
Genomic Features of the Bundle-Forming Heliobacterium Heliophilum fasciatum

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. General Genomic Properties
3.2. Phototrophy
3.3. Central Carbon Metabolism
3.4. Nitrogen Metabolism: Utilization of Fixed Nitrogen
3.5. Nitrogen Metabolism: Nitrogen Fixation
3.6. Sulfur Metabolism
3.7. Motility
3.8. Sporulation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gest, H.; Favinger, J.L. Heliobacterium chlorum, an anoxygenic brownish-green photosynthetic bacterium containing a “new” form of bacteriochlorophyll. Arch. Microbiol. 1983, 136, 11–16. [Google Scholar] [CrossRef]
- Madigan, M.T.; Ormerod, J.G. Taxonomy, physiology, and ecology of heliobacteria. In Anoxygenic Photosynthetic Bacteria; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; Volume 2, pp. 17–30. ISBN 0-306-47954-0. [Google Scholar]
- Brockmann, H.; Lipinski, A. Bacteriochlorophyll g. A new bacteriochlorophyll from Heliobacterium chlorum. Arch. Microbiol. 1983, 136, 17–19. [Google Scholar] [CrossRef]
- Sattley, W.M.; Madigan, M.T. The Family Heliobacteriaceae. In The Prokaryotes—Firmicutes and Tenericutes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 185–196. ISBN 978-3-642-30120-9. [Google Scholar]
- Orf, G.S.; Redding, K.E. The Heliobacteria. In Encyclopedia of Biological Chemistry; Joseph, J., Ed.; Elsevier: Oxford, UK, 2021; Volume 2, pp. 352–364. [Google Scholar]
- Kimble-Long, L.K.; Madigan, M.T. Molecular evidence that the capacity for endosporulation is universal among phototrophic heliobacteria. FEMS Microbiol. Lett. 2001, 199, 191–195. [Google Scholar] [CrossRef]
- Stevenson, A.K.; Kimble, L.K.; Woese, C.R.; Madigan, M.T. Characterization of new phototrophic heliobacteria and their habitats. Photosynth. Res. 1997, 53, 1–12. [Google Scholar] [CrossRef]
- Kyndt, J.A.; Salama, D.M.; Meyer, T.E.; Imhoff, J.F. Phylogenetic relationship of phototrophic heliobacteria and systematic reconsideration of species and genus assignments based on genome sequences of eight species. Int. J. Syst. Evol. Microbiol. 2021, 71, 4729. [Google Scholar] [CrossRef]
