
Analysis of S-Adenosylmethionine and S-Adenosylhomocysteine: Method Optimisation and Profiling in Healthy Adults upon Short-Term Dietary Intervention

 ,
,  , , ,
, , ,
Abstract
:
1. Introduction
2. Results
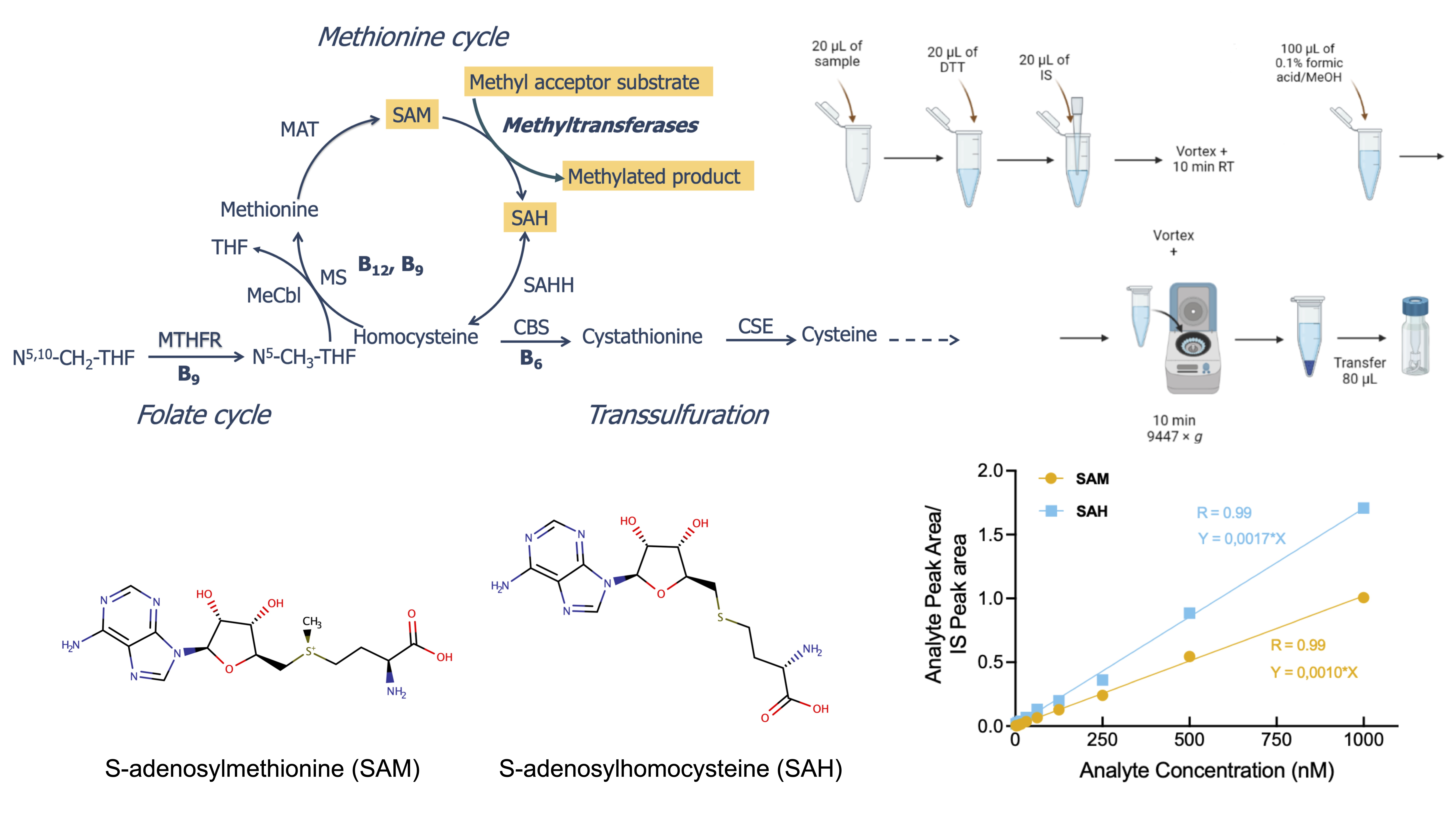
2.1. Separation and Quantification of SAM and SAH by LC-MS/MS
2.2. Linearity Range and Limit of Quantification
2.3. Recovery, Carry Over, and Analysis of Matrix Effect
2.4. Determination of SAM and SAH in the Plasma of Healthy Adults
2.5. Determination of SAM and SAH in the Plasma of Patients with Inborn Errors of Metabolism
2.6. Profile of SAM and SAH in a Randomised Control Trial in Healthy Subjects with Dietary Intervention with a Vegan (VD) or a Meat-Rich (MD) Diet
2.6.1. Concentration of SAM, SAH, and Creatinine in the Plasma and Urine of Healthy Omnivores at the Start of the Trial
2.6.2. Effect of Dietary Intervention on the Concentration of SAM and SAH in the Plasma and Urine of Healthy Subjects
2.6.3. Associations of Plasma and Urinary SAM and SAH before and after Dietary Intervention
3. Discussion
Strengths and Limitations of the Study
4. Materials and Methods
4.1. Specimen Collection, Pre-Analytics, and Storage
4.2. Cell Lysis and Protein Quantification
4.3. Preparation of Stock Solutions, Internal Standard (IS) Solution, and Extraction Solution
4.4. Preparation of Calibration Curves
4.5. Preparation of Samples and Normalisation
4.6. Preparation of Quality Controls (QCs)
4.7. LC-MS/MS Method, Signal Processing and Quantification, and Validation
4.8. Matrix Effects
4.9. Ethical Approval
4.10. Cohort Characteristics of the Randomised Controlled trial (RCT) with Intervention with Vegan (VD) or Meat-Rich (MD) Diets
4.11. Statistical Analysis
4.12. Data Visualisation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mato, J.M.; Corrales, F.J.; Lu, S.C.; Avila, M.A. S-Adenosylmethionine: A control switch that regulates liver function. FASEB J. 2002, 16, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauerle, M.R.; Schwalm, E.L.; Booker, S.J. Mechanistic diversity of radical S-adenosylmethionine (SAM)-dependent methylation. J. Biol. Chem. 2015, 290, 3995–4002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broderick, W.E.; Broderick, J.B. Radical SAM enzymes: Surprises along the path to understanding mechanism. J. Biol. Inorg. Chem. 2019, 24, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.P.; Abdelwahed, S.H.; Mahanta, N.; Fedoseyenko, D.; Philmus, B.; Cooper, L.E.; Liu, Y.; Jhulki, I.; Ealick, S.E.; Begley, T.P. Radical S-adenosylmethionine (SAM) enzymes in cofactor biosynthesis: A treasure trove of complex organic radical rearrangement reactions. J. Biol. Chem. 2015, 290, 3980–3986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.C.; Frey, P.A. S-adenosylmethionine as an oxidant: The radical SAM superfamily. Trends Biochem. Sci. 2007, 32, 101–110. [Google Scholar] [CrossRef]
- Hannibal, L.; Blom, H.J. Homocysteine and disease: Causal associations or epiphenomenons? Mol. Aspects Med. 2017, 53, 36–42. [Google Scholar] [CrossRef]
- Hannibal, L.; DiBello, P.M.; Jacobsen, D.W. Proteomics of vitamin B12 processing. Clin. Chem. Lab. Med. 2013, 51, 477–488. [Google Scholar] [CrossRef] [Green Version]
- McCorvie, T.J.; Kopec, J.; Hyung, S.J.; Fitzpatrick, F.; Feng, X.; Termine, D.; Strain-Damerell, C.; Vollmar, M.; Fleming, J.; Janz, J.M.; et al. Inter-domain communication of human cystathionine beta-synthase: Structural basis of S-adenosyl-L-methionine activation. J. Biol. Chem. 2014, 289, 36018–36030. [Google Scholar] [CrossRef] [Green Version]
