proc. Calif. Acad. Sci. 1984, 43, 179–220. [Google Scholar]
- Dou, S.; Chen, D. Taxonomy, biology and abundance of icefishes, or noodlefishes (Salangidae), in the Yellow River estuary of the Bohai Sea, China. J. Fish Biol. 1994, 45, 737–748. [Google Scholar]
- Zhang, Y.; Qiao, X. Study on phylogeny and zoogeography of fishes of the family Salangidae. Acta Zool. Taiwanica 1994, 5, 95–113. [Google Scholar]
- Saruwatari, T.; Oohara, I.; Kobayashi, T. Salangid fishes: Their past, present and future. Fish. Sci. 2002, 68, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Lin, R. Occurrence of neoteny in Hemisalanx and its evolutionary significance. Acta Hydrobiol. Sin. 1965, 5, 239–245. [Google Scholar]
- Tang, F.J.; Liu, W.; Wang, J.; Froese, R.; Xie, S. Growth, length-weight relationship and biological information on the clearhead icefish (Protosalanx hyalocranius Abbott, 1901) in Lake Khanka (Xingkai). J. Appl. Ichtgyol. 2012, 28, 842–844. [Google Scholar] [CrossRef]
- Ilves, K.L.; Taylor, E.B. Molecular resolution of the systematics of a problematic group of fishes (Teleostei: Osmeridae) and evidence for morphological homoplasy. Mol. Phylogenet. Evol. 2009, 50, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L. A taxonomic study on the Chinese icefishes of the genus Neosalanx (Pisces: Salangidae), with description of a new species from the Lake Taihu. Zool. Res. 1987, 8, 277–286. [Google Scholar]
- Xie, Y.H.; Xie, H. Classification, distribution, and population ecology of Salangidae fishes. Chin. J. Fish. 1997, 10, 11–19. [Google Scholar]
- Fu, C.; Luo, J.; Wu, J.; López, J.A.; Zhong, Y.; Lei, G.; Chen, J. Phylogenetic relationships of salangid fishes (Osmeridae, Salanginae) with comments on phylogenetic placement of the salangids based on mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2005, 35, 76–84. [Google Scholar] [CrossRef]
- Fu, C.; Guo, L.; Xia, R.; Li, J.; Lei, G. A multilocus phylogeny of Asian noodlefishes Salangidae (Teleostei: Osmeriformes) with a revised classification of the family. Mol. Phylogenet. Evol. 2012, 62, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.; Xu, M.; Takita, T.; Wei, F. Molecular phylogeny of icefish Salangidae based on complete mtDNA cytochrome b sequences, with comments on estuarine fish evolution. Biol. J. Linn. Soc. 2007, 91, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Li, J.; Wang, Z.; Fu, C. Phylogenetic relationships of noodle-fishes (Osmeriformes: Salangidae) based on four mitochondrial genes. Acta Hydrobiol. Sin. 2011, 35, 449–459. [Google Scholar]
- Xia, D.Q.; Cao, Y.; Wu, T.T.; Yang, H. Genetic structure of Protosalanx hyalocrsanius, Neosalanx tangkahkeii and Neosalanx oligodontis populations in Lake Taihu. J. Fish. Sci. China 1999, 23, 254–260. [Google Scholar]
- Xia, D.Q.; Cao, Y.; Wu, T.T.; Yang, H. Study on lineages of Protosalanx chinensis, Neosalanx taihuensis and N. oligodontis in Taihu Lake with RAPD technique. J. Fish. Sci. China 2000, 7, 12–15. [Google Scholar]
- Gao, T.; Chen, X.; Han, Z.; Liu, J.; Zhang, Y. Analysis of Protosalanx hyalocranius and Salangichthys microdon mitochondrial cytochrome b and part of gene 16S rRNA sequences. J. Ocean Univ. Chin. 2004, 34, 791–794. [Google Scholar]
- Zhang, Y.; Dong, S.; Wang, Q.; Sun, Z. The genetic composition of isozymes of Protosalanx hyalocranius and Neosalanx tangkahkeii. J. Dalian Fish. Univ. 2005, 20, 111–115. [Google Scholar]
- Zhang, J.; Wang, C.; Wang, S.; Wang, C. Cloning and sequence analysis of gene 18S rRNA of Macrura reevesii, Neosalanx tangkahkeii and Protosalanx hyalocranius. J. Wuhan Univ. 2010, 56, 87–92. [Google Scholar]
- Zhao, L.; Gao, G.; Zhang, X. Preliminary study on screening of Protosalanx hyalocranius microsatellite markers by magnetic beads enrichment method. J. Suzhou Univ. 2010, 25, 24–26. [Google Scholar]
- Xiao, M.; Bao, F.; Cui, F.; Wang, S. Mitochondrial DNA variation of Protosalanx hyalocranius in the Huaihe River. Anim. Genet. 2016, 47, 388–389. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Li, D.; Liu, W.; Li, H.; Wang, J.; Lu, C.; Jin, X. Evolutionary tendency of clearhead icefish Protosalanx hyalocranius inferring mitochondrial DNA variation analyses in Amur (Heilongjiang) River catchment, China. Int. J. Agric. Biol. 2018, 20, 2329–2334. [Google Scholar]
- Zhang, J.; Waryani, B.; Zhou, Q. Isolation and characterization of microsatellite loci from an icefish, Neosalanx tangkahkeii (Osmeriformes, Salangidae). Pakistan J. Zool. 2019, 51, 1853–1857. [Google Scholar] [CrossRef]
- Lu, C.; Gu, Y.; Li, C.; Cheng, L.; Sun, X.; Tang, F. Complete mitochondrial genome of clearhead icefish Protosalanx hyalocranius (Salmoniformes: Salangidae). Mitochondrial DNA 2016, 27, 514–515. [Google Scholar] [CrossRef]
- Li, J.; Xia, R.; McDowall, R.M.; Lopez, J.A.; Lei, G.; Fu, C. Phylogenetic position of the enigmatic Lepidogalaxias salamandroides with comment on the orders of lower euteleostean fishes. Mol. Phylogenet. Evol. 2010, 57, 932–936. [Google Scholar] [CrossRef]
- Zhong, L.; Wang, M.; Li, D.; Tang, S.; Zhang, T.; Bian, W.; Chen, X. Complete mitochondrial genome of Chinese icefish Neosalanx tangkahkeiis (Salmoniformes, Salangidae): Comparison reveals Neosalanx taihuensis not a valid name. Mitochondrial DNA Part A DNA Map. Seq. Anal. 2016, 27, 3303–3305. [Google Scholar] [CrossRef]
- Liu, K.; Xu, D.; Li, J.; Bian, C.; Duan, J.; Zhou, Y.; Zhang, M.; You, X.; You, Y.; Chen, J.; et al. Whole genome sequencing of Chinese clearhead icefish, Protosalanx hyalocranius. GigaScience 2017, 6, giw012. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xing, T.; Liu, J. Genome-wide association analyses based on whole genome sequencing of Protosalanx hyalocranius provide insights into sex determination of Salangid fishes. Mol. Ecol. Res. 2020, 20, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qi, J.; Shi, F.; Pan, H.; Liu, M.; Tian, R.; Geng, Y.; Li, H.; Qu, Y.; Chen, J.; et al. Insights into the evolution of neoteny from the genome of the Asian icefish Protosalanx chinensis. iScience 2020, 23, 101267. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Doiuchi, R.; Nakabo, T. Molecular and morphological differences between two geographic populations of Salanx ariakensis (Salangidae) from Korea and Japan. Ichthyol. Res. 2006, 53, 52–62. [Google Scholar] [CrossRef]
