Metabolite Changes in an Estuarine Annelid Following Sublethal Exposure to a Mixture of Zinc and Boscalid

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
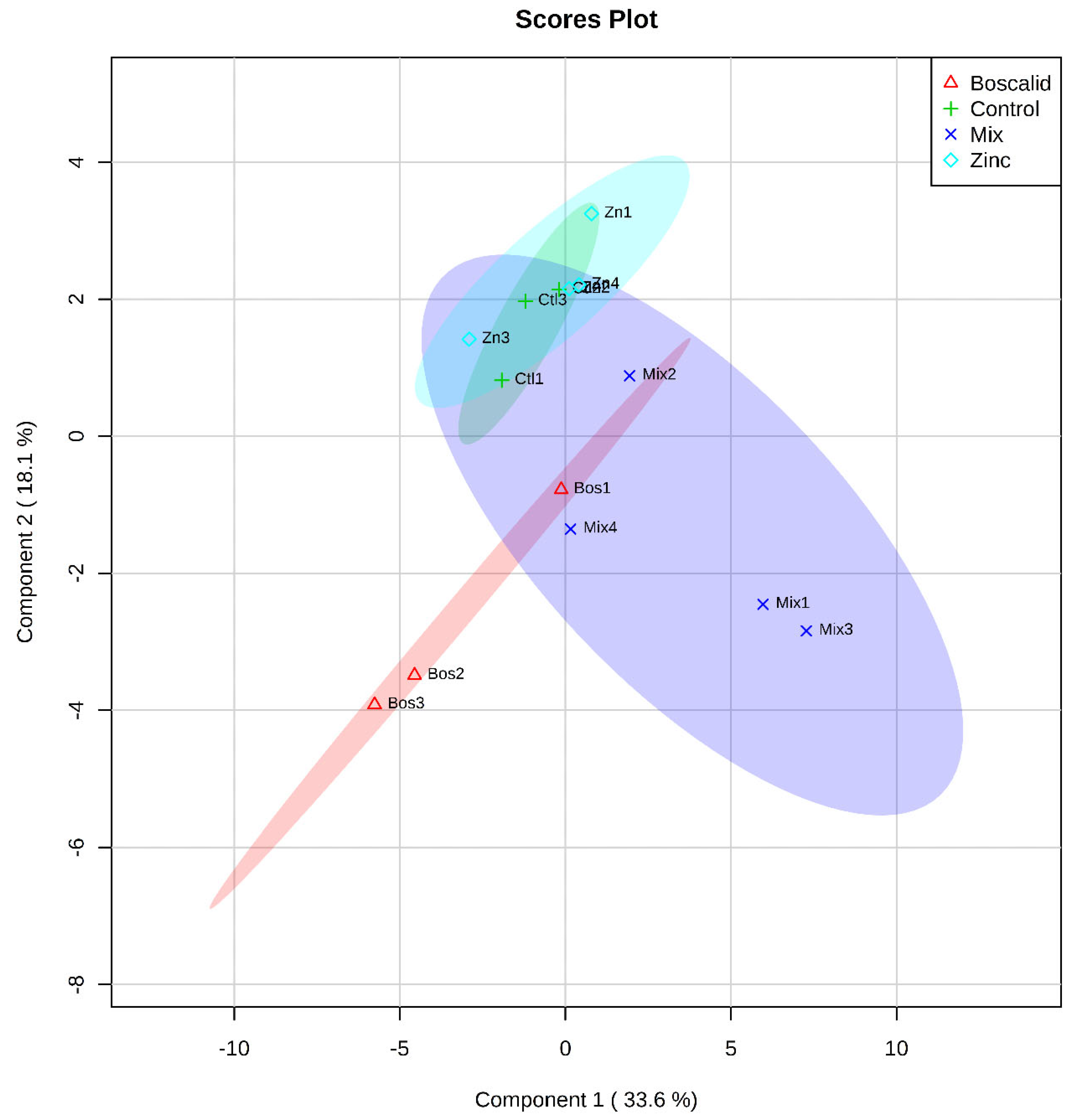
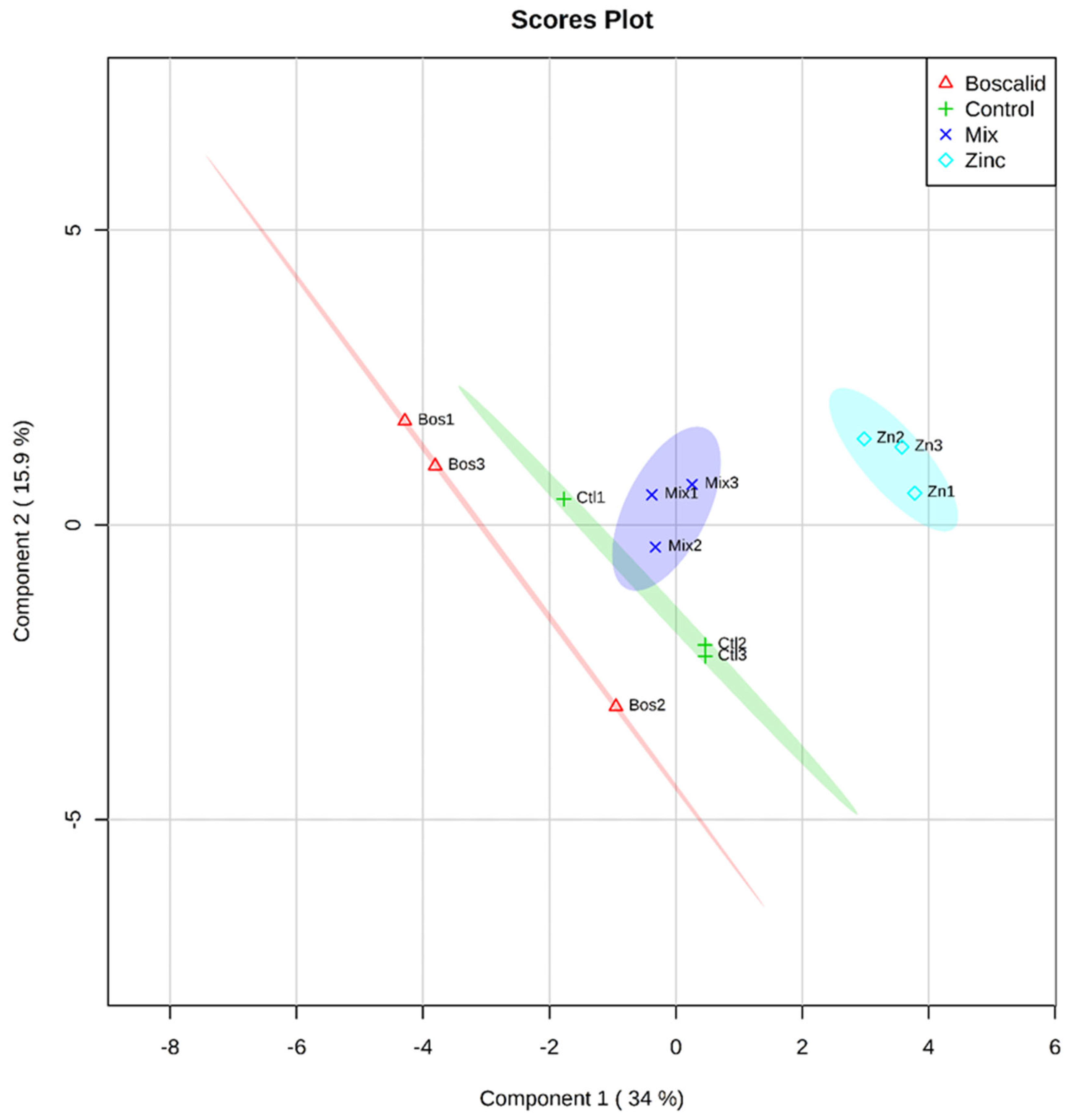
2.1. Multivariate Analysis of Metabolites
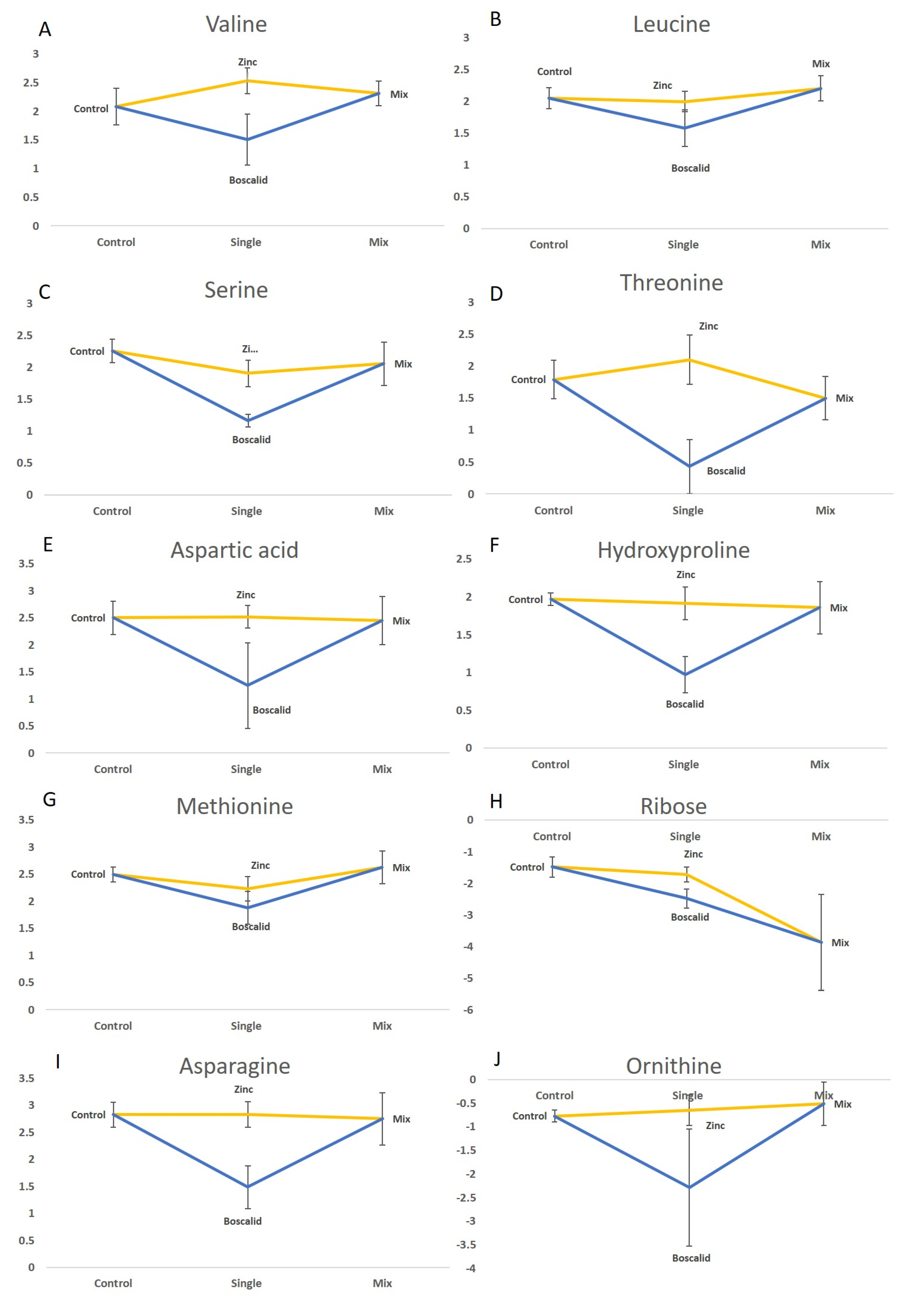
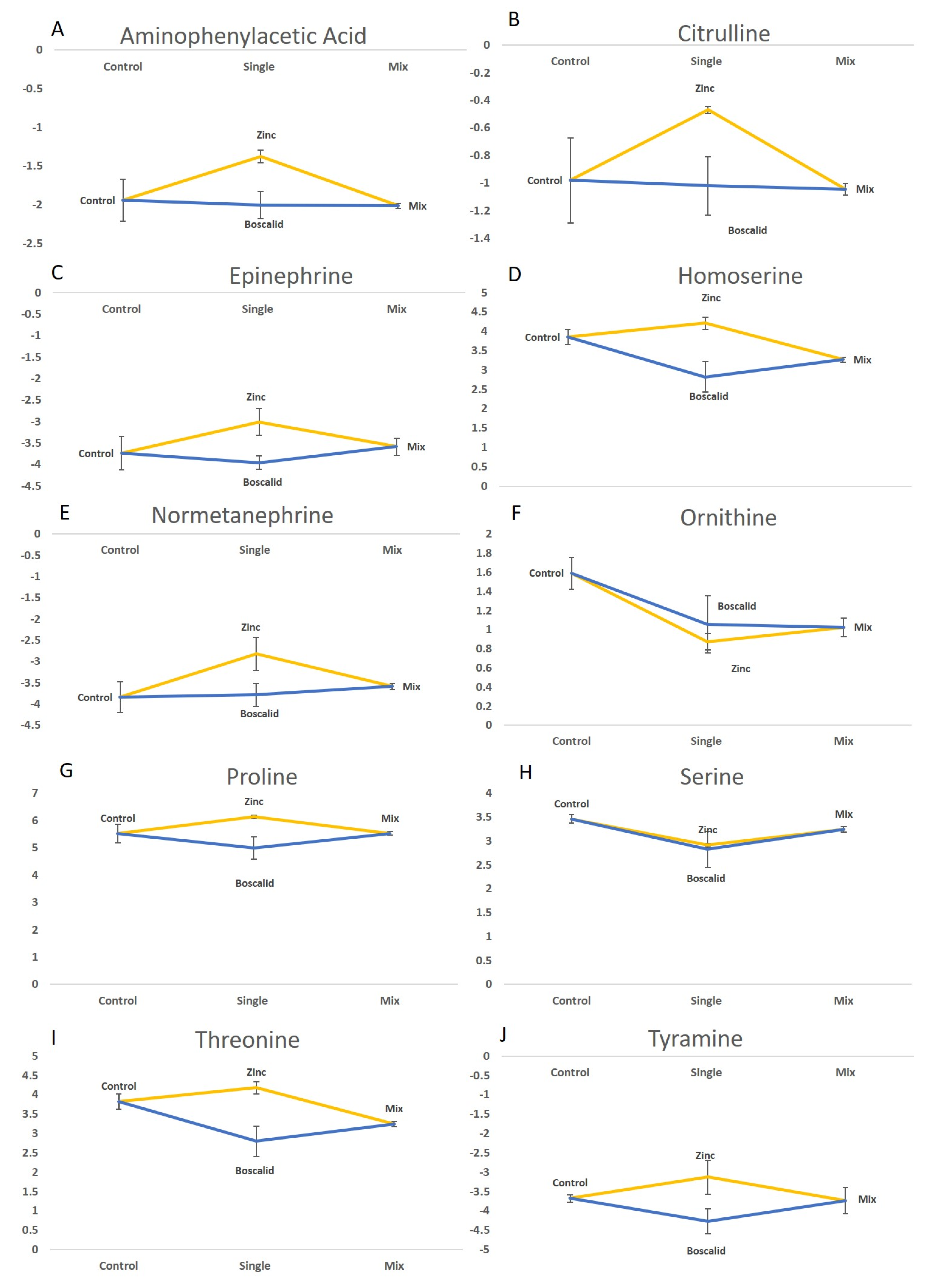
2.2. Univariate Analysis of Treatment Effects on Metabolites Responses
2.3. GC–MS Data
2.4. LC–MS Data
2.5. Combined Metabolite Pathway Responses
3. Conclusions
4. Materials and Methods
4.1. Test Species
4.2. Mixture Exposure
4.3. Sample Preparation
4.4. Metabolite Extraction
4.5. Gas Chromatography-–Mass Spectrometry (GC–MS) and Liquid Chromatography–Mass Spectrometry (LC–MS)—Amine Compounds
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spurgeon, D.J.; Jones, O.A.H.; Dorne, J.-L.C.M.; Svendsen, C.; Swain, S.; Stürzenbaum, S.R. Systems Toxicology Approaches for Understanding the Joint Effects of Environmental Chemical Mixtures. Sci. Total Environ. 2010, 408, 3725–3734. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Kirwan, J.A.; Johnson, C.; Yan, N.D.; Viant, M.R.; Gunn, J.M.; McGeer, J.C. Predicting Chronic Copper and Nickel Reproductive Toxicity to Daphnia Pulex-Pulicaria from Whole-Animal Metabolic Profiles. Environ. Pollut. 2016, 212, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.S.H.; El-Masri, H.A.; Thomas, R.S.; Dobrev, I.D.; Dennison, J.E.; Bae, D.S.; Campain, J.A.; Liao, K.H.; Reisfeld, B.; Andersen, M.E.; et al. Chemical Mixture Toxicology: From Descriptive to Mechanistic, and Going on to in Silico Toxicology. Environ. Toxicol. Pharmacol. 2004, 18, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, K.C.; Lingenfelter, R.; Cizmas, L.; Falahatpisheh, M.H.; Qian, Y.; Tang, Y.; Garcia, S.; Ramos, K.; Tiffany-Castiglioni, E.; Mumtaz, M.M. Toxicity Assessment of Complex Mixtures Remains a Goal. Environ. Toxicol. Pharmacol. 2004, 18, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Jeppe, K.J.; Carew, M.E.; Pettigrove, V.; Hoffmannz, A.A.; Hoffmann, A.A. Toxicant Mixtures in Sediment Alter Gene Expression in The Cysteine Metabolism of Chironomus tepperi. Environ. Toxicol. Chem. 2017, 36, 691–698. [Google Scholar] [CrossRef]