- Froese, D.S.; Kopec, J.; Rembeza, E.; Bezerra, G.A.; Oberholzer, A.E.; Suormala, T.; Lutz, S.; Chalk, R.; Borkowska, O.; Baumgartner, M.R.; et al. Structural basis for the regulation of human 5,10-methylenetetrahydrofolate reductase by phosphorylation and S-adenosylmethionine inhibition. Nat. Commun. 2018, 9, 2261. [Google Scholar] [CrossRef] [Green Version]
- Chiang, P.K.; Gordon, R.K.; Tal, J.; Zeng, G.C.; Doctor, B.P.; Pardhasaradhi, K.; McCann, P.P. S-Adenosylmethionine and methylation. FASEB J. 1996, 10, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Fontecave, M.; Atta, M.; Mulliez, E. S-adenosylmethionine: Nothing goes to waste. Trends Biochem. Sci. 2004, 29, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Baric, I.; Fumic, K.; Glenn, B.; Cuk, M.; Schulze, A.; Finkelstein, J.D.; James, S.J.; Mejaski-Bosnjak, V.; Pazanin, L.; Pogribny, I.P.; et al. S-adenosylhomocysteine hydrolase deficiency in a human: A genetic disorder of methionine metabolism. Proc. Natl. Acad. Sci. USA 2004, 101, 4234–4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baric, I.; Staufner, C.; Augoustides-Savvopoulou, P.; Chien, Y.H.; Dobbelaere, D.; Grunert, S.C.; Opladen, T.; Petkovic Ramadza, D.; Rakic, B.; Wedell, A.; et al. Consensus recommendations for the diagnosis, treatment and follow-up of inherited methylation disorders. J. Inherit. Metab. Dis. 2017, 40, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahl, W.A.; Bernardini, I.; Finkelstein, J.D.; Tangerman, A.; Martin, J.J.; Blom, H.J.; Mullen, K.D.; Mudd, S.H. Transsulfuration in an adult with hepatic methionine adenosyltransferase deficiency. J. Clin. Invest. 1988, 81, 390–397. [Google Scholar] [CrossRef]
- Mudd, S.H. Hypermethioninemias of genetic and non-genetic origin: A review. Am. J. Med Genet. Part C Semin. Med Genet. 2011, 157, 3–32. [Google Scholar] [CrossRef]
- Chang, C.A.; Wei, X.C.; Martin, S.R.; Sinasac, D.S.; Al-Hertani, W. Transiently elevated plasma methionine, S-adenosylmethionine and S-adenosylhomocysteine: Unreported laboratory findings in a patient with NGLY1 deficiency, a congenital disorder of deglycosylation. JIMD Rep. 2019, 49, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Elshorbagy, A.K.; Jerneren, F.; Samocha-Bonet, D.; Refsum, H.; Heilbronn, L.K. Serum S-adenosylmethionine, but not methionine, increases in response to overfeeding in humans. Nutr. Diabetes 2016, 6, e192. [Google Scholar] [CrossRef] [Green Version]
- Lind, M.V.; Lauritzen, L.; Pedersen, O.; Vestergaard, H.; Stark, K.D.; Hansen, T.; Ross, A.B.; Kristensen, M. Higher intake of fish and fat is associated with lower plasma s-adenosylhomocysteine: A cross-sectional study. Nutr. Res. 2017, 46, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Poirier, L.A.; Wise, C.K.; Delongchamp, R.R.; Sinha, R. Blood determinations of S-adenosylmethionine, S-adenosylhomocysteine, and homocysteine: Correlations with diet. Cancer Epidemiol. Biomark. Prev. 2001, 10, 649–655. [Google Scholar]
