
Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut–Brain Axis

 , , , and
, , , and 
Abstract
:1. Introduction
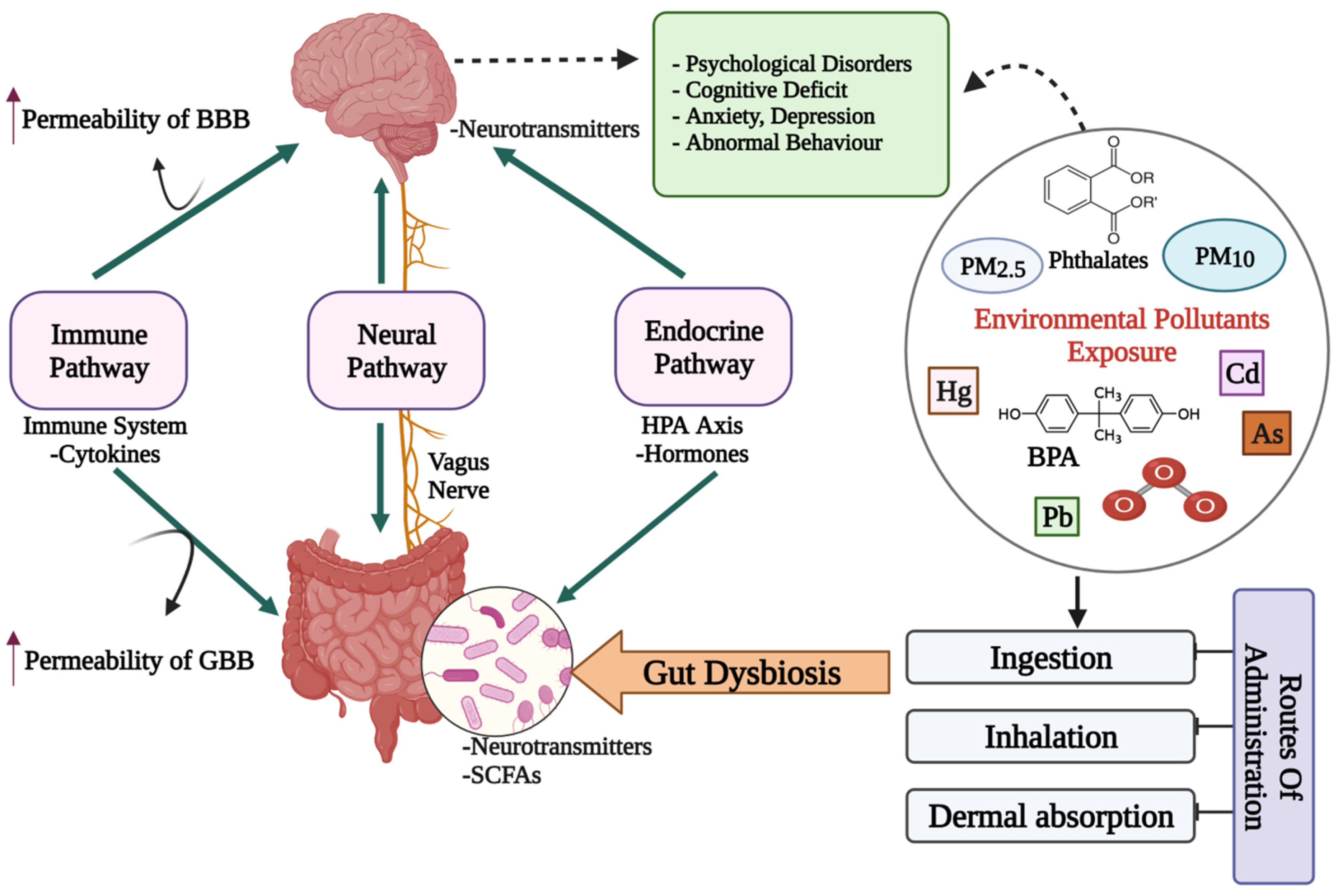
2. The Microbiota–Gut–Brain Axis
Crosstalk between Microbiota, Gut and Brain
3. Impact of Environmental Pollutants on Gut Dysbiosis and Mental Health
3.1. Heavy Metals
3.2. Phthalates
3.3. Bisphenol A
3.4. Air Pollutants
4. Microbiota-Targeted Interventions for Mental Health
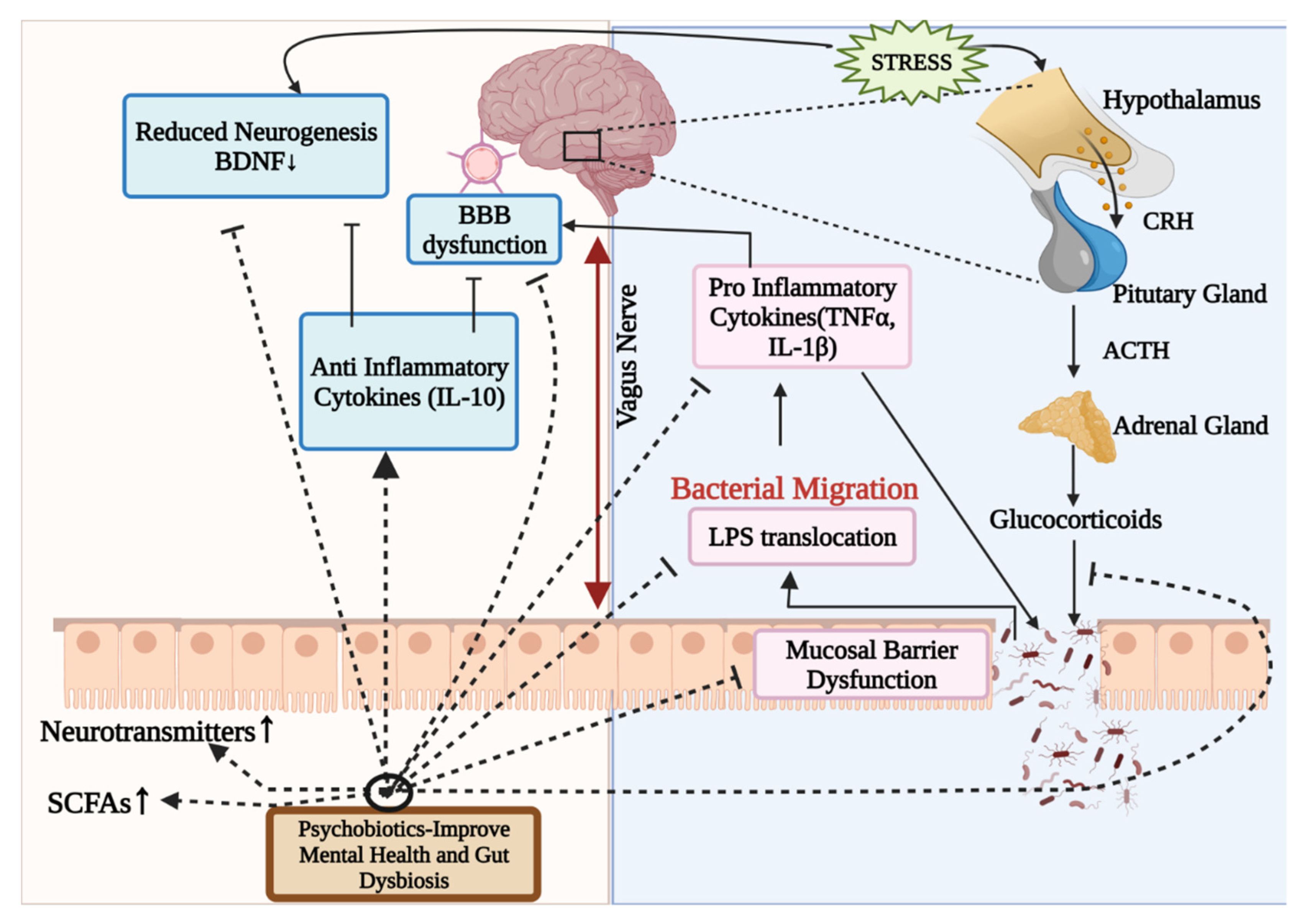
4.1. Psychobiotics
Possible Mode of Action of Psychobiotics
5. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsythe, P.; Kunze, W.A. Voices from within: Gut microbes and the CNS. Cell. Mol. Life Sci. 2012, 70, 55–69. [Google Scholar] [CrossRef]
- Liu, L.; Huh, J.R.; Shah, K. Microbiota and the gut-brain-axis: Implications for new therapeutic design in the CNS. eBioMedicine 2022, 77, 103908. [Google Scholar] [CrossRef] [PubMed]
- Bezek, K.; Petelin, A.; Pražnikar, J.; Nova, E.; Redondo, N.; Marcos, A.; Pražnikar, Z.J. Obesity Measures and Dietary Parameters as Predictors of Gut Microbiota Phyla in Healthy Individuals. Nutrients 2020, 12, 2695. [Google Scholar] [CrossRef]
- Davenport, E.R.; Sanders, J.G.; Song, S.J.; Amato, K.R.; Clark, A.G.; Knight, R. The human microbiome in evolution. BMC Biol. 2017, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- Groussin, M.; Mazel, F.; Alm, E.J. Co-evolution and Co-speciation of Host-Gut Bacteria Systems. Cell Host Microbe 2020, 28, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Long, D.; Liu, M.; Li, H.; Song, J.; Jiang, X.; Wang, G.; Yang, X. Dysbacteriosis induces abnormal neurogenesis via LPS in a pathway requiring NF-κB/IL-6. Pharmacol. Res. 2021, 167, 105543. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.; Young, R.; Wong, M.-L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Mental Health. Available online: https://www.cdc.gov/mentalhealth/index.htm (accessed on 6 May 2022).