- Kim, J.K.; Kai, Y.; Nakabo, T. Genetic diversity of Salanx ariakensis (Salangidae) from Korea and Japan inferred from AFLP. Ichthyol. Res. 2007, 54, 416–419. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, J.; Liu, Z.; Funk, S.M.; Wei, F.; Xu, M.; Li, M. Complex population genetic and demographic history of the Salangid, Neosalanx taihuensis, based on cytochrome b sequences. BMC Evol. Biol. 2008, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, J.; Liu, Z.J.; Xu, M.Q.; Li, M. Population genetic structure and demographic history of Neosalanx jordani based on cytochrome b sequences. J. Biodivers. Sci. 2010, 18, 251–261. [Google Scholar]
- Zhao, L.; Zhang, J.; Liu, Z.; Li, M. Multiple unrelated founding events for the long-distance Pleistocene dispersal of the Salangid, Neosalanx taihuensis: A general demographic model for inshore-orientated freshwater fish. Mol. Phylogenet. Evol. 2011, 58, 142–147. [Google Scholar] [CrossRef]
- Hua, X.; Wang, W.; Yin, W.; He, Q.; Jin, B.; Li, J.; Chen, J.; Fu, C. Phylogeographical analysis of an estuarine fish, Salanx ariakensis (Osmeridae: Salanginae) in the north-western Pacific. J. Fish. Biol. 2009, 75, 354–367. [Google Scholar] [CrossRef]
- Liu, H.; Li, C.; Xiong, F. Population genetic structure of Neosalanx taihuensis between invasive and original areas revealed by microsatellite DNA. J. Fish. China 2016, 40, 1521–1530. [Google Scholar]
- Armani, A.; Castigliego, L.; Tinacci, L.; Gianfaldoni, D.; Guidi, A. Molecular characterization of icefish, (Salangidae family), using direct sequencing of mitochondrial cytochrome b gene. Food Control 2011, 22, 888–895. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, C.; Lei, G. China icefish biodiversity and its protection strategies. Biodivers. Sci. 2002, 10, 416–424. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, C.; Hu, H.; Zhou, Y.; Xu, C.; Lei, G. Freshwater icefishes (Salangidae) in the Yangtze River basin of China: Spatial distribution patterns and environmental determinants. Environ. Biol. Fish. 2005, 73, 253–262. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, J.; Xu, C.; Lei, G. Degradation of icefishes (Salangidae) in the Yangtze River basin of China: Threats and strategies. Environ. Biol. Fish. 2009, 86, 109–117. [Google Scholar] [CrossRef]
- Tang, F.; Gao, W.; Li, H.; Liu, W. Biology and fishery ecology of Protosalanx chinensis: A review. J. Fish. China 2020, 44, 2100–2111. [Google Scholar]
- Kang, B.; Deng, J.; Wang, Z.; Zhang, J. Transplantation of icefish (Salangidae) in China: Glory or disaster? Rev. Aquacult. 2015, 7, 13–27. [Google Scholar] [CrossRef]
- Gu, L.W. The national status of the salangids introduction from Taihu Lake. Modern Fish. Inform. 1998, 13, 23–25. [Google Scholar]
- Hu, C.L.; Liu, J.S.; Peng, J.H.; Yu, F.H. Salangids in China and their ecological management principles for transplantation. J. Lake Sci. 2001, 13, 204–210. [Google Scholar] [CrossRef]
- Li, S.; Wang, D.; Xu, Y.; Luo, M.; Jiang, G. China’s Protosalanx hyalocranius transplantation and proliferation overview and development countermeasures. J. Zhanjiang Ocean Univ. 2002, 22, 78–82. [Google Scholar]
- Wang, Y.; Chen, J.; Wang, H.; Zhang, K.; Li, H. Study on the technique of artificial propagation of large ice fish (Protosalanx hyalocranius). J. Aquacult. 2019, 12, 18. [Google Scholar]
- Kang, B.; Huang, X.; Li, J.; Liu, M.; Guo, L.; Han, C.-C. Inland fisheries in China: Past, present, and future. Rev. Fish. Sci. Aquac. 2017, 25, 270–285. [Google Scholar] [CrossRef]
- Ding, C.; Jiang, X.; Xie, Z.; Brosse, S. Seventy-five years of biodiversity decline of fish assemblages in Chinese isolated plateau lakes: Widespread introductions and extirpations of narrow endemics lead to regional loss of dissimilarity. Divers. Distrib. 2017, 23, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.-Y.; Ye, S.-W.; Li, Z.-J.; Zhang, T.-L.; Yuan, J.; Guo, Z.-Q.; Tang, J.-F.; Liu, J.-S. Growth and reproduction of the non-native icefish Neosalanx taihuensis Chen, 1956 (Salangidae) in a plateau lake, southwestern China. J. Appl. Ichthyol. 2014, 30, 862–869. [Google Scholar] [CrossRef]
- Huang, G.; Wang, Q.; Du, X.; Feng, K.; Ye, S.; Yuan, J.; Liu, J.; Li, Z.; De Silva, S.S. Modeling trophic interactions and impacts of introduced icefish (Neosalanx taihuensis Chen) in three large reservoirs in the Yangtze River basin, China. Hydrobiologia 2020, 847, 3637–3657. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Tang, W.; Sun, Z.; Pan, B. Non-native freshwater fish species in the Yellow River Basin: Origin, distribution and potential risk. Environ. Biol. Fish. 2021, 104, 253–264. [Google Scholar] [CrossRef]
- Butova, E.V.; Novomodny, G.V. Fresh water clearhead icefish Protosalanx hyalogranius (Aabbott, 1901) from the Amur river. Vladimir Ya. Levanidov’s Bienn. Meml. Meet. 2014, 6, 124–133. [Google Scholar]
- Kong, L.; Zou, M.; Zhang, Z.; Zhang, Y. Status of clearhead icefish Protosalanx hyalocranius introduction in Heilongjiang province and the developing countermeasures. Heilongjiang Fish. 2007, 122, 27–28. [Google Scholar]
- Tang, F.J.; Liu, W.; Wang, J.L.; Xie, S.G. Fish composition in Lake Xingkai (Khanka) and Lake Mini-Xingkai. Chin. J. Fish. 2011, 24, 40–47. [Google Scholar]
- Tang, F.; Liu, W.; Wang, J.; Brown, A. On the occurrence of the clearhead icefish, Protosalanx hyalocranius (Salmoniformes: Salangidae), in the Songhua River, the largest tributary of Amur River. J. Appl. Ichthyol. 2013, 29, 237–238. [Google Scholar] [CrossRef]
- Tang, F.J.; Liu, W.; Wang, J.L.; Henne, J.; Cui, X.S. Clearhead icefish (Protosalanx hyalocranius Abbott, 1901) (Salmoniformes, Salangidae), a new non-native species has established a population in the Amur River, China. J. Appl. Ichthyol. 2015, 31, 177–179. [Google Scholar] [CrossRef]
- Svirskiy, V.G.; Barabanshchikov, E.I. Biological invasions as an element of anthropogenic pressure upon the aquatic biotic communities in Lake Khanka. Rus. J. Biol. Invas. 2009, 1, 21–25. [Google Scholar] [CrossRef]