- Kimble, L.K.; Mandelco, L.; Woese, C.R.; Madigan, M.T. Heliobacterium modesticaldum, sp. nov., a thermophilic heliobacterium of hot springs and volcanic soils. Arch. Microbiol. 1995, 163, 259–267. [Google Scholar] [CrossRef]
- Sattley, W.M.; Madigan, M.T.; Swingley, W.D.; Cheung, P.C.; Clocksin, K.M.; Conrad, A.L.; Dejesa, L.C.; Honchak, B.M.; Jung, D.; Karbach, L.E.; et al. The Genome of Heliobacterium modesticaldum, a Phototrophic Representative of the Firmicutes Containing the Simplest Photosynthetic Apparatus. J. Bacteriol. 2008, 190, 4687–4696. [Google Scholar] [CrossRef] [Green Version]
- Sattley, W.M.; Swingley, W.D. Properties and evolutionary implications of the heliobacterial genome. In Genome Evolution of Photosynthetic Bacteria; Beatty, T.J., Ed.; Academic Press: San Diego, CA, USA, 2013; Volume 66, pp. 67–98. ISBN 9780123979230. [Google Scholar]
- Asao, M.; Jung, D.; Achenbach, L.A.; Madigan, M.T. Heliorestis convoluta sp. nov., a coiled, alkaliphilic heliobacterium from the Wadi El Natroun, Egypt. Extremophiles 2006, 10, 403–410. [Google Scholar] [CrossRef]
- Dewey, E.D.; Stokes, L.M.; Burchell, B.M.; Shaffer, K.N.; Huntington, A.M.; Baker, J.M.; Nadendla, S.; Giglio, M.G.; Bender, K.S.; Touchman, J.W.; et al. Analysis of the Complete Genome of the Alkaliphilic and Phototrophic Firmicute Heliorestis convoluta Strain HHT. Microorganisms 2020, 8, 313. [Google Scholar] [CrossRef] [Green Version]
- Asao, M.; Takaichi, S.; Madigan, M.T. Amino acid-assimilating phototrophic heliobacteria from soda lake environments: Heliorestis acidaminivorans sp. nov. and ‘Candidatus Heliomonas lunata’. Extremophiles 2012, 16, 585–595. [Google Scholar] [CrossRef]
- Kimble, L.K.; Madigan, M.T. Nitrogen fixation and nitrogen metabolism in heliobacteria. Arch. Microbiol. 1992, 158, 155–161. [Google Scholar] [CrossRef]
