Intracellular Delivery of Biologically-Active Fungal Metabolite Gliotoxin Using Magnetic Nanoparticles

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
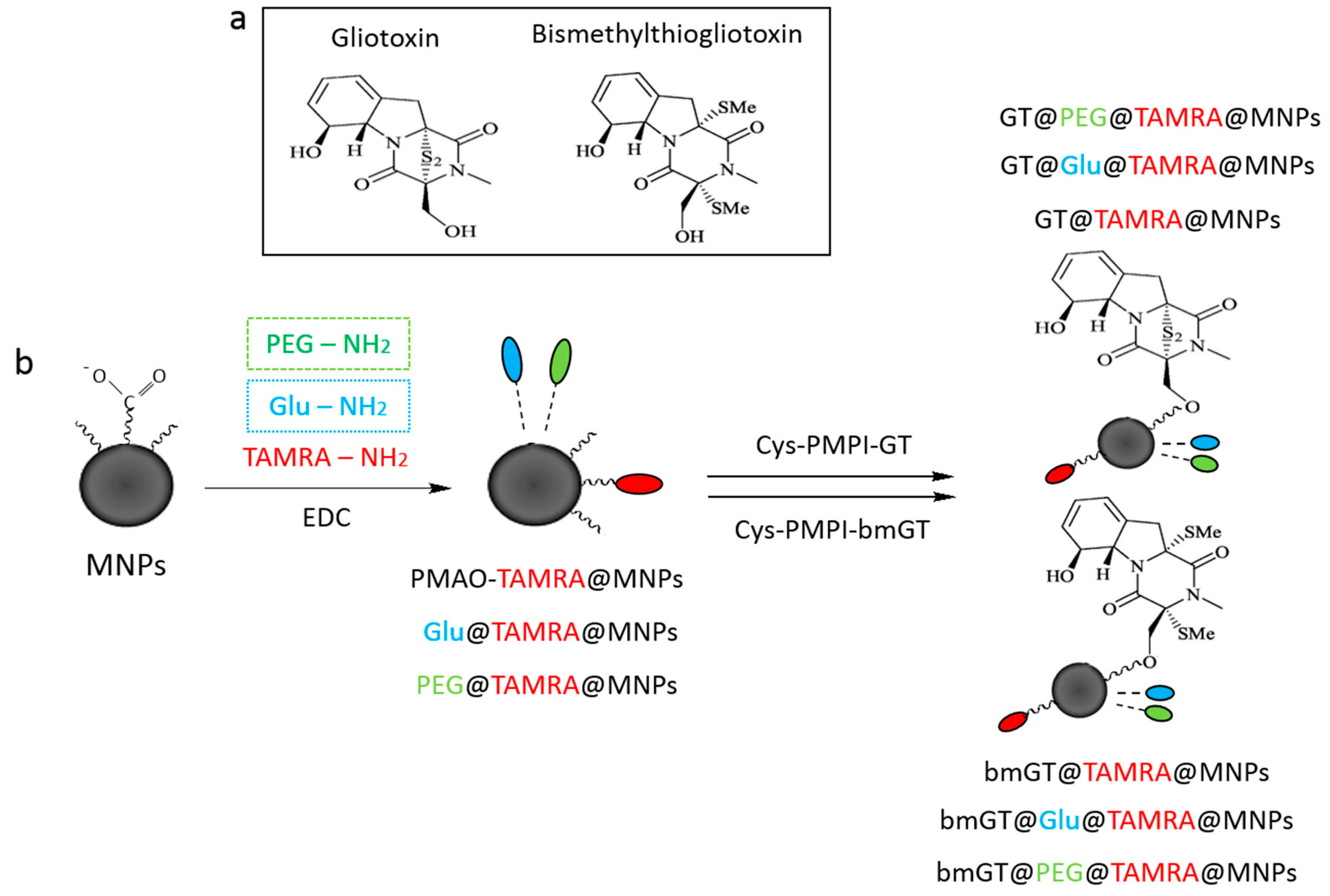
2.1. Synthesis of MNPs
2.2. Functionalization of MNPs with Gliotoxin
2.3. Characterization of MNPs
2.4. Cell Culture and Reagents
2.5. Cell Viability, Cell Proliferation, and Cytotoxicity Assay
2.6. Flow Cytometry
2.7. Confocal Microscopy
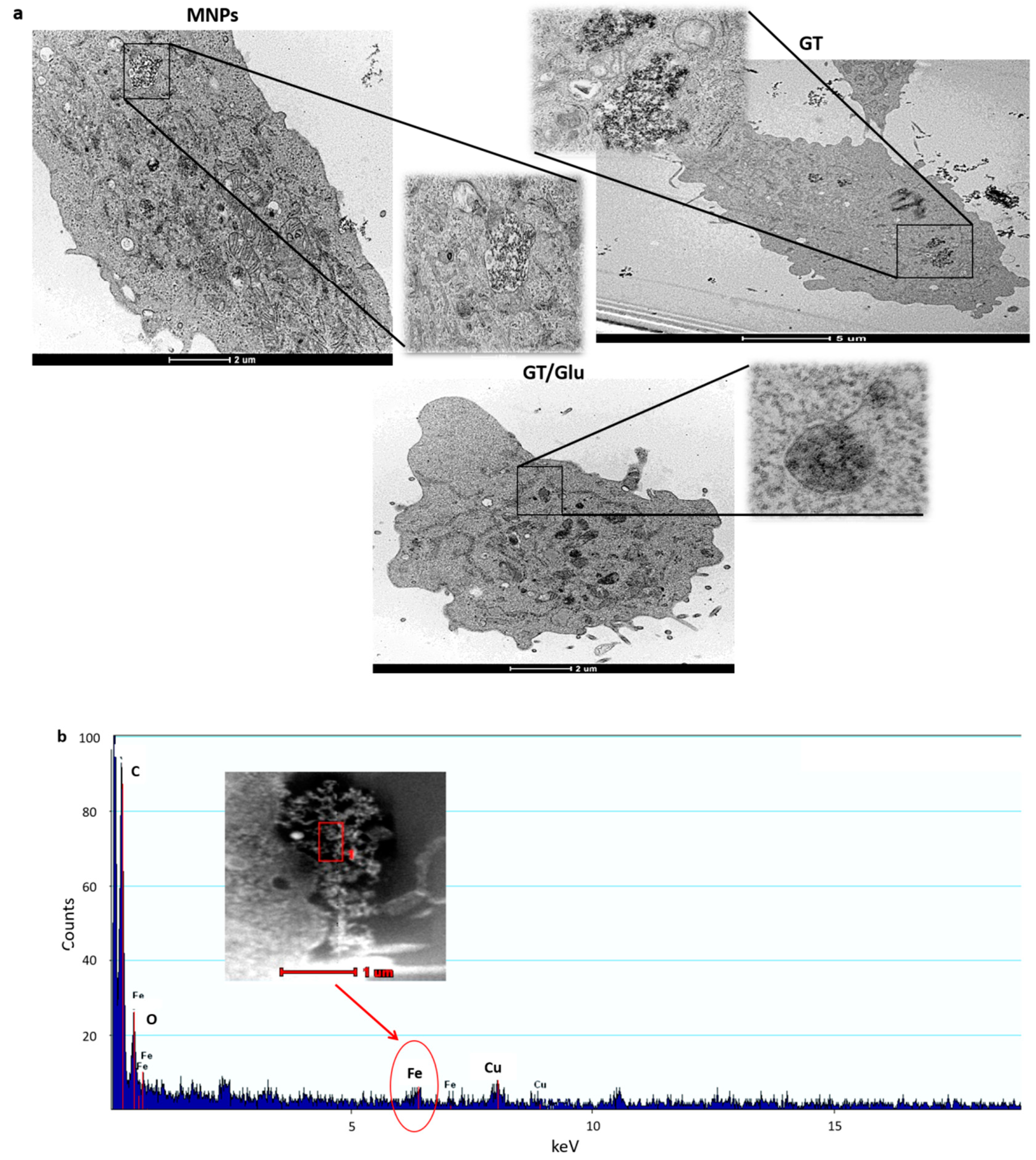
2.8. Transmission Electron Microscopy
2.9. Statistics
3. Results and Discussion
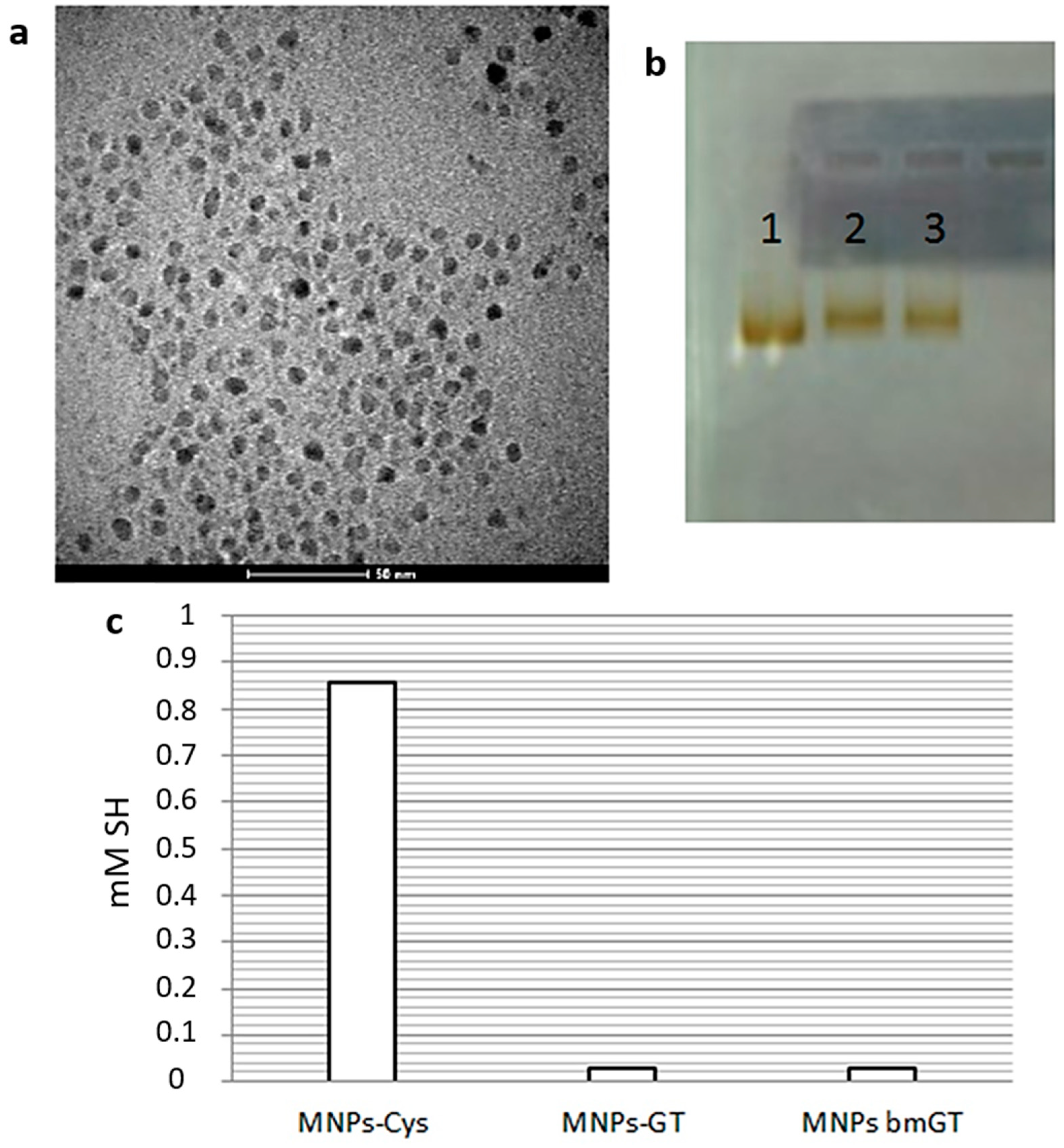
3.1. Characterization of Magnetic NPs Functionalized with GT and Inactive bmGT
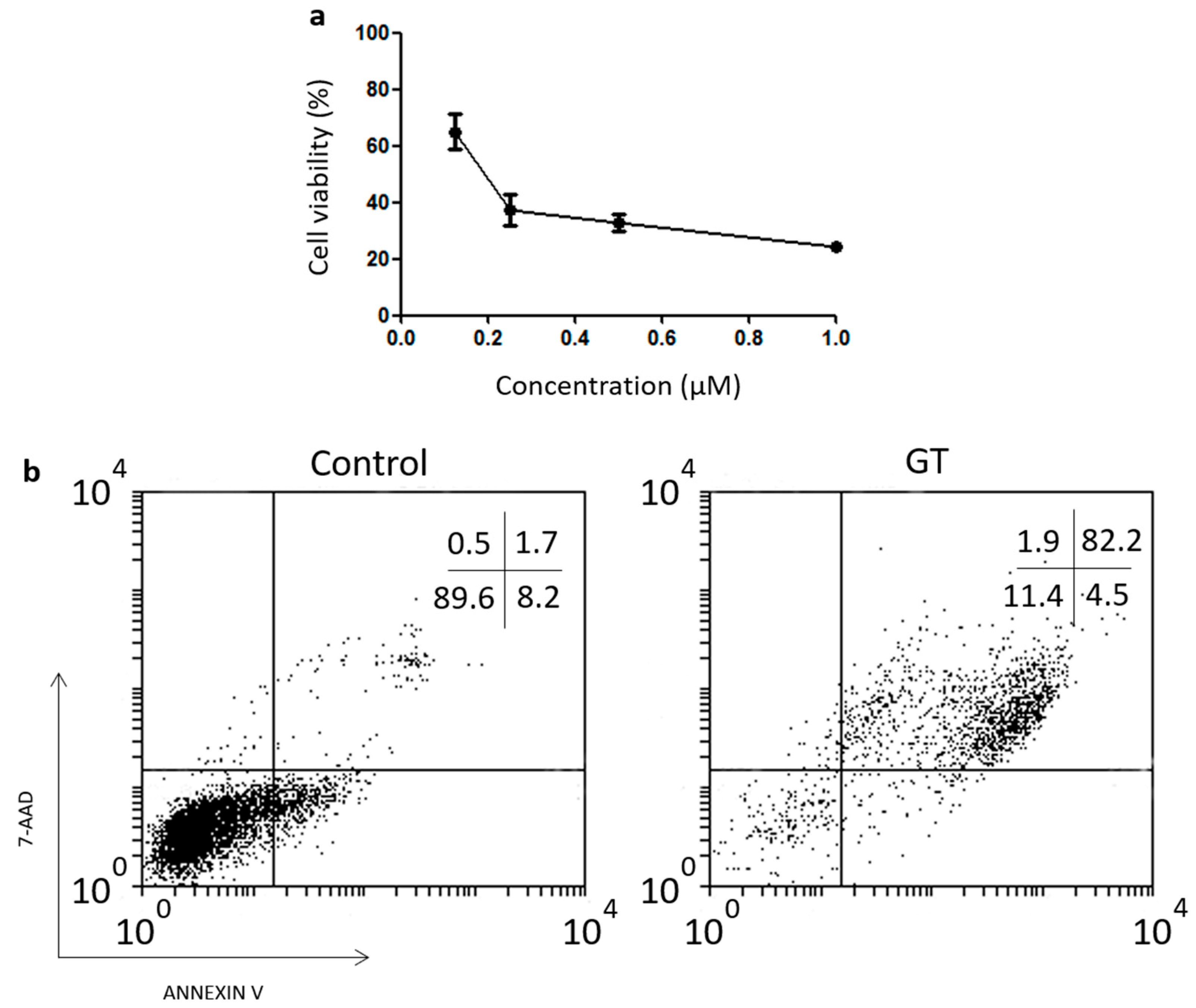
3.2. Magnetic NPs Functionalized with GT Kill L929 Sarcoma Cells
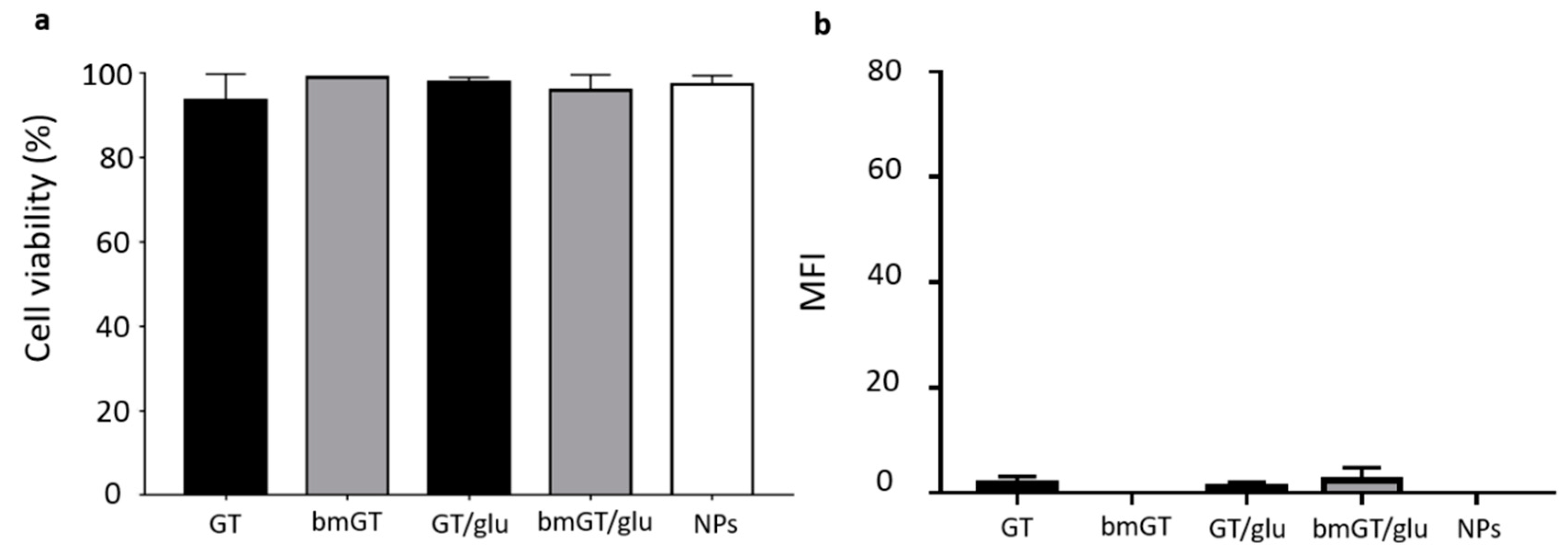
3.3. Glucose Enhances the Anti-Tumor Activity of Magnetic NPs Functionalized with GT against L929 Sarcoma Cells
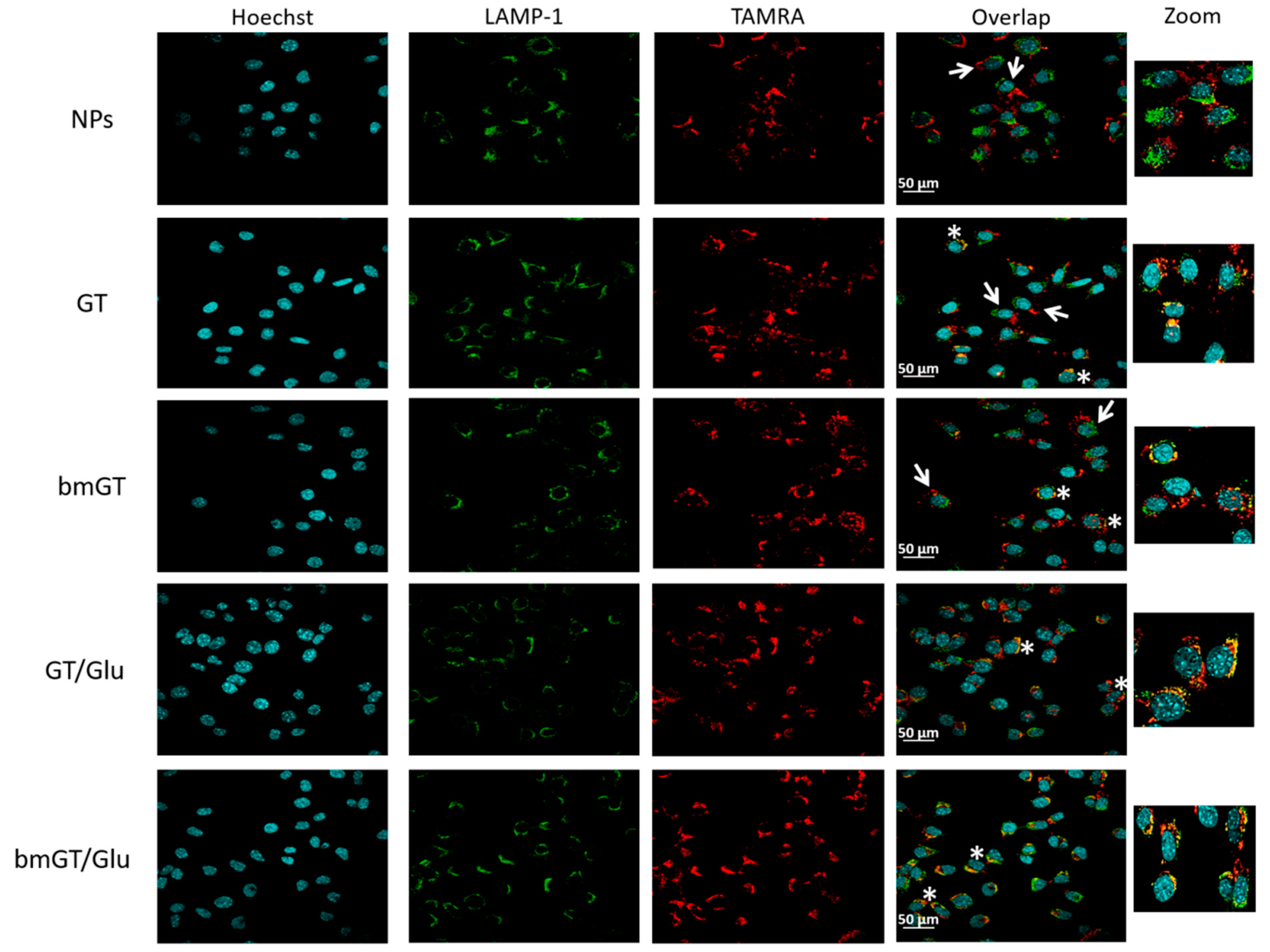
3.4. Glucose Enhances Cellular Interaction of Magnetic NPs Functionalized with GT in L929 Sarcoma Cells
3.5. Glucose Enhances Intracellular Delivery of Magnetic NPs Functionalized with GT in L929 Sarcoma Cells
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Taylor, A. The toxicology of sporidesmins and other epipolythiodioxopiperazines. Microb. Toxins 1971, 337–376. [Google Scholar]
- Müllbacher, A.; Waring, P.; Tiwari-Palni, U.; Eichner, R.D. Structural relationship of epipolythiodioxopiperazines and their immunomodulating activity. Mol. Immunol. 1986, 23, 231–235. [Google Scholar] [CrossRef]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology 1932, 22, 837–845. [Google Scholar]
- Fukuyama, T.; Kishi, Y. A Total Synthesis of Gliotoxin. J. Am. Chem. Soc. 1976, 98, 6723–6724. [Google Scholar] [CrossRef]