- Wang, X.; Xiang, J.; Liu, J.; Liu, M.; Wu, L.; Murphy, B.R.; Xie, S. Reduced growth and reproductive investment of Hemiculter leucisculus (Cyprinidae) in a reservoir with introduced icefish Neosalanx taihuensis (Salangidae). Environ. Biol. Fish. 2013, 96, 895–903. [Google Scholar] [CrossRef]
- Rhymer, J.M.; Simberloff, D. Extinction by hybridization and introgression. Annu. Rev. Ecol. Syst. 1996, 27, 83–109. [Google Scholar] [CrossRef]
- Simberloff, D. Hybridization between native and introduced wildlife species: Importance for conservation. Wildlife Biol. 1996, 2, 143–150. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Leary, R.F.; Spruell, P.; Wenburg, J.K. The problems with hybrids: Setting conservation guidelines. Trends Ecol. Evol. 2001, 16, 613–622. [Google Scholar] [CrossRef]
- Vitule, J.R.S.; Freire, C.A.; Simberloff, D. Introduction of non- native freshwater fish can certainly be bad. Fish Fish. 2009, 10, 98–108. [Google Scholar] [CrossRef]
- Crispo, E.; Moore, J.-S.; Lee-Yaw, J.A.; Gray, S.M.; Haller, B.C. Broken barriers: Human-induced changes to gene flow and introgression in animals: An examination of the ways in which humans increase genetic exchange among populations and species and the consequences for biodiversity. BioEssays 2011, 33, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Hayden, B.; Pulcini, D.; Kelly-Quinn, M.; O’Grady, M.; Caffrey, J.; McGrath, A.; Mariani, S. Hybridization between two cyprinid fishes in a novel habitat: Genetics, morphology and life-history traits. BMC Evol. Biol. 2010, 10, 169. [Google Scholar] [CrossRef] [Green Version]
- Lamer, J.T.; Dolan, C.R.; Petersen, J.L.; Chick, J.H.; Epifanio, J.M. Introgressive hybridization between Bighead Carp and Silver Carp in the Mississippi and Illinois Rivers. N. Am. J. Fish. Manag. 2010, 30, 1452–1461. [Google Scholar] [CrossRef]
- Lamer, J.T.; Sass, G.G.; Boone, J.Q.; Arbieva, Z.H.; Green, S.J.; Epifanio, J.M. Restriction site-associated DNA sequencing generates high-quality single nucleotide polymorphisms for assessing hybridization between bighead and silver carp in the United States and China. Mol. Ecol. Res. 2014, 14, 79–86. [Google Scholar] [CrossRef]
- Lamer, J.T.; Ruebush, B.C.; Arbieva, Z.H.; McClelland, M.A.; Epifanio, J.M.; Sass, G.G. Diagnostic SNPs reveal widespread introgressive hybridization between introduced bighead and silver carp in the Mississippi River Basin. Mol. Ecol. 2015, 24, 3931–3943. [Google Scholar] [CrossRef]
- Haynes, G.D.; Gongora, J.; Gilligan, D.M.; Grewe, P.; Moran, C.; Nicholas, F.W. Cryptic hybridization and introgression between invasive Cyprinid species Cyprinus carpio and Carassius auratus in Australia: Implications for invasive species management. Anim. Conserv. 2012, 15, 83–94. [Google Scholar] [CrossRef]
- Lindner, K.; Cerwenka, A.F.; Brandner, J.; Gertzen, S.; Borcherding, J.; Schliewen, U.K. First evidence for interspecific hybridization between invasive goby species Neogobius fluviatilis and Neogobius melanostomus (Teleostei: Gobiidae: Benthophilinae). J. Fish Biol. 2013, 82, 2128–2134. [Google Scholar] [CrossRef] [PubMed]
- Chouvenc, T.; Helmick, E.E.; Su, N.-Y. Hybridization of two major termite invaders as a consequence of human activity. PLoS ONE 2015, 10, e0120745. [Google Scholar] [CrossRef] [PubMed]
- Coulter, A.A.; Brey, M.K.; Lamer, J.T.; Whitledge, G.W.; Garvey, J.E. Early generation hybrids may drive range expansion of two invasive fishes. Freshw. Biol. 2020, 65, 716–730. [Google Scholar] [CrossRef]
- Wang, J.; Gaughan, S.; Lamer, J.T.; Deng, C.; Hu, W.; Wachholtz, M.; Qin, S.; Nie, H.; Liao, X.; Ling, Q.; et al. Resolving the genetic paradox of invasions: Preadapted genomes and postintroduction hybridization of bigheaded carps in the Mississippi River Basin. Evol. Appl. 2020, 13, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Kondo, R.; Satta, Y.; Matsuura, E.T.; Takahata, N.; Chigusa, S.I. Incomplete maternal transmission of mitochondrial DNA in Drosophila. Genetics 1990, 126, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Gyllensten, U.; Wharton, D.; Josefsson, A.; Wilson, A.C. Paternal inheritance of mitochondrial DNA in mice. Nature 1991, 352, 255–257. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, H.; Hayashi, J.; Takahama, S.; Taya, C.; Lindahl, K.F.; Yonekawa, H. Elimination of paternal mitochondrial DNA in intraspecific crosses during early mouse embryogenesis. Proc. Natl. Acad. Sci. USA 1995, 92, 4542–4546. [Google Scholar] [CrossRef] [Green Version]
- Rokas, A.; Ladoukakis, E.; Zouros, E. Animal mitochondrial DNA recombination revisited. Trends Ecol. Evol. 2003, 18, 411–417. [Google Scholar] [CrossRef]
- Kmiec, B.; Woloszynska, M.; Janska, H. Heteroplasmy as a common state of mitochondrial genetic information in plants and animals. Curr. Genet. 2006, 50, 149–159. [Google Scholar] [CrossRef]
- Ladoukakis, E.D.; Zouros, E. Evolution and inheritance of animal mitochondrial DNA: Rules and exceptions. J. Biol. Res. 2017, 24, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parakatselaki, M.-E.; Ladoukakis, E.D. mtDNA heteroplasmy: Origin, detection, significance, and evolutionary consequences. Life 2021, 11, 633. [Google Scholar] [CrossRef] [PubMed]
- Poláková, S.B.; Lichtner, Ž.; Szemes, T.; Smolejová, M.; Sulo, P. Mitochondrial DNA duplication, recombination, and introgression during interspecific hybridization. Sci. Rep. 2021, 11, 12726. [Google Scholar] [CrossRef] [PubMed]
- Balakirev, E.S.; Krupnova, T.N.; Ayala, F.J. DNA variation in the phenotypically-diverse brown alga Saccharina japonica. BMC Plant Biol. 2012, 12, 108. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo-Correa, J.P.; Bousquet, J. Mitochondrial genome recombination in the zone of contact between two hybridizing conifers. Genetics 2005, 171, 1951–1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banguera-Hinestroza, E.; Sawall, Y.; Al-Sofyani, A.; Mardulyn, P.; Fuertes-Aguilar, J.; Cárdenas-Henao, H.; Jimenez-Infante, F.; Voolstra, C.R.; Flot, J.-F. mtDNA recombination indicative of hybridization suggests a role of the mitogenome in the adaptation of reef-building corals to extreme environments. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Ciborowski, K.L.; Consuegra, S.; De Leániz, C.G.; Beaumont, M.A.; Wang, J.; Jordan, W.C. Rare and fleeting: An example of interspecific recombination in animal mitochondrial DNA. Biol. Lett. 2007, 3, 554–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilgrim, B.L.; Perry, R.C.; Barron, J.L.; Marshall, H.D. Nucleotide variation in the mitochondrial genome provides evidence for dual routes of postglacial recolonization and genetic recombination in the northeastern brook trout (Salvelinus fontinalis). Genet. Mol. Res. 2012, 11, 3466–3481. [Google Scholar] [CrossRef] [Green Version]
