
Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. The Ocean Habitat and Microbial Diversity: A New Leaf in Drug Discovery
3. Actinomycetes: A Teeming Wealth of Chemical and Biological Warfare
4. Marine Actinomycetes: A Potential Frontier of Bioactive Compounds
5. Genomic Insight of Marine Actinomycetes
6. Cultivation Techniques of Marine Actinomycetes
7. Fermentation and Extraction of Bioactive Compounds from Actinomycetes
8. The Biology of Coral
8.1. Coral Morphology and Distribution
8.2. Coral Significance and Bleaching
8.3. Coral Defense Mechanism
8.4. Coral Symbiosis
9. The Order Scleractinia
10. Association between Scleractinian and Actinomycetes
11. Novel Species and Compounds of Actinobacteria Recovered from Coral (Order Scleractinia)
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Subramani, R.; Aalbersberg, W. Marine actinomycetes: An ongoing source of novel bioactive metabolites. Microbiol. Res. 2012, 167, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; Salamat, M.K.F.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. Available online: http://www.ncbi.nlm.nih.gov/pubmed/30349322 (accessed on 19 January 2019). [CrossRef] [PubMed] [Green Version]
- Abdallah, E. Plants: An alternative source for antimicrobials. J. Appl. Pharm. Sci. 2011, 1, 16–20. [Google Scholar]
- Foxlee, N.D.; Townell, N.; McIver, L.; Lau, C.L. Antibiotic Resistance in Pacific Island Countries and Territories: A Systematic Scoping Review. Antibiotics 2019, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Khan, S.N.; Khan, A.U. Breaking the Spell: Combating Multidrug Resistant ‘Superbugs’. Front. Microbiol. 2016, 7, 174. [Google Scholar] [CrossRef]
- Carvalho, I.T.; Santos, L. Antibiotics in the aquatic environments: A review of the European scenario. Environ. Int. 2016, 94, 736–757. [Google Scholar] [CrossRef]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef]
- Choudhary, A.; Naughton, L.M.; Montánchez, I.; Dobson, A.D.W.; Rai, D.K. Current Status and Future Prospects of Marine Natural Products (MNPs) as Antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Sogin, M.L.; Morrison, H.G.; Huber, J.A.; Welch, D.M.; Huse, S.M.; Neal, P.R.; Arrieta, J.M.; Herndl, G.J. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. USA 2006, 103, 12115–12120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. Prokaryote diversity and taxonomy: Current status and future challenges. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 623–638. [Google Scholar] [CrossRef] [PubMed]
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The unseen majority. Proc. Natl. Acad. Sci. USA 1998, 95, 6578–6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial metabolites in nutrition, healthcare and agriculture. 3 Biotech 2017, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Hu, Y.; Odermatt, P.D.; Gonzalez, C.G.; Zhang, L.; Elias, J.E.; Chang, F.; Huang, K.C. Precise regulation of the relative rates of surface area and volume synthesis in bacterial cells growing in dynamic environments. Nat. Commun. 2021, 12, 1975. [Google Scholar] [CrossRef] [PubMed]
- Zarraonaindia, I.; Smith, D.P.; Gilbert, J.A. Beyond the genome: Community-level analysis of the microbial world. Biol. Philos. 2012, 28, 261–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.K.; Macdonald, C. Drug discovery from uncultivable microorganisms. Drug Discov. Today 2010, 15, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Sanchez, S. Microbial drug discovery: 80 years of progress. J. Antibiot. 2009, 62, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenical, W.; Jensen, P. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Gasol, J.M.; McDonough, N.; Calewaert, J.B. Marine Microbial Diversity and Its Role in Eco-System Functioning and Environmental Change; Calewaert, J.B., McDonough, N., Eds.; Marine Board European Scientific Foundation: Ostend, Belgium, 2012; pp. 1–80. [Google Scholar] [CrossRef]
- Jagannathan, S.; Manemann, E.; Rowe, S.; Callender, M.; Soto, W. Marine Actinomycetes, New Sources of Biotechnological Products. Mar. Drugs 2021, 19, 365. [Google Scholar] [CrossRef]
- Mahmoud, H.M.; Kalendar, A.A. Coral-Associated Actinobacteria: Diversity, Abundance, and Biotechnological Potentials. Front. Microbiol. 2016, 7, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, S.; Mathivanan, N. Screening of marine actinomycetes isolated from the Bay of Bengal, India for antimicrobial activity and industrial enzymes. World J. Microbiol. Biotechnol. 2009, 25, 2103–2111. [Google Scholar] [CrossRef]
- Subramani, R.; Aalbersberg, W. Culturable rare Actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donia, M.; Hamann, M.T. Marine natural products and their potential applications as anti-infective agents. Lancet Infect. Dis. 2003, 3, 338–348. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the Past and Charting the Future of Marine Natural Products Drug Discovery and Chemical Biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortorella, E.; Tedesco, P.; Esposito, F.P.; January, G.G.; Fani, R.; Jaspars, M.; de Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, C.C.C.R.; Fernandes, P. Production of Metabolites as Bacterial Responses to the Marine Environment. Mar. Drugs 2010, 8, 705–727. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive Compounds from Marine Bacteria and Fungi. Microb. Biotechnol. 2010, 3, 544–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, C.-L.; Xia, J.-M.; Wang, J.-S.; Lin, D.-H.; Yang, X.-W. Metabolomic Investigations on Nesterenkonia flava Revealed Significant Differences between Marine and Terrestrial Actinomycetes. Mar. Drugs 2018, 16, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigwart, J.D.; Blasiak, R.; Jaspars, M.; Jouffray, J.-B.; Tasdemir, D. Unlocking the potential of marine biodiscovery. Nat. Prod. Rep. 2020, 38, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. 2016, 96, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Rotter, A.; Barbier, M.; Bertoni, F.; Bones, A.M.; Cancela, M.L.; Carlsson, J.; Carvalho, M.F.; Cegłowska, M.; Chirivella-Martorell, J.; Dalay, M.C.; et al. The Essentials of Marine Biotechnology. Front. Mar. Sci. 2021, 8, 158. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Anandan, R.; Dharumadurai, D.; Manogaran, G.P. Anandan, R.; Dharumadurai, D.; Manogaran, G.P. An introduction to actinobacteria. In Actinobacteria-Basics and Biotechnological Applications; Dhanasekaran., D., Jiang, Y., Eds.; IntechOpen: London, UK, 2016; pp. 3–37. [Google Scholar] [CrossRef] [Green Version]
- Jüttner, F.; Watson, S.B. Biochemical and Ecological Control of Geosmin and 2-Methylisoborneol in Source Waters. Appl. Environ. Microbiol. 2007, 73, 4395–4406. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Cross, T. Actinomycetes. In Biology of Plant Litter Decomposition; Dickinson, C.H., Pugh, G.J.F., Eds.; Academic Press: London, UK, 2012; pp. 269–302. [Google Scholar]