- Jones, O.A.H.; Murfitt, S.; Svendsen, C.; Turk, A.; Turk, H.; Spurgeon, D.J.; Walker, L.A.; Shore, R.F.; Long, S.M.; Griffin, J.L. Comparisons of Metabolic and Physiological Changes in Rats Following Short Term Oral Dosing with Pesticides Commonly Found in Food. Food Chem. Toxicol. 2013, 59, 438–445. [Google Scholar] [CrossRef]
- Tang, G.; Yao, J.; Zhang, X.; Lu, N.; Zhu, K.Y. Comparison of Gene Expression Profiles in the Aquatic Midge (Chironomus tentans) Larvae Exposed to Two Major Agricultural Pesticides. Chemosphere 2018, 194, 745–754. [Google Scholar] [CrossRef]
- Sinclair, G.M.; O’Brien, A.L.; Keough, M.; De Souza, D.P.; Dayalan, S.; Kanojia, K.; Kouremenos, K.; Tull, D.L.; Coleman, R.A.; Jones, O.A.H.; et al. Using Metabolomics to Assess the Sub-Lethal Effects of Zinc and Boscalid on an Estuarine Polychaete Worm over Time. Metabolomics 2019, 15, 108. [Google Scholar] [CrossRef]
- Jones, O.A.H.H.; Swain, S.C.; Svendsen, C.; Griffin, J.L.; Sturzenbaum, S.R.; Spurgeon, D.J. Potential New Method of Mixture Effects Testing Using Metabolomics and Caenorhabditis elegans. J. Proteome Res. 2012, 11, 1446–1453. [Google Scholar] [CrossRef]
- Long, S.M.; Tull, D.L.; Jeppe, K.J.; De Souza, D.P.; Dayalan, S.; Pettigrove, V.J.; McConville, M.J.; Hoffmann, A.A. A Multi-Platform Metabolomics Approach Demonstrates Changes in Energy Metabolism and the Transsulfuration Pathway in Chironomus tepperi Following Exposure to Zinc. Aquat. Toxicol. 2015, 162, 54–65. [Google Scholar] [CrossRef]
- Lankadurai, B.P.; Nagato, E.G.; Simpson, M.J. Environmental Metabolomics: An Emerging Approach to Study Organism Responses to Environmental Stressors. Environ. Rev. 2013, 21, 180–205. [Google Scholar] [CrossRef]
- Baylay, A.J.; Spurgeon, D.J.; Svendsen, C.; Griffin, J.L.; Swain, S.C.; Sturzenbaum, S.R.; Jones, O.A.H. A Metabolomics Based Test of Independent Action and Concentration Addition Using the Earthworm Lumbricus rubellus. Ecotoxicology 2012, 21, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Vandenbrouck, T.; Jones, O.A.H.; Dom, N.; Griffin, J.L.; De