- Benito, S.; Sanchez-Ortega, A.; Unceta, N.; Jansen, J.J.; Postma, G.; Andrade, F.; Aldamiz-Echevarria, L.; Buydens, L.M.C.; Goicolea, M.A.; Barrio, R.J. Plasma biomarker discovery for early chronic kidney disease diagnosis based on chemometric approaches using LC-QTOF targeted metabolomics data. J. Pharm. Biomed. Anal. 2018, 149, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Kruglova, M.P.; Ivanov, A.V.; Virus, E.D.; Bulgakova, P.O.; Samokhin, A.S.; Fedoseev, A.N.; Grachev, S.V.; Kubatiev, A.A. Urine S-Adenosylmethionine are Related to Degree of Renal Insufficiency in Patients with Chronic Kidney Disease. Lab. Med. 2021, 52, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Linnebank, M.; Popp, J.; Smulders, Y.; Smith, D.; Semmler, A.; Farkas, M.; Kulic, L.; Cvetanovska, G.; Blom, H.; Stoffel-Wagner, B.; et al. S-adenosylmethionine is decreased in the cerebrospinal fluid of patients with Alzheimer’s disease. Neurodegener. Dis. 2010, 7, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liao, R.; Dai, X.; Guo, H.; Wang, D.; Xia, M.; Ling, W.; Xiao, Y. Association between plasma S-adenosylmethionine and risk of mortality in patients with coronary artery disease: A cohort study. Am. J. Clin. Nutr. 2021, 114, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Wexler, O.; Gough, M.S.; Morgan, M.A.M.; Mack, C.M.; Apostolakos, M.J.; Doolin, K.P.; Mooney, R.A.; Arning, E.; Bottiglieri, T.; Pietropaoli, A.P. Methionine Metabolites in Patients with Sepsis. J. Intensive Care Med. 2018, 33, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gerbarg, P.; Bottiglieri, T.; Massoumi, L.; Carpenter, L.L.; Lavretsky, H.; Muskin, P.R.; Brown, R.P.; Mischoulon, D. As Work Group of the American Psychiatric Association Council on, R. S-Adenosylmethionine (SAMe) for Neuropsychiatric Disorders: A Clinician-Oriented Review of Research. J. Clin. Psychiatry 2017, 78, e656–e667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gellekink, H.; van Oppenraaij-Emmerzaal, D.; van Rooij, A.; Struys, E.A.; den Heijer, M.; Blom, H.J. Stable-isotope dilution liquid chromatography-electrospray injection tandem mass spectrometry method for fast, selective measurement of S-adenosylmethionine and S-adenosylhomocysteine in plasma. Clin. Chem. 2005, 51, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Alae-Carew, C.; Green, R.; Stewart, C.; Cook, B.; Dangour, A.D.; Scheelbeek, P.F.D. The role of plant-based alternative foods in sustainable and healthy food systems: Consumption trends in the UK. Sci. Total Environ. 2021, 807, 151041. [Google Scholar] [CrossRef] [PubMed]
- Eustachio Colombo, P.; Elinder, L.S.; Lindroos, A.K.; Parlesak, A. Designing Nutritionally Adequate and Climate-Friendly Diets for Omnivorous, Pescatarian, Vegetarian and Vegan Adolescents in Sweden Using Linear Optimization. Nutrients 2021, 13, 2507. [Google Scholar] [CrossRef]
- Magkos, F.; Tetens, I.; Bugel, S.G.; Felby, C.; Schacht, S.R.; Hill, J.O.; Ravussin, E.; Astrup, A. A Perspective on the Transition to Plant-Based Diets: A Diet Change May Attenuate Climate Change, but Can It Also Attenuate Obesity and Chronic Disease Risk? Adv. Nutr. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moreno, L.A.; Meyer, R.; Donovan, S.M.; Goulet, O.; Haines, J.; Kok, F.J.; Van’t Veer, P. Perspective: Striking a Balance between Planetary and Human Health: Is There a Path Forward? Adv. Nutr. 2021, 13, 355–375. [Google Scholar] [CrossRef]
- Springmann, M.; Clark, M.A.; Rayner, M.; Scarborough, P.; Webb, P. The global and regional costs of healthy and sustainable dietary patterns: A modelling study. Lancet Planet. Health 2021, 5, e797–e807. [Google Scholar] [CrossRef]