- De Simeis, D.; Serra, S. Actinomycetes: A Never-Ending Source of Bioactive Compounds—An Overview on Antibiotics Production. Antibiotics 2021, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Grasso, L.L.; Martino, D.C.; Alduina, R. Production of Antibacterial Compounds from Actinomycetes. Actinobacteria-Basics Biotechnol. Appl. 2016, 214, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Mak, S.; Xu, Y.; Nodwell, J.R. The expression of antibiotic resistance genes in antibiotic-producing bacteria. Mol. Microbiol. 2014, 93, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dangi, P.; Choudhary, M. Actinomycetes: Source, identification, and their applications. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 801–832. [Google Scholar]
- Subramani, R.; Sipkema, D. Marine Rare Actinomycetes: A Promising Source of Structurally Diverse and Unique Novel Natural Products. Mar. Drugs 2019, 17, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.-K. RETRACTED: Marine actinobacterial metabolites: Current status and future perspectives. Microbiol. Res. 2013, 168, 311–332. [Google Scholar] [CrossRef]
- Baskaran, R.; Subramanian, T.; Zuo, W.; Qian, J.; Wu, G.; Kumar, A. Major source of marine actinobacteria and its biomedical application. In Microbial Applications; Kalia, C.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 2, pp. 55–82. [Google Scholar]
- Janardhan, A.; Kumar, A.P.; Viswanath, B.; Saigopal, D.V.R.; Narasimha, G. Production of Bioactive Compounds by Actinomycetes and Their Antioxidant Properties. Biotechnol. Res. Int. 2014, 2014, 217030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary metabolites and biodiversity of actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine Bacterial Secondary Metabolites: A Treasure House for Structurally Unique and Effective Antimicrobial Compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Biswas, K.; Soalnki, V.; Kumar, P.; Tarafdar, A. Actinomycetes: Potential bioresource for human welfare: A review. Res. J. Chem. Environ. Sci. 2014, 2, 5–16. [Google Scholar]
- Zhang, H.; Wang, Y.; Pfeifer, B.A. Bacterial Hosts for Natural Product Production. Mol. Pharm. 2008, 5, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Helmke, E.; Weyland, H. Rhodococcus marinonascens sp. nov., an Actinomycete from the Sea. Int. J. Syst. Bacteriol. 1984, 34, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Haynes, J.A. Actinomycetes in marine sediments. Biol. Biochem. Biomed. Asp. Actinomycetes 1984, 453–472. [Google Scholar] [CrossRef]
- Jensen, P.R.; Dwight, R.; Fenical, W. Distribution of actinomycetes in near-shore tropical marine sediments. Appl. Environ. Microbiol. 1991, 57, 1102–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, M.A.; Rutherford, L.T.; Hodson, R.E. Evidence for indigenous Streptomyces populations in a marine environment determined with a 16S rRNA probe. Appl. Environ. Microbiol. 1995, 61, 3695–3700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mincer, T.J.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Widespread and Persistent Populations of a Major New Marine Actinomycete Taxon in Ocean Sediments. Appl. Environ. Microbiol. 2002, 68, 5005–5011. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, L.A.; Fenical, W.; Jensen, P.; Kauffman, C.; Mincer, T.; Ward, A.; Bull, A.T.; Goodfellow, M. Salinispora arenicola gen. nov., sp. nov. and Salinispora tropica sp. nov., obligate marine actinomycetes belonging to the family Micromonosporaceae. Int. J. Syst. Evol. Microbiol. 2005, 55, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; El-Wahed, A.A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [Green Version]
- Voser, T.M.; Campbell, M.D.; Carroll, A.R. How different are marine microbial natural products compared to their terrestrial counterparts? Nat. Prod. Rep. 2021, 39, 7–19. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural Product Drug Discovery in the Next Millennium. Pharm. Biol. 2001, 39, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Rathore, D.S.; Sharma, A.K.; Dobariya, A.; Ramavat, H.; Singh, S.P. Cultivation and diversity of marine actinomycetes: Molecu-lar approaches and bioinformatics tools. In Actinobacteria: Microbiology to Synthetic Biology; Karthik, L., Ed.; Springer: Singapore, 2022; pp. 215–240. [Google Scholar]
- Palazzotto, E.; Weber, T. Omics and multi-omics approaches to study the biosynthesis of secondary metabolites in microorganisms. Curr. Opin. Microbiol. 2018, 45, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.-K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Alanjary, M.; Weber, T. The evolution of genome mining in microbes—A review. Nat. Prod. Rep. 2016, 33, 988–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Undabarrena, A.; Ugalde, J.A.; Seeger, M.; Cámara, B. Genomic data mining of the marine actinobacteriaStreptomycessp. H-KF8 unveils insights into multi-stress related genes and metabolic pathways involved in antimicrobial synthesis. PeerJ 2017, 5, e2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letzel, A.-C.; Li, J.; Amos, G.C.A.; Millán-Aguiñaga, N.; Ginigini, J.; Abdelmohsen, U.R.; Gaudêncio, S.P.; Ziemert, N.; Moore, B.S.; Jensen, P.R. Genomic insights into specialized metabolism in the marine actinomyceteSalinispora. Environ. Microbiol. 2017, 19, 3660–3673. [Google Scholar] [CrossRef]
- Xu, L.; Ye, K.-X.; Dai, W.-H.; Sun, C.; Xu, L.-H.; Han, B.-N. Comparative Genomic Insights into Secondary Metabolism Biosynthetic Gene Cluster Distributions of Marine Streptomyces. Mar. Drugs 2019, 17, 498. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Lee, N.; Jeong, Y.; Lee, Y.; Kim, W.; Cho, S.; Palsson, B.O.; Cho, B.-K. Primary transcriptome and translatome analysis determines transcriptional and translational regulatory elements encoded in the Streptomyces clavuligerus genome. Nucleic Acids Res. 2019, 47, 6114–6129. [Google Scholar] [CrossRef] [Green Version]
- Chu, L.; Huang, J.; Muhammad, M.; Deng, Z.; Gao, J. Genome mining as a biotechnological tool for the discovery of novel marine natural products. Crit. Rev. Biotechnol. 2020, 40, 571–589. [Google Scholar] [CrossRef]
- Mu, D.-S.; Ouyang, Y.; Chen, G.-J.; Du, Z.-J. Strategies for culturing active/dormant marine microbes. Mar. Life Sci. Technol. 2020, 3, 121–131. [Google Scholar] [CrossRef]
- Stewart, E.J. Growing Unculturable Bacteria. J. Bacteriol. 2012, 194, 4151–4160. [Google Scholar] [CrossRef] [Green Version]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zengler, K.; Toledo, G.; Rappé, M.; Elkins, J.; Mathur, E.J.; Short, J.M.; Keller, M. Cultivating the uncultured. Proc. Natl. Acad. Sci. USA 2002, 99, 15681–15686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Li, Q.; Chen, X.; Jiang, C. Isolation and cultivation methods of actinobacteria. In Actinobacteria-Basics and Biotechnological Applications; Dhanasekaran, D., Jiang, Y., Eds.; InTechOpen: London, UK, 2016; pp. 39–50. [Google Scholar]
- Mohamed, S.S.; Abdelhamid, S.A.; Ali, R.H. Isolation and identification of marine microbial products. J. Genet. Eng. Biotechnol. 2021, 19, 162. [Google Scholar] [CrossRef] [PubMed]