- Balakirev, E.S.; Romanov, N.S.; Mikheev, P.B.; Ayala, F.J. Mitochondrial DNA variation and introgression in Siberian taimen Hucho taimen. PLoS ONE 2013, 8, e71147. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Liu, S.; Liu, Y. Evidence for recombination of mitochondrial DNA in triploid crucian carp. Genetics 2006, 172, 1745–1749. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Jiao, N.; Zhao, L.; Zhang, M.; Zhou, P.; Huang, X. Evidence for the paternal mitochondrial DNA in the crucian carp-like fish lineage with hybrid origin. Sci. China Life Sci. 2019, 63, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Piganeau, G.; Gardner, M.; Eyre-Walker, A. A broad survey of recombination in animal mitochondria. Mol. Biol. Evol. 2004, 21, 2319–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simberloff, D. Invasive Species. In Conservation Biology for All; Sodhi, N.S., Ehrlich, P.R., Eds.; Oxford University Press Inc.: New York, NY, USA, 2010; pp. 131–152. [Google Scholar]
- The National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 16 March 2022).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Filatov, D.A. PROSEQ: A software for preparation and evolutionary analysis of DNA sequence data sets. Mol. Ecol. Notes 2002, 2, 621–624. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Schwarz, G.E. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Bruen, T.C.; Philippe, H.; Bryant, D. A simple and robust statistical test for detecting the presence of recombination. Genetics 2006, 172, 2665–2681. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Rybicki, E. RDP: Detection of recombination amongst aligned sequences. Bioinformatics 2000, 16, 562–563. [Google Scholar] [CrossRef]
- Martin, D.P.; Posada, D.; Crandall, K.A.; Williamson, C. A modified bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. Aids Res. Hum. Retrovir. 2005, 21, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Bredell, H.; Martin, D.P.; Van Harmelen, J.; Varsani, A.; Sheppard, H.W.; Donovan, R.; Gray, C.M.; Williamson, C.; Team, H.S. HIV type 1 subtype C gag and nef diversity in southern Africa. Aids Res. Hum. Retrovir. 2007, 23, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible emergence of new geminiviruses by frequent recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.M. Analyzing the mosaic structure of genes. J. Mol. Evol. 1992, 34, 126–129. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, M.J.; Armstrong, J.S.; Gibbs, A.J. Sister-Scanning: A Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformatics 2000, 16, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Boni, M.F.; Posada, D.; Feldman, M.W. An exact nonparametric method for inferring mosaic structure in sequence triplets. Genetics 2006, 176, 1035–1047. [Google Scholar] [CrossRef] [Green Version]
- Hudson, R.R.; Kaplan, N.L. The coalescent process in models with selection and recombination. Genetics 1988, 120, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Kartavtsev, Y.P.; Sharina, S.N.; Goto, T.; Balanov, A.; Hanzawa, N. Sequence diversity at cytochrome oxidase 1 (Co1) gene among sculpins (Scorpaeniformes, Cottidae) and some other scorpionfish of Russia Far East with phylogenetic and taxonomic insights. Genes Genom. 2009, 31, 183–197. [Google Scholar] [CrossRef]
- Jacobsen, M.W.; Hansen, M.M.; Orlando, L.; Bekkevold, D.; Bernatchez, L.; Willerslev, E.; Gilbert, M.T.P. Mitogenome sequencing reveals shallow evolutionary histories and recent divergence time between morphologically and ecologically distinct European whitefish (Coregonus spp.). Mol. Ecol. 2012, 21, 2727–2742. [Google Scholar] [CrossRef]
- Teacher, A.G.; André, C.; Merilä, J.; Wheat, C.W. Whole mitochondrial genome scan for population structure and selection in the Atlantic herring. BMC Evol. Biol. 2012, 12, 248. [Google Scholar] [CrossRef] [Green Version]
- Lait, L.A.; Carr, S.M. Intraspecific mitogenomics of three marine species-at-risk: Atlantic, spotted, and northern wolffish (Anarhichas spp.). Genome 2018, 61, 625–634. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Kravchenko, A.Y.; Semenchenko, A.A. Genetic evidence for a mixed composition of the genus Myoxocephalus (Cottoidei: Cottidae) necessitates generic realignment. Genes 2020, 11, 1071. [Google Scholar] [CrossRef]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Saveliev, P.A.; Ayala, F.J. Complete mitochondrial genomes of the Cherskii’s sculpin Cottus czerskii and Siberian taimen Hucho taimen reveal GenBank entry errors: Incorrect species identification and recombinant mitochondrial genome. Evol. Bioinform. 2017, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleinik, A.G.; Skurikhina, L.A.; Kukhlevsky, A.D. Clarification of taxonomic assignment of smelt complete mitochondrial genome: GenBank accession number KP281293.1 (NC_026566.1). Mitochondrial DNA Part B 2019, 4, 1696–1697. [Google Scholar] [CrossRef]
- Sangster, G.; Luksenburg, J.A. The published complete mitochondrial genome of the milk shark (Rhizoprionodon acutus) is a misidentified Pacific spadenose shark (Scoliodon macrorhynchos) (Chondrichthyes: Carcharhiniformes). Mitochondrial DNA Part B 2021, 6, 828–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, C.; Chen, J. The mitochondrial genome of clearhead icefish Protosalanx chinensis (Salmoniformes: Salangidae). Mitochondrial DNA Part B 2018, 3, 774–775. [Google Scholar] [CrossRef] [Green Version]
- Pääbo, S.; Irwin, D.M.; Wilson, A.C. DNA damage promotes jumping between templates during enzymatic amplification. J. Biol. Chem. 1990, 265, 4718–4721. [Google Scholar] [CrossRef]