- Fukuyama, T.; Nakatsuka, S.I.; Kishi, Y. Total synthesis of gliotoxin, dehydrogliotoxin and hyalodendrin. Tetrahedron 1981, 37, 2045–2078. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Lu, M.; Totokotsopoulos, S.; Heretsch, P.; Giguère, D.; Sun, Y.P.; Sarlah, D.; Nguyen, T.H.; Wolf, I.C.; Smee, D.F. Synthesis and biological evaluation of epidithio-, epitetrathio-, and bis-(methylthio)diketopiperazines: Synthetic methodology, enantioselective total synthesis of epicoccin G, 8,8′-epi-ent-rostratin B, gliotoxin, gliotoxin G, emethallicin E, and haematoc. J. Am. Chem. Soc. 2012, 134, 17320–17332. [Google Scholar] [CrossRef]
- Korzybski, T.; Kowszyk-Gindifer, Z.; Kuryłowicz, W.; Paryski, E. Antibiotics. In Volume II: Origin, Nature and Properties; Paryski, E., Translator; Pergamon Press: Oxford, UK, 1967; pp. 450–715. [Google Scholar]
- Van Der Pyl, D.; Yang, H.; Inokoshi, J.; Shiomi, K.; Takeshima, H.; Omura, S. Inhibition of Farnesyl-protein transferase by gliotoxin and acetylgliotixin. J. Antibiot 1992, 45, 1802–1805. [Google Scholar]
- Müllbacher, A.; Eichner, R.D. Immunosuppression in vitro by a metabolite of a human pathogenic fungus. Proc. Natl. Acad. Sci. USA 1984, 81, 3835–3837. [Google Scholar] [CrossRef]
- Eichner, R.D.; Mullbacher, A. Hypothesis: Fungal toxins are involved in aspergillosis and AIDS. Aust. J. Exp. Biol. Med. Sci. 1984, 62, 479–484. [Google Scholar] [CrossRef]
- Sutton, P.; Waring, P.; Mullbacher, A. Exacerbation of invasive aspergillosis by the immunosuppressive fungal metabolite, gliotoxin. Immunol. Cell Biol. 1996, 74, 318–322. [Google Scholar] [CrossRef]
- Waring, P.; Eichner, R.D.; Mullbacher, A.; Sjaarda, A. Gliotoxin induces apoptosis in macrophages unrelated to its antiphagocytic properties. J. Biol. Chem. 1988, 263, 18493–18499. [Google Scholar] [PubMed]
- Tsunawaki, S.; Yoshida, L.S.; Nishida, S.; Kobayashi, T.; Shimoyama, T. Fungal metabolite gliotoxin inhibits assembly of the human respiratory burst NADPH oxidase. Infect. Immun. 2004, 72, 3373–3382. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; Owens, R.A.; O’Keeffe, G.; Hammel, S.; Fitzpatrick, D.A.; Jones, G.W.; Doyle, S. Regulation of nonribosomal peptide synthesis: Bis-thiomethylation attenuates gliotoxin biosynthesis in Aspergillus fumigatus. Chem. Biol. 2014, 21, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Y.; Zhu, T.J.; Han, X.X.; Fan, G.T.; Liu, H.B.; Zhu, W.M.; Gu, Q.Q. A new gliotoxin analogue from a marine-derived fungus Aspergillus fumigatus Fres. Nat. Prod. Res. 2009, 23, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Maschmeyer, G.; Haas, A.; Cornely, O.A. Invasive aspergillosis: Epidemiology, diagnosis and management in immunocompromised patients. Drugs 2007, 67, 1567–1601. [Google Scholar] [CrossRef] [PubMed]
- Domingo, M.P.; Colmenarejo, C.; Martínez-Lostao, L.; Müllbacher, A.; Jarne, C.; Revillo, M.J. Bis(methyl)gliotoxin proves to be a more stable and reliable marker for invasive aspergillosis than gliotoxin and suitable for use in diagnosis. Diagn. Microbiol. Infect. Dis. 2012, 73, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Vidal-García, M.; Domingo, M.P.; De Rueda, B.; Roc, L.; Delgado, M.P.; Revillo, M.J.; Pardo, J.; Gálvez, E.M.; Rezusta, A. Clinical validity of bis(methylthio)gliotoxin for the diagnosis of invasive aspergillosis. Appl. Microbiol. Biotechnol. 2016, 100, 2327–2334. [Google Scholar] [CrossRef]
- Gladue, R.P.; Carroll, L.A.; Milici, A.J.; Scampoli, D.N.; Stukenbrok, H.A.; Pettipher, E.R. Inhibition of leukotriene B4-receptor interaction suppresses eosinophil infiltration and disease pathology in a murine model of experimental allergic encephalomyelitis. J. Exp. Med. 1996, 183, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Sugui, J.A. What do we know about the role of gliotoxin in the pathobiology of Aspergillus fumigatus? Med. Mycol. 2009, 47 (Suppl. 1), S97–S103. [Google Scholar] [CrossRef]
- McMinn, P.C.; Halliday, G.M.; Waring, P.; Konrad Muller, H. Langerhans cell depletion in gliotoxin-treated murine epidermis. Pathology 1991, 23, 39–44. [Google Scholar] [CrossRef]