- Alexy, U.; Fischer, M.; Weder, S.; Langler, A.; Michalsen, A.; Sputtek, A.; Keller, M. Nutrient Intake and Status of German Children and Adolescents Consuming Vegetarian, Vegan or Omnivore Diets: Results of the VeChi Youth Study. Nutrients 2021, 13, 1707. [Google Scholar] [CrossRef]
- Hovinen, T.; Korkalo, L.; Freese, R.; Skaffari, E.; Isohanni, P.; Niemi, M.; Nevalainen, J.; Gylling, H.; Zamboni, N.; Erkkola, M.; et al. Vegan diet in young children remodels metabolism and challenges the statuses of essential nutrients. EMBO Mol. Med. 2021, 13, e13492. [Google Scholar] [CrossRef] [PubMed]
- Kahleova, H.; McCann, J.; Alwarith, J.; Rembert, E.; Tura, A.; Holubkov, R.; Barnard, N.D. A plant-based diet in overweight adults in a 16-week randomized clinical trial: The role of dietary acid load. Clin. Nutr. ESPEN 2021, 44, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Rees, K.; Al-Khudairy, L.; Takeda, A.; Stranges, S. Vegan dietary pattern for the primary and secondary prevention of cardiovascular diseases. Cochrane Database Syst. Rev. 2021, 2, CD013501. [Google Scholar] [CrossRef]
- Kohnert, E.; Kreutz, C.; Binder, N.; Hannibal, L.; Gorkiewicz, G.; Muller, A.; Storz, M.A.; Huber, R.; Lederer, A.K. Changes in Gut Microbiota after a Four-Week Intervention with Vegan vs. Meat-Rich Diets in Healthy Participants: A Randomized Controlled Trial. Microorganisms 2021, 9, 727. [Google Scholar] [CrossRef]
- Lederer, A.K.; Hannibal, L.; Hettich, M.; Behringer, S.; Spiekerkoetter, U.; Steinborn, C.; Grundemann, C.; Zimmermann-Klemd, A.M.; Muller, A.; Simmet, T.; et al. Vitamin B12 Status Upon Short-Term Intervention with a Vegan Diet-A Randomized Controlled Trial in Healthy Participants. Nutrients 2019, 11, 2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lederer, A.K.; Maul-Pavicic, A.; Hannibal, L.; Hettich, M.; Steinborn, C.; Grundemann, C.; Zimmermann-Klemd, A.M.; Muller, A.; Sehnert, B.; Salzer, U.; et al. Vegan diet reduces neutrophils, monocytes and platelets related to branched-chain amino acids-A randomized, controlled trial. Clin. Nutr. 2020, 39, 3241–3250. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Zimmermann-Klemd, A.M.; Lederer, A.K.; Hannibal, L.; Kowarschik, S.; Huber, R.; Storz, M.A. A Vegan Diet Is Associated with a Significant Reduction in Dietary Acid Load: Post Hoc Analysis of a Randomized Controlled Trial in Healthy Individuals. Int. J. Environ. Res. Public Health 2021, 18, 9998. [Google Scholar] [CrossRef] [PubMed]
- Behringer, S.; Wingert, V.; Oria, V.; Schumann, A.; Grunert, S.; Cieslar-Pobuda, A.; Kolker, S.; Lederer, A.K.; Jacobsen, D.W.; Staerk, J.; et al. Targeted Metabolic Profiling of Methionine Cycle Metabolites and Redox Thiol Pools in Mammalian Plasma, Cells and Urine. Metabolites 2019, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Kirsch, S.H.; Knapp, J.P.; Geisel, J.; Herrmann, W.; Obeid, R. Simultaneous quantification of S-adenosyl methionine and S-adenosyl homocysteine in human plasma by stable-isotope dilution ultra performance liquid chromatography tandem mass spectrometry. J. Chromatogr. B 2009, 877, 3865–3870. [Google Scholar] [CrossRef] [PubMed]