- Lee, W.-J.; Conroy, L.; Howell, W.H.; Kocher, T. Structure and evolution of teleost mitochondrial control regions. J. Mol. Evol. 1995, 41, 54–66. [Google Scholar] [CrossRef]
- Chu, Z.; Guo, W.; Hu, W.; Mei, J. Delayed elimination of paternal mtDNA in the interspecific hybrid of Pelteobagrus fulvidraco and Pelteobagrus vachelli during early embryogenesis. Gene 2019, 704, 1–7. [Google Scholar] [CrossRef]
- Westermann, B. Mitochondrial membrane fusion. Biochim. Biophys. Acta 2003, 1641, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Isobe, K.; Nakada, K.; Hayashi, J.-I. Human cells are protected from mitochondrial dysfunction by complementation of DNA products in fused mitochondria. Nat. Genet. 2001, 28, 272–275. [Google Scholar] [CrossRef]
- Chen, X.J. Mechanism of homologous recombination and implications for aging-related deletions in mitochondrial DNA. Microbiol. Mol. Biol. Rev. 2013, 77, 476–496. [Google Scholar] [CrossRef] [Green Version]
- Ladoukakis, E.; Zouros, E. Recombination in animal mitochondrial DNA: Evidence from published sequences. Mol. Biol. Evol. 2001, 18, 2127–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballard, J.W.O.; Whitlock, M.C. The incomplete natural history of mitochondria. Mol. Ecol. 2004, 13, 729–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis-Rogers, N.; Crandall, K.A.; Posada, D. Evolutionary analyses of genetic recombination. In Dynamical Genetics; Parisi, V., De Fonzo, V., Aluffi-Pentini, F., Eds.; Research Signpost: Kerala, India, 2004; pp. 49–78. ISBN 81-7736-231-3. [Google Scholar]
- Barr, C.M.; Neiman, M.; Taylor, D.R. Inheritance and recombination of mitochondrial genomes in plants, fungi and animals. New Phytol. 2005, 168, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Tsaousis, A.D.; Martin, D.P.; Ladoukakis, E.D.; Posada, D.; Zouros, E. Widespread recombination in published animal mtDNA sequences. Mol. Biol. Evol. 2005, 22, 925–933. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Wolff, J.N.; Pierson, M.; Gemmell, N.J. Revealing the hidden complexities of mtDNA inheritance. Mol. Ecol. 2008, 17, 4925–4942. [Google Scholar] [CrossRef]
- Galtier, N.; Nabholz, B.; Glémin, S.; Hurst, G.D.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- Mazunin, I.O.; Levitskii, S.A.; Patrushev, M.V.; Kamenski, P.A. Mitochondrial matrix processes. Biochemistry 2015, 80, 1418–1428. [Google Scholar] [CrossRef]
- Chen, N.S. On the salangid fishes of Lake Taihu. Acta Hydrobiol. Sin. 1956, 2, 324–335. [Google Scholar]
- Buehrens, T.W.; Glasgow, J.; Ostberg, C.O.; Quinn, T.P. Spatial segregation of spawning habitat limits hybridization between sympatric native steelhead and coastal cutthroat trout. Transact. Amer. Fish. Soc. 2013, 142, 221–233. [Google Scholar] [CrossRef]
- Scribner, K.T.; Page, K.S.; Bartron, M.L. Hybridization in freshwater fishes: A review of case studies and cytonuclear methods of biological inference. Rev. Fish Biol. Fish. 2000, 10, 293–323. [Google Scholar] [CrossRef]
- Hasselman, D.J.; Argo, E.E.; McBride, M.C.; Bentzen, P.; Schultz, T.F.; Perez-Umphrey, A.A.; Palkovacs, E.P. Human disturbance causes the formation of a hybrid swarm between two naturally sympatric fish species. Mol. Ecol. 2014, 23, 1137–1152. [Google Scholar] [CrossRef] [PubMed]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Novomeská, A.; Kováč, V. 2009: Life-history traits of non-native black bullhead Ameiurus melas with comments on its invasive potential. J. Appl. Ichthyol. 2009, 25, 79–84. [Google Scholar] [CrossRef]
- Lee, A.L.; Dafforn, K.A.; Hutchings, P.A.; Johnston, E.L. Reproductive strategy and gamete development of an invasive fanworm, Sabella spallanzanii (Polychaeta: Sabellidae), a field study in Gulf St Vincent, South Australia. PLoS ONE 2018, 13, e0200027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A. Evolution of mitochondrial DNA in fishes. In Biochemistry and Molecular Biology of Fishes. Molecular Biology Frontiers; Hochachka, P.W., Mommsen, T.P., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1993; Volume 2, pp. 1–38. [Google Scholar]
- Zhang, J.; Ding, M.; Qi, J.; Chen, H.; Zhang, B. Complete mitochondrial genome of Hemisalanx brachyrostralis (Osteichthyes: Salangidae). Mitochondrial DNA Part A 2016, 27, 836–837. [Google Scholar] [CrossRef]
- Xiong, F.; Zhai, D.; Yu, J.; Chen, Y.; Wang, Y.; Liu, H. The complete mitochondrial genome of the Chinese noodlefish Leucosoma chinensis and phylogenetic analysis of Salangidae (Osteichthyes: Osmeriformes). Mitochondrial DNA Part B 2021, 6, 176–177. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, H.; Yao, J.; Sun, K.; Wang, W.; Liu, H.; Yang, D.; Zhang, F.; Xiong, F. Complete mitochondrial genome of the Salangid icefish Neosalanx taihuensis (Actinopterygii: Osmeriformes: Salangidae). Mitochondrial DNA Part B 2018, 3, 1040–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Sui, Z.; Liu, K.; Liu, Y. The complete mitochondrial DNA sequence of Linyi large icefish (Protosalanx chinensis). GenBank 2021. submitted for publication. [Google Scholar]
- Zhang, J.F.; Rui, C.F. The complete mitochondrial genome of Neosalanx taihuensis and its phylogenetic analysis. GenBank 2021. submitted for publication. [Google Scholar]
- Balakirev, E.S.; Kravchenko, A.Y.; Romanov, N.S.; Ayala, F.J. Complete mitochondrial genome of the Arctic rainbow smelt Osmerus dentex (Osmeriformes, Osmeridae). Mitochondrial DNA Part B 2018, 3, 881–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, N.; Miya, M.; Nishida, M. Complete mitochondrial DNA sequence of ayu Plecoglossus altivelis. Fish. Sci. 2001, 67, 474–481. [Google Scholar] [CrossRef]
- Fu, J.; Xue, T.; Li, J. Complete mitochondrial genome of large icefish, Protosalanx chinensis (Salmoniformes: Salangidae). GenBank 2015. submitted for publication. [Google Scholar]
- Ishiguro, N.B.; Miya, M.; Nishida, M. Basal euteleostean relationships: A mitogenomic perspective on the phylogenetic reality of the ‘Protacanthopterygii’. Mol. Phylogenet. Evol. 2003, 27, 476–488. [Google Scholar] [CrossRef]