Coen, W. Mixtures of Similarly Acting Compounds in Daphnia magna: From Gene to Metabolite and Beyond. Environ. Int. 2010, 36, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.T.; Keough, M.J.; Long, S.M.; Pettigrove, V.J. Effects of The Boscalid Fungicide Filan 1 on The Marine Amphipod Allorchestes compressa At Environmentally Relevant Concentrations. Env. Toxicol Chem 2016, 35, 1130–1137. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.; Pettigrove, V.; Carew, M.; Hoffmann, A. Isolating the Impact of Sediment Toxicity in Urban Streams. Environ. Pollut. 2010, 158, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Pettigrove, V.; Hoffmann, A. A Field-Based Microcosm Method to Assess the Effects of Polluted Urban Stream Sediments on Aquatic Macroinvertebrates. Environ. Toxicol. Chem. 2005, 24, 170. [Google Scholar] [CrossRef]
- Kellar, C.R.; Hassell, K.L.; Long, S.M.; Myers, J.H.; Golding, L.; Rose, G.; Kumar, A.; Hoffmann, A.A.; Pettigrove, V. Ecological Evidence Links Adverse Biological Effects to Pesticide and Metal Contamination in an Urban Australian Watershed. J. Appl. Ecol. 2014, 51, 426–439. [Google Scholar] [CrossRef]
- O’Brien, A.L.; Dafforn, K.A.; Chariton, A.A.; Johnston, E.L.; Mayer-Pinto, M. After Decades of Stressor Research in Urban Estuarine Ecosystems the Focus Is Still on Single Stressors: A Systematic Literature Review and Meta-Analysis. Sci. Total Environ. 2019, 684, 753–764. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). Wiley Statsref Stat. Ref. Online 2017, 1–15. [Google Scholar] [CrossRef]
- Szymańska, E.; Saccenti, E.; Smilde, A.K.; Westerhuis, J.A. Double-Check: Validation of Diagnostic Statistics for PLS-DA Models in Metabolomics Studies. Metabolomics 2012, 8, 3–16. [Google Scholar] [CrossRef]
- Relyea, R.; Hoverman, J. Assessing the Ecology in Ecotoxicology: A Review and Synthesis in Freshwater Systems. Ecol. Lett. 2006, 9, 1157–1171. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.A.; Hill, C.B.; Jayasinghe, N.S.; Atieno, J.; Sutton, T.; Roessner, U. Quantitative Profiling of Polar Primary Metabolites of Two Chickpea Cultivars with Contrasting Responses to Salinity. J. Chromatogr. B 2015, 1000, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Nunes-Nesi, A.; Nikoloski, Z.; Sweetlove, L.J.; Fernie, A.R. Metabolic Control and Regulation of the Tricarboxylic Acid Cycle in Photosynthetic and Heterotrophic Plant Tissues. Plant. Cell Environ. 2012, 35, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, J.H.F.; Esteves-Ferreira, A.A.; Quinhones, C.G.S.; Pereira-Lima, I.A.; Nunes-Nesi, A.; Fernie, A.R.; Araujo, W.L. Evolution and Functional Implications of the Tricarboxylic Acid Cycle as Revealed by Phylogenetic Analysis. Genome Biol. Evol. 2014, 6, 2830–2848. [Google Scholar] [CrossRef] [Green Version]
- Suárez, M.; Caimari, A.; del Bas, J.M.; Arola, L. Metabolomics: An Emerging Tool to Evaluate the Impact of Nutritional and Physiological Challenges. TrAC Trends Anal. Chem. 2017, 96, 79–88. [Google Scholar]
- Felföldi, G.; Eleftherianos, I. Plays a Central Role in Insect Immunity A Serine Proteinase Homologue, SPH-3, Ffrench-Constant and István Venekei. J. Immunol. Ref. 2011, 186, 4828–4834. [Google Scholar] [CrossRef]
- Watanabe, M.; Tohge, T.; Fernie, A.R.; Hoefgen, R. The Effect of Single and Multiple SERAT Mutants on Serine and Sulfur Metabolism. Front. Plant Sci. 2018, 9, 702. [Google Scholar] [CrossRef]
- Fuchs, S.; Bundy, J.G.; Davies, S.K.; Viney, J.M.; Swire, J.S.; Leroi, A.M. A Metabolic Signature of Long Life in Caenorhabditis elegans. BMC Biol. 2010, 8, 1–2. [Google Scholar] [CrossRef]
- Pant, A.; Cao, S.; Yang, Z.; Shisler, J.L. Asparagine Is a Critical Limiting Metabolite for Vaccinia Virus Protein Synthesis during Glutamine Deprivation Virus-Cell Interactions. J. Virol. 2019, 93, 1834–1852. [Google Scholar] [CrossRef]
- Kruger, N.J.; von Schaewen, A. The Oxidative Pentose Phosphate Pathway: Structure and Organisation. Curr. Opin. Plant Biol. 2003, 6, 236–246. [Google Scholar] [CrossRef]
- Deutz, N.E.; Simbo, S.Y.; Ligthart-Melis, G.C.; Cynober, L.; Smriga, M.; Engelen, M.P. Tolerance to Increased Supplemented Dietary Intakes of Methionine in Healthy Older Adults. Am. J. Clin. Nutr. 2017, 106, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Viant, M.R. Metabolomics of Aquatic Organisms: The New ‘Omics’ on the Block. Mar. Ecol. Prog. Ser. 2007, 332, 301–306. [Google Scholar] [CrossRef]