- Davies-Bolorunduro, O.; Osuolale, O.; Saibu, S.; Adeleye, I.; Aminah, N. Bioprospecting marine actinomycetes for antileishmanial drugs: Current perspectives and future prospects. Heliyon 2021, 7, e07710. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Diversity and isolation of rare actinomycetes: An overview. Crit. Rev. Microbiol. 2012, 39, 256–294. [Google Scholar] [CrossRef] [PubMed]
- Jose, P.A.; Jha, B. Intertidal marine sediment harbours Actinobacteria with promising bioactive and biosynthetic potential. Sci. Rep. 2017, 7, 10041. [Google Scholar] [CrossRef] [PubMed]
- Claverías, F.P.; Undabarrena, A.N.; Egonzález, M.; Eseeger, M.; Cámara, B.P. Culturable diversity and antimicrobial activity of Actinobacteria from marine sediments in Valparaíso bay, Chile. Front. Microbiol. 2015, 6, 737. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.-H.; Lee, L.-H.; Goh, B.-H. Critical review of fermentation and extraction of anti-Vibrio compounds from Streptomyces. Prog. Microbes Mol. Biol. 2020, 3, 1–14. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, X.; An, F.; Wang, G.; Zhang, X. Improvement of antibiotic activity of Xenorhabdus bovienii by medium optimization using response surface methodology. Microb. Cell Factories 2011, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, D.H.; Abdallah, N.A.; Abolmaaty, A.; Tolba, S.; Wellington, E.M.H. Microbiological and molecular insights on rare Actinobacteria harboring bioactive prospective. Bull. Natl. Res. Cent. 2020, 44, 5. [Google Scholar] [CrossRef]
- Feng, R.; Chen, L.; Chen, K. Fermentation trip: Amazing microbes, amazing metabolisms. Ann. Microbiol. 2018, 68, 717–729. [Google Scholar] [CrossRef]
- Kumar, V.; Ahluwalia, V.; Saran, S.; Kumar, J.; Patel, A.K.; Singhania, R.R. Recent developments on solid-state fermentation for production of microbial secondary metabolites: Challenges and solutions. Bioresour. Technol. 2020, 323, 124566. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A. Solid-state fermentation. Biochem. Eng. J. 2003, 13, 81–84. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, M.; Ramteke, P.W.; Mishra, P.K. Solid-state fermentation strategy for microbial metabolites pro-duction: An overview. In New And Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–354. [Google Scholar] [CrossRef]
- Subramaniyam, R.; Vimala, R. Solid state and submerged fermentation for the production of bioactive substances: A comparative study. Int. J. Sci. Nat. 2012, 3, 480–486. [Google Scholar]
- Balagurunathan, R.; Radhakrishnan, M.; Shanmugasundaram, T.; Gopikrishnan, V.; Jerrine, J. Production of bioproducts from actinobacteria. In Protocols in Actinobacterial Research; Springer: Berlin/Heidelberg, Germany, 2020; pp. 113–128. [Google Scholar]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Rasul, M.G. Extraction, isolation and characterization of natural products from medicinal plants. Int. J. Basic Sci. Appl. Comput. 2018, 2, 1–6. [Google Scholar]
- Balagurunathan, R.; Radhakrishnan, M.; Shanmugasundaram, T.; Gopikrishnan, V.; Jerrine, J. Bioassay-guided isolation and characterization of metabolites from actinobacteria. In Protocols in Actinobacterial Research; Springer: Berlin/Heidelberg, Germany, 2020; pp. 147–163. [Google Scholar]
- Buddemeier, R.W.; Kleypas, J.; Aronson, R. Coral Reefs and Global Climate Change: Potential Contributions of Climate Change to Stresses on Coral Reef Ecosystems; Pew Center on Global Climate Change: Arlington, VA, USA, 2004. [Google Scholar]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral reef biodiversity. In Life in the World’s Oceans: Diversity Distribution and Abundance; Mclntyre, A.D., Ed.; Wiley-Blackwell: Singapore, 2010; pp. 65–74. [Google Scholar]
- Spalding, M.D.; Ravilious, C.; Green, E.P. World Atlas of Coral Reefs; University of California Press: Berkeley, CA, USA, 2001; pp. 9–404. [Google Scholar]
- Barott, K.L.; Venn, A.A.; Perez, S.O.; Tambutté, S.; Tresguerres, M. Coral host cells acidify symbiotic algal microenvironment to promote photosynthesis. Proc. Natl. Acad. Sci. USA 2014, 112, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Feely, R.; Sabine, C.; Takahashi, T.; Wanninkhof, R. Uptake and Storage of Carbon Dioxide in the Ocean: The Global CO2 Survey. Oceanography 2001, 14, 18–32. [Google Scholar] [CrossRef]
- Kleypas, J.; Feely, R.; Fabry, V.J.; Langdon, C.; Christopher, S.; Robbins, L.L. Impacts of Ocean Acidification on Coral Reefs and Other Marine Calcifiers: A Guide for Future Research; Report of a Workshop Sponsored; National Science Foundation, National Oceanic and Atmospheric Administration, and United States Geological Survey: St. Petersburg, FL, USA, 2006. [Google Scholar]
- Souter, D.W.; Lindén, O. The health and future of coral reef systems. Ocean Coast. Manag. 2000, 43, 657–688. [Google Scholar] [CrossRef]
- Dutra, L.; Haywood, M.D.E.; Singh, S.S.; Piovano, S.; Ferreira, M.; Johnson, J.E.; Morris, C.W. Effects of climate change on corals relevant to the Pacific Islands. In Pacific Marine Climate Change Report Card; Townhill, B., Buckley, P., Eds.; Commonwealth Marine Economics Programme: London, UK, 2018; pp. 132–158. [Google Scholar]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschel, U.; Hirt, H.; Hülter, N.F.; Lachnit, T.; et al. Metaorganisms in extreme environments: Do microbes play a role in organismal adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Douglas, A. Coral bleaching—How and why? Mar. Pollut. Bull. 2003, 46, 385–392. [Google Scholar] [CrossRef]
- Mendes, J.M.; Woodley, J.D. Effect of the 1995-1996 bleaching event on polyp tissue depth, growth, reproduction and skeletal band formation in Montastraea annularis. Mar. Ecol. Prog. Ser. 2002, 235, 93–102. [Google Scholar] [CrossRef]
- Spalding, M.D.; Brown, B.E. Warm-water coral reefs and climate change. Science 2015, 350, 769–771. [Google Scholar] [CrossRef] [PubMed]
- Hume, B.; D’Angelo, C.; Burt, J.; Baker, A.; Riegl, B.; Wiedenmann, J. Corals from the Persian/Arabian Gulf as models for thermotolerant reef-builders: Prevalence of clade C3 Symbiodinium, host fluorescence and ex situ temperature tolerance. Mar. Pollut. Bull. 2013, 72, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.E.; Bythell, J.C. Perspectives on mucus secretion in reef corals. Mar. Ecol. Prog. Ser. 2005, 296, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Bythell, J.C.; Wild, C. Biology and ecology of coral mucus release. J. Exp. Mar. Biol. Ecol. 2011, 408, 88–93. [Google Scholar] [CrossRef]
- Huettel, M.; Wild, C.; Gonelli, S. Mucus trap in coral reefs: Formation and temporal evolution of particle aggregates caused by coral mucus. Mar. Ecol. Prog. Ser. 2006, 307, 69–84. [Google Scholar] [CrossRef]
- Ritchie, K. Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar. Ecol. Prog. Ser. 2006, 322, 1–14. [Google Scholar] [CrossRef]
- Shnit-Orland, M.; Kushmaro, A. Coral mucus-associated bacteria: A possible first line of defense. FEMS Microbiol. Ecol. 2009, 67, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, J.; Kannapiran, E.; Manikandan, B.; Francis, K.; Arora, S.; Karunya, E.; Kumar, A.; Singh, S.K.; Jose, J. UV-absorbing bacteria in coral mucus and their response to simulated temperature elevations. Coral Reefs 2013, 32, 1043–1050. [Google Scholar] [CrossRef]