- Ormerod, J.G.; Kimble, L.K.; Nesbakken, T.; Torgersen, Y.A.; Woese, C.R.; Madigan, M.T. Heliophilum fasciatum gen. nov. sp. nov. and Heliobacterium gestii sp. nov.: Endospore-forming heliobacteria from rice field soils. Arch. Microbiol. 1996, 165, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.-H.; Yue, H.; Blankenship, R.E. Energy metabolism of Heliobacterium modesticaldum during phototrophic and chemotrophic growth. BMC Microbiol. 2010, 10, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gappled BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Raymond, J.; Siefert, J.L.; Staples, C.R.; Blankenship, R.E. The Natural History of Nitrogen Fixation. Mol. Biol. Evol. 2004, 21, 541–554. [Google Scholar] [CrossRef] [Green Version]
- North, J.A.; Narrowe, A.B.; Xiong, W.; Byerly, K.M.; Zhao, G.; Young, S.J.; Murali, S.; Wildenthal, J.A.; Cannon, W.R.; Wrighton, K.C.; et al. A nitrogenase-like enzyme system catalyzes methionine, ethylene, and methane biogenesis. Science 2020, 369, 1094–1098. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Bryantseva, I.A.; Gorlenko, V.M.; Kompantseva, E.I.; Achenbach, L.A.; Madigan, M.T. Heliorestis daurensis, gen. nov. sp. nov., an alkaliphilic rod-to-coiled-shaped phototrophic heliobacterium from a Siberian soda lake. Arch. Microbiol. 1999, 172, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Bryantseva, I.A.; Gorlenko, V.M.; Kompantseva, E.I.; Tourova, T.P.; Kuznetsov, B.B.; Osipov, G.A. Alkaliphilic heliobacterium Heliorestis baculata sp. nov. and emended description of the genus Heliorestis. Arch. Microbiol. 2000, 174, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Briantseva, I.A.; Gorlenko, V.M.; Turova, T.P.; Kuznetsov, B.B.; Lysenko, A.M.; Bykova, S.A.; Gal’Chenko, V.F.; Mitiushina, L.L.; Osipov, G.A. Heliobacterium sulfidophilum sp. nov. and Heliobacterum undosum sp. nov.: Sulfide-oxidizing heliobacteria from thermal sulfidic springs. Mikrobiologia 2000, 69, 396–406. [Google Scholar]
- Kimble, L.K.; Stevenson, A.K.; Madigan, M.T. Chemotrophic growth of heliobacteria in darkness. FEMS Microbiol. Lett. 1994, 115, 51–55. [Google Scholar] [CrossRef]
- Pickett, M.W.; Williamson, M.P.; Kelly, D.J. An enzyme and13C-NMR study of carbon metabolism in heliobacteria. Photosynth. Res. 1994, 41, 75–88. [Google Scholar] [CrossRef]