- Stanzani, M.; Orciuolo, E.; Lewis, R.; Kontoyiannis, D.P.; Martins, S.L.R.; St. John, L.S.; Komanduri, K.V. Aspergillus fumigatus suppresses the human cellular immune response via gliotoxin-mediated apoptosis of monocytes. Blood 2005, 105, 2258–2265. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; O’Keeffe, G.; Jones, G.W.; Doyle, S. Resistance is not futile: Gliotoxin biosynthesis, functionality and utility. Trends Microbiol. 2015, 23, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Sutton, P.; Newcombe, N.R.; Waring, P.; Mullbacher, A. In vivo immunosuppressive activity of gliotoxin, a metabolite produced by human pathogenic fungi. Infect. Immun. 1994, 62, 1192–1198. [Google Scholar] [PubMed]
- Eichner, R.D.; Waring, P.; Geue, A.M.; Braithwaite, A.W.; Mullbacher, A. Gliotoxin causes oxidative damage to plasmid and cellular DNA. J. Biol. Chem. 1988, 263, 3772–3777. [Google Scholar] [PubMed]
- Kweon, Y.-O.; Paik, Y.-H.; Schnabl, B.; Qian, T.; Lemasters, J.J.; Brenner, D.A. Gliotoxin-mediated apoptosis of activated human hepatic stellate cells. J. Hepatol. 2003, 39, 38–46. [Google Scholar] [CrossRef]
- Pardo, J.; Urban, C.; Galvez, E.M.; Ekert, P.G.; Müller, U.; Kwon-Chung, J. The mitochondrial protein Bak is pivotal for gliotoxin-induced apoptosis and a critical host factor of Aspergillus fumigatus virulence in mice. J. Cell. Biol. 2006, 174, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Suen, Y.K.; Fung, K.P.; Lee, C.Y.; Kong, S.K. Gliotoxin induces apoptosis in cultured macrophages via production of reactive oxygen species and cytochrome c release without mitochondrial depolarization. Free Radic Res. 2001, 35, 1–10. [Google Scholar] [CrossRef]
- Zhou, X.; Zhao, A.; Goping, G.; Hirszel, P. Gliotoxin-induced cytotoxicity proceeds via apoptosis and is mediated by caspases and reactive oxygen species in LLC-PK1 cells. Toxicol. Sci. 2000, 54, 194–202. [Google Scholar] [CrossRef]
- Pahl, H.L.; Krauss, B.; Schulze-Osthoff, K.; Decker, T.; Traenckner, E.B.; Vogt, M. The immunosuppressive fungal metabolite gliotoxin specifically inhibits transcription factor NF-kappaB. J. Exp. Med. 1996, 183, 1829–1840. [Google Scholar] [CrossRef]