- Palmer, C.V.; Traylor-Knowles, N.G.; Willis, B.L.; Bythell, J.C. Corals Use Similar Immune Cells and Wound-Healing Processes as Those of Higher Organisms. PLoS ONE 2011, 6, e23992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, K.M.; Peters, E.C.; Harvell, C.D. Coral resistance to disease. In Coral Health and Disease; Rosenberg, E., Loya, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 377–399. [Google Scholar]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Koren, O.; Reshef, L.; Efrony, R.; Zilber-Rosenberg, I. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Genet. 2007, 5, 355–362. [Google Scholar] [CrossRef]
- Webster, N.S.; Reusch, T.B.H. Microbial contributions to the persistence of coral reefs. ISME J. 2017, 11, 2167–2174. [Google Scholar] [CrossRef]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the Core Microbiome in Corals’ Microbial Soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Marchioro, G.M.; Glasl, B.; Engelen, A.H.; Serrão, E.A.; Bourne, D.G.; Webster, N.S.; Frade, P.R. Microbiome dynamics in the tissue and mucus of acroporid corals differ in relation to host and environmental parameters. PeerJ 2020, 8, e9644. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Iida, Y.; Uthicke, S.; Smith-Keune, C. Changes in coral-associated microbial communities during a bleaching event. ISME J. 2007, 2, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Glasl, B.; Herndl, G.J.; Frade, P.R. The microbiome of coral surface mucus has a key role in mediating holobiont health and survival upon disturbance. ISME J. 2016, 10, 2280–2292. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.C. Flexibility and Specificity in Coral-Algal Symbiosis: Diversity, Ecology, and Biogeography of Symbiodinium. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 661–689. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O. Coral reefs in a century of rapid environmental change. Symbiosis 2004, 37, 1–31. [Google Scholar]
- Padilla-Gamiño, J.L.; Pochon, X.; Bird, C.; Concepcion, G.T.; Gates, R.D. From Parent to Gamete: Vertical Transmission of Symbiodinium (Dinophyceae) ITS2 Sequence Assemblages in the Reef Building Coral Montipora capitata. PLoS ONE 2012, 7, e38440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumbo, V.R.; Baird, A.H.; van Oppen, M. The promiscuous larvae: Flexibility in the establishment of symbiosis in corals. Coral Reefs 2012, 32, 111–120. [Google Scholar] [CrossRef]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell Biology of Cnidarian-Dinoflagellate Symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 229–261. [Google Scholar] [CrossRef] [Green Version]
- Stat, M.; Loh, W.K.W.; Hoegh-Guldberg, O.; Carter, D.A. Symbiont acquisition strategy drives host–symbiont associations in the southern Great Barrier Reef. Coral Reefs 2008, 27, 763–772. [Google Scholar] [CrossRef] [Green Version]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pochon, X.; Montoya-Burgos, J.I.; Stadelmann, B.; Pawlowski, J. Molecular phylogeny, evolutionary rates, and divergence timing of the symbiotic dinoflagellate genus Symbiodinium. Mol. Phylogenetics Evol. 2006, 38, 20–30. [Google Scholar] [CrossRef]
- Blackall, L.L.; Wilson, B.; van Oppen, M. Coral-the world’s most diverse symbiotic ecosystem. Mol. Ecol. 2015, 24, 5330–5347. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, K.E.; Lichtenberg, M.; Ralph, P.; Kühl, M.; Wangpraseurt, D. Radiative energy budget reveals high photosynthetic efficiency in symbiont-bearing corals. J. R. Soc. Interface 2014, 11, 20130997. [Google Scholar] [CrossRef]
- Alieva, N.O.; Konzen, K.A.; Field, S.F.; Meleshkevitch, E.A.; Hunt, M.E.; Beltran-Ramirez, V.; Miller, D.J.; Wiedenmann, J.; Salih, A.; Matz, M.V. Diversity and Evolution of Coral Fluorescent Proteins. PLoS ONE 2008, 3, e2680. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, C.R.C.; Davy, S.K.; Pilling, G.M.; Graham, N.A.J. The Biology of Coral Reefs; Oxford University Press: Oxford, UK, 2018; pp. 1–341. [Google Scholar]
- Burriesci, M.S.; Raab, T.K.; Pringle, J.R. Evidence that glucose is the major transferred metabolite in dinoflagellate–cnidarian symbiosis. J. Exp. Biol. 2012, 215, 3467–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellowlees, D.; Rees, T.A.V.; Leggat, W. Metabolic interactions between algal symbionts and invertebrate hosts. Plant Cell Environ. 2008, 31, 679–694. [Google Scholar] [CrossRef] [PubMed]
- Kayal, E.; Roure, B.; Philippe, H.; Collins, A.G.; Lavrov, D.V. Cnidarian phylogenetic relationships as revealed by mitogenomics. BMC Evol. Biol. 2013, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitahara, M.V.; Fukami, H.; Benzoni, F.; Huang, D. The new systematics of Scleractinia: Integrating molecular and morphological evidence. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 41–59. [Google Scholar]
- Stanley, G.D. Photosymbiosis and the Evolution of Modern Coral Reefs. Science 2006, 312, 857–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, G.D.; Fautin, D.G. The Origins of Modern Corals. Science 2001, 291, 1913–1914. [Google Scholar] [CrossRef]
- Sutherland, K.P.; Porter, J.; Torres, C. Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals. Mar. Ecol. Prog. Ser. 2004, 266, 273–302. [Google Scholar] [CrossRef] [Green Version]
- Varijakzhan, D.; Loh, J.-Y.; Yap, W.-S.; Yusoff, K.; Seboussi, R.; Lim, S.-H.; Lai, K.-S.; Chong, C.-M. Bioactive Compounds from Marine Sponges: Fundamentals and Applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Ward, A.C.; Bora, N. Diversity and biogeography of marine actinobacteria. Curr. Opin. Microbiol. 2006, 9, 279–286. [Google Scholar] [CrossRef]
- Raimundo, I.; Silva, S.G.; Costa, R.; Keller-Costa, T. Bioactive Secondary Metabolites from Octocoral-Associated Microbes—New Chances for Blue Growth. Mar. Drugs 2018, 16, 485. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Li, P.; Tang, X. Natural products from corals. In Symbiotic Microbiomes of Coral Reefs Sponges and Corals; Li, Z., Ed.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 465–505. [Google Scholar]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionísio, G.; Rocha, R.J.M.; Silva, B.; Rosa, R.; Calado, R. Marine Microorganism-Invertebrate Assemblages: Perspectives to Solve the “Supply Problem” in the Initial Steps of Drug Discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef] [Green Version]
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.-M.; Xu, R.-F.; Gu, Y.-C.; Wang, C.-Y.; Shao, C.-L. Biological and Chemical Diversity of Coral-Derived Microorganisms. Curr. Med. Chem. 2015, 22, 3707–3762. [Google Scholar] [CrossRef] [PubMed]