- Dos Santos, P.C.; Fang, Z.; Mason, S.W.; Setubal, J.C.; Dixon, R. Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes. BMC Genom. 2012, 13, 162. [Google Scholar] [CrossRef] [Green Version]
- Enkh-Amgalan, J.; Kawasaki, H.; Seki, T. NifH and NifD sequences of heliobacteria: A new lineage in the nitrogenase phylogeny. FEMS Microbiol. Lett. 2005, 243, 73–79. [Google Scholar] [CrossRef]
- Garcia, A.K.; McShea, H.; Kolaczkowski, B.; Kaçar, B. Reconstructing the evolutionary history of nitrogenases: Evidence for ancestral molybdenum-cofactor utilization. Geobiology 2020, 18, 394–411. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Nishihara, A.; Haruta, S. Nitrogen-fixing Ability and Nitrogen Fixation-related Genes of Thermophilic Fermentative Bacteria in the Genus Caldicellulosiruptor. Microbes Environ. 2021, 36, ME21018. [Google Scholar] [CrossRef]
- Lie, T.J.; Kuo, Y.P.; Leite, M.; Costa, K.C.; Harwood, C.S.; Leigh, J.A. A genetic study of Nif-associated genes in a hyperthermophilic methanogen. Microbiol. Spectr. 2022, 10, e02093-21. [Google Scholar] [CrossRef]
- Zheng, H.; Dietrich, C.; Radek, R.; Brune, A. Endomicrobium proavitum, the first isolate of Endomicrobia class. nov. (phylum Elusimicrobia)—An ultramicrobacterium with an unusual cell cycle that fixes nitrogen with a Group IV nitrogenase. Environ. Microbiol. 2015, 18, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.S.; Anbar, A.D.; Miller, S.; Hamilton, T.L.; Lavin, M.; Peters, J.W. A late methanogen origin for molybdenum-dependent nitrogenase. Geobiology 2011, 9, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.S. Iron-only and vanadium nitrogenases: Fail-safe enzymes or something more? Ann. Rev. Microbiol. 2020, 74, 247–266. [Google Scholar] [CrossRef] [PubMed]
- Kimble, L.K.; Madigan, M.T. Evidence for an alternative nitrogenase in Heliobacterium gestii. FEMS Microbiol. Lett. 1992, 100, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Nomata, J.; Mizoguchi, T.; Tamiaki, H.; Fujita, Y. A second nitrogenase-like enzyme for bacteriochlorophyll biosynthesis: Reconstitution of chlorophyllide a reductase with purified X-protein (BchX) and YZ-protein (BchY-BchZ) from Rhodobacter capsulatus. J. Biol. Chem. 2006, 281, 15021–15028. [Google Scholar] [CrossRef] [Green Version]