- World Health Organisation. Mental Health. Available online: https://www.who.int/health-topics/mental-health#tab=tab_2 (accessed on 31 December 2021).
- World Health Organisation. The World Health Report 2001: Mental Health: New Understanding, New Hope; World Health Organisation: Geneva, Switzerland, 2001. [Google Scholar]
- Ayuso-Álvarez, A.; Simón, L.; Nuñez, O.; Rodríguez-Blázquez, C.; Martín-Méndez, I.; Bel-Lán, A.; López-Abente, G.; Merlo, J.; Fernandez-Navarro, P.; Galán, I. Association between heavy metals and metalloids in topsoil and mental health in the adult population of Spain. Environ. Res. 2019, 179, 108784. [Google Scholar] [CrossRef]
- World Health Organisation. Mental Disorders. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-disorders#:~:text=They%20are%20generally%20characterized%20by,and%20developmental%20disorders%20including%20autism (accessed on 8 June 2022).
- WebMD. Causes of Mental Illness. Available online: https://www.webmd.com/mental-health/mental-health-causes-mental-illness (accessed on 30 June 2022).
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, H.-R. Vagal and hormonal gut-brain communication: From satiation to satisfaction. Neurogastroenterol. Motil. 2008, 20, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Taché, Y.; Vale, W.; Rivier, J.; Brown, M. Brain regulation of gastric secretion: Influence of neuropeptides. Proc. Natl. Acad. Sci. USA 1980, 77, 5515–5519. [Google Scholar] [CrossRef] [Green Version]
- Agustí, A.; García-Pardo, M.P.; López-Almela, I.; Campillo, I.; Maes, M.; Romani-Pérez, M.; Sanz, Y. Interplay Between the Gut-Brain Axis, Obesity and Cognitive Function. Front. Neurosci. 2018, 12, 155. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, A.M.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. Q. Publ. Hell. Soc. Gastroenterol. 2015, 28, 203. [Google Scholar]
- Rhee, H.S.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain–gut–enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, S.; Chen, R. Chapter One—Metaproteomic analysis of human gut microbiome in digestive and metabolic diseases. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–12. [Google Scholar]
- Balaguer-Trias, J.; Deepika, D.; Schuhmacher, M.; Kumar, V. Impact of Contaminants on Microbiota: Linking the Gut–Brain Axis with Neurotoxicity. Int. J. Environ. Res. Public Health 2022, 19, 1368. [Google Scholar] [CrossRef]
- Hattori, N.; Yamashiro, Y. Gut Brain Axis (GBA). Ann. Nutr. Metab. 2021, 77, 1–3. [Google Scholar] [CrossRef]
- Forsythe, P.; Bienenstock, J.; Kunze, W.A. Vagal pathways for microbiome-brain-gut axis communication. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014; pp. 115–133. [Google Scholar]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.-N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef]
- Mudd, A.T.; Berding, K.; Wang, M.; Donovan, S.M.; Dilger, R.N. Serum cortisol mediates the relationship between fecal Ruminococcus and brain N-acetylaspartate in the young pig. Gut Microbes 2017, 8, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, E.; Dinan, T.G.; Cryan, J.F. Recent developments in understanding the role of the gut microbiota in brain health and disease. Ann. N. Y. Acad. Sci. 2017, 1420, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef]
- Calvani, R.; Picca, A.; Lo Monaco, M.R.; Landi, F.; Bernabei, R.; Marzetti, E. Of Microbes and Minds: A Narrative Review on the Second Brain Aging. Front. Med. 2018, 5, 53. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The Role of Short-Chain Fatty Acids in Health and Disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Del Toro-Barbosa, M.; Hurtado-Romero, A.; Garcia-Amezquita, L.E.; García-Cayuela, T. Psychobiotics: Mechanisms of Action, Evaluation Methods and Effectiveness in Applications with Food Products. Nutrients 2020, 12, 3896. [Google Scholar] [CrossRef]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dockray, G.J. Gastrointestinal hormones and the dialogue between gut and brain. J. Physiol. 2014, 592, 2927–2941. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, U.; Woods, S.C.; Langhans, W.; Meyer, U. PYY3–36: Beyond food intake. Front. Neuroendocrinol. 2015, 38, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Anderberg, R.H.; Anefors, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. Dopamine signaling in the amygdala, increased by food ingestion and GLP-1, regulates feeding behavior. Physiol. Behav. 2014, 136, 135–144. [Google Scholar] [CrossRef]
- Ventriglio, A.; Bellomo, A.; di Gioia, I.; Di Sabatino, D.; Favale, D.; De Berardis, D.; Cianconi, P. Environmental pollution and mental health: A narrative review of literature. CNS Spectrums 2020, 26, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Simkin, D.R. Microbiome and Mental Health, Specifically as It Relates to Adolescents. Curr. Psychiatry Rep. 2019, 21, 93. [Google Scholar] [CrossRef]
- Obrenovich, M.E.M. Leaky Gut, Leaky Brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.M.; Bercik, P. The Relationship Between Intestinal Microbiota and the Central Nervous System in Normal Gastrointestinal Function and Disease. Gastroenterology 2009, 136, 2003–2014. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Silbergeld, E.K.; Goldberg, A. Pharmacological and neurochemical investigations of lead-induced hyperactivity. Neuropharmacology 1975, 14, 431–444. [Google Scholar] [CrossRef]
- Yang, M.-Y.; Kim, S.-H.; Kim, J.-C.; Shin, T.-K.; Moon, C.-J. Toluene Induces Depression-Like Behaviors in Adult Mice. Toxicol. Res. 2010, 26, 315–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanninen, H.; Mantere, P.; Hernberg, S.; Seppalainen, A.M.; Kock, B. Subjective symptoms in low-level exposure to lead. Neurotoxicology 1979, 1, 333–347. [Google Scholar]
- Opler, M.G.; Buka, S.L.; Groeger, J.; McKeague, I.; Wei, C.; Factor-Litvak, P.; Bresnahan, M.; Graziano, J.; Goldstein, J.M.; Seidman, L.J.; et al. Prenatal exposure to lead, delta-aminolevulinic acid, and schizophrenia: Further evidence. Environ. Health Perspect. 2008, 116, 1586–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, P.; Hossain, M.I.; Sadat, A.N.; Nahar, Z.; Hossain, M.K.; Hasnat, A. Serum levels of cadmium, calcium, lead and iron in schizophrenic patients. Dhaka Univ. J. Pharm. Sci. 2006, 5, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Hanninen, H.; Aitio, A.; Kovala, T.; Luukkonen, R.; Matikainen, E.; Mannelin, T.; Erkkila, J.; Riihimaki, V. Occupational exposure to lead and neuropsychological dysfunction. Occup. Environ. Med. 1998, 55, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Jurczak, A.; Brodowska, A.; Szkup, M.; Prokopowicz, A.; Karakiewicz, B.; Łój, B.; Kotwas, A.; Brodowska, A.; Grochans, E. Influence of Pb and Cd levels in whole blood of postmenopausal women on the incidence of anxiety and depressive symptoms. Ann. Agric. Environ. Med. 2018, 25, 219–223. [Google Scholar] [CrossRef]