- Sang, V.T.; Dat, T.T.H.; Vinh, L.B.; Cuong, L.C.V.; Oanh, P.T.T.; Ha, H.; Kim, Y.H.; Anh, H.L.T.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific source of Potential Bioactive Natural Products. Mar. Drugs 2019, 17, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainsworth, T.D.; Krause, L.; Bridge, T.; Torda, G.; Raina, J.-B.; Zakrzewski, M.; Gates, R.D.; Padilla-Gamiño, J.L.; Spalding, H.L.; Smith, C.; et al. The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME J. 2015, 9, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Anbuchezhian, R.; Li, Z. Association of coral-microbes, and the ecological roles of microbial symbionts in corals. In The Cnidaria, Past, Present and Future; Goffredo, S., Dubinsky, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 347–357. [Google Scholar]
- Cárdenas, A.; Rodriguez-r, L.M.; Pizarro, V.; Cadavid, L.F.; Arévalo-Ferro, C. Shifts in bacterial communities of two caribbean reef-building coral species affected by white plague disease. ISME J. 2011, 6, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaneveld, J.R.; Burkepile, D.E.; Shantz, A.; Pritchard, C.E.; McMinds, R.; Payet, J.P.; Welsh, R.; Correa, A.M.S.; Lemoine, N.P.; Rosales, S.; et al. Overfishing and nutrient pollution interact with temperature to disrupt coral reefs down to microbial scales. Nat. Commun. 2016, 7, 11833. [Google Scholar] [CrossRef] [PubMed]
- Lampert, Y.; Kelman, D.; Dubinsky, Z.; Nitzan, Y.; Hill, R.T. Diversity of culturable bacteria in the mucus of the Red Sea coral Fungia scutaria. FEMS Microbiol. Ecol. 2006, 58, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Lampert, Y.; Kelman, D.; Nitzan, Y.; Dubinsky, Z.; Behar, A.; Hill, R.T. Phylogenetic diversity of bacteria associated with the mucus of Red Sea corals. FEMS Microbiol. Ecol. 2008, 64, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Nithyanand, P.; Pandian, S.K. Phylogenetic characterization of culturable bacterial diversity associated with the mucus and tissue of the coral Acropora digitifera from the Gulf of Mannar. FEMS Microbiol. Ecol. 2009, 69, 384–394. [Google Scholar] [CrossRef]
- Yang, S.; Sun, W.; Tang, C.; Jin, L.; Zhang, F.; Li, Z. Phylogenetic Diversity of Actinobacteria Associated with Soft Coral Alcyonium gracllimum and Stony Coral Tubastraea coccinea in the East China Sea. Microb. Ecol. 2013, 66, 189–199. [Google Scholar] [CrossRef]
- Nithyanand, P.; Manju, S.; Pandian, S.K. Phylogenetic characterization of culturable actinomycetes associated with the mucus of the coral Acropora digitifera from Gulf of Mannar. FEMS Microbiol. Lett. 2010, 314, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Valliappan, K.; Sun, W.; Li, Z. Marine actinobacteria associated with marine organisms and their potentials in producing pharmaceutical natural products. Appl. Microbiol. Biotechnol. 2014, 98, 7365–7377. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Calado, R.; Sheridan, C.; Alimonti, A.; Osinga, R. Coral aquaculture to support drug discovery. Trends Biotechnol. 2013, 31, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, J.-D.; Yang, J.; Luo, X.-M.; Zhang, S. Detection of polyketide synthase and nonribosomal peptide synthetase biosynthetic genes from antimicrobial coral-associated actinomycetes. Antonie Van Leeuwenhoek 2014, 106, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Limsuwan, S.; Voravuthikunchai, S.P. Boesenbergia pandurata (Roxb.) Schltr., Eleutherine americana Merr. and Rhodomyrtus tomentosa (Aiton) Hassk. as antibiofilm producing and antiquorum sensing in Streptococcus pyogenes. FEMS Immunol. Med Microbiol. 2008, 53, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nithyanand, P.; Thenmozhi, R.; Rathna, J.; Pandian, S.K. Inhibition of Streptococcus pyogenes Biofilm Formation by Coral-Associated Actinomycetes. Curr. Microbiol. 2010, 60, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Al-Dahash, L.M.; Mahmoud, H.M. Harboring oil-degrading bacteria: A potential mechanism of adaptation and survival in corals inhabiting oil-contaminated reefs. Mar. Pollut. Bull. 2013, 72, 364–374. [Google Scholar] [CrossRef]
- Li, J.; Kuang, W.; Long, L.; Zhang, S. Production of quorum-sensing signals by bacteria in the coral mucus layer. Coral Reefs 2017, 36, 1235–1241. [Google Scholar] [CrossRef]
- Sabdono, A.; Radjasa, O. Phylogenetic Diversity of Organophosphorous Pesticide-Degrading Coral Bacteria from Mid-West Coast of Indonesia. Biotechnology 2008, 7, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Braña, A.J.F.; Fiedler, H.-P.; Nava, H.; González, V.; Sarmiento-Vizcaíno, A.; Molina, A.; Acuña, J.L.; Garcia, L.; Blanco, M.G.B. Two Streptomyces Species Producing Antibiotic, Antitumor, and Anti-Inflammatory Compounds Are Widespread Among Intertidal Macroalgae and Deep-Sea Coral Reef Invertebrates from the Central Cantabrian Sea. Microb. Ecol. 2014, 69, 512–524. [Google Scholar] [CrossRef]
- Sarmiento-Vizcaíno, A.; González, V.; Braña, A.J.F.; Palacios-Gutiérrez, J.-J.; Otero, L.; Fernández, J.L.A.; Molina, A.; Kulik, A.; Vázquez, F.; Acuña, J.L.; et al. Pharmacological Potential of Phylogenetically Diverse Actinobacteria Isolated from Deep-Sea Coral Ecosystems of the Submarine Avilés Canyon in the Cantabrian Sea. Microb. Ecol. 2016, 73, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Rajasabapathy, R.; Ghadi, S.C.; Manikandan, B.; Mohandass, C.; Surendran, A.; Dastager, S.G.; Meena, R.M.; James, R.A. Antimicrobial profiling of coral reef and sponge associated bacteria from southeast coast of India. Microb. Pathog. 2020, 141, 103972. [Google Scholar] [CrossRef] [PubMed]
- Riyanti, R.; Nurkhasanah, W.; Radjasa, O.K. Diversity and Antifungal Activity of Actinomycetes Symbiont Hard Coral Mucus of Genera Goniopora and Porites. Makara J. Sci. 2016, 20, 193–198. [Google Scholar] [CrossRef]
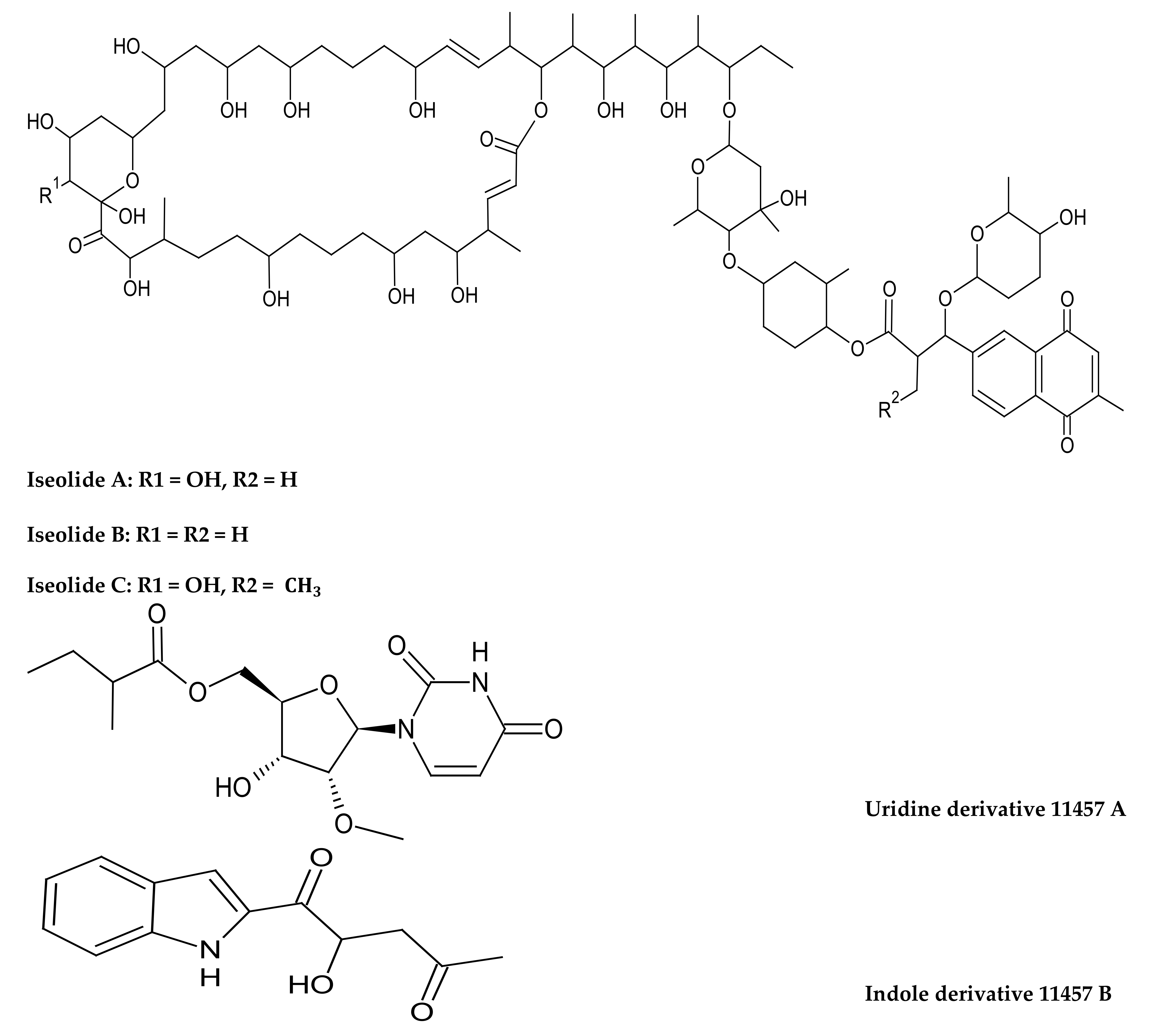
- Zhang, Z.; Zhou, T.; Harunari, E.; Oku, N.; Igarashi, Y. Iseolides A–C, antifungal macrolides from a coral-derived actinomycete of the genus Streptomyces. J. Antibiot. 2020, 73, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.S.; Ling, C.Y.; Zhou, Z.B.; Dong, Y.L.; Sun, C.L.; Song, Y.X.; Wong, N.K.; Ju, J.H. Chemical Diversity of Metabolites and Antibacterial Potential of Actinomycetes Associated with Marine Invertebrates from Intertidal Regions of Daya Bay and Nansha Islands. Microbiology 2020, 89, 483–492. [Google Scholar] [CrossRef]
- Nofiani, R.; Weisberg, A.J.; Tsunoda, T.; Panjaitan, R.G.P.; Brilliantoro, R.; Chang, J.H.; Philmus, B.; Mahmud, T. Antibacterial Potential of Secondary Metabolites from Indonesian Marine Bacterial Symbionts. Int. J. Microbiol. 2020, 2020, 8898631. [Google Scholar] [CrossRef]
- Sharma, A.R.; Harunari, E.; Oku, N.; Matsuura, N.; Trianto, A.; Igarashi, Y. Two antibacterial and PPARα/γ-agonistic unsaturated keto fatty acids from a coral-associated actinomycete of the genus Micrococcus. Beilstein J. Org. Chem. 2020, 16, 297–304. [Google Scholar] [CrossRef]
- Ocampo-Alvarez, H.; Canales, I.D.M.; Mateos-Salmón, C.; Rios-Jara, E.; Zaragoza, F.A.R.; Robles-Murguía, C.; Muñoz-Urias, A.; Hernández-Herrera, R.M.; Choix-Ley, F.J.; Becerril-Espinosa, A. Diving Into Reef Ecosystems for Land-Agriculture Solutions: Coral Microbiota Can Alleviate Salt Stress During Germination and Photosynthesis in Terrestrial Plants. Front. Plant Sci. 2020, 11, 648. [Google Scholar] [CrossRef]