- Schmidt, D.J.; Islam, R.; Hughes, J.M. Complete mitogenomes for two lineages of the Australian smelt, Retropinna semoni (Osmeriformes: Retropinnidae). Mitochondrial DNA B Resources 2016, 1, 615–616. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, N.B.; Miya, M.; Inoue, J.G.; Nishida, M. Sundasalanx (Sundasalangidae) is a progenetic clupeiform, not a closelyrelated group of salangids (Osmeriformes): Mitogenomic evidence. J. Fish Biol. 2005, 67, 561–569. [Google Scholar] [CrossRef]
- Zhang, H.; Xian, W. The complete mitochondrial genome of the larvae Salanx ariakensis (Osmeriformes, Salangidae) from Yangtze estuary. Mitochondrial DNA Part A 2016, 27, 1902–1903. [Google Scholar] [CrossRef]
- Ding, M.; Wang, Z.; Qi, J.; Zhang, B.; Zhang, J. Mitochondrial genome of the Salanx cuvieri (Osteichthyes: Salangidae). Mitochondrial DNA 2014, 27, 834–835. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood tree for the clearhead icefish Protosalanx chinensis and other members of the family Salangidae inferred from the complete mitochondrial (mt) genomes. The numerals at the nodes are bootstrap percent probability values based on 10,000 replications (values below 75% are omitted). Plecoglossus altivelis (Plecoglossidae) and Retropiunna retropinna (Retropinnidae) are used as outgroup. The species name P. hyalocranius was used for the originally published mt genome KJ499917 [30]. However, P. hyalocranius is a synonym of P. chinensis [19]. Consequently, we use the species name P. chinensis for the mt genome KJ499917 to avoid any confusion. The P. chinensis KP306787 genome is indicated as “Unverified” in GenBank. However, this mt genome shows very close affinity to all other P. chinensis genomes; the p-distance between this mt genome and the other P. chinensis mt genomes is 0.0111 ± 0.0007, which is even lower than the average p-distance for the rest of the P. chinensis mt genomes (0.0126 ± 0.0007). Consequently, we included the KP306787 mt genome in further analysis.
Figure 1.
Maximum likelihood tree for the clearhead icefish Protosalanx chinensis and other members of the family Salangidae inferred from the complete mitochondrial (mt) genomes. The numerals at the nodes are bootstrap percent probability values based on 10,000 replications (values below 75% are omitted). Plecoglossus altivelis (Plecoglossidae) and Retropiunna retropinna (Retropinnidae) are used as outgroup. The species name P. hyalocranius was used for the originally published mt genome KJ499917 [30]. However, P. hyalocranius is a synonym of P. chinensis [19]. Consequently, we use the species name P. chinensis for the mt genome KJ499917 to avoid any confusion. The P. chinensis KP306787 genome is indicated as “Unverified” in GenBank. However, this mt genome shows very close affinity to all other P. chinensis genomes; the p-distance between this mt genome and the other P. chinensis mt genomes is 0.0111 ± 0.0007, which is even lower than the average p-distance for the rest of the P. chinensis mt genomes (0.0126 ± 0.0007). Consequently, we included the KP306787 mt genome in further analysis.

Figure 2.
Sliding window plots of nucleotide variability along the complete mitochondrial genomes of Protosalanx chinensis. (a) Intraspecific polymorphism inferred from the five genomes studied; (b–h) Pairwise comparisons between the genomes are as follows: MW291629–MH330683 and KP306787–MH330683 (b); KJ499917–MH330683 (c); KJ499917–HM106494 (d); MW291629–KP306787 (e); MW291629–HM106494 (f); MH330683–HM106494 (g); MW291629–KJ499917 and KP306787–KJ499917 (h). Window sizes are 100 nucleotides with 25-nucleotide increments. Note the four significant peaks of divergence centered at the COI, ND4, and ND5 genes, and also at CR.
Figure 2.
Sliding window plots of nucleotide variability along the complete mitochondrial genomes of Protosalanx chinensis. (a) Intraspecific polymorphism inferred from the five genomes studied; (b–h) Pairwise comparisons between the genomes are as follows: MW291629–MH330683 and KP306787–MH330683 (b); KJ499917–MH330683 (c); KJ499917–HM106494 (d); MW291629–KP306787 (e); MW291629–HM106494 (f); MH330683–HM106494 (g); MW291629–KJ499917 and KP306787–KJ499917 (h). Window sizes are 100 nucleotides with 25-nucleotide increments. Note the four significant peaks of divergence centered at the COI, ND4, and ND5 genes, and also at CR.

Figure 3.
Maximum likelihood trees of the clearhead icefish Protosalanx chinensis and other members of the family Salangidae, as inferred from the analysis of the (a) COI; (b) ND4; (c) ND5; and (d) CR high divergence (HD) regions; and (e) the complete mitochondrial genomes with the HD regions excluded. The discordant phylogenetic signals between P. chinensis and the Neosalanx species are highlighted in bold. For other notes, see Figure 1.
Figure 3.
Maximum likelihood trees of the clearhead icefish Protosalanx chinensis and other members of the family Salangidae, as inferred from the analysis of the (a) COI; (b) ND4; (c) ND5; and (d) CR high divergence (HD) regions; and (e) the complete mitochondrial genomes with the HD regions excluded. The discordant phylogenetic signals between P. chinensis and the Neosalanx species are highlighted in bold. For other notes, see Figure 1.

Figure 4.
Schematic representation of the recombination events in the mitochondrial genomes of Protosalanx chinensis (a) KJ499917; (b) MH330683; and (c) HM106494. The major parental sequence (in white) is from P. chinensis (MW291629); the minor parental sequences (in black) are from N. tangkahkeii KP170510 (the COI, ND4, and CR recombination events) and N. anderssoni HM106492 (the ND5 recombination event). The recombinant fragments are indicated by black boxes. For other notes, see Table 2.
Figure 4.
Schematic representation of the recombination events in the mitochondrial genomes of Protosalanx chinensis (a) KJ499917; (b) MH330683; and (c) HM106494. The major parental sequence (in white) is from P. chinensis (MW291629); the minor parental sequences (in black) are from N. tangkahkeii KP170510 (the COI, ND4, and CR recombination events) and N. anderssoni HM106492 (the ND5 recombination event). The recombinant fragments are indicated by black boxes. For other notes, see Table 2.