- Homme, K.G.; Kern, J.K.; Haley, B.E.; Geier, D.A.; King, P.G.; Sykes, L.K.; Geier, M.R. New science challenges old notion that mercury dental amalgam is safe. Biometals 2014, 27, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Harakeh, S.; Sabra, N.; Kassak, K.; Doughan, B. Factors influencing total mercury levels among Lebanese dentists. Sci. Total Environ. 2002, 297, 153–160. [Google Scholar] [CrossRef]
- Shiue, I. Urinary heavy metals, phthalates and polyaromatic hydrocarbons independent of health events are associated with adult depression: USA NHANES, 2011–2012. Environ. Sci. Pollut. Res. 2015, 22, 17095–17103. [Google Scholar] [CrossRef]
- Lahouaoui, H.; Aimrane, A.; Khamsi, Y.; Zouhairi, N.; Benammi, H.; El Hidan, M.A.; Draoui, A.; Alahyane, H.; Bouazza, A. Depression and Anxiety Emerging From Heavy Metals: What Relationship? In Handbook of Research on Global Environmental Changes and Human Health; IGI Global: Pennsylvania, PA, USA, 2019; pp. 305–321. [Google Scholar]
- Tshala-Katumbay, D.; Mwanza, J.-C.; Rohlman, D.S.; Maestre, G.E.; Oriá, R. A global perspective on the influence of environmental exposures on the nervous system. Nature 2015, 527, S187–S192. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Yu, L.; Tian, F.; Zhai, Q.; Fan, L.; Chen, W. Gut microbiota: A target for heavy metal toxicity and a probiotic protective strategy. Sci. Total Environ. 2020, 742, 140429. [Google Scholar] [CrossRef] [PubMed]
- Giambò, F.; Italia, S.; Teodoro, M.; Briguglio, G.; Furnari, N.; Catanoso, R.; Costa, C.; Fenga, C. Influence of toxic metal exposure on the gut microbiota (Review). World Acad. Sci. J. 2021, 3, 19. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Gut Dysbiosis in Animals Due to Environmental Chemical Exposures. Front. Cell. Infect. Microbiol. 2017, 7, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.; Abo, R.P.; Schlieper, K.A.; Graffam, M.E.; Levine, S.S.; Wishnok, J.S.; Swenberg, J.A.; Tannenbaum, S.R.; Fox, J.G. Arsenic Exposure Perturbs the Gut Microbiome and Its Metabolic Profile in Mice: An Integrated Metagenomics and Metabolomics Analysis. Environ. Health Perspect. 2014, 122, 284–291. [Google Scholar] [CrossRef]
- Lu, K.; Mahbub, R.; Cable, P.H.; Ru, H.; Parry, N.M.A.; Bodnar, W.M.; Wishnok, J.S.; Styblo, M.; Swenberg, J.A.; Fox, J.G.; et al. Gut Microbiome Phenotypes Driven by Host Genetics Affect Arsenic Metabolism. Chem. Res. Toxicol. 2014, 27, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Liu, S.; Wang, Z.; Zhang, X.-X.; Li, M.; Wu, B. Metagenomic profiles and antibiotic resistance genes in gut microbiota of mice exposed to arsenic and iron. Chemosphere 2014, 112, 1–8. [Google Scholar] [CrossRef]
- Dheer, R.; Patterson, J.; Dudash, M.; Stachler, E.N.; Bibby, K.J.; Stolz, D.B.; Shiva, S.; Wang, Z.; Hazen, S.L.; Barchowsky, A.; et al. Arsenic induces structural and compositional colonic microbiome change and promotes host nitrogen and amino acid metabolism. Toxicol. Appl. Pharmacol. 2015, 289, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Chi, L.; Bian, X.; Gao, B.; Ru, H.; Tu, P.; Lu, K. Sex-Specific Effects of Arsenic Exposure on the Trajectory and Function of the Gut Microbiome. Chem. Res. Toxicol. 2016, 29, 949–951. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wen, X.; Faulk, C.; Boehnke, K.; Zhang, H.; Dolinoy, D.C.; Xi, C. Perinatal Lead Exposure Alters Gut Microbiota Composition and Results in Sex-specific Bodyweight Increases in Adult Mice. Toxicol. Sci. 2016, 151, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhou, C.; Guo, X.; Hu, G.; Li, G.; Zhuang, Y.; Cao, H.; Li, L.; Xing, C.; Zhang, C.; et al. Exposed to Mercury-Induced Oxidative Stress, Changes of Intestinal Microflora, and Association between them in Mice. Biol. Trace Element Res. 2020, 199, 1900–1907. [Google Scholar] [CrossRef]
- Breton, J.Ô.; Daniel, C.; Dewulf, J.; Pothion, S.; Froux, N.; Sauty, M.; Thomas, P.; Pot, B.; Foligne, B. Gut microbiota limits heavy metals burden caused by chronic oral exposure. Toxicol. Lett. 2013, 222, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chi, L.; Mahbub, R.; Bian, X.; Tu, P.; Ru, H.; Lu, K. Multi-Omics Reveals that Lead Exposure Disturbs Gut Microbiome Development, Key Metabolites, and Metabolic Pathways. Chem. Res. Toxicol. 2017, 30, 996–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ning, Z.; Mayne, J.; Moore, J.I.; Li, J.; Butcher, J.; Deeke, S.A.; Chen, R.; Chiang, C.K.; Wen, M.; et al. MetaPro-IQ: A universal metaproteomic approach to studying human and mouse gut microbiota. Microbiome 2016, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, Y.; Xia, Y.; Liu, K.; Ren, L.; Ji, Y. The Dysbiosis of Gut Microbiota Caused by Low-Dose Cadmium Aggravate the Injury of Mice Liver through Increasing Intestinal Permeability. Microorganisms 2020, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabec, J.; Wright, J.; Ly, T.; Wong, H.T.; McClimans, C.J.; Tokarev, V.; Lamendella, R.; Sherchand, S.; Shrestha, D.; Uprety, S.; et al. Arsenic disturbs the gut microbiome of individuals in a disadvantaged community in Nepal. Heliyon 2020, 6, e03313. [Google Scholar] [CrossRef] [Green Version]
- Shao, M.; Zhu, Y. Long-term metal exposure changes gut microbiota of residents surrounding a mining and smelting area. Sci. Rep. 2020, 10, 4453–4459. [Google Scholar] [CrossRef]
- Kish, L.; Hotte, N.; Kaplan, G.; Vincent, R.; Tso, R.; Gänzle, M.; Rioux, K.P.; Thiesen, A.; Barkema, H.; Wine, E.; et al. Environmental Particulate Matter Induces Murine Intestinal Inflammatory Responses and Alters the Gut Microbiome. PLoS ONE 2013, 8, e62220. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; He, F.; Liao, B.; Zhou, Y.; Li, B.; Ran, P. Exposure to ambient particulate matter alters the microbial composition and induces immune changes in rat lung. Respir. Res. 2017, 18, 143. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, E.A.; Comba, I.Y.; Cho, T.; Engen, P.A.; Yazıcı, C.; Soberanes, S.; Hamanaka, R.B.; Niğdelioğlu, R.; Meliton, A.Y.; Ghio, A.J.; et al. Inhalational exposure to particulate matter air pollution alters the composition of the gut microbiome. Environ. Pollut. 2018, 240, 817–830. [Google Scholar] [CrossRef]