- Ramalingam, V.; Varunkumar, K.; Ravikumar, V.; Rajaram, R. Production and structure elucidation of anticancer potential surfactin from marine actinomycete Micromonospora marina. Process Biochem. 2019, 78, 169–177. [Google Scholar] [CrossRef]
- González, I.S.; Juarez, O.P.; Contreras, M.R.; Roa, M.A.C.; Juárez, R.V. Adhesion ability to coral mucus of isolated bacteria from Pocillopora sp. and Porites panamensis of California. CICIMAR Ocean 2019, 34, 17–27. [Google Scholar] [CrossRef]
- Chiu, J.M.; Li, S.; Li, A.; Po, B.; Zhang, R.; Shin, P.K.; Qiu, J.-W. Bacteria associated with skeletal tissue growth anomalies in the coral Platygyra carnosus. FEMS Microbiol. Ecol. 2011, 79, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Aeby, G.S.; Work, T.; Bourne, D.G. Bacterial communities associated with healthy and Acropora white syndrome-affected corals from American Samoa. FEMS Microbiol. Ecol. 2012, 80, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Kalimutho, M.; Ahmad, A.; Kassim, Z. Isolation, characterization and identification of bacteria associated with mucus of Acropora cervicornis coral from Bidong island, Terengganu, Malaysia. Malays. J. Sci. 2007, 26, 27–39. [Google Scholar]
- Kageyama, A.; Takahashi, Y.; Yasumoto-Hirose, M.; Kasai, H.; Shizuri, Y.; Omura, S. Janibacter corallicola sp. nov., isolated from coral in Palau. J. Gen. Appl. Microbiol. 2007, 53, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-F.; Li, J.; You, Z.-Q.; Zhang, S. Prauserella coralliicola sp. nov., isolated from the coral Galaxea fascicularis. Int. J. Syst. Evol. Microbiol. 2014, 64, 3341–3345. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Pei, S.; Xie, F.; Gu, L.; Zhang, G. Saccharopolyspora coralli sp. nov. a novel actinobacterium isolated from the stony coral Porites. Int. J. Syst. Evol. Microbiol. 2020, 70, 3241–3246. [Google Scholar] [CrossRef]
- Ramaprasad, E.V.V.; Mahidhara, G.; Sasikala, C.; Ramana, C.V. Rhodococcus electrodiphilus sp. nov., a marine electro active actinobacterium isolated from coral reef. Int. J. Syst. Evol. Microbiol. 2018, 68, 2644–2649. [Google Scholar] [CrossRef]
- Li, F.; Xie, Q.; Zhou, S.; Kong, F.; Xu, Y.; Ma, Q.; Wu, W.; Huang, D.; Zhao, Y.; Huang, X. Nocardiopsis coralli sp. nov. a novel actinobacterium isolated from the coral Galaxea astreata. Int. J. Syst. Evol. Microbiol. 2021, 71, 004817. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Y.; Guo, A.; Chen, Y.; Zhang, S.; Li, J. Pseudokineococcus galaxeicola sp. nov., isolated from mucus of a stony coral. Int. J. Syst. Evol. Microbiol. 2020, 70, 5671–5675. [Google Scholar] [CrossRef]
- Buangrab, K.; Sutthacheep, M.; Yeemin, T.; Harunari, E.; Igarashi, Y.; Sripreechasak, P.; Kanchanasin, P.; Tanasupawat, S.; Phongsopitanun, W. Streptomyces corallincola and Kineosporia corallincola sp. nov., two new coral-derived marine actinobacteria. Int. J. Syst. Evol. Microbiol. 2022, 72, 005249. [Google Scholar] [CrossRef]
- Sarmiento-Vizcaíno, A.; González, V.; Braña, A.F.; Molina, A.; Acuña, J.L.; Garcia, L.; Blanco, G. Myceligenerans cantabricum sp. nov., a barotolerant actinobacterium isolated from a deep cold-water coral. Int. J. Syst. Evol. Microbiol. 2015, 65, 1328–1334. [Google Scholar] [CrossRef]
- Ben-Dov, E.; Ben Yosef, D.Z.; Pavlov, V.; Kushmaro, A. Corynebacterium maris sp. nov., a marine bacterium isolated from the mucus of the coral Fungia granulosa. Int. J. Syst. Evol. Microbiol. 2009, 59, 2458–2463. [Google Scholar] [CrossRef] [Green Version]
- Das, L.; Deb, S.; Das, S.K. Glutamicibacter mishrai sp. nov., isolated from the coral Favia veroni from Andaman Sea. Arch. Microbiol. 2019, 202, 733–745. [Google Scholar] [CrossRef]
- Raina, J.-B.; Tapiolas, D.; Motti, C.A.; Foret, S.; Seemann, T.; Tebben, J.; Willis, B.L.; Bourne, D.G. Isolation of an antimicrobial compound produced by bacteria associated with reef-building corals. PeerJ 2016, 4, e2275. [Google Scholar] [CrossRef] [Green Version]
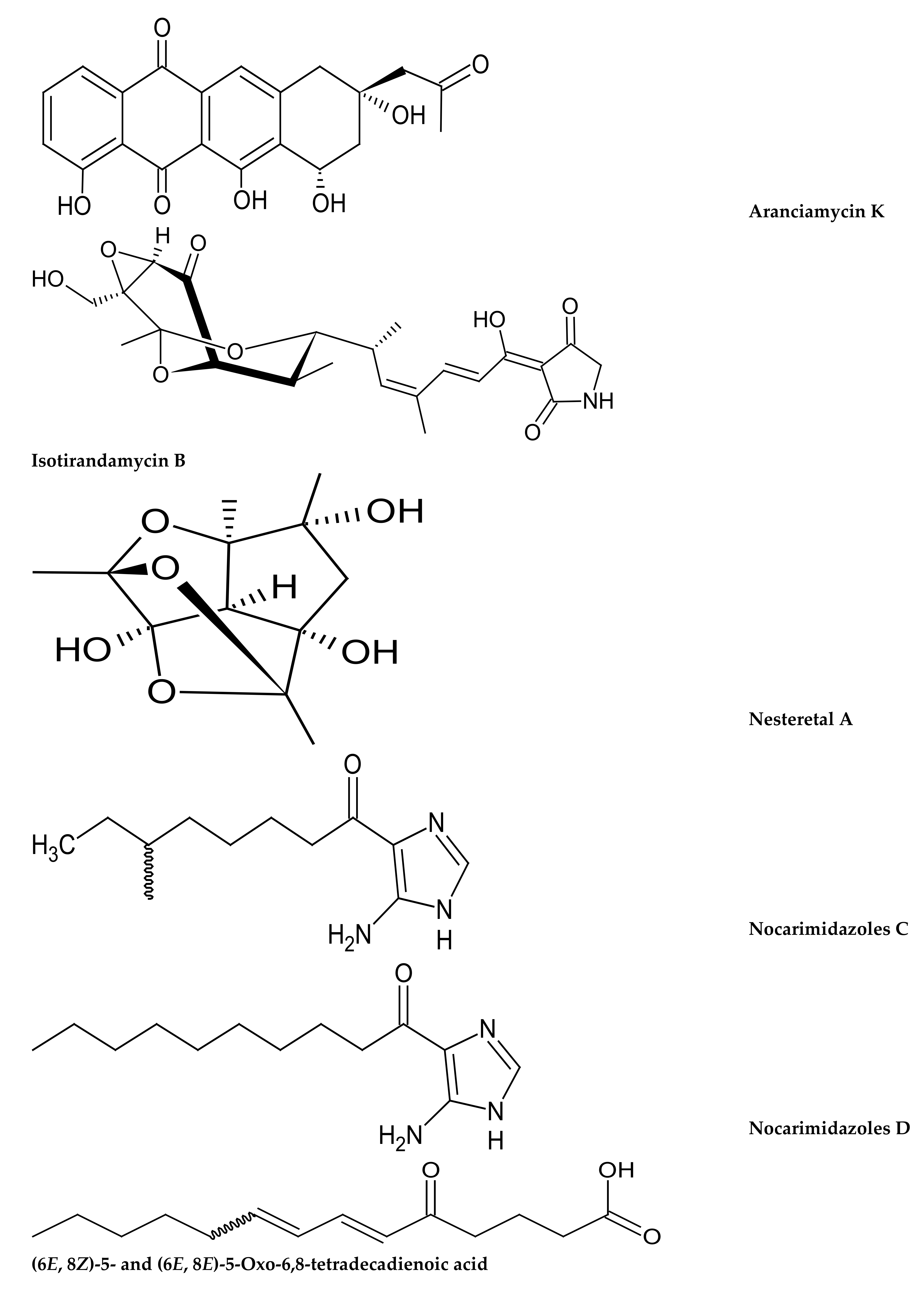
- Ul-Karim, M.R.; Harunari, E.; Sharma, A.R.; Oku, N.; Akasaka, K.; Urabe, D. Nocarimidazoles C and D, antimicrobial alkanoylimidazoles from a coral-derived actinomycete Kocuria sp.: Application of (1)J(C,H) coupling constants for the unequivocal determination of substituted imidazoles and stereochemical diversity of anteisoalkyl chains in microbial metabolites. Beilstein J. Org. Chem. 2020, 16, 2719–2727. [Google Scholar] [CrossRef]
- Fang, Z.; Chen, S.; Zhu, Y.; Li, J.; Khan, I.; Zhang, Q.; Zhang, C. A new uridine derivative and a new indole derivative from the coral-associated actinomycete Pseudonocardia sp. SCSIO 11457. Nat. Prod. Res. 2019, 35, 188–194. [Google Scholar] [CrossRef] [PubMed]
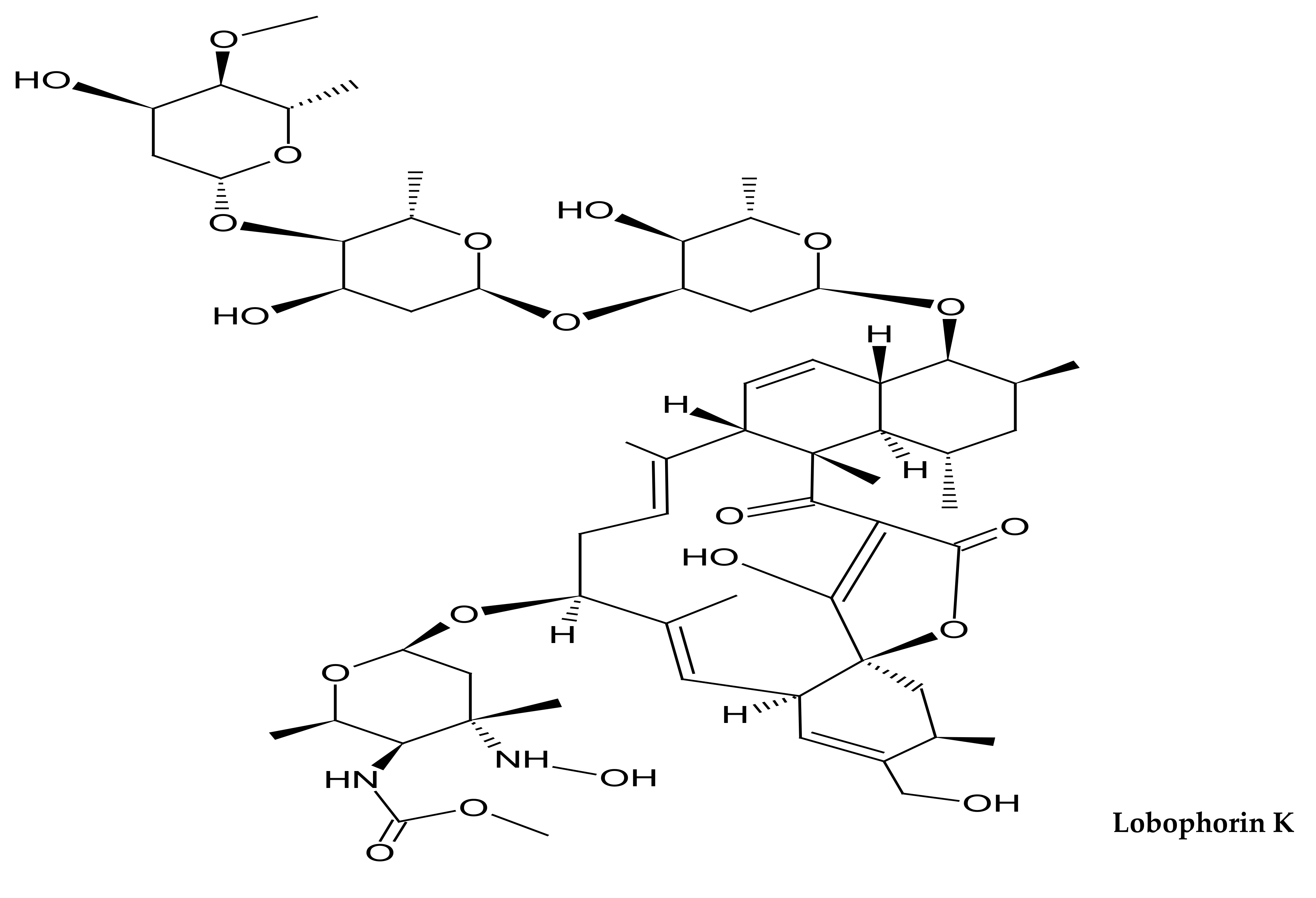
- Braña, A.F.; Sarmiento-Vizcaíno, A.; Osset, M.; Pérez-Victoria, I.; Martín, J.; de Pedro, N.; de la Cruz, M.; Díaz, C.; Vicente, F.; Reyes, F.; et al. Lobophorin K, a New Natural Product with Cytotoxic Activity Produced by Streptomyces sp. M-207 Associated with the Deep-Sea Coral Lophelia pertusa. Mar. Drugs 2017, 15, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Yang, B.; Zhou, X.; Huang, D.; Wang, J. Cytotoxic anthracycline and antibacterial tirandamycin analogues from a marine-derived Streptomyces sp. SCSIO 41399. J. Antibiot. 2018, 72, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-L.; Chen, R.; Yang, S.; Xia, J.-M.; Zhang, G.-Y.; Chen, C.-H.; Zhang, Y.; Yang, X.-W. Nesteretal A, A Novel Class of Cage-Like Polyketide from Marine-Derived Actinomycete Nesterenkonia halobia. Org. Lett. 2019, 21, 8174–8177. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genus/Species | Family | Coral Species and Nature of Sample | Isolation Medium, Incubation Temperature/Time, Pretreatment and Bioactivity | Country/Sampling Location | Ref. |
|---|---|---|---|---|---|
| Micrococcus luteus Dermatophilus congolensis Kytococcus sedentarius Kocuria sp. | Micrococcaceae Dermatophilaceae Kytococcaceae | Fungia scutaria (depth of 1–2 m) (mucus) | Bacto Marine Agar 2216 + salt concentration (3.6%). Incubated at 25 °C for 3–7 days. (No bioactivity test) | Gulf of Eilat, northern Red Sea. | [156] |
| Actinomyces sp. Micrococcus roseus Micrococcus sp. Micrococcus varians | Actinomycetaceae Micrococcaceae | Acropora cervicornis (depth of 4–5 m) (mucus) | Sucrose seawater medium. Incubated at 28 °C for 24 h. (No bioactivity test) | North of Bidong Island, Terengganu, Malaysia | [182] |
| Brachybacterium sp. Brevibacterium sp. Kytococcus sp. | Dermabacteriaceae Brevibacteriaceae Kytococcaceae | Porites sp. Galaxea sp. Acopora sp. Pavona sp. (depth of 2–3 m) (tissue) | Half-strength Zobell 2216E marine agar. Incubated at room temperature for 48 h. (Capable of degrading organophosphorous pesticides) | Java, Bali, Sulawesi and Komodo Island | [168] |
| Brachybacterium paraconglomeratum Brevibacterium linesis Kocuria rosea Kocuria flavus | Dermabacteriaceae Brevibacteriaceae Micrococcaceae | Acropora digitifera (mucus and tissue slurry) | Zobell Marine agar. Incubated at 27 °C for 7–12 days. (Antibacterial activity) | India (gulf of manner) | [158] |