Figure 5.
Alignment of variable nucleotide sites in the (a) COI; (b,c) ND4; (d) ND5; and (e) CR recombinant fragments and flanking regions for the salangid fishes Protosalanx chinensis (KJ499917, MW291629, and HM106494), Neosalanx tangkahkeii (KP170510), and N. anderssoni (HM106492). Dots indicate nucleotide identity with the recombinant sequence. Rec: recombinant sequence, Minor: minor parent sequence, Major: major parent sequence (see also Table 2 and Figure 4). The two mismatches in the ND4 alignment and the four mismatches in the ND5 alignment are highlighted in bold. The ND4 recombination event is represented in two parts, illustrating the (b) beginning and (c) ending breakpoint regions, respectively. The alignments of variable sites in the ND4 recombinant fragment are identical for the P. chinensis KJ499917 and MH330683 recombinant sequences. The polymorphic sites 10,391–11,558 of the ND4 region alignment, containing no differences between the recombinant and the minor parent sequences, were deleted to fit onto the page.
Figure 5.
Alignment of variable nucleotide sites in the (a) COI; (b,c) ND4; (d) ND5; and (e) CR recombinant fragments and flanking regions for the salangid fishes Protosalanx chinensis (KJ499917, MW291629, and HM106494), Neosalanx tangkahkeii (KP170510), and N. anderssoni (HM106492). Dots indicate nucleotide identity with the recombinant sequence. Rec: recombinant sequence, Minor: minor parent sequence, Major: major parent sequence (see also Table 2 and Figure 4). The two mismatches in the ND4 alignment and the four mismatches in the ND5 alignment are highlighted in bold. The ND4 recombination event is represented in two parts, illustrating the (b) beginning and (c) ending breakpoint regions, respectively. The alignments of variable sites in the ND4 recombinant fragment are identical for the P. chinensis KJ499917 and MH330683 recombinant sequences. The polymorphic sites 10,391–11,558 of the ND4 region alignment, containing no differences between the recombinant and the minor parent sequences, were deleted to fit onto the page.



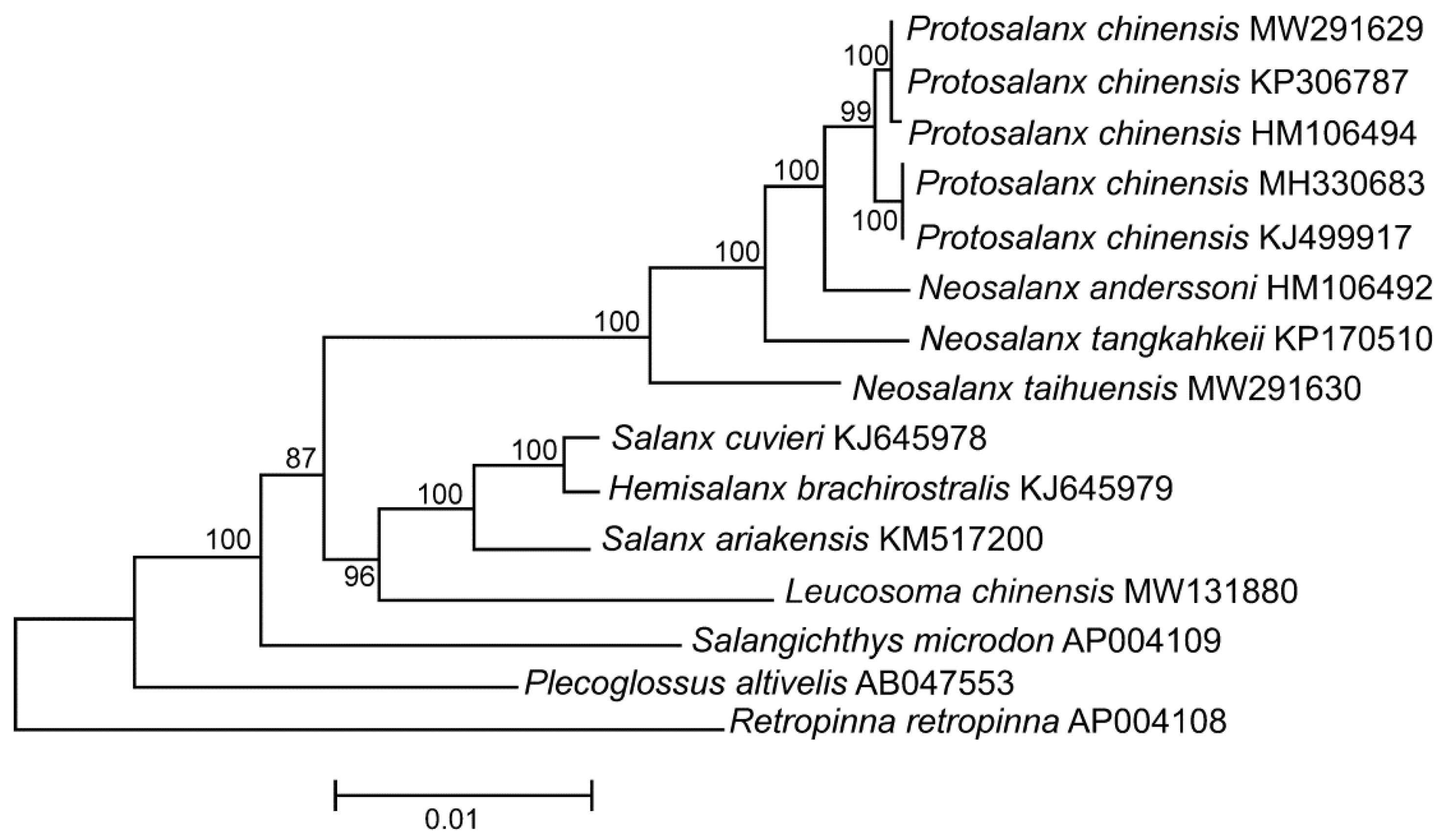{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Pairwise distances (Dxy ± SE) between Protosalanx chinensis and closely related species inferred from different segments of mitochondrial genomes.
Table 1.