- Longnecker, K.; Futrelle, J.; Coburn, E.; Kido Soule, M.C.; Kujawinski, E.B. Environmental Metabolomics: Databases and Tools for Data Analysis. Mar. Chem. 2015, 177, 366–373. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, R.; Zhang, Y.; Su, H.; Wang, P.; Jenkins, A.; Ferrier, R.C.; Bailey, M.; Squire, G. Ecosystem Health towards Sustainability. Ecosyst. Heal. Sustain. 2015, 1, 1–15. [Google Scholar] [CrossRef]
- Dondero, F.; Banni, M.; Negri, A.; Boatti, L.; Dagnino, A.; Viarengo, A. Interactions of a Pesticide/Heavy Metal Mixture in Marine Bivalves: A Transcriptomic Assessment. Bmc Genomics. 2011, 12, 195. [Google Scholar] [CrossRef]
- Song, Q.; Zheng, P.; Qiu, L.; Jiang, X.; Zhao, H.; Zhou, H.; Han, Q.; Diao, X. Toxic Effects of Male Perna Viridis Gonad Exposed to BaP, DDT and Their Mixture: A Metabolomic and Proteomic Study of the Underlying Mechanism. Toxicol. Lett. 2016, 240, 185–195. [Google Scholar] [CrossRef]
- Søfteland, L.; Kirwan, J.A.; Hori, T.S.F.; Størseth, T.R.; Sommer, U.; Berntssen, M.H.G.; Viant, M.R.; Rise, M.L.; Waagbø, R.; Torstensen, B.E.; et al. Toxicological Effect of Single Contaminants and Contaminant Mixtures Associated with Plant Ingredients in Novel Salmon Feeds. Food Chem. Toxicol. 2014. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy Homeostasis as an Integrative Tool for Assessing Limits of Environmental Stress Tolerance in Aquatic Invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef]
- Watford, M. The Urea Cycle: Teaching Intermediary Metabolism in a Physiological Setting. Biochem. Mol. Biol. Educ. 2003, 31, 289–297. [Google Scholar] [CrossRef]
- Jones, O.A.H.; Dondero, F.; Viarengo, A.; Griffin, J.L. Metabolic Profiling of Mytilus Galloprovincialis and Its Potential Applications for Pollution Assessment. Mar. Ecol. Prog. Ser. 2008, 369, 169–179. [Google Scholar] [CrossRef]