| Streptomyces akiyoshinensis Actinobacterium sp. | Streptomycetaceae Actinomycetaceae | Acropora digitifera (mucus) | Starch casein agar. Incubated at 27 °C for 7–12 days. (Antibiofilm activity) | India (gulf of manner) | [165] |
| Streptomyces Micrococcus Curtobacterium Propionibacterium | Streptomycetaceae Micrococcaceae Microbacteriaceae Propionibacteriaceae | Acropora digitifera (mucus) | Starch casein agar + actidione (40 µgml−1) and nalidixic acid (10 µgml−1). Incubated at 28 °C for 10–14 days (Antibacterial activity) | India (gulf of manner) | [160] |
| Micrococcus luteus Mycrobacterium paraoxydans Brachybacterium paraconglomeratum Kocuria rhizophila Kytococcus sedentarius | Micrococcaceae Microbacteriaceae Dermabacteraceae Kytococcaceae | Acropora hyacinthus (depth < 10 m) (mucus) | Marine agar. Incubated at 28 °C for 48 h. (No bioactivity test) | Vatia Bay, American Samoa | [181] |
| Nocardiopsis alba Micrococcus yunnanensis | Nocardiopsaceae Micrococcaceae | Diploria strigosa (depth of 15 m) (crushed coral) | Luria Bertani agar and Marine agar. Incubated at 28 °C for 48 h. (No bioactivity test) | Aguja Island, Panama | [154] |
| Brevibacterium linens Leucobacter komagatae Microbacterium arabinogalactanolyticum Microbacterium oxydans Brevibacterium fermense Dietzia sp. | Brevibacteriaceae Microbacteriaceae Dietziaceae | Siderastrea siderea (depth of 15 m) (crushed coral) | |||
| Rothia amarae | Micrococcaceae | Platygra carnosus (crushed coral) | Heterotrophic bacterial media + artificial seawater. Incubated at 20 °C for 48–72 h. (No bioactivity test) | Hoi Marine Park, China | [180] |
| Dietzia maris Gordonia bronchialis Gordonia lacunae Kocuria flava Micrococcus luteus Mycobacterium Chlorophenolicum | Dietziaceae Gordoniaceae Micrococcaceae Microbacteriaceae | Acropora clathrata Porites compressa (mucus and tissue) | Basal Inorganic Medium + 0.5% light crude oil (carbon source). Incubated at 30 °C for 1 week. (Oil degraders) | Kuwait | [166] |
| Dermacoccus sp. Serinicoccus sp. Rhodococcus sp. Nacardioides sp. Micromonospora sp. Leucobacter sp. Brevibacterium sp. | Dermacoccaceae Ornithinimicrobiaceae Nocardiaceae Nocardioidaceae Micromonosporaceae Microbacteriaceae Brevibacteriaceae | Tubastraea coccinea (depth of 5–10 m) (crushed coral) | Glycerol arginine agar (GAA), asparagine peptone agar (APA) and R2A + ASW + potassium dichromate (50 μgml−1) with nalidixic acid (15 μgml−1). Incubated at 28 °C for 3–6 weeks. (No bioactivity test) | Zhao’an Bay, East China Sea | [159] |
| Streptomyces variabilis Streptomyces rutgersensis Streptomyces viridodiastaticus Mycobacterium gilvum Mycobacterium parafortuitum Mycobacterium vanbaalenii Nocardiopsis yanglingensis Micromonospora aurantiaca Brevibacterium epidermidis Brevibacterium picturae Gordonia sp. Gordonia westfalica Brachybacterium paraconglomeratum Cellulosimicrobium funkei | Streptomycetaceae Mycobacteriaceae Nocardiopsaceae Micromonosporaceae Brevibacteriaceae Gordoniaceae Dermabacteraceae Promicromonosporaceae | Porites lutea (depth of 3–5 m) (tissue) | Marine agar, yeast extract agar, trehalose proline agar, raffinose histidine agar and pyruvic acid sodium asparagine agar + natural seawater. Media pH-7.5, Incubated at 28 °C for 3–4 weeks. (Antimicrobial activity) | South China sea | [163] |
| Streptomyces variabilis Streptomyces fimicarius Micrococcus yunnanensis Microbacterium aerolatum Nocardiopsis flavescens Nocardiopsis dassonvillei Nocardiopsis alba Micromonospora aurantiaca Brevibacterium epidermidis Brevibacterium picturae Tsukamurella pulmonis Gordonia sp. Amycolatopsis sp. Jiangella alba Jiangella gansuensis Brachybacterium paraconglomeratum Prauserella marina Pseudonocardia kongjuensis | Streptomycetaceae Micrococcaceae Microbacteriaceae Nocardiopsaceae Micromonosporaceae Brevibacteriaceae Tsukamurellaceae Gordoniaceae Pseudonocardiaceae Jiangellaceae Dermabacteraceae | Galaxea fascicularies (depth of 3–5 m) (tissue) | |||
| Streptomyces fimicarius Microbacterium ginsengisoli Microbacterium paraoxidans Nocardiopsis flavescens Nocardiopsis yanglingensis Micromonospora aurantiaca Brevibacterium epidermidis Gordonia sp. Jiangella gansuensis Pseudonocardia carboxydivorans Pseudonocardia ammoniooxydans Cellulosmicrobium funkei | Streptomycetaceae Microbacteriaceae Nocardiopsaceae Micromonosporaceae Brevibacteriaceae Gordoniaceae Jiangellaceae Pseudonocardiaceae Promicromonosporaceae | Acropora millepora (depth of 3–5 m) (tissue) | |||
| Streptomyces cyaneofuscatus Streptomyces carnosus | Streptomycetaceae | Deep sea corals; Lophelia pertusa, and Desmophyllum sp. (depth of 1800 m) (crushed coral) | One-third tryptic soy agar (TSA) and 1/6 MOPS BLEB agar (Oxoid) + seawater + cycloheximide (80 µgml−1) and nalidixic acid (20 µgm−1). Incubated at 28 °C for 2 weeks. (antibacterial, antifungal, cytotoxic, antiinflammatory activities) | Aviles Canyon, Cantabrian sea. | [169] |
| Streptomyces sp. Rhodococcus sp. Nocardia sp. Brevibacterium sp. Micrococcus sp. Devriesea sp. Kocuria sp. Cellulomonas sp. Arthrobacter sp. Dermacoccus sp. | Streptomycetaceae Nocardiaceae Brevibacteriaceae Micrococcaceae Dermabacteraceae Cellulomonadaceae Dermacoccaceae | Coscinaraea columna (mucus, tissue and skeleton) | R2A medium, M2 medium, M4 medium and Starch Casein Agar (SCA). R2A and SCA media + 3% (w/v) NaCl + potassium dichromate (50 µgml−1), nalidixic acid (15 µg ml−1), cycloheximide (75 µgml−1) and nystatin (75 µgml−1). Media pH-7.6, incubated at 28–30 °C for 3–6 weeks. (Antimicrobial activity) | Kuwait | [23] |
| Streptomyces sp. Rhodococcus sp. Micromonospora sp. Dietzia sp. Brevibacterium sp. Micrococcus sp. Brachybacterium sp. Kocuria sp. Microbacterium sp. Cellulomonas sp. Arthrobacter sp. Ornithinimicrobium sp. Kineococcus sp. Agrococcus sp. | Streptomycetaceae Nocardiaceae Micromonosporaceae Dietziaceae Brevibacteriaceae Micrococcaceae Dermabacteraceae Microbacteriaceae Cellulomonadaceae Ornithinimicrobiaceae Kineosporiaceae | Platygyra daedalea (mucus, tissue and skeleton) | |||
| Streptomyces sp. Rhodococcus sp. Nocardia sp. Brevibacterium sp. Micrococcus sp. Kocuria sp. Microbacterium sp. Arthrobacter sp. Marmoricola sp. | Streptomycetaceae Nocardiaceae Brevibacteriaceae Micrococcaceae Microbacteriaceae Nocardioidaceae | Porites harrisoni (mucus, tissue and tissue) | |||
| Not specified; Actinomycete isolates with different codes including a prominent antifungal code SCAS324 | Goniopora spp. and Porites spp. (mucus) | Starch nitrate agar and starch casein agar + seawater. Media pH-7.2 and 7.4, respectively. Incubated at 30 °C for 4 weeks. (Antifungal activity) | Indonesia | [172] | |
| Rothia amarae | Micrococcaceae | Galaxea sp. Porites lutea (mucus) | Marine agar 2216. Incubated at 30 °C for 1–2 weeks. (Produces autoinductor 2) | China | [167] |