- Struys, E.A.; Jansen, E.E.; de Meer, K.; Jakobs, C. Determination of S-adenosylmethionine and S-adenosylhomocysteine in plasma and cerebrospinal fluid by stable-isotope dilution tandem mass spectrometry. Clin. Chem. 2000, 46, 1650–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzideh, F.; Wosniok, W.; Haeckel, R. Reference limits of plasma and serum creatinine concentrations from intra-laboratory data bases of several German and Italian medical centres: Comparison between direct and indirect procedures. Clin. Chim. Acta 2010, 411, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Lind, M.V.; Lauritzen, L.; Vestergaard, H.; Hansen, T.; Pedersen, O.; Kristensen, M.; Ross, A.B. One-carbon metabolism markers are associated with cardiometabolic risk factors. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 402–410. [Google Scholar] [CrossRef]
- Melnyk, S.; Pogribna, M.; Pogribny, I.P.; Yi, P.; James, S.J. Measurement of plasma and intracellular S-adenosylmethionine and S-adenosylhomocysteine utilizing coulometric electrochemical detection: Alterations with plasma homocysteine and pyridoxal 5′-phosphate concentrations. Clin. Chem. 2000, 46, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Klepacki, J.; Brunner, N.; Schmitz, V.; Klawitter, J.; Christians, U.; Klawitter, J. Development and validation of an LC-MS/MS assay for the quantification of the trans-methylation pathway intermediates S-adenosylmethionine and S-adenosylhomocysteine in human plasma. Clin. Chim. Acta 2013, 421, 91–97. [Google Scholar] [CrossRef] [Green Version]
- James, S.J.; Cutler, P.; Melnyk, S.; Jernigan, S.; Janak, L.; Gaylor, D.W.; Neubrander, J.A. Metabolic biomarkers of increased oxidative stress and impaired methylation capacity in children with autism. Am. J. Clin. Nutr. 2004, 80, 1611–1617. [Google Scholar] [CrossRef] [Green Version]
- Stabler, S.P.; Allen, R.H. Quantification of serum and urinary S-adenosylmethionine and S-adenosylhomocysteine by stable-isotope-dilution liquid chromatography-mass spectrometry. Clin. Chem. 2004, 50, 365–372. [Google Scholar] [CrossRef]
- Guiraud, S.P.; Montoliu, I.; Da Silva, L.; Dayon, L.; Galindo, A.N.; Corthesy, J.; Kussmann, M.; Martin, F.P. High-throughput and simultaneous quantitative analysis of homocysteine-methionine cycle metabolites and co-factors in blood plasma and cerebrospinal fluid by isotope dilution LC-MS/MS. Anal. Bioanal. Chem. 2017, 409, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Kruglova, M.P.; Virus, E.D.; Bulgakova, P.O.; Vital’evich Grachev, S.; Kubatiev, A.A. Determination of S-adenosylmethionine, S-adenosylhomocysteine, and methylthioadenosine in urine using solvent-modified micellar electrokinetic chromatography. Electrophoresis 2020, 41, 209–214. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Virus, E.D.; Nikiforova, K.A.; Kushlinskii, N.E.; Luzyanin, B.P.; Maksimova, M.Y.; Piradov, M.A.; Kubatiev, A.A. Capillary electrophoresis and phenylboronic acid solid phase extraction for the determination of S-adenosylmethionine/S-adenosylhomocysteine ratio in human urine. Electrophoresis 2016, 37, 2663–2669. [Google Scholar] [CrossRef] [PubMed]
- Kruglova, M.P.; Grachev, S.V.; Bulgakova, P.O.; Ivanov, A.V.; Virus, E.D.; Nikiforova, K.A.; Fedoseev, A.N.; Savina, G.D.; Kubatiev, A.A. Low S-adenosylmethionine/S-adenosylhomocysteine Ratio in Urine is Associated with Chronic Kidney Disease. Lab. Med. 2020, 51, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.L.; Hoover, E.; Wang, Y.; Zanetti, K.A. Pre-Analytical Factors that Affect Metabolite Stability in Human Urine, Plasma, and Serum: A Review. Metabolites 2019, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Ntasi, G.; Tsarbopoulos, A.; Mikros, E.; Gikas, E. Targeted Metabolomics: The LC-MS/MS Based Quantification of the Metabolites Involved in the Methylation Biochemical Pathways. Metabolites 2021, 11, 416. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Mato, J.M. S-adenosylmethionine in liver health, injury, and cancer. Physiol. Rev. 2012, 92, 1515–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worcester, E.M.; Bergsland, K.J.; Gillen, D.L.; Coe, F.L. Mechanism for higher urine pH in normal women compared with men. Am. J. Physiol. Renal Physiol. 2018, 314, F623–F629. [Google Scholar] [CrossRef] [PubMed]