Pairwise distances (Dxy ± SE) between Protosalanx chinensis and closely related species inferred from different segments of mitochondrial genomes.
| Species Pair | Full Genomes | COI | ND4 | ND5 | CR | HD Regions Excluded |
|---|---|---|---|---|---|---|
| P. chinensis MW291629—P. chinensis KJ499917 | 0.0162 ± 0.0009 | 0.0638 ± 0.0196 | 0.1164 ± 0.0077 | 0.0000 ± 0.0000 | 0.1293 ± 0.0169 | 0.0019 ± 0.0003 |
| P. chinensis MW291629—P. chinensis MH330683 | 0.0143 ± 0.0009 | 0.0142 ± 0.0096 | 0.1164 ± 0.0077 | 0.0000 ± 0.0000 | 0.0201 ± 0.0071 | 0.0023 ± 0.0004 |
| P. chinensis MW291629—P. chinensis HM106494 | 0.0047 ± 0.0005 | 0.0071 ± 0.0070 | 0.0006 ± 0.0006 | 0.0682 ± 0.0116 | 0.0057 ± 0.0040 | 0.0034 ± 0.0005 |
| P. chinensis KJ499917—P. chinensis MH330683 | 0.0031 ± 0.0003 | 0.0496 ± 0.0180 | 0.0000 ± 0.0000 | 0.0000 ± 0.0000 | 0.1264 ± 0.0169 | 0.0011 ± 0.0002 |
| P. chinensis KJ499917—P. chinensis HM106494 | 0.0193 ± 0.0009 | 0.0567 ± 0.0192 | 0.1158 ± 0.0078 | 0.0682 ± 0.0116 | 0.1293 ± 0.0170 | 0.0037 ± 0.0005 |
| P. chinensis MH330683—P. chinensis HM106494 | 0.0170 ± 0.0009 | 0.0071 ± 0.0069 | 0.1158 ± 0.0078 | 0.0682 ± 0.0116 | 0.0144 ± 0.0059 | 0.0035 ± 0.0005 |
| P. chinensis MW291629—N. anderssoni HM106492 | 0.0472 ± 0.0015 | 0.0496 ± 0.0186 | 0.0649 ± 0.0055 | 0.0588 ± 0.0108 | 0.0776 ± 0.0144 | 0.0439 ± 0.0018 |
| P. chinensis KJ499917—N. anderssoni MH330683 | 0.0546 ± 0.0017 | 0.0638 ± 0.0196 | 0.1228 ± 0.0077 | 0.0588 ± 0.0108 | 0.1408 ± 0.0174 | 0.0446 ± 0.0018 |
| P. chineesis MH330683—N. anderssoni HM106492 | 0.0540 ± 0.0016 | 0.0496 ± 0.0182 | 0.1228 ± 0.0077 | 0.0588 ± 0.0108 | 0.0833 ± 0.0145 | 0.0448 ± 0.0018 |
| P. chinensis HM106494—N. anderssoni HM106492 | 0.0459 ± 0.0015 | 0.0426 ± 0.0173 | 0.0643 ± 0.0054 | 0.0094 ± 0.0044 | 0.0718 ± 0.0140 | 0.0442 ± 0.0017 |
| P. chinensis MW291629—N. tangkahkeii KP170510 | 0.0893 ± 0.0018 | 0.0638 ± 0.0196 | 0.1152 ± 0.0077 | 0.1012 ± 0.0136 | 0.1293 ± 0.0169 | 0.0859 ± 0.0024 |
| P. chinensis KJ499917—N. tangkahkeii KP170510 | 0.0753 ± 0.0018 | 0.0000 ± 0.0000 | 0.0012 ± 0.0007 | 0.1012 ± 0.0136 | 0.0000 ± 0.0000 | 0.0863 ± 0.0023 |
| P. chinensis MH330683—N. tangkahkeii KP170510 | 0.0776 ± 0.0017 | 0.0496 ± 0.0180 | 0.0012 ± 0.0007 | 0.1012 ± 0.0136 | 0.1264 ± 0.0169 | 0.0864 ± 0.0025 |
| P. chinensis HM106494—N. tangkahkeii KP170510 | 0.0899 ± 0.0019 | 0.0567 ± 0.0192 | 0.1146 ± 0.0078 | 0.1035 ± 0.0143 | 0.1293 ± 0.0170 | 0.0866 ± 0.0023 |
HD regions excluded: full mt genomes with deleted high divergence (HD) fragments located at the COI, ND4, ND5, and CR regions. The erroneously high or low pairwise p-distances due to the presence or absence of the N. tangkahkeii and N. anderssoni fragments in the P. chinensis genomes are highlighted in bold (see the text for details. The P. chinensis KP306787 mt genome with the redundant data concerning the pairwise distances are not included in the table.
Table 2.
A summary of the recombination events detected in the Protosalanx chinensis mitochondrial genome.
Table 2.
A summary of the recombination events detected in the Protosalanx chinensis mitochondrial genome.
| Recombination Event | Beginning Breakpoint | Ending Breakpoint | p-Value |
|---|---|---|---|
| COI (140 bp) Recombinant: Protosalanx chinensis (KJ499917) Major parent: Protosalanx chinensis (MW291629) Minor parent: Neosalanx tangkahkeii (KP170510) | 5566 | 5705 | 1.730 × 10−13 (G) 1.380 × 10−14 (B) 1.977 × 10−02 (M) 1.954 × 10−02 (C) 1.611 × 10−05 (Q) |
| ND4 (1710 bp) Recombinant 1: Protosalanx chinensis (KJ499917) Recombinant 2: Protosalanx chinensis (MH330683) Major parent: Protosalanx chinensis (MW291629) Minor parent: Neosalanx tangkahkeii (KP170510) | 10,167 | 11,876 | 3.865 × 10−146 (R) 6.784 × 10−143 (G) 2.238 × 10−110 (B) 3.884 × 10−36 (M) 2.448 × 10−36 (C) 4.440 × 10−16 (Q) |
| ND5 (425 bp) Recombinant: Protosalanx chinensis (HM106494) Major parent: Protosalanx chinensis (MW291629) Minor parent: Neosalanx anderssoni (HM106492) | 12,946 | 13,372 | 1.491 × 10−26 (R) 8.703 × 10−25 (G) 1.310 × 10−26 (B) 4.644 × 10−10 (M) 1.099 × 10−09 (C) 2.220 × 10−16 (Q) |
| CR (359 bp excluding alignment gaps) Recombinant: Protosalanx chinensis (KJ499917) Major parent: Protosalanx chinensis (MW291629) Minor parent: Neosalanx tangkahkeii (KP170510) | 15,606 | 16,193 | 3.719 × 10−49 (R) 8.960 × 10−55 (G) 3.878 × 10−49 (B) 3.155 × 10−13 (M) 8.322 × 10−13 (C) 3.330 × 10−16 (Q) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balakirev, E.S. Recombinant Mitochondrial Genomes Reveal Recent Interspecific Hybridization between Invasive Salangid Fishes. Life 2022, 12, 661. https://doi.org/10.3390/life12050661
AMA Style
Balakirev ES. Recombinant Mitochondrial Genomes Reveal Recent Interspecific Hybridization between Invasive Salangid Fishes. Life. 2022; 12(5):661. https://doi.org/10.3390/life12050661
Chicago/Turabian StyleBalakirev, Evgeniy S. 2022. "Recombinant Mitochondrial Genomes Reveal Recent Interspecific Hybridization between Invasive Salangid Fishes" Life 12, no. 5: 661. https://doi.org/10.3390/life12050661
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.