- Muraki, N.; Nomata, J.; Ebata, K.; Mizoguchi, T.; Shiba, T.; Tamiaki, H.; Kurisu, G.; Fujita, Y. X-ray crystal structure of the light-independent protochlorophyllide reductase. Nature 2010, 465, 110–114. [Google Scholar] [CrossRef]
- Moore, S.; Sowa, S.T.; Schuchardt, C.; Deery, E.; Lawrence, A.D.; Ramos, J.V.; Billig, S.; Birkemeyer, C.; Chivers, P.; Howard, M.; et al. Elucidation of the biosynthesis of the methane catalyst coenzyme F430. Nature 2017, 543, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Zheng, K.; Ngo, P.D.; Owens, V.L.; Yang, X.-P.; Mansoorabadi, S.O. The biosynthetic pathway of coenzyme F430 in methanogenic and methanotrophic archaea. Science 2016, 354, 339–342. [Google Scholar] [CrossRef] [Green Version]
- Méheust, R.; Castelle, C.; Carnevali, P.B.M.; Farag, I.F.; He, C.; Chen, L.-X.; Amano, Y.; Hug, L.; Banfield, J.F. Groundwater Elusimicrobia are metabolically diverse compared to gut microbiome Elusimicrobia and some have a novel nitrogenase paralog. ISME J. 2020, 14, 2907–2922. [Google Scholar] [CrossRef]
- Fernandes, G.D.C.; Trarbach, L.J.; de Campos, S.B.; Beneduzi, A.; Passaglia, L.M. Alternative nitrogenase and pseudogenes: Unique features of the Paenibacillus riograndensis nitrogen fixation system. Res. Microbiol. 2014, 165, 571–580. [Google Scholar] [CrossRef]
- McRose, D.L.; Zhang, X.; Kraepiel, A.M.L.; Morel, F.M.M. Diversity and Activity of Alternative Nitrogenases in Sequenced Genomes and Coastal Environments. Front. Microbiol. 2017, 8, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahak, Y.; Schütz, M.; Bronstein, M.; Greisbeck, C.; Hauska, G.; Padan, E. Sulfide-dependent anoxygenic photosynthesis in prokaryotes—Sulfide-Quinone Reductase (SQR), the initial step. In The Phototrophic Prokaryotes; Peschek, G.A., Loffelhardt, W.L., Schmetterer, G., Eds.; Kluwer Academic/Plenum: New York, NY, USA, 1999; pp. 217–228. [Google Scholar]