- Pigoli, G.; Dorizzi, R.M.; Ferrari, F. Variations of the urinary pH values in a population of 13.000 patients addressing to the National Health System. Minerva Ginecol. 2010, 62, 85–90. [Google Scholar]
- Schumann, A.; Belche, V.; Schaller, K.; Grunert, S.C.; Kaech, A.; Baumgartner, M.R.; Kolker, S.; Hannibal, L.; Spiekerkoetter, U. Mitochondrial damage in renal epithelial cells is potentiated by protein exposure in propionic aciduria. J. Inherit. Metab. Dis. 2021, 44, 1330–1342. [Google Scholar] [CrossRef]
- European Medicines Agency. Note for Guidance on Validation of Analytical Procedures: Text and Methodology (CPMP/ICH/381/95); European Medicines Agency: London, UK, 1995. [Google Scholar]
- Chen, D.; Han, W.; Su, X.; Li, L.; Li, L. Overcoming Sample Matrix Effect in Quantitative Blood Metabolomics Using Chemical Isotope Labeling Liquid Chromatography Mass Spectrometry. Anal. Chem. 2017, 89, 9424–9431. [Google Scholar] [CrossRef]
- Matuszewski, B.K.; Constanzer, M.L.; Chavez-Eng, C.M. Strategies for the assessment of matrix effect in quantitative bioanalytical methods based on HPLC-MS/MS. Anal. Chem. 2003, 75, 3019–3030. [Google Scholar] [CrossRef]
- Trufelli, H.; Palma, P.; Famiglini, G.; Cappiello, A. An overview of matrix effects in liquid chromatography-mass spectrometry. Mass Spectrom. Rev. 2011, 30, 491–509. [Google Scholar] [CrossRef] [PubMed]
- Deutsche Gesellschaft für Ernährung Vollwertige Ernährung. Available online: https://www.dge.de/ernaehrungspraxis/vollwertige-ernaehrung/ (accessed on 14 February 2022).
- R Core Team. 2021 R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 15 March 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Precursor Ion (Da) | Product Ion (Da) | Dwell Time (msec) | RT (min) | DP (V) | EP (V) | CE (V) | CXP (V) |
|---|---|---|---|---|---|---|---|---|
| SAM | 399.1 | 136.1 | 20 | 0.83 | 81 | 10 | 37 | 22 |
| D3-SAM | 402.1 | 136.1 | 20 | 0.83 | 51 | 10 | 32 | 14 |
| SAH | 385.1 | 136.1 | 20 | 1.53 | 46 | 10 | 23 | 12 |
| 13C5-SAH | 390.1 | 136.1 | 20 | 1.53 | 11 | 10 | 23 | 6 |
| SAM (nM) | SAH (nM) | Creatinine (µM) a | SAM/SAH | |
|---|---|---|---|---|
| Mean ± standard deviation | 120.6 ± 18.1 | 21.5 ± 3.3 | 42.8 ± 14.9 | 5.6 ± 1.0 |
| Median (1st quartile–3rd quartile) | 124.0 (106.0–134.0) | 20.9 (19.5–22.9) | 43.2 (32.5–53.8) | 6.0 (4.8–6.5) |
| Specimen * | Plasma | Urine |
|---|---|---|
| Number of subjects | n = 53 | n = 51 |
| SAM (µM) | 0.125 (0.112–0.151) | 0.832 (0.504–1.160) |
| SAH (µM) | 0.055 (0.051–0.061) | 0.079 (0.057–0.109) |
| Creatinine (µM) | 55.5 (47.0–69.4) | 509.0 (323.0–774.5) |
| SAM/SAH | 2.3 (2.0–2.7) | 10.0 (8.8–11.3) |
| SAM/Creatinine (nmol/µmol) | - | 1.609 (1.434–1.906) |
| SAH/Creatinine (nmol/µmol) | - | 0.156 (0.134–0.183) |
| Study | Lind, M. V. et al. [18] | Struys, E. A. et al. [42] (*) | Lind, M. V. et al. [44] | Melnik, S. et al. [45] | Kirsch, S. H. et al. [41] | Klepacki, J. et al. [46] | James, S.J. et al. [47] | Stabler, S. and Allen, R. [48] | Guiraud, S.P. et al. [49] | This Study |