- Li, R.; Yang, J.; Saffari, A.; Jacobs, J.; Baek, K.I.; Hough, G.; Larauche, M.H.; Ma, J.; Jen, N.; Moussaoui, N.; et al. Ambient Ultrafine Particle Ingestion Alters Gut Microbiota in Association with Increased Atherogenic Lipid Metabolites. Sci. Rep. 2017, 7, 42906. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Zhang, C.; Zhao, J.; Li, D.; Chen, J. Exposure to concentrated ambient PM2.5 (CAPM) induces intestinal disturbance via inflammation and alternation of gut microbiome. Environ. Int. 2022, 161, 107138. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, J.; Chen, M.; Huang, X.; Xie, X.; Li, W.; Cao, Q.; Kan, H.; Xu, Y.; Ying, Z. Exposure to concentrated ambient PM2.5 alters the composition of gut microbiota in a murine model. Part. Fibre Toxicol. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Chen, X.; Xu, Y.; Wu, W.; Tang, W.; Chen, Z.; Ji, G.; Peng, J.; Jiang, Q.; Xiao, J.; et al. Gut microbiota partially mediates the effects of fine particulate matter on type 2 diabetes: Evidence from a population-based epidemiological study. Environ. Int. 2019, 130, 104882. [Google Scholar] [CrossRef]
- Alderete, T.L.; Jones, R.B.; Chen, Z.; Kim, J.S.; Habre, R.; Lurmann, F.; Gilliland, F.D.; Goran, M.I. Exposure to traffic-related air pollution and the composition of the gut microbiota in overweight and obese adolescents. Environ. Res. 2017, 161, 472–478. [Google Scholar] [CrossRef]
- Liu, Y.; Yao, Y.; Li, H.; Qiao, F.; Wu, J.; Du, Z.-Y.; Zhang, M. Influence of Endogenous and Exogenous Estrogenic Endocrine on Intestinal Microbiota in Zebrafish. PLoS ONE 2016, 11, e0163895. [Google Scholar] [CrossRef] [PubMed]
- Javurek, A.B.; Spollen, W.G.; Johnson, S.A.; Bivens, N.J.; Bromert, K.H.; Givan, S.A.; Rosenfeld, C.S. Effects of exposure to bisphenol A and ethinyl estradiol on the gut microbiota of parents and their offspring in a rodent model. Gut Microbes 2016, 7, 471–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, D.; Zhang, H.; Jiang, X.; Zou, J.; Li, Q.; Mai, H.; Su, D.; Ling, W.; Feng, X. Bisphenol A exposure induces gut microbiota dysbiosis and consequent activation of gut-liver axis leading to hepatic steatosis in CD-1 mice. Environ. Pollut. 2020, 265, 114880. [Google Scholar] [CrossRef]
- Lai, K.-P.; Chung, Y.-T.; Li, R.; Wan, H.-T.; Wong, C.K.-C. Bisphenol A alters gut microbiome: Comparative metagenomics analysis. Environ. Pollut. 2016, 218, 923–930. [Google Scholar] [CrossRef]
- Wang, Y.; Rui, M.; Nie, Y.; Lu, G. Influence of gastrointestinal tract on metabolism of bisphenol A as determined by in vitro simulated system. J. Hazard. Mater. 2018, 355, 111–118. [Google Scholar] [CrossRef]
- Koestel, Z.L.; Backus, R.C.; Tsuruta, K.; Spollen, W.G.; Johnson, S.A.; Javurek, A.B.; Ellersieck, M.R.; Wiedmeyer, C.E.; Kannan, K.; Xue, J.; et al. Bisphenol A (BPA) in the serum of pet dogs following short-term consumption of canned dog food and potential health consequences of exposure to BPA. Sci. Total Environ. 2017, 579, 1804–1814. [Google Scholar] [CrossRef]
- Hu, J.; Raikhel, V.; Gopalakrishnan, K.; Fernandez-Hernandez, H.; Lambertini, L.; Manservisi, F.; Falcioni, L.; Bua, L.; Belpoggi, F.; Teitelbaum, S.L.; et al. Effect of postnatal low-dose exposure to environmental chemicals on the gut microbiome in a rodent model. Microbiome 2016, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Han, H.; Li, Y.; Xu, B.; Dai, W.; Zhang, Y.; Zhou, F.; Ma, H.; Pei, X. Di-(2-ethylhexyl) phthalate exposure induces female reproductive toxicity and alters the intestinal microbiota community structure and fecal metabolite profile in mice. Environ. Toxicol. 2021, 36, 1226–1242. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Zeng, Y.; Zhou, J.; Shu, R.; Xie, X.; Fu, Z. Exposure to dibutyl phthalate impairs lipid metabolism and causes inflammation via disturbing microbiota-related gut–liver axis. Acta Biochim. Biophys. Sin. 2020, 52, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Menon, R.; Manteiga, S.; Alden, N.; Hunt, C.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Environmental Chemical Diethylhexyl Phthalate Alters Intestinal Microbiota Community Structure and Metabolite Profile in Mice. mSystems 2019, 4, e00724-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, K.S. CDC Updates Guidelines for Children’s Lead Exposure. Environ. Health Perspect. 2012, 120, a268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchard, M.F.; Bellinger, D.C.; Weuve, J.; Matthews-Bellinger, J.; Gilman, S.; Wright, R.O.; Schwartz, J.; Weisskopf, M.G. Blood Lead Levels and Major Depressive Disorder, Panic Disorder, and Generalized Anxiety Disorder in US Young Adults. Arch. Gen. Psychiatry 2009, 66, 1313–1319. [Google Scholar] [CrossRef]
- Gilbert, S.G.; Weiss, B. A rationale for lowering the blood lead action level from 10 to 2 μg/dL. NeuroToxicology 2006, 27, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Bressler, J.; Kim, K.-A.; Chakraborti, T.; Goldstein, G. Molecular Mechanisms of Lead Neurotoxicity. Neurochem. Res. 1999, 24, 595–600. [Google Scholar] [CrossRef]
- Virgolini, M.B.; Chen, K.; Weston, D.D.; Bauter, M.R.; Cory-Slechta, D.A. Interactions of Chronic Lead Exposure and Intermittent Stress: Consequences for Brain Catecholamine Systems and Associated Behaviors and HPA Axis Function. Toxicol. Sci. 2005, 87, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Cory-Slechta, D.A.; Virgolini, M.B.; Rossi-George, A.; Thiruchelvam, M.; Lisek, R.; Weston, D. Lifetime Consequences of Combined Maternal Lead and Stress. Basic Clin. Pharmacol. Toxicol. 2008, 102, 218–227. [Google Scholar] [CrossRef]
- Lidsky, T.I.; Schneider, J.S. Lead neurotoxicity in children: Basic mechanisms and clinical correlates. Brain 2003, 126, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Fattal, O.; Budur, K.; Vaughan, A.J.; Franco, K. Review of the Literature on Major Mental Disorders in Adult Patients with Mitochondrial Diseases. Psychosomatics 2006, 47, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jou, S.-H.; Chiu, N.-Y.; Liu, C.-S. Mitochondrial dysfunction and psychiatric disorders. Chang Gung Med. J. 2009, 32, 370–379. [Google Scholar] [PubMed]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-Chain Amino Acid Supplementation Promotes Survival and Supports Cardiac and Skeletal Muscle Mitochondrial Biogenesis in Middle-Aged Mice. Cell Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Valerio, A.; D’Antona, G.; Nisoli, E. Branched-chain amino acids, mitochondrial biogenesis, and healthspan: An evolutionary perspective. Aging 2011, 3, 464. [Google Scholar] [CrossRef] [Green Version]
- Monachese, M.; Burton, J.P.; Reid, G. Bioremediation and tolerance of humans to heavy metals through microbial processes: A potential role for probiotics? Appl. Environ. Microbiol. 2012, 78, 6397–6404. [Google Scholar] [CrossRef] [Green Version]
- Counter, S.A.; Buchanan, L.H. Mercury exposure in children: A review. Toxicol. Appl. Pharmacol. 2004, 198, 209–230. [Google Scholar] [CrossRef]
- Huang, X.; Law, S.; Li, D.; Yu, X.; Li, B. Mercury Poisoning: A Case of a Complex Neuropsychiatric Illness. Am. J. Psychiatry 2014, 171, 1253–1256. [Google Scholar] [CrossRef]
- Ekino, S.; Susa, M.; Ninomiya, T.; Imamura, K.; Kitamura, T. Minamata disease revisited: An update on the acute and chronic manifestations of methyl mercury poisoning. J. Neurol. Sci. 2007, 262, 131–144. [Google Scholar] [CrossRef]