- Kearns, D.; Losick, R. Swarming motility in undomesticated Bacillus subtilis. Mol. Microbiol. 2004, 49, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Genet. 2010, 8, 634–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apel, D.; Surette, M.G. Bringing order to a complex molecular machine: The assembly of the bacterial flagella. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 1851–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanna, A.; Alomari, A. Comparison 2D flagellar proteins structure in gram positive vs. gram-negative motile bacteria. EurAsian J. Biosci. 2020, 17, 6275–6281. [Google Scholar]
- Nakamura, S.; Minamino, T. Flagella-Driven Motility of Bacteria. Biomolecules 2019, 9, 279. [Google Scholar] [CrossRef] [Green Version]
- Stafford, G.P.; Hughes, C. Salmonella typhimurium flhE, a conserved flagellar regulon gene required for swarming. Microbiology 2007, 153, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Girgis, H.S.; Liu, Y.; Ryu, W.S.; Tavazoie, S. A Comprehensive Genetic Characterization of Bacterial Motility. PLoS Genet. 2007, 3, e154–e160. [Google Scholar] [CrossRef]
- Herlihey, F.A.; Moynihan, P.; Clarke, A.J. The Essential Protein for Bacterial Flagella Formation FlgJ Functions as a β-N-Acetylglucosaminidase. J. Biol. Chem. 2014, 289, 31029–31042. [Google Scholar] [CrossRef] [Green Version]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Szurmant, H.; Ordal, G.W. Diversity in Chemotaxis Mechanisms among the Bacteria and Archaea. Microbiol. Mol. Biol. Rev. 2004, 68, 301–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.V.; Young, R.; Mahenthiralingam, E.; Stickler, D.J. Ultrastructure of Proteus mirabilis Swarmer Cell Rafts and Role of Swarming in Catheter-Associated Urinary Tract Infection. Infect. Immun. 2004, 72, 3941–3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traag, B.A.; Pugliese, A.; Eisen, J.A.; Losick, R. Gene Conservation among Endospore-Forming Bacteria Reveals Additional Sporulation Genes in Bacillus subtilis. J. Bacteriol. 2013, 195, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piggot, P.J.; Hilbert, D.W. Sporulation of Bacillus subtilis. Curr. Opin. Microbiol. 