- French, C.J.; Mommsen, T.P.; Hochachka, P.W. Amino Acid Utilisation in Isolated Hepatocytes from Rainbow Trout. Eur. J. Biochem. 1981, 113, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.L.; Bundy, J.G.; Want, E.J.; Kille, P.; Stürzenbaum, S.R. The Metabolomic Responses of Caenorhabditis elegans to Cadmium Are Largely Independent of Metallothionein Status, but Dominated by Changes in Cystathionine and Phytochelatins. J. Proteome Res. 2009, 8, 3512–3519. [Google Scholar] [CrossRef] [PubMed]
- Elskus, A.A. Toxicity, Sublethal Effects, and Potential Modes of Action of Select Fungicides on Freshwater Fish and Invertebrates Open-File Report 2012–1213. U.S. Dep. Inter. U.S. Geol. Surv. 2017. [Google Scholar] [CrossRef]
- Van den Brink, P.J.; Bracewell, S.A.; Bush, A.; Chariton, A.; Choung, C.B.; Compson, Z.G.; Dafforn, K.A.; Korbel, K.; Lapen, D.R.; Mayer-Pinto, M.; et al. Towards a General Framework for the Assessment of Interactive Effects of Multiple Stressors on Aquatic Ecosystems: Results from the Making Aquatic Ecosystems Great Again (MAEGA) Workshop. Sci. Total Environ. 2019, 684, 722–726. [Google Scholar] [CrossRef]
- O’Brien, A.L. What Are the Roadblocks to Using Population Models in Ecotoxicology Studies? Mar. Pollut. Bull. 2017, 124, 5–8. [Google Scholar] [CrossRef]
- Saunders, E.C.; de Souza, D.P.; Chambers, J.M.; Ng, M.; Pyke, J.; McConville, M.J. Use of 13C Stable Isotope Labelling for Pathway and Metabolic Flux Analysis in Leishmania Parasites. Hum. Pressnew Yorkny. 2015, 1201, 281–296. [Google Scholar]
- Jones, O.A.H.; Spurgeon, D.J.; Svendsen, C.; Griffin, J.L. A Metabolomics Based Approach to Assessing the Toxicity of the Polyaromatic Hydrocarbon Pyrene to the Earthworm Lumbricus rubellus. Chemosphere 2008, 71, 601–609. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental Metabolomics: A Critical Review and Future Perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Nel, L.; Strydom, N.A.; Bouwman, H. Preliminary Assessment of Contaminants in the Sediment and Organisms of the Swartkops Estuary, South Africa. Mar. Pollut. Bull. 2015, 101, 878–885. [Google Scholar] [CrossRef]
- Overgaard, A.J.; Weir, J.M.; De Souza, D.P.; Tull, D.; Haase, C.; Meikle, P.J.; Pociot, F. Lipidomic and Metabolomic Characterization of a Genetically Modified Mouse Model of the Early Stages of Human Type 1 Diabetes Pathogenesis. Metabolomics 2016, 12, 13. [Google Scholar] [CrossRef]