- Kroll, M.; Arenzana-Seisdedos, F.; Bachelerie, F.; Thomas, D.; Friguet, B.; Conconi, M. The secondary fungal metabolite gliotoxin targets proteolytic activities of the proteasome. Chem. Biol. 1999, 6, 689–698. [Google Scholar] [CrossRef]
- Hubmann, R.; Sieghart, W.; Schnabl, S.; Araghi, M.; Hilgarth, M.; Reiter, M.; Demirtas, D.; Valent, P.; Zielinski, C.; Jäger, U. Gliotoxin targets nuclear NOTCH2 in human solid tumor derived cell lines in vitro and inhibits melanoma growth in xenograft mouse model. Front. Pharmacol. 2017, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, C.; Lan, W.; Huang, C.; Lin, M.; Wang, Z.; Liang, W.; Iwamoto, A.; Yang, X.; Liu, H. Gliotoxin inhibits proliferation and induces apoptosis in colorectal cancer cells. Mar. Drugs. 2015, 13, 6259–6273. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, L.R.; Wang, J.; Le, T. In Vitro and In Vivo effects oggliotoxinEfficacy Against Dextran Sodium Sulfate-Induced Colitis in rat. Dig. Dis. Sci. 2000, 45, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Herfarth, H.; Brand, K.; Rath, H.C.; Rogler, G.; Scholmerich, J.; Falk, W. Nuclear factor-kB activity and intestinal inflammation in dextran sulphate sodium (DSS)-induced colitis in mice is suppressed by gliotoxin. Clin. Exp. Immunol. 2000, 120, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, X.; Yu, B.; Ren, K. Gliotoxin potentiates osteoblast differentiation by inhibiting nuclear factor-κB signaling. Mol. Med. Rep. 2015, 12, 877–884. [Google Scholar] [CrossRef]
- Müllbacher, A.; Hume, D.; Braithwaite, A.W.; Waring, P.; Eichner, R.D. Selective resistance of bone marrow-derived hemopoietic progenitor cells to gliotoxin. Proc. Natl. Acad. Sci. USA 1987, 84, 3822–3825. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.H.; Arias, J.L.; Nicolas, J.; Couvreur, P. Magnetic nanoparticles: Design and characterization, toxicity and biocompatibility, pharmaceutical and biomedical applications. Chem. Rev. 2012, 112, 5818–5878. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.L.; Zhang, Y.F.; Gao, F.; Li, G.L.; He, Y.; Peng, M.L.; Fan, H.M. Magnetic nanoparticles based cancer therapy: Current status and applications. Sci. China Life Sci. 2018, 61, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Thanou, M. Targeting nanoparticles to cancer. Pharmacol. Res. 2010, 62, 90–99. [Google Scholar] [CrossRef]
- Moros, M.; Pelaz, B.; López-Larrubia, P.; García-Martin, M.L.; Grazú, V.; de la Fuente, J.M. Engineering biofunctional magnetic nanoparticles for biotechnological applications. Nanoscale 2010, 2, 1746. [Google Scholar] [CrossRef]
- Sun, S.; Zeng, H.; Robinson, D.B.; Raoux, S.; Rice, P.M.; Wang, S.X.; Li, G. Monodisperse MFe2O4 (M = Fe, Co, Mn) Nanoparticles. J. Am. Chem. Soc. 2004, 126, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Moros, M.; Hernáez, B.; Garet, E.; Dias, J.T.; Sáez, B.; Grazú, V. Monosaccharides versus PEG-functionalized NPs: Influence in the cellular uptake. ACS Nano 2012, 6, 1565–1577. [Google Scholar] [CrossRef] [PubMed]
- Catalán, E.; Charni, S.; Jaime, P.; Aguiló, J.I.; Enríquez, J.A.; Naval, J.; Pardo, J.; Villalba, M.; Anel, A. MHC-I modulation due to changes in tumor cell metabolism regulates tumor sensitivity to CTL and NK cells. Oncoimmunology 2015, 4, e985924. [Google Scholar] [CrossRef] [PubMed]
- Pardo, J.; Perez-Galan, P.; Gamen, S.; Marzo, I.; Monleon, I.; Kaspar, A.A. A Role of the Mitochondrial Apoptosis-Inducing Factor in Granulysin-Induced Apoptosis. J. Immunol. 2001, 167, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hernández, M.; Del Pino, P.; Mitchell, S.G.; Moros, M.; Stepien, G.; Pelaz, B.; Parak, W.J.; Gálvez, E.M.; Pardo, J.; de la Fuente, J.M. Dissecting the molecular mechanism of apoptosis during photothermal therapy using gold nanoprisms. ACS Nano 2015, 9, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Beola, L.; Asín, L.; Fratila, R.M.; Herrero, V.; de la Fuente, J.M.; Grazú, V.; Gutiérrez, L. Dual Role of Magnetic Nanoparticles as Intracellular Hotspots and Extracellular Matrix Disruptors Triggered by Magnetic Hyperthermia in 3D Cell Culture Models. ACS Appl. Mater. Interfaces 2018, 10, 44301–44313. [Google Scholar] [CrossRef] [PubMed]