2004, 7, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Mechanisms for the prevention of damage to DNA in spores of bacillus species. Annu. Rev. Microbiol. 1995, 49, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Bagyan, I.; Setlow, B.; Setlow, P. New Small, Acid-Soluble Proteins Unique to Spores of Bacillus subtilis: Identification of the Coding Genes and Regulation and Function of Two of These Genes. J. Bacteriol. 1998, 180, 6704–6712. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.O.; Moran, C.P., Jr. Structure, Assembly, and Function of the Spore Surface Layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef]
- Habte, M.; Alexander, M. Nitrogen Fixation by Photosynthetic Bacteria in Lowland Rice Culture. Appl. Environ. Microbiol. 1980, 39, 342–347. [Google Scholar] [CrossRef] [Green Version]
- Maeda, I. Potential of Phototrophic Purple Nonsulfur Bacteria to Fix Nitrogen in Rice Fields. Microorganisms 2021, 10, 28. [Google Scholar] [CrossRef]






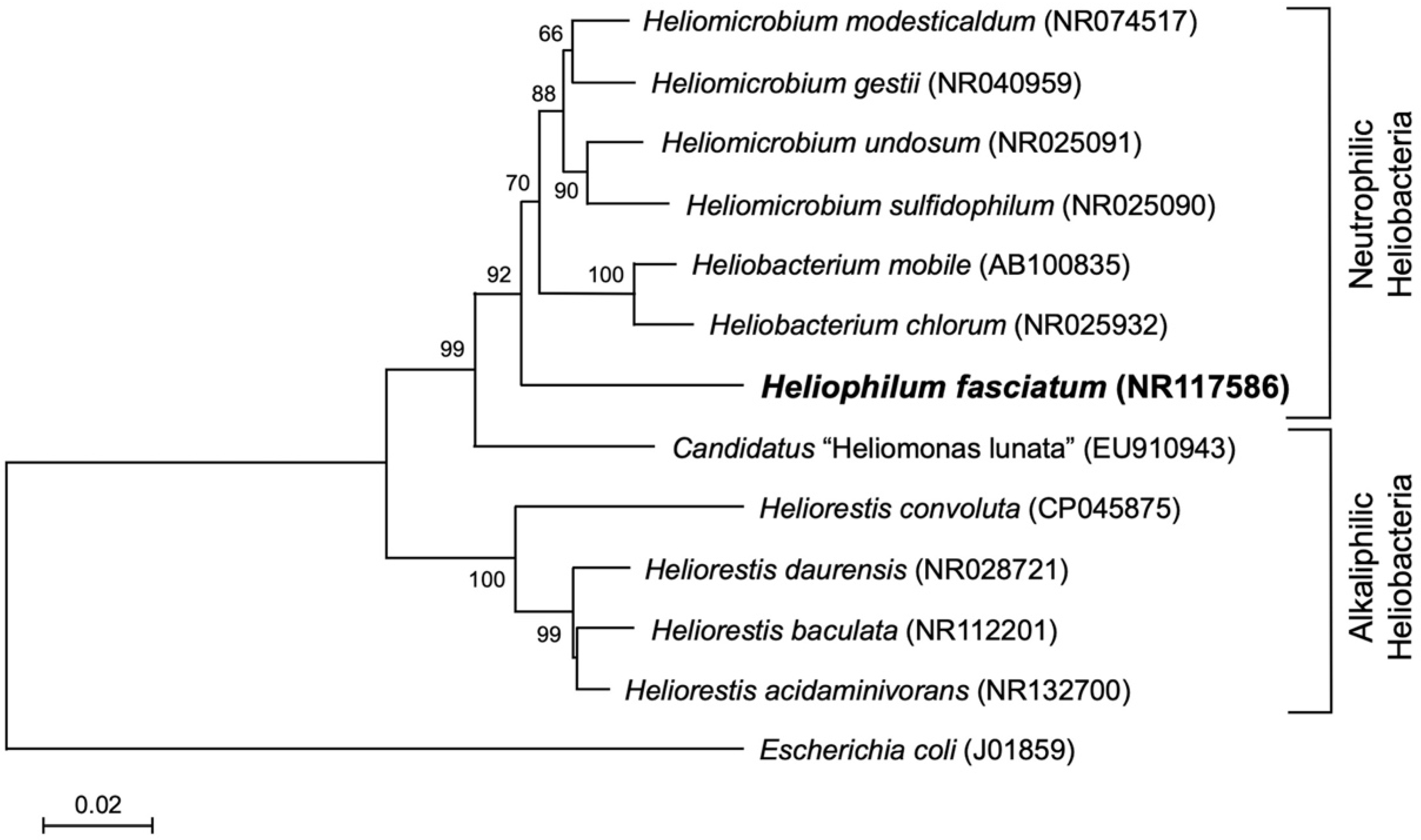{kind=link}
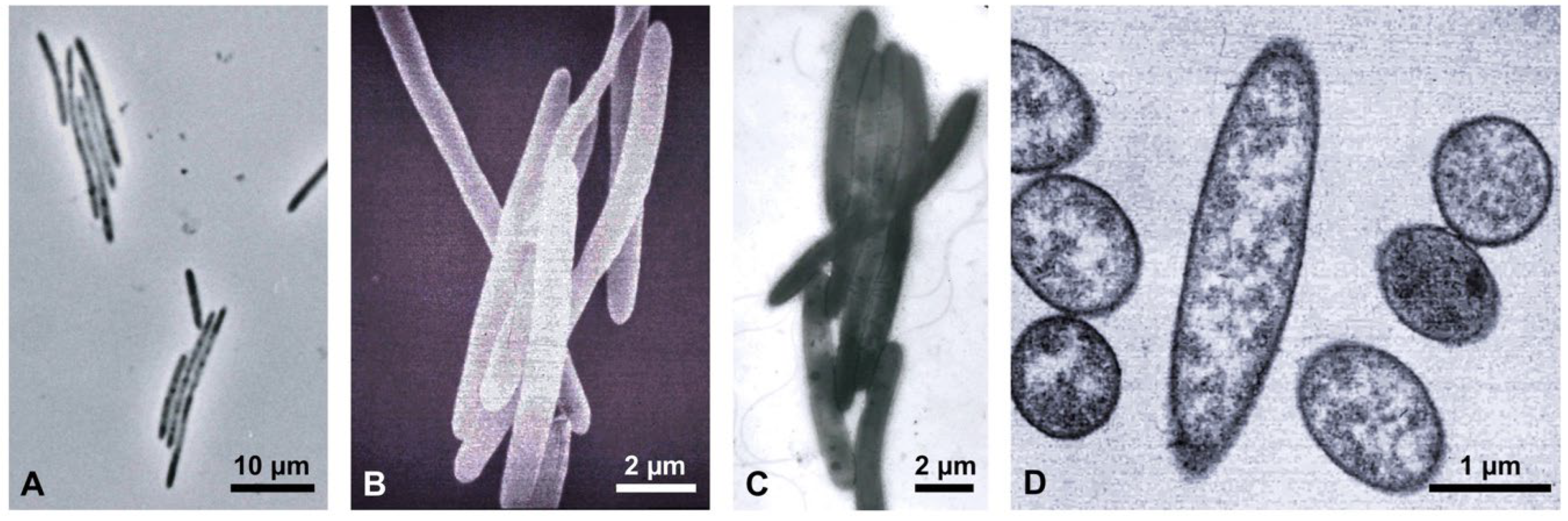{kind=link}
{kind=link}
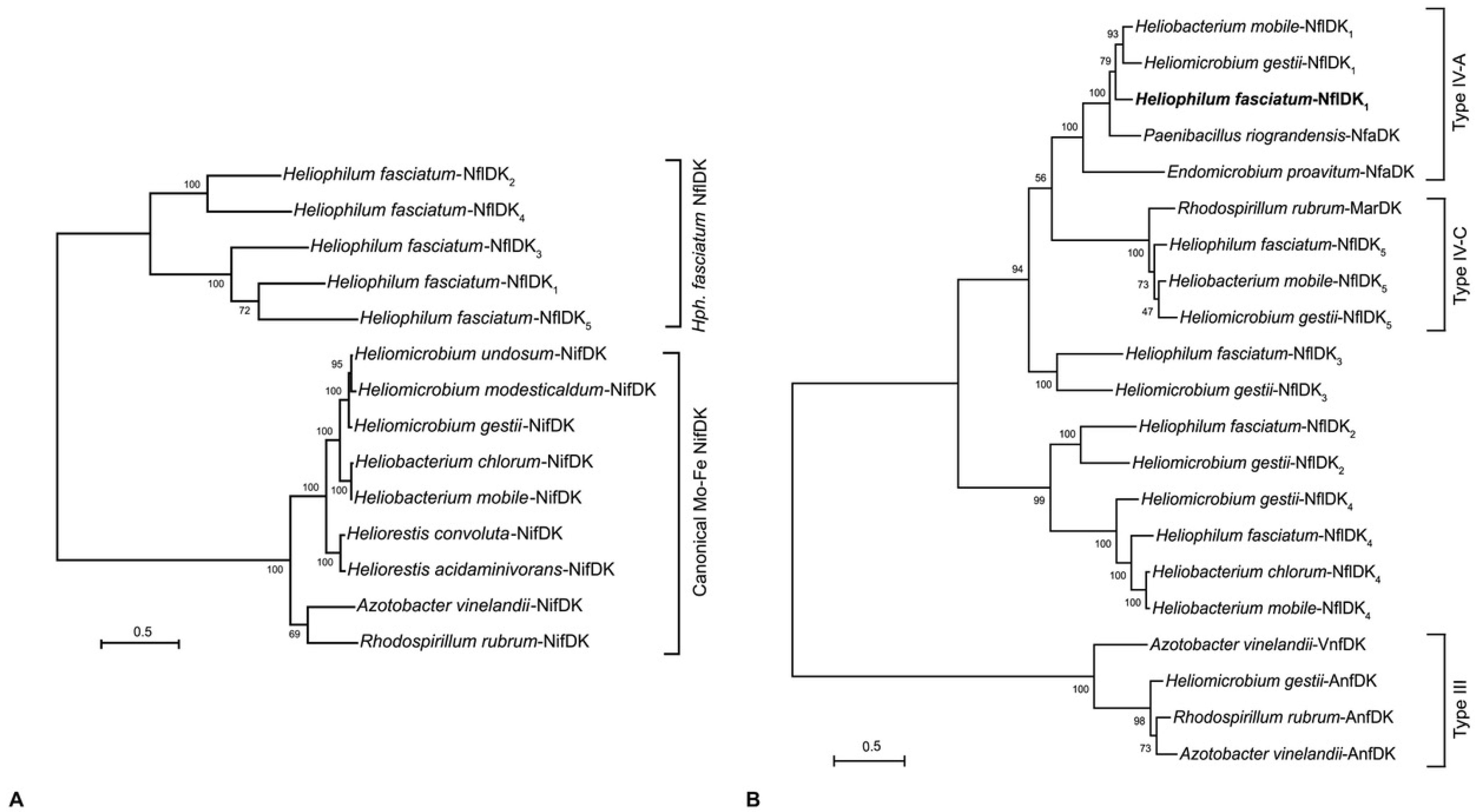{kind=link}
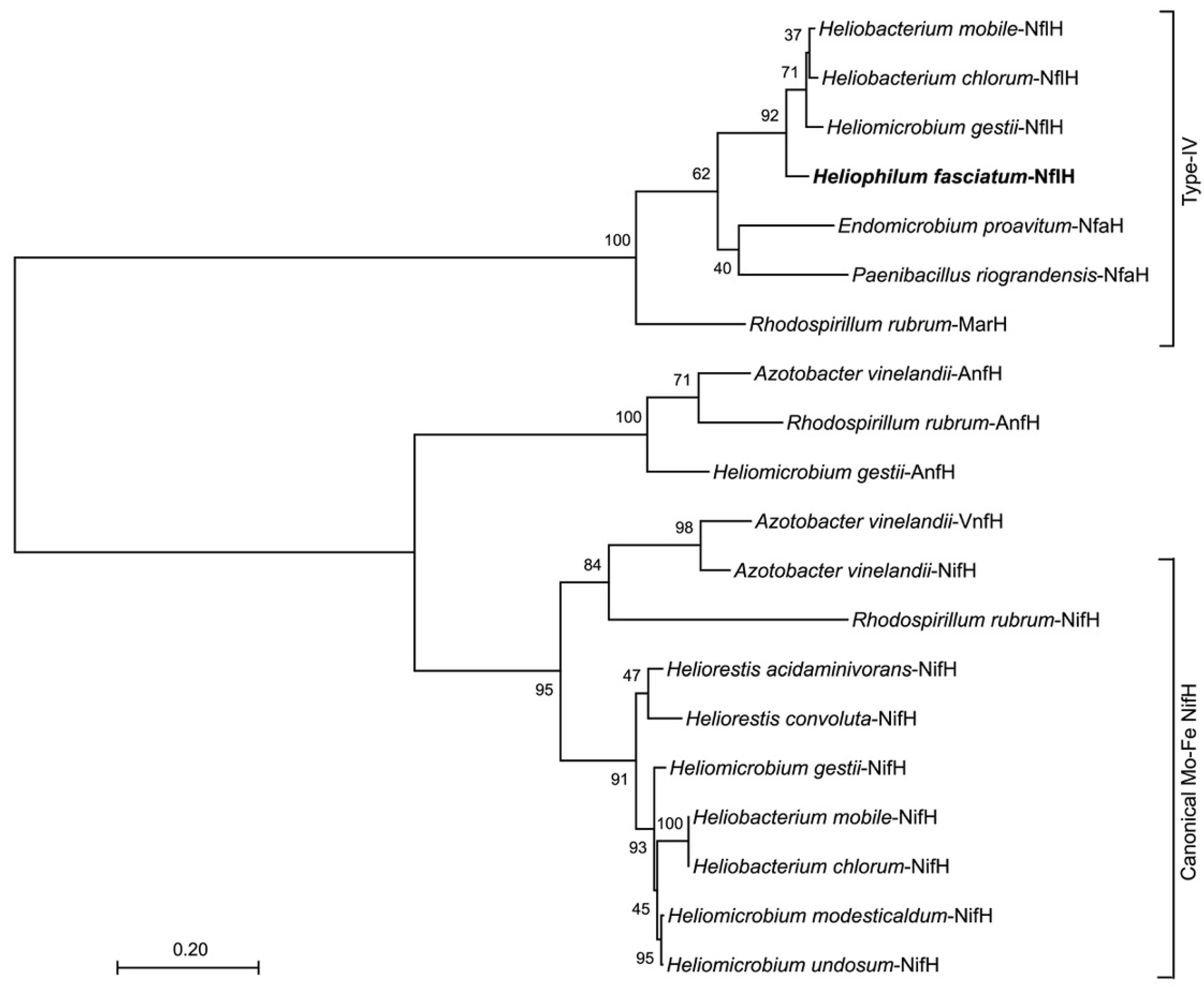{kind=link}
| Property a | TanzaniaT | HHT | Ice1T |
|---|---|---|---|
| Cell morphology | Rods in bundles | Ring-shaped coils | Rods |
| Flagellar motility | Yes | Yes | Yes |
| Absorption maxima b | 792 nm | 786 nm | 788 nm |
| Characteristic carotenoid | 4,4′-diaponeu-rosporene | OH-diaponeurosporene glucoside ester | 4,4′diaponeu-rosporene |
| Growth temperature optima (°C)/pH optima | 37/7 | 33/8.5−9 | 50−52/6.5 |
| Photoassimilation of acetate or pyruvate | Yes | Yes | Yes |
| Other C sources photoassimilated | Lactate, Butyrate c, ethanol c | Propionate, butyrate | Lactate, glucose, fructose, ribose d |
| Chemotrophic (dark, fermentative) growth | Yes | No | Yes |
| 16S rRNA gene sequence identity to TanzaniaT (%) | 100 | 90 | 93 |
| Genus/Species | Habitat/pH Optimum | Protein-Encoding Genes a |
|---|---|---|
| Heliobacterium chlorum | Garden soil/7 | 3876 (NZ_JACVHF000000000.1) |
| Heliobacterium mobile | Paddy soil, Thailand/7 | 3699 (NZ_WNKU00000000.1) |