|---|---|---|---|---|---|---|---|---|---|---|
| SAM (nM) | 89.7 ± 15.3 | 104 ± 4.4 | 90.2 ± 14.5 | 73.5 ± 6.7 | 83.6 ± 7.8 | 95.2 ± 21.6 | 96.9 ± 12 | 109 (71–168) | 88.5 ± 18.1 | 120 ± 18 |
| SAH (nM) | 16.5 ± 6.5 | 46.5 ± 1.9 | 16.3 ± 5.6 | 22.6 ± 1.3 | 12.7 ± 1.5 | 30.4 ± 6.2 | 19.4 ± 3.4 | 15 (8–26) | 25.7 ± 9.9 | 21.5 ± 3.3 |
| SAM/SAH | 5.9 | 2.2 | 5.9 | 3.2 | 6.6 | 3.1 | 5.2 | 7.4 | 3.4 | 5.6 |
| N | 118, 47 men and 71 women | 15 women | 118, 47 men and 71 women | 58 healthy women | 31 healthy individuals of which 6 are men | 8 healthy volunteers (47 de novo kidney transplant patients) | 33 healthy children | 48 healthy controls | 6 healthy controls | 33 adult healthy controls |
| Method | LC-MS/MS | LC-MS/MS | LC-MS/MS, | HPLC with coulometric detection | UPLC-MS/MS | LC-MS/MS | HPLC with electrochemical detection | LC-MS/MS | LC-MS/MS | LC-MS/MS |
| Amount of sample | 100 µL plasma | 500 µL plasma | 100 µL plasma | 200 µL plasma | 500 µL plasma | 200 µL plasma | 200 µL plasma | 500 µL plasma, serum or CSF 50 µL urine | 50 µL plasma or CSF | 20 µL plasma (or 20 µL urine, cell lysate, culture medium) |
| Comments | Average age 49 years old, overweight, slightly elevated metabolic features | Pre-menopausal women with normal Hcy | Average age 49 years old, overweight, slightly elevated metabolic features | Healthy adult females, mean age 37.2 years old | Developed by diagnostic laboratory in Germany | FDA validated study USA-Germany cooperation | Children aged 7.4 ± 1.3 years old | Recommended storage: −80 °C and thawing in ice on the day of the assay | SAM and SAH along with other metabolites by SRM | QC: internal. Commercial human plasma spiked with SAM and SAH |
| Sample cleanup | Acidic extraction Neutralisation SPE | Acidic extraction Neutralisation SPE | Acidic extraction Neutralisation SPE | Acidic extraction 0.22 µm filtration | Neutral pH SPE | Acetone extraction (only IS in acidic solution) | Reduction and acidic extraction | Acidic extraction SPE | Reduction Acidic extraction 0.22 µm filtration | Acidic extraction |
| Study | Stabler, S. and Allen, R. [48] | Ivanov, A.V. et al. [50] | Ivanov, A.V. et al. [51] | Kruglova, M. P. et al. [52] | This Study |
|---|---|---|---|---|---|
| SAM (µM) | 19.1–124.8 | 12.1 | 30–60 | 10.2 | 22.1 ± 11.0 |
| SAH (µM) | 0.08–3.1 | 0.73 | 6–30 | 0.89 | 2.2 ± 1.0 |
| SAM/SAH | 4.5–94.4 | 16.58 | 2–5 | 11 | 10.0 ± 1.9 (6.1–14.8) |
| N | 48 (only n = 6 for urine) | 30 urine samples from healthy volunteers | 40 healthy volunteers (20–75 years old) | 20 healthy controls | 53 adult healthy controls (18–60 years old) |
| Method | Stable isotope dilution LC-MS (SIM) | Micellar electrokinetic chromatography (MEKC/CE) | CE/UV detection | CE/UV detection | LC-MS/MS |
| Amount of sample | 50 µL of urine | 300 µL of urine | 2000 µL | 2200 µL of urine | 20 µL urine 1:25 |
| Comments | Recommended storage: −80 °C and thawing in ice on the day of the assay | Median SAM/Creatinine (µM/mM): 1.17 Median SAH/Creatinine (µM/mM): 0.083 | LOD SAM and SAH: 0.07 µmol/L (S/N = 3) LOQ SAM and SAH: 0.2 µmol/L (S/N = 10) Linearity SAM: 0.06-40 µM Linerity SAH: 0.09-7 µM | Mean age of 58.5 years (range, 46–71 years; 43.5% male) SAM/Creatinine 2.33(µmol/mmol) SAH/Creatinine 0.201 (µmol/mmol) | SAM/Creatinine (nmol/µmol): 1.609 (1.434–1.906) SAH/Creatinine (nmol/µmol): 0.156 (0.134–0.183) SAM/SAH: 10.0 (8.8–11.3) |