- Stepien, G.; Moros, M.; Pérez-Hernández, M.; Monge, M.; Gutiérrez, L.; Fratila, R.M.; Las Heras, M.; Menao Guillén, S.; Puente Lanzarote, J.J.; Solans, C. Effect of Surface Chemistry and Associated Protein Corona on the Long-Term Biodegradation of Iron Oxide Nanoparticles In Vivo. ACS Appl. Mater. Interfaces 2018, 10, 4548–4560. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.; Moros, M.; del Pino, P.; Fernández, S.; Grazú, V.; de la Fuente, J.M. DNA as a Molecular Local Thermal Probe for Magnetic Hyperthermia Analysis. Angew. Chem. 2013, 52, 11526–11529. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Ambrosone, A.; Sanz, V.; Hernandez, Y.; Marchesano, V.; Tian, F. Design of multifunctional gold nanoparticles for in vitro and in vivo gene silencing. ACS Nano 2012, 6, 8316–8324. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Niestroj, M.; Yuan, D.; Chang, S.; Chen, J. Treating cancer stem cells and cancer metastasis using glucose-coated gold nanoparticles. Int. J. Nanomed. 2015, 10, 2065–2077. [Google Scholar]
- Xie, J.; Xu, C.; Kohler, N.; Hou, Y.; Sun, S. Controlled PEGylation of monodisperse Fe3O4 nanoparticles for reduced non-specific uptake by macrophage cells. Adv. Mater. 2007, 19, 3163–3166. [Google Scholar] [CrossRef]
- Cho, E.C.; Zhang, Q.; Xia, Y. The effect of sedimentation and diffusion on cellular uptake of gold nanoparticles. Nat. Nanotechnol. 2011, 24, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, P.H.; Brasch, N.; Chai, C.L.L.; Waring, P. A Novel Redox Mechanism for the Glutathione-dependent Reversible Uptake of a Fungal Toxin in Cells. J. Biol. Chem. 2003, 278, 46549–46555. [Google Scholar] [CrossRef] [PubMed]