- Boughton, B.A.; Callahan, D.L.; Silva, C.; Bowne, J.; Nahid, A.; Rupasinghe, T.; Tull, D.L.; Mcconville, M.J.; Bacic, A.; Roessner, U.; et al. Comprehensive Profiling and Quantitation of Amine Group Containing Metabolites. Anal. Chem. 2011, 83, 7523–7530. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 3rd ed.; Primer-E Ltd.: Plymouth, UK, 2015; pp. 1–17. [Google Scholar]
- Chong, J.; Yamamoto, M.; Xia, J.; Chong, J.; Yamamoto, M.; Xia, J. MetaboAnalystR 2.0: From Raw Spectra to Biological Insights. Metabolites 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinforma. 2016, 55, 14.10.1-14.10.91. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | GC–MS Metabolites | LC–MS Metabolites | ||||
|---|---|---|---|---|---|---|
| Df | MS | P | df | MS | P | |
| Treatment | 3 | 72.14 | 0.0691 | 3 | 5.69 × 1015 | 0.0006 1 |
| Residual | 10 | 34.973 | 8 | 1.17 × 1015 | ||
| Pairwise | t | P | T | P | ||
| Ctl, Bos | 1.0406 | 0.3024 | 1.5462 | 0.2041 | ||
| Mix, Bos | 1.5246 | 0.2004 | 2.4663 | 0.0999 | ||
| Zn, Bos | 1.3158 | 0.1404 | 2.1502 | 0.0994 | ||
| Ctl, Mix | 1.5209 | 0.1722 | 1.8296 | 0.1028 | ||
| Ctl, Zn | 0.7768 | 0.7968 | 1.7654 | 0.1960 | ||
| Zn, Mix | 1.6489 | 0.0301 1 | 4.2502 | 0.0975 | ||
| Metabolite | MS Residual | Treatment | Comparison | ||||||
|---|---|---|---|---|---|---|---|---|---|
| GC–MS | df | 10 | 3 | Ctl–Bos | Mix–Bos | Zn–Bos | Mix–Ctl | Zn–Ctl | Zn–Mix |
| Valine | 0.128 | 0.022 2 | 0.259 | 0.061 | 0.017 2 | 0.843 | 0.409 | 0.821 | |
| Leucine | 0.062 | 0.043 2 | 0.161 | 0.034 2 | 0.103 | 0.837 | 0.999 | 0.873 | |
| Serine | 0.076 | 0.003 2 | 0.003 | 0.008 2 | 0.023 2 | 0.780 | 0.387 | 0.862 | |
| Threonine | 0.187 | 0.003 2 | 0.014 | 0.037 2 | 0.002 2 | 0.816 | 0.780 | 0.260 | |
| Aspartic acid | 0.311 | 0.044 2 | 0.081 | 0.074 | 0.056 | 0.999 | 1.000 | 0.997 | |
| Hydroxyproline | 0.086 | 0.005 2 | 0.009 2 | 0.012 2 | 0.008 2 | 0.957 | 0.995 | 0.992 | |
| Methionine | 0.089 | 0.040 2 | 0.115 | 0.035 2 | 0.452 | 0.938 | 0.660 | 0.296 | |
| Ribose | 1.006 | 0.034 2 | 0.629 | 0.317 | 0.764 | 0.045 2 | 0.988 | 0.052 | |