- Ratcliffe, H.E.; Swanson, G.M.; Fischer, L.J. Human Exposure to Mercury: A Critical Assessment of the Evidence of Adverse Health Effects. J. Toxicol. Environ. Health Part A 1996, 49, 221–270. [Google Scholar] [CrossRef]
- Ozuah, P.O. Mercury poisoning. Curr. Probl. Pediatr. 2000, 30, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Li, X.; dos Santos, A.P.M.; Farina, M.; da Rocha, J.B.T.; Avila, D.S.; Soldin, O.P.; Lu, R.; Aschner, M. Chapter 35—Mercury. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 451–459. [Google Scholar]
- Park, D.-J.; Zheng, W. Human exposure and health effects of inorganic and elemental mercury. J. Prev. Med. Public Health—Yebang Uihakhoe Chi 2012, 45, 344–352. [Google Scholar] [CrossRef]
- Lin, X.; Zhao, J.; Zhang, W.; He, L.; Wang, L.; Chang, D.; Cui, L.; Gao, Y.; Li, B.; Chen, C.; et al. Acute oral methylmercury exposure perturbs the gut microbiome and alters gut-brain axis related metabolites in rats. Ecotoxicol. Environ. Saf. 2020, 190, 110130. [Google Scholar] [CrossRef] [PubMed]
- Sarawagi, A.; Soni, N.D.; Patel, A.B. Glutamate and GABA Homeostasis and Neurometabolism in Major Depressive Disorder. Front. Psychiatry 2021, 12, 637863. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.M.; Zhang, Y.; Curran, T.E.; Magnuson, J.T.; Venables, B.J.; Durrer, K.E.; Allen, M.S.; Roberts, A.P. Alterations to the Intestinal Microbiome and Metabolome of Pimephales promelas and Mus musculus Following Exposure to Dietary Methylmercury. Environ. Sci. Technol. 2018, 52, 8774–8784. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Su, C.-T.; Shiue, H.-S.; Chen, W.-J.; Chen, Y.-H.; Choy, C.-S.; Chiou, H.-Y.; Han, B.-C.; Hsueh, Y.-M. The Methylation Capacity of Arsenic and Insulin Resistance are Associated with Psychological Characteristics in Children and Adolescents. Sci. Rep. 2017, 7, 3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karri, V.; Schuhmacher, M.; Kumar, V. Heavy metals (Pb, Cd, As and MeHg) as risk factors for cognitive dysfunction: A general review of metal mixture mechanism in brain. Environ. Toxicol. Pharmacol. 2016, 48, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Chi, L.; Bian, X.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The Effects of an Environmentally Relevant Level of Arsenic on the Gut Microbiome and Its Functional Metagenome. Toxicol. Sci. 2017, 160, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. S2), S1–S63. [Google Scholar] [CrossRef] [Green Version]
- Romero, E.S.; Cotoner, C.A.; Camacho, C.P.; Bedmar, M.C.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 108, 686–696. [Google Scholar]
- Wang, J.; Hu, W.; Yang, H.; Chen, F.; Shu, Y.; Zhang, G.; Liu, J.; Liu, Y.; Li, H.; Guo, L. Arsenic concentrations, diversity and co-occurrence patterns of bacterial and fungal communities in the feces of mice under sub-chronic arsenic exposure through food. Environ. Int. 2020, 138, 105600. [Google Scholar] [CrossRef]
- Forero-Rodríguez, L.J.; Josephs-Spaulding, J. Parkinson’s Disease and the Metal-Microbiome-Gut-Brain Axis: A Systems Toxicology Approach. Antioxidants 2021, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Phthalates Factsheet. Available online: https://www.cdc.gov/biomonitoring/Phthalates_FactSheet.html#:~:text=Phthalates%20are%20a%20group%20of,%2C%20shampoos%2C%20hair%20sprays (accessed on 5 April 2022).
- Przybylińska, P.A.; Wyszkowski, M. Environmental contamination with phthalates and its impact on living organisms. Ecol. Chem. Eng. S 2016, 23, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.N.; Yang, Y.C.S.; Lin, I.H.; Chen, Y.Y.; Lin, H.Y.; Wu, C.Y.; Su, Y.T.; Yang, Y.J.; Yang, S.N.; Suen, J.L. Phthalate exposure alters gut microbiota composition and IgM vaccine response in human newborns. Food Chem. Toxicol. 2019, 132, 110700. [Google Scholar] [CrossRef] [PubMed]
- Whyatt, R. Prenatal Exposure to Phthalates Linked to Decreased Mental and Motor Development. Available online: https://www.publichealth.columbia.edu/public-health-now/news/prenatal-exposure-phthalates-linked-decreased-mental-and-motor-development#:~:text=Among%20girls%2C%20one%20of%20the,somatic%20complaints%20and%20withdrawn%20behavior (accessed on 7 June 2022).
- Hu, D.; Wang, Y.-X.; Chen, W.-J.; Zhang, Y.; Li, H.-H.; Xiong, L.; Zhu, H.-P.; Chen, H.-Y.; Peng, S.-X.; Wan, Z.-H.; et al. Associations of phthalates exposure with attention deficits hyperactivity disorder: A case-control study among Chinese children. Environ. Pollut. 2017, 229, 375–385. [Google Scholar] [CrossRef]
- Ejaredar, M.; Nyanza, E.C.; Eycke, K.T.; Dewey, D. Phthalate exposure and childrens neurodevelopment: A systematic review. Environ. Res. 2015, 142, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-W.; Kim, M.-S.; Lim, Y.-H.; Lee, N.; Hong, Y.-C. Prenatal and postnatal exposure to di-(2-ethylhexyl) phthalate and neurodevelopmental outcomes: A systematic review and meta-analysis. Environ. Res. 2018, 167, 558–566. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, X.-Z.; Huang, X.; Wang, M.; Wu, J. The association between prenatal exposure to phthalates and cognition and neurobehavior of children-evidence from birth cohorts. NeuroToxicology 2019, 73, 199–212. [Google Scholar] [CrossRef]
- Lucaccioni, L.; Trevisani, V.; Passini, E.; Righi, B.; Plessi, C.; Predieri, B.; Iughetti, L. Perinatal Exposure to Phthalates: From Endocrine to Neurodevelopment Effects. Int. J. Mol. Sci. 2021, 22, 4063. [Google Scholar] [CrossRef]
- Xu, X.; Yang, Y.; Wang, R.; Wang, Y.; Ruan, Q.; Lu, Y. Perinatal exposure to di-(2-ethylhexyl) phthalate affects anxiety- and depression-like behaviors in mice. Chemosphere 2015, 124, 22–31. [Google Scholar] [CrossRef]
- Brown, J.S. Effects of Bisphenol-A and Other Endocrine Disruptors Compared with Abnormalities of Schizophrenia: An Endocrine-Disruption Theory of Schizophrenia. Schizophr. Bull. 2008, 35, 256–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.-C.; Chen, T.-J.; Lin, M.-L.; Jhong, Y.-C.; Chen, S.-C. Exercise prevents the increased anxiety-like behavior in lactational di-(2-ethylhexyl) phthalate-exposed female rats in late adolescence by improving the regulation of hypothalamus-pituitary-adrenal axis. Horm. Behav. 2014, 66, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Perera, F.; Nolte, E.L.R.; Wang, Y.; Margolis, A.E.; Calafat, A.M.; Wang, S.; Garcia, W.; Hoepner, L.A.; Peterson, B.S.; Rauh, V.; et al. Bisphenol A exposure and symptoms of anxiety and depression among inner city children at 10–12 years of age. Environ. Res. 2016, 151, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Environmental Health Sciences. Bisphenol A (BPA). Available online: https://www.niehs.nih.gov/health/topics/agents/sya-bpa/index.cfm (accessed on 26 November 2021).