| Heliomicrobium modesticaldum | Hot springs, Reykjanes, Iceland/6-7 b | 2662 (NC_010337.2) |
| Heliomicrobium gestii | Paddy soil, Dar es Salaam, Tanzania/7 | 3266 (NZ_WXEX00000000.1) |
| Heliomicrobium undosum | Microbial mat, Garga Hot Springs, Siberia/7.5 | 3356 (NZ_WXEY00000000.1) |
| Heliorestis acidaminivorans | Lake Hamra, Wadi Natroun Egypt/9 | 2765 (NZ_WBXO00000000.1) |
| Heliorestis convoluta | Lake Hamra, Wadi, Natroun Egypt/8.5 | 2909 (NZ_CP045875.1) |
| Heliophilum fasciatum | Paddy soil, Dar es Salaam, Tanzania/7 | c 2834 (NZ_SLXT00000000.1)/2951 |
| Characteristic | TanzaniaT | HHT | Ice1T |
|---|---|---|---|
| Accession number | Ga043916 b | CP045875 b | CP000930 b |
| Genome size (bp) c | 3,141,306 | 3,218,981 | 3,075,407 |
| Contigs | 75 | 1 | 1 |
| Genome G+C (%) | 50.9 | 43.1 | 56 |
| Coding DNA (%) | 95.7 | 90.1 | 90.6 |
| Total ORFs | 2951 | 3263 | 3138 |
| Hypothetical proteins (%) | 22.9 | 27.5 | 23.8 |
| rRNAs | 15 | 9 | 24 |
| tRNAs | 89 | 105 | 104 |
| Average nucleotide identity (ANI) | |||
| to genome of TanzaniaT (%) d | 100 | 66.7 | 68.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bender, K.S.; Madigan, M.T.; Williamson, K.L.; Mayer, M.H.; Parenteau, M.N.; Jahnke, L.L.; Welander, P.V.; Sanguedolce, S.A.; Brown, A.C.; Sattley, W.M. Genomic Features of the Bundle-Forming Heliobacterium Heliophilum fasciatum. Microorganisms 2022, 10, 869. https://doi.org/10.3390/microorganisms10050869
Bender KS, Madigan MT, Williamson KL, Mayer MH, Parenteau MN, Jahnke LL, Welander PV, Sanguedolce SA, Brown AC, Sattley WM. Genomic Features of the Bundle-Forming Heliobacterium Heliophilum fasciatum. Microorganisms. 2022; 10(5):869. https://doi.org/10.3390/microorganisms10050869
Chicago/Turabian StyleBender, Kelly S., Michael T. Madigan, Kyleigh L. Williamson, Marisa H. Mayer, Mary N. Parenteau, Linda L. Jahnke, Paula V. Welander, Sophia A. Sanguedolce, Abigail C. Brown, and W. Matthew Sattley. 2022. "Genomic Features of the Bundle-Forming Heliobacterium Heliophilum fasciatum" Microorganisms 10, no. 5: 869. https://doi.org/10.3390/microorganisms10050869