| Asparagine | 0.178 | 0.006 2 | 0.0132 | 0.013 2 | 0.009 2 | 0.995 | 1.000 | 0.994 | |
| Ornithine | 0.587 | 0.050 2 | 0.136 | 0.053 | 0.076 | 0.076 | 0.997 | 0.994 | |
| LC–MS | df | 8 | 3 | ||||||
| Aminophenylacetic acid | 0.043 2 | 0.014 2 | 0.983 | 1.000 | 0.024 2 | 0.973 | 0.039 2 | 0.022 2 | |
| Citrulline | 0.053 | 0.046 2 | 0.997 | 0.999 | 0.074 | 0.985 | 0.099 | 0.061 | |
| Epinephrine | 0.116 | 0.046 2 | 0.859 | 0.572 | 0.038 2 | 0.460 | 0.114 | 0.240 | |
| Homoserine | 0.081 | 0.002 2 | 0.009 2 | 0.290 | 0.002 2 | 0.132 | 0.468 | 0.016 2 | |
| Normetanephrine | 0.135 | 0.032 2 | 0.997 | 0.910 | 0.050 2 | 0.833 | 0.038 2 | 0.125 | |
| Ornithine | 0.048 | 0.014 2 | 0.060 | 1.000 | 0.608 | 0.063 | 0.011 2 | 0.589 | |
| Proline | 0.106 | 0.018 2 | 0.257 | 0.260 | 0.011 2 | 1.000 | 0.180 | 0.177 | |
| Serine | 0.059 | 0.042 2 | 0.050 2 | 0.231 | 0.968 | 0.701 | 0.094 | 0.404 | |
| Threonine | 0.083 | 0.002 2 | 0.010 2 | 0.300 | 0.002 2 | 0.138 | 0.473 | 0.017 2 | |
| Tyramine | 0.156 | 0.047 2 | 0.321 | 0.401 | 0.0312 | 0.998 | 0.386 | 0.308 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinclair, G.M.; O’Brien, A.L.; Keough, M.; de Souza, D.P.; Dayalan, S.; Kanojia, K.; Kouremenos, K.; Tull, D.L.; Coleman, R.A.; Jones, O.A.H.; et al. Metabolite Changes in an Estuarine Annelid Following Sublethal Exposure to a Mixture of Zinc and Boscalid. Metabolites 2019, 9, 229. https://doi.org/10.3390/metabo9100229
Sinclair GM, O’Brien AL, Keough M, de Souza DP, Dayalan S, Kanojia K, Kouremenos K, Tull DL, Coleman RA, Jones OAH, et al. Metabolite Changes in an Estuarine Annelid Following Sublethal Exposure to a Mixture of Zinc and Boscalid. Metabolites. 2019; 9(10):229. https://doi.org/10.3390/metabo9100229
Chicago/Turabian StyleSinclair, Georgia M., Allyson L. O’Brien, Michael Keough, David P. de Souza, Saravanan Dayalan, Komal Kanojia, Konstantinos Kouremenos, Dedreia L. Tull, Rhys A. Coleman, Oliver A.H. Jones, and et al. 2019. "Metabolite Changes in an Estuarine Annelid Following Sublethal Exposure to a Mixture of Zinc and Boscalid" Metabolites 9, no. 10: 229. https://doi.org/10.3390/metabo9100229