- Malaisé, Y.; Menard, S.; Cartier, C.; Gaultier, E.; Lasserre, F.; Lencina, C.; Harkat, C.; Geoffre, N.; Lakhal, L.; Castan-Laurell, I.; et al. Gut dysbiosis and impairment of immune system homeostasis in perinatally-exposed mice to Bisphenol A precede obese phenotype development. Sci. Rep. 2017, 7, 14472. [Google Scholar] [CrossRef] [Green Version]
- Khokhlova, E.V.; Smeianov, V.V.; Efimov, B.A.; Kafarskaia, L.I.; Pavlova, S.I.; Shkoporov, A.N. Anti-inflammatory properties of intestinal Bifidobacterium strains isolated from healthy infants. Microbiol. Immunol. 2011, 56, 27–39. [Google Scholar] [CrossRef]
- Perera, F.; Vishnevetsky, J.; Herbstman, J.B.; Calafat, A.M.; Xiong, W.; Rauh, V.; Wang, S. Prenatal Bisphenol A Exposure and Child Behavior in an Inner-City Cohort. Environ. Health Perspect. 2012, 120, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Harley, K.G.; Gunier, R.B.; Kogut, K.; Johnson, C.; Bradman, A.; Calafat, A.M.; Eskenazi, B. Prenatal and early childhood bisphenol A concentrations and behavior in school-aged children. Environ. Res. 2013, 126, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Braun, J.M.; Kalkbrenner, A.E.; Calafat, A.M.; Yolton, K.; Ye, X.; Dietrich, K.N.; Lanphear, B.P. Impact of Early-Life Bisphenol A Exposure on Behavior and Executive Function in Children. Pediatrics 2011, 128, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Braun, J.M.; Yolton, K.; Dietrich, K.N.; Hornung, R.; Ye, X.; Calafat, A.M.; Lanphear, B.P. Prenatal bisphenol A exposure and early childhood behavior. Environ. Health Perspect. 2009, 117, 1945–1952. [Google Scholar] [CrossRef]
- Evans, S.F.; Kobrosly, R.W.; Barrett, E.S.; Thurston, S.W.; Calafat, A.M.; Weiss, B.; Stahlhut, R.; Yolton, K.; Swan, S.H. Prenatal bisphenol A exposure and maternally reported behavior in boys and girls. NeuroToxicology 2014, 45, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Roen, E.L.; Wang, Y.; Calafat, A.M.; Wang, S.; Margolis, A.; Herbstman, J.; Hoepner, L.A.; Rauh, V.; Perera, F.P. Bisphenol A exposure and behavioral problems among inner city children at 7–9 years of age. Environ. Res. 2015, 142, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, B.C.; Vandenbergh, J.G. Developmental exposure to environmental estrogens alters anxiety and spatial memory in female mice. Horm. Behav. 2006, 50, 85–93. [Google Scholar] [CrossRef]
- Xu, X.; Liu, Y.; Sadamatsu, M.; Tsutsumi, S.; Akaike, M.; Ushijima, H.; Kato, N. Perinatal bisphenol A affects the behavior and SRC-1 expression of male pups but does not influence on the thyroid hormone receptors and its responsive gene. Neurosci. Res. 2007, 58, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Cox, K.H.; Gatewood, J.D.; Howeth, C.; Rissman, E.F. Gestational exposure to bisphenol A and cross-fostering affect behaviors in juvenile mice. Horm. Behav. 2010, 58, 754–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patisaul, H.B.; Fortino, A.E.; Polston, E.K. Neonatal genistein or bisphenol-A exposure alters sexual differentiation of the AVPV. Neurotoxicol. Teratol. 2006, 28, 111–118. [Google Scholar] [CrossRef]
- Tian, Y.-H.; Baek, J.-H.; Lee, S.-Y.; Jang, C.-G. Prenatal and postnatal exposure to bisphenol a induces anxiolytic behaviors and cognitive deficits in mice. Synapse 2010, 64, 432–439. [Google Scholar] [CrossRef]
- Yu, C.; Tai, F.; Song, Z.; Wu, R.; Zhang, X.; He, F. Pubertal exposure to bisphenol A disrupts behavior in adult C57BL/6J mice. Environ. Toxicol. Pharmacol. 2011, 31, 88–99. [Google Scholar] [CrossRef]
- Xu, X.; Dong, F.; Yang, Y.; Wang, Y.; Wang, R.; Shen, X. Sex-specific effects of long-term exposure to bisphenol-A on anxiety- and depression-like behaviors in adult mice. Chemosphere 2015, 120, 258–266. [Google Scholar] [CrossRef]
- Luo, G.; Wang, S.; Li, Z.; Wei, R.; Zhang, L.; Liu, H.; Wang, C.; Niu, R.; Wang, J. Maternal Bisphenol A Diet Induces Anxiety-Like Behavior in Female Juvenile with Neuroimmune Activation. Toxicol. Sci. 2014, 140, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Prakash, A.; Tiwari, R. Environmental Endocrine-Disrupting Chemical Exposure: Role in Non-Communicable Diseases. Front. Public Health 2020, 8, 553850. [Google Scholar] [CrossRef]
- Chen, F.; Zhou, L.; Bai, Y.; Zhou, R.; Chen, L. Hypothalamic-pituitary-adrenal axis hyperactivity accounts for anxiety- and depression-like behaviors in rats perinatally exposed to bisphenol A. J. Biomed. Res. 2015, 29, 250–258. [Google Scholar] [PubMed] [Green Version]
- Panagiotidou, E.; Zerva, S.; Mitsiou, D.J.; Alexis, M.N.; Kitraki, E. Perinatal exposure to low-dose bisphenol A affects the neuroendocrine stress response in rats. J. Endocrinol. 2013, 220, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddivari, L.; Veeramachaneni, D.N.R.; Walters, W.A.; Lozupone, C.; Palmer, J.; Hewage, M.K.K.; Bhatnagar, R.; Amir, A.; Kennett, M.J.; Knight, R.; et al. Perinatal Bisphenol A Exposure Induces Chronic Inflammation in Rabbit Offspring via Modulation of Gut Bacteria and Their Metabolites. mSystems 2017, 2, e00093-17. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.; Zhao, Z.; Chen, S.; Zhang, L.; Wu, D.; Chen, Z. Bisphenol A suppresses proliferation and induces apoptosis in colonic epithelial cells through mitochondrial and MAPK/AKT pathways. Life Sci. 2018, 208, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Qu, W.; Wang, K.; Chen, S.; Zhang, L.; Wu, D.; Chen, Z. Bisphenol A inhibits mucin 2 secretion in intestinal goblet cells through mitochondrial dysfunction and oxidative stress. Biomed. Pharmacother. 2019, 111, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Hu, L.; Yang, S.; Ni, L.; Ma, L.; Zhao, Y.; Zheng, A.; Jin, Y.; Fu, Z. Bisphenol A impairs cognitive function and 5-HT metabolism in adult male mice by modulating the microbiota-gut-brain axis. Chemosphere 2021, 282, 130952. [Google Scholar] [CrossRef]
- Sun, Q.; Hong, X.; Wold, L.E. Cardiovascular Effects of Ambient Particulate Air Pollution Exposure. Circulation 2010, 121, 2755–2765. [Google Scholar] [CrossRef]
- MohanKumar, S.M.; Campbell, A.; Block, M.; Veronesi, B. Particulate matter, oxidative stress and neurotoxicity. Neurotoxicology 2008, 29, 479–488. [Google Scholar] [CrossRef]
- Szyszkowicz, M.; Rowe, B.H.; Colman, I. Air pollution and daily emergency department visits for depression. Int. J. Occup. Med. Environ. Health 2009, 22, 355–362. [Google Scholar] [CrossRef]
- Chen, S.; Oliva, P.; Zhang, P. Air Pollution and Mental Health: Evidence from China; National Bureau of Economic Research, Inc.: Cambridge, MA, USA, 2018. [Google Scholar]
- Bakolis, I.; Hammoud, R.; Stewart, R.; Beevers, S.; Dajnak, D.; MacCrimmon, S.; Broadbent, M.; Pritchard, M.; Shiode, N.; Fecht, D.; et al. Mental health consequences of urban air pollution: Prospective population-based longitudinal survey. Soc. Psychiatry 2020, 56, 1587–1599. [Google Scholar] [CrossRef]