| Streptomyces sp. | Streptomycetaceae | Deep sea corals (depth of 2000 m) (crushed coral) | One-third tryptic soy agar (TSA) and 1/6 MOPS BLEB agar (Oxoid) + seawater + cycloheximide (80 µgml−1) and nalidixic acid (20 µgml−1). Incubated at 28 °C for 2 weeks. (Antifungal and antibacterial activities) | Aviles Canyon, Cantabrian sea. | [170] |
| Kocuria turfanensis | Micrococcaceae | Porites panamensis (depth of 2–10 m) (crushed coral) | Marine agar. (No bioactivity test) | Gulf of California, Mexico | [179] |
| Micromonospora marina | Micromonosporaceae | Acropora formosa (mucus) | Starch casein agar + cycloheximide and nalidixic acid. Incubated for 7 days. Pre-heat treatment before serial dilution. (Potent biosurfactant-anticancer) | Not stated | [178] |
| Streptomyces sp. | Streptomycetaceae | Dendrophyllia sp. (depth of 20 to 25 m) (crushed coral) | ISP4 agar medium. Incubated at 23 °C for 14 days. (Antifungal activity) | Japan | [173] |
| Micrococcus sp. | Micrococcaceae | Catalaphyllia sp. (crushed coral) | Marine agar 2216 (Difco). Incubated at 23 °C for 2 days. (Antibacterial activity) | Japan | [176] |
| Salinispora arenicola | Micromonosporaceae | Porites lobata Porites panamensis (tissue) | Ten percent A1 culture medium + cycloheximide (100 μgml−1) and gentamicin (5 μgml−1). Incubated at 28 °C for 2 weeks. Coral tissue dried in laminar flow hood for 72 h before serial dilution. (Plant biostimulant activity) | Tropical central Pacific | [177] |
| Streptomyces sp. | Streptomycetaceae | Unidentified stony corals (crushed coral) | Nine different enrichment media; M1 (Mycose agar), M2 (Actinomycete isolation agar), M3 (Glucose asparagine agar), M4 (International Streptomyces Project), M5 (Humic Vitamin acid agar), M6 (Glycerin agar), M7 (Chitin agar), M8 (Gauze’s no.1 agar) and M9 (Marine agar) + nystatin (50 µgml−1) and trimethoprim (50 µgml−1). Incubated at room temperature for 3–5 weeks. Wet heat treatment (55 °C for 5 min) and incubated at 28 °C in a rotary shaker at 200 rpm for 30 min to facilitate the growth of actinomycetes. (Antibacterial activity) | South China Sea | [174] |
| Streptomyces sp. | Streptomycetaceae | Unidentified hard coral sample (crushed coral) | ISP2 medium + 0–5% NaCl + nystatin (100 µgml−1) and nalidixic (100 µgml−1). Incubated at 27–32 °C (Antibacterial activity) | Indonesia | [175] |
| Glutamicibacter mysorens | Micrococcaceae | Favites halicora (mucus) | Nutrient agar, marine agar, R2A agar, starch casein agar and International Streptomyces Project medium-2 (ISP-2) agar + seawater + cycloheximide (100 μgml−1) and nalidixic acid (25 μgml−1). Incubated at 26 ± 2 °C for 3 weeks. (Antimicrobial activity) | Southeast coast of India | [171] |
| Novel Species | Family | Coral Species and Nature of Sample | Isolation Medium and Incubation Temperature/Time | Ref. |
|---|---|---|---|---|
| Janibacter corallicola | Intrasporangiaceae | Acropora gemmifera (depth of 3–5 m) (crushed coral) | One-tenth MA + Ca medium. Incubated at 25 °C for about 1–3 weeks. | [183] |
| Corynebacterium maris | Corynebacteriaceae | Fungia granulosa (depth of 10–15 m) (coral mucus) | Marine agar, LB agar and nutrient agar. Incubated at 30 °C for 48 to 72 h. | [191] |
| Prauserella coralliicola | Pseudonocardiaceae | Galaxea fascicularis (depth of 5 m) (tissue slurry) | Isolation medium (yeast extract 0.25 g, 0.5 g, agar 12 g, 500 mL seawater and 500 mL distilled water; pH 7.0). Incubated at 28 °C for 4 weeks. | [184] |
| Myceligenerans cantabricum | Promicromonosporaceae | Deep sea coral (order: Scleractinia, Family: Caryophillidae) (depth of 1500 m) (crushed coral) | Selective media; 1/3 tryptic soy (TSA, Merck) and 1/6 M-BLEB agar + seawater + cycloheximide (80 µgml−1) and nalidixic acid (20 µgml−1). | [190] |
| Rhodococcus electrodiphilus | Nocardiaceae | An unidentified stony coral sample (crushed coral) | Marine agar (MA; Hi-Media) incubated at 25 °C for 1 week. | [186] |
| Saccharopolyspora coralli. | Pseudonocardiaceae | Porites sp. (depth of 5 m) (crushed coral) | ISP media 2 & 4 (fast growth), 3 & 6 (slow growth) and TSA (fast growth). Incubated at 25–30 °C for 3 weeks. | [185] |
| Pseudokineococcus galaxeicola | Kineosporiaceae | Galaxea sp. (depth of 4.2 m) (coral mucus) | Mucus agar medium. Cultivated at 25 °C for 30 days. | [188] |
| Glutamicibacter mishrai | Micrococcaceae | Favia veroni | Isolation medium; Marine agar 2216. Incubated at 25 °C for 4 days. | [192] |
| Nocardiopsis coralli | Nocardiopsaceae | Galaxea astreata (crushed coral) | Gause modified medium 1 + potassium dichromate (75 µgml−1). Cultivated at 28 °C for 4 weeks. | [187] |
| Streptomyces corallincola Kineosporia corallincola | Streptomycetaceae Kineosporiaceae | Favites pentagona | [189] |
| New Compound | Chemical Class | Source | Bioactivity | Ref. |
|---|---|---|---|---|
| Lobophorin K | Spirotetronate | Streptomyces sp. from deep sea coral Lophelia pertusa | Cytotoxic activity | [196] |
| Aranciamycin K Isotirandamycin B | Anthracycline Tirandamycin | Streptomyces sp. from Porites sp. | Isotirandamycin B showed antimicrobial activity against Streptococcus agalactiae with a MIC of 11.5 μM | [197] |
| Nesteretal A | Polyketide | Nesterenkonia halobia from scleratinian coral Platygyra | Showed a weak retinoid X receptor-α transcriptional activation effect | [198] |
| Nocarimidazoles C Nocarimidazoles D | Alkanoylimidazoles | Kocuria sp from stony coral mycedium sp. | Moderate antimicrobial activity against gram-positive bacteria and fungi (MIC—6.25–25 μg/mL) | [194] |
| (6E, 8Z)-5- and (6E, 8E)-5- Oxo-6,8-tetradecadienoic acid | Keto fatty acids | Actinomycete from the genera Micrococcus sp. which is associated with stony coral Catalaphyllia sp. | Showed antibacterial activity against the plant pathogen Rhizobium radiobacter and the fish pathogen Tenacibaculum maritimum. | [176] |
| Iseolide A Iseolide B Iseolide C | Macrolides | Streptomyces sp. associated with the stony coral of the genus Dendrophyllia | Showed antifungal activity against the plant pathogen Glomerella cingulate and human pathogens Candida albicans and Trichophyton rubrum. (MIC—6.25–25 μg/mL) | [173] |
| Uridine derivative 11457 A Indole derivative 11457 B | Alkaloids | Pseudonocardia sp. from the stony coral Galaxea fascicularis | Showed no antibacterial activity against pathogenic bacteria and cytotoxic against human cancer cell lines | [195] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siro, G.; Pipite, A.; Christi, K.; Srinivasan, S.; Subramani, R. Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites. Microorganisms 2022, 10, 1349. https://doi.org/10.3390/microorganisms10071349
Siro G, Pipite A, Christi K, Srinivasan S, Subramani R. Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites. Microorganisms. 2022; 10(7):1349. https://doi.org/10.3390/microorganisms10071349
Chicago/Turabian StyleSiro, Galana, Atanas Pipite, Ketan Christi, Sathiyaraj Srinivasan, and Ramesh Subramani. 2022. "Marine Actinomycetes Associated with Stony Corals: A Potential Hotspot for Specialized Metabolites" Microorganisms 10, no. 7: 1349. https://doi.org/10.3390/microorganisms10071349