| Sample Cleanup | Acidic extraction, SPE | SPE | SPE | SPE | Acidic extraction |
| Plasma | GLMM for Plasma | ||||||
| MD | VD | Fixed Effects and Interactions χ2 (p-Value) | |||||
| Baseline | End | Baseline | End | ||||
| (n = 27) | (n = 27) | (n = 26) | (n = 26) | Time | Diet | Time × Diet | |
| SAM (µM) | 0.122 (0.108–0.139) | 0.141 (0.127–0.160) | 0.135 (0.112–0.154) | 0.149 (0.137–0.158) | 4.363 (0.036) | 2.976 (0.084) | 2.029 (0.154) |
| SAH (µM) | 0.054 (0.049–0.061) | 0.057 (0.045–0.060) | 0.055 (0.051–0.060) | 0.057 (0.051–0.063) | 0.073 (0.786) | 1.463 (0.226) | 1.129 (0.287) |
| Creatinine (µM) | 57.7 (43.9–79.5) | 49.1 (34.8–68.5) | 54.5 (48.3–61.7) | 40.7 (30.2–63.1) | 5.009 (0.025) | 1.790 (0.180) | 0.0009 (0.975) |
| SAM/SAH | 2.2 (2.0–2.5) | 2.7 (2.2–3.2) | 2.4 (2.1–2.7) | 2.6 (2.2–3.0) | 4.692 (0.030) | 0.269 (0.603) | 2.996 (0.083) |
| Urine | GLMM for Urine | ||||||
| MD | VD | Fixed Effects and Interactions χ2 (p-Value) | |||||
| Baseline | End | Baseline | End | ||||
| (n = 26) | (n = 26) | (n = 25) | (n = 25) | Time | Diet | Time × Diet | |
| SAM/Creatinine (nmol/µmol) | 1.569 (1.394–1.738) | 1.792 (1.626–2.180) | 1.627 (1.483–2.161) | 2.113 (1.716–3.423) | 26.976 (< 0.001) | 7.101 (0.007) | 2.725 (0.098) |
| SAH/Creatinine (nmol/µmol) | 0.153 (0.128–0.165) | 0.163 (0.132–0.182) | 0.165 (0.143–0.200) | 0.195 (0.160–0.248) | 5.356 (0.020) | 8.025 (0.004) | 2.387 (0.122) |
| SAM/SAH | 10.4 (8.9–11.6) | 11.8 (10.8–12.9) | 9.9 (8.8–10.5) | 11.5 (10.2–12.4) | 23.687 (< 0.001) | 0.374 (0.540) | 0.024 (0.8761) |
| SAM in Plasma | SAH in Plasma | SAM/SAH in Plasma | ||||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| Metabolite_Urine | 0.005 | 0.939 | 5.687 | 0.019 | 0.920 | 0.339 |
| Time | 12.038 | 0.0008 | 0.467 | 0.495 | 11.775 | 0.0009 |
| Diet | 0.735 | 0.393 | 0.678 | 0.412 | 0.000 | 0.975 |
| Metabolite_Urine × Time | 0.53 | 0.468 | 0.220 | 0.639 | 2.162 | 0.144 |
| Metabolite_Urine × Diet | 0.002 | 0.961 | 0.478 | 0.491 | 0.761 | 0.385 |
| Time × Diet | 0.022 | 0.880 | 0.055 | 0.814 | 1.608 | 0.20 |
| Metabolite_Urine × Time × Diet | 0.154 | 0.695 | 2.506 | 0.116 | 0.002 | 0.959 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bravo, A.C.; Aguilera, M.N.L.; Marziali, N.R.; Moritz, L.; Wingert, V.; Klotz, K.; Schumann, A.; Grünert, S.C.; Spiekerkoetter, U.; Berger, U.; et al. Analysis of S-Adenosylmethionine and S-Adenosylhomocysteine: Method Optimisation and Profiling in Healthy Adults upon Short-Term Dietary Intervention. Metabolites 2022, 12, 373. https://doi.org/10.3390/metabo12050373
Bravo AC, Aguilera MNL, Marziali NR, Moritz L, Wingert V, Klotz K, Schumann A, Grünert SC, Spiekerkoetter U, Berger U, et al. Analysis of S-Adenosylmethionine and S-Adenosylhomocysteine: Method Optimisation and Profiling in Healthy Adults upon Short-Term Dietary Intervention. Metabolites. 2022; 12(5):373. https://doi.org/10.3390/metabo12050373
Chicago/Turabian StyleBravo, Aida Corrillero, Maria Nieves Ligero Aguilera, Nahuel R. Marziali, Lennart Moritz, Victoria Wingert, Katharina Klotz, Anke Schumann, Sarah C. Grünert, Ute Spiekerkoetter, Urs Berger, and et al. 2022. "Analysis of S-Adenosylmethionine and S-Adenosylhomocysteine: Method Optimisation and Profiling in Healthy Adults upon Short-Term Dietary Intervention" Metabolites 12, no. 5: 373. https://doi.org/10.3390/metabo12050373