- Lim, Y.-H.; Kim, H.; Kim, J.H.; Bae, S.; Park, H.Y.; Hong, Y.-C. Air Pollution and Symptoms of Depression in Elderly Adults. Environ. Health Perspect. 2012, 120, 1023–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vert, C.; Sánchez-Benavides, G.; Martínez, D.; Gotsens, X.; Gramunt, N.; Cirach, M.; Molinuevo, J.L.; Sunyer, J.; Nieuwenhuijsen, M.J.; Crous-Bou, M.; et al. Effect of long-term exposure to air pollution on anxiety and depression in adults: A cross-sectional study. Int. J. Hyg. Environ. Health 2017, 220, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Borroni, E.; Pesatori, A.C.; Bollati, V.; Buoli, M.; Carugno, M. Air pollution exposure and depression: A comprehensive updated systematic review and meta-analysis. Environ. Pollut. 2021, 292, 118245. [Google Scholar] [CrossRef] [PubMed]
- SShin, J.; Park, J.Y.; Choi, J. Long-term exposure to ambient air pollutants and mental health status: A nationwide population-based cross-sectional study. PLoS ONE 2018, 13, e0195607. [Google Scholar]
- Bakian, A.V.; Huber, R.S.; Coon, H.; Gray, D.; Wilson, P.; McMahon, W.M.; Renshaw, P.F. Acute Air Pollution Exposure and Risk of Suicide Completion. Am. J. Epidemiol. 2015, 181, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Jung, S.H.; Kang, D.R.; Kim, H.C.; Moon, K.T.; Hur, N.W.; Shin, D.C.; Suh, I. Ambient Particulate Matter as a Risk Factor for Suicide. Am. J. Psychiatry 2010, 167, 1100–1107. [Google Scholar] [CrossRef]
- Perera, F.P.; Tang, D.; Wang, S.; Vishnevetsky, J.; Zhang, B.; Diaz, D.; Camann, D.; Rauh, V. Prenatal Polycyclic Aromatic Hydrocarbon (PAH) Exposure and Child Behavior at Age 6–7 Years. Environ. Health Perspect. 2012, 120, 921–926. [Google Scholar] [CrossRef] [Green Version]
- Power, M.C.; Kioumourtzoglou, M.-A.; Hart, J.E.; Okereke, O.I.; Laden, F.; Weisskopf, M.G. The relation between past exposure to fine particulate air pollution and prevalent anxiety: Observational cohort study. BMJ 2015, 350, h1111. [Google Scholar] [CrossRef] [Green Version]
- Forns, J.; Dadvand, P.; Foraster, M.; Alvarez-Pedrerol, M.; Rivas, I.; López-Vicente, M.; Suades-Gonzalez, E.; Garcia-Esteban, R.; Esnaola, M.; Cirach, M.; et al. Traffic-Related Air Pollution, Noise at School, and Behavioral Problems in Barcelona Schoolchildren: A Cross-Sectional Study. Environ. Health Perspect. 2016, 124, 529–535. [Google Scholar] [CrossRef]
- Yorifuji, T.; Kashima, S.; Diez, M.H.; Kado, Y.; Sanada, S.; Doi, H. Prenatal exposure to outdoor air pollution and child behavioral problems at school age in Japan. Environ. Int. 2017, 99, 192–198. [Google Scholar] [CrossRef]
- Campbell, A.; Oldham, M.; Becaria, A.; Bondy, S.; Meacher, D.; Sioutas, C.; Misra, C.; Mendez, L.; Kleinman, M. Particulate Matter in Polluted Air May Increase Biomarkers of Inflammation in Mouse Brain. NeuroToxicology 2005, 26, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Chen, L.; Guo, Z.; Sang, N.; Li, G. In vivo screening to determine neurological hazards of nitrogen dioxide (NO2) using Wistar rats. J. Hazard. Mater. 2012, 225-226, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Yue, H.; Yun, Y.; Sang, N. Chronic SO2 inhalation above environmental standard impairs neuronal behavior and represses glutamate receptor gene expression and memory-related kinase activation via neuroinflammation in rats. Environ. Res. 2015, 137, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Coccaro, E.F.; Lee, R.; Coussons-Read, M. Elevated Plasma Inflammatory Markers in Individuals with Intermittent Explosive Disorder and Correlation with Aggression in Humans. JAMA Psychiatry 2014, 71, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 2015, 16, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and psychiatric illness. J. Neuroinflamm. 2013, 10, 816. [Google Scholar] [CrossRef] [Green Version]
- Guxens, M.; Sunyer, J. A review of epidemiological studies on neuropsychological effects of air pollution. Swiss Med. Wkly. 2012, 141, w13322. [Google Scholar]
- Calderón-Garcidueñas, L.; Solt, A.C.; Henríquez-Roldán, C.; Torres-Jardón, R.; Nuse, B.; Herritt, L.; Villarreal-Calderón, R.; Osnaya, N.; Stone, I.; García, R.; et al. Long-term air pollution exposure is associated with neuroinflammation, an altered innate immune response, disruption of the blood-brain barrier, ultrafine particulate deposition, and accumulation of amyloid β-42 and α-synuclein in children and young adults. Toxicol. Pathol. 2008, 36, 289–310. [Google Scholar]
- Roberts, S.; Arseneault, L.; Barratt, B.; Beevers, S.; Danese, A.; Odgers, C.; Moffitt, T.; Reuben, A.; Kelly, F.J.; Fisher, H.L. Exploration of NO2 and PM2.5 air pollution and mental health problems using high-resolution data in London-based children from a UK longitudinal cohort study. Psychiatry Res. 2019, 272, 8–17. [Google Scholar] [CrossRef]
- Block, L.M.; Calderón-Garcidueñas, L. Air pollution: Mechanisms of neuroinflammation and CNS disease. Trends Neurosci. 2009, 32, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Salim, Y.S.; Kaplan, G.G.; Madsen, K.L. Air pollution effects on the gut microbiota: A link between exposure and inflammatory disease. Gut Microbes 2014, 5, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Mutlu, E.A.; Engen, P.; Soberanes, S.; Urich, D.; Forsyth, C.B.; Nigdelioglu, R.; Chiarella, S.E.; Radigan, K.A.; Gonzalez, A.; Jakate, S.; et al. Particulate matter air pollution causes oxidant-mediated increase in gut permeability in mice. Part. Fibre Toxicol. 2011, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, E.M. Air Pollution, Stress, and Allostatic Load: Linking Systemic and Central Nervous System Impacts. J. Alzheimer’s Dis. 2019, 69, 597–614. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Niu, Y.; Cai, J.; Liu, C.; Meng, X.; Chen, R.; Kan, H. Personal ozone exposure and stress hormones in the hypothalamus–pituitary–adrenal and sympathetic-adrenal-medullary axes. Environ. Int. 2021, 159, 107050. [Google Scholar] [CrossRef]
- Hughes, D.T.; Clarke, M.B.; Yamamoto, K.; Rasko, D.A.; Sperandio, V. The QseC Adrenergic Signaling Cascade in Enterohemorrhagic E. coli (EHEC). PLOS Pathog. 2009, 5, e1000553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouladi, F.; Bailey, M.J.; Patterson, W.B.; Sioda, M.; Blakley, I.C.; Fodor, A.A.; Jones, R.B.; Chen, Z.; Kim, J.S.; Lurmann, F.; et al. Air pollution exposure is associated with the gut microbiome as revealed by shotgun metagenomic sequencing. Environ. Int. 2020, 138, 105604. [Google Scholar] [CrossRef]
- Li, T.; Fang, J.; Tang, S.; Du, H.; Zhao, L.; Wang, Y.; Deng, F.; Liu, Y.; Du, Y.; Cui, L.; et al. PM2.5 exposure associated with microbiota gut-brain axis: Multi-omics mechanistic implications from the BAPE study. Innovation 2022, 3, 100213. [Google Scholar] [CrossRef]
- World Health Organization. AIR POLLUTION. Available online: https://www.who.int/health-topics/air-pollution#tab=tab_1 (accessed on 22 September 2021).