- Kong, T.; Zeng, J.; Wang, X.; Yang, X.; Yang, J.; McQuarrie, S.; McEwan, A.; Roa, W.; Chen, J.; Xing, J.Z. Enhancement of radiation cytotoxicity in breast-cancer cells by localized attachment of gold nanoparticles. Small 2008, 4, 1537–1543. [Google Scholar] [CrossRef]
- Pelaz, B.; Grazu, V.; Ibarra, A.; Magen, C.; Del Pino, P.; De La Fuente, J.M. Tailoring the synthesis and heating ability of gold nanoprisms for bioapplications. Langmuir 2012, 28, 8965–8970. [Google Scholar] [CrossRef] [PubMed]
- Evelina, M.; Gian Paolo, S.; Federica, T.; Angelo, Z.; De Marinis, F.; Giulia, P.; Luigi, R.Z.; Silverio, T. Positron Emission Tomography (PET) radiotracers in oncology—Utility of 18F-Fluoro-deoxy-glucose (FDG)-PET in the management of patients with non-small-cell lung cancer (NSCLC). J. Exp. Clin. Cancer Res. 2008, 27, 52. [Google Scholar]
- Arvizo, R.R.; Miranda, O.R.; Thompson, M.A.; Pabelick, C.M.; Bhattacharya, R.; Robertson, J.D.; Rotello, V.M.; Prakash, Y.S.; Mukherjee, P. Effect of nanoparticle surface charge at the plasma membrane and beyond. Nano Lett. 2010, 10, 2543–2548. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comas, L.; Polo, E.; Domingo, M.P.; Hernández, Y.; Arias, M.; Esteban, P.; Martínez-Lostao, L.; Pardo, J.; Martínez de la Fuente, J.; Gálvez, E.M. Intracellular Delivery of Biologically-Active Fungal Metabolite Gliotoxin Using Magnetic Nanoparticles. Materials 2019, 12, 1092. https://doi.org/10.3390/ma12071092
Comas L, Polo E, Domingo MP, Hernández Y, Arias M, Esteban P, Martínez-Lostao L, Pardo J, Martínez de la Fuente J, Gálvez EM. Intracellular Delivery of Biologically-Active Fungal Metabolite Gliotoxin Using Magnetic Nanoparticles. Materials. 2019; 12(7):1092. https://doi.org/10.3390/ma12071092
Chicago/Turabian StyleComas, Laura, Esther Polo, M Pilar Domingo, Yulán Hernández, Maykel Arias, Patricia Esteban, Luis Martínez-Lostao, Julián Pardo, Jesús Martínez de la Fuente, and Eva M Gálvez. 2019. "Intracellular Delivery of Biologically-Active Fungal Metabolite Gliotoxin Using Magnetic Nanoparticles" Materials 12, no. 7: 1092. https://doi.org/10.3390/ma12071092