- Allen, A.P.; Hutch, W.; Borre, Y.E.; Kennedy, P.J.; Temko, A.; Boylan, G.; Murphy, E.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Bifidobacterium longum 1714 as a translational psychobiotic: Modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 2016, 6, e939. [Google Scholar] [CrossRef] [Green Version]
- Benton, D.; Williams, C.; Brown, A. Impact of consuming a milk drink containing a probiotic on mood and cognition. Eur. J. Clin. Nutr. 2006, 61, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Patterson, E.; Griffin, S.M.; Ibarra, A.; Ellsiepen, E.; Hellhammer, J. Lacticaseibacillus paracasei Lpc-37® improves psychological and physiological markers of stress and anxiety in healthy adults: A randomized, double-blind, placebo-controlled and parallel clinical trial (the Sisu study). Neurobiol. Stress 2020, 13, 100277. [Google Scholar] [CrossRef]
- Wu, S.-I.; Wu, C.-C.; Tsai, P.-J.; Cheng, L.-H.; Hsu, C.-C.; Shan, I.-K.; Chan, P.-Y.; Lin, T.-W.; Ko, C.-J.; Chen, W.-L.; et al. Psychobiotic Supplementation of PS128TM Improves Stress, Anxiety, and Insomnia in Highly Stressed Information Technology Specialists: A Pilot Study. Front. Nutr. 2021, 8, 614105. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, N.; Milesi, C.; Burn, O.; van den Bogert, B.; Nauta, A.; Hart, K.; Sowden, P.; Burnet, P.W.J.; Kadosh, K.C. Anxiolytic effects of a galacto-oligosaccharides prebiotic in healthy females (18–25 years) with corresponding changes in gut bacterial composition. Sci. Rep. 2021, 11, 8302. [Google Scholar] [CrossRef] [PubMed]
- Donoso, F.; Egerton, S.; Bastiaanssen, T.F.; Fitzgerald, P.; Gite, S.; Fouhy, F.; Ross, R.P.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Polyphenols selectively reverse early-life stress-induced behavioural, neurochemical and microbiota changes in the rat. Psychoneuroendocrinology 2020, 116, 104673. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; Rajabi, S.; Mohammadshahi, M. Effect of synbiotic and probiotic supplementation on serum brain-derived neurotrophic factor level, depression and anxiety symptoms in hemodialysis patients:A randomized, double-blinded, clinical trial. Nutr. Neurosci. 2019, 24, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Moludi, J.; Khedmatgozar, H.; Nachvak, S.M.; Abdollahzad, H.; Moradinazar, M.; Tabaei, A.S. The effects of co-administration of probiotics and prebiotics on chronic inflammation, and depression symptoms in patients with coronary artery diseases: A randomized clinical trial. Nutr. Neurosci. 2021, 24, 1–10. [Google Scholar] [CrossRef]
- Quero, C.; Manonelles, P.; Fernández, M.; Abellán-Aynés, O.; López-Plaza, D.; Andreu-Caravaca, L.; Hinchado, M.; Gálvez, I.; Ortega, E. Differential Health Effects on Inflammatory, Immunological and Stress Parameters in Professional Soccer Players and Sedentary Individuals after Consuming a Synbiotic. A Triple-Blinded, Randomized, Placebo-Controlled Pilot Study. Nutrients 2021, 13, 1321. [Google Scholar] [CrossRef]
- Wei, C.-L.; Wang, S.; Yen, J.-T.; Cheng, Y.-F.; Liao, C.-L.; Hsu, C.-C.; Wu, C.-C.; Tsai, Y.-C. Antidepressant-like activities of live and heat-killed Lactobacillus paracasei PS23 in chronic corticosterone-treated mice and possible mechanisms. Brain Res. 2019, 1711, 202–213. [Google Scholar] [CrossRef]
- Nishida, K.; Sawada, D.; Kuwano, Y.; Tanaka, H.; Rokutan, K. Health Benefits of Lactobacillus gasseri CP2305 Tablets in Young Adults Exposed to Chronic Stress: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2019, 11, 1859. [Google Scholar] [CrossRef] [Green Version]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus plantarum 299v decreases kynurenine concentration and improves cognitive functions in patients with major depression: A double-blind, randomized, placebo controlled study. Psychoneuroendocrinology 2018, 100, 213–222. [Google Scholar] [CrossRef]
- Lew, L.-C.; Hor, Y.-Y.; Yusoff, N.A.A.; Choi, S.-B.; Yusoff, M.S.; Roslan, N.S.; Ahmad, A.; Mohammad, J.A.; Abdullah, M.F.I.; Zakaria, N.; et al. Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study. Clin. Nutr. 2018, 38, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, L.; Sellaro, R.; van Hemert, S.; Bosch, J.A.; Colzato, L.S. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain Behav. Immun. 2015, 48, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillisch, K.; Labus, J.; Kilpatrick, L.; Jiang, Z.; Stains, J.; Ebrat, B.; Guyonnet, D.; Legrain-Raspaud, S.; Trotin, B.; Naliboff, B.; et al. Consumption of Fermented Milk Product with Probiotic Modulates Brain Activity. Gastroenterology 2013, 144, 1394–1401.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messaoudi, M.; Violle, N.; Bisson, J.-F.; Desor, D.; Javelot, H.; Rougeot, C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes 2011, 2, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.-H.; Chuang, H.-L.; Huang, Y.-T.; Wu, C.-C.; Chou, G.-T.; Wang, S.; Tsai, Y.-C. Alteration of behavior and monoamine levels attributable to Lactobacillus plantarum PS128 in germ-free mice. Behav. Brain Res. 2015, 298, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Liu, W.-H.; Wu, C.-C.; Juan, Y.-C.; Wu, Y.-C.; Tsai, H.-P.; Wang, S.; Tsai, Y.-C. Psychotropic effects of Lactobacillus plantarum PS128 in early life-stressed and naïve adult mice. Brain Res. 2016, 1631, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Agusti, A.; Moya-Pérez, A.; Campillo, I.; La Paz, S.M.-D.; Cerrudo, V.; Perez-Villalba, A.; Sanz, Y. Bifidobacterium pseudocatenulatum CECT 7765 Ameliorates Neuroendocrine Alterations Associated with an Exaggerated Stress Response and Anhedonia in Obese Mice. Mol. Neurobiol. 2017, 55, 5337–5352. [Google Scholar] [CrossRef]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Jin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef]
- Wang, T.; Hu, X.; Liang, S.; Li, W.; Wu, X.; Wang, L.; Jin, F. Lactobacillus fermentum NS9 restores the antibiotic induced physiological and psychological abnormalities in rats. Benef. Microbes 2015, 6, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.J.; Emge, J.R.; Berzins, K.; Lung, L.; Khamishon, R.; Shah, P.; Rodrigues, D.M.; Sousa, A.J.; Reardon, C.; Sherman, P.; et al. Probiotics normalize the gut-brain-microbiota axis in immunodeficient mice. Am. J. Physiol. Liver Physiol. 2014, 307, G793–G802. [Google Scholar] [CrossRef]
- Luo, J.; Wang, T.; Liang, S.; Hu, X.; Li, W.; Jin, F. Ingestion of Lactobacillus strain reduces anxiety and improves cognitive function in the hyperammonemia rat. Sci. China Life Sci. 2014, 57, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-Y.; Chen, L.-H.; Wang, M.-F.; Hsu, C.-C.; Chan, C.-H.; Li, J.-X.; Huang, H.-Y. Lactobacillus paracasei PS23 Delays Progression of Age-Related Cognitive Decline in Senescence Accelerated Mouse Prone 8 (SAMP8) Mice. Nutrients 2018, 10, 894. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Hsu, C.; Chou, G.; Hsu, J.; Liong, M.; Tsai, Y. Lactobacillus paracasei PS23 reduced early-life stress abnormalities in maternal separation mouse model. Benef. Microbes 2019, 10, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savignac, H.M.; Kiely, B.; Dinan, T.G.; Cryan, J.F. Bifidobacteria exert strain-specific effects on stress-related behavior and physiology in BALB/c mice. Neurogastroenterol. Motil. 2014, 26, 1615–1627. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D.; et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. 2011, 23, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Tarr, A.J.; Galley, J.D.; Fisher, S.E.; Chichlowski, M.; Berg, B.M.; Bailey, M.T. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: Evidence for effects on the gut–brain axis. Brain Behav. Immun. 2015, 50, 166–177. [Google Scholar] [CrossRef] [Green Version]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W. Psychobiotics and the Manipulation of Bacteria-Gut-Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Bienenstock, J.; Dinan, T.G. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatr. Res. 2008, 43, 164–174. [Google Scholar] [CrossRef]
- Soderholm, J.D.; Perdue, M.H., II. Stress and intestinal barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G7–G13. [Google Scholar] [CrossRef] [PubMed]
- De Vries, H.E.; Blom-Roosemalen, M.C.; van Oosten, M.; de Boer, A.G.; van Berkel, T.J.; Breimer, D.D.; Kuiper, J. The influence of cytokines on the integrity of the blood-brain barrier in vitro. J. Neuroimmunol. 1996, 64, 37–43. [Google Scholar] [CrossRef]
- Banks, W.A.; Erickson, M.A. The blood–brain barrier and immune function and dysfunction. Neurobiol. Dis. 2010, 37, 26–32. [Google Scholar] [CrossRef]
- Ek, M.; Engblom, D.; Saha, S.; Blomqvist, A.; Jakobsson, P.J.; Ericsson-Dahlstrand, A. Pathway across the blood–brain barrier. Nature 2001, 410, 430–431. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Mohanty, D. Psychobiotics: A new approach for treating mental illness? Crit. Rev. Food Sci. Nutr. 2019, 59, 1230–1236. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study Model | Dosing Regimen | Impact on the Gut Microbiome | Reference |
|---|---|---|---|
| Heavy Metals | |||
| Six-week-old female C57Bl/6 mice | Mice were treated with 10 ppm as in the drinking water for 4 weeks |
| Lu et al. [62] |
| Wild-type and IL10−/− mice | Mice were treated with 10 ppm as in the drinking water for 4 weeks |
| Lu et al. [63] |
| Five-week-old ICR mice | Mice were treated with as (3 mg/L), Fe (5 mg/L), or in combination in drinking water, for 90 days |
| Guo et al. [64] |
| Six- to eight-week-old C57Bl/6 Tac male mice | Mice exposed for 2, 5, or 10 weeks to 0, 10, or 250 ppb arsenite (As (III)) |
| Dheer et al. [65] |
| C57/BL6 male and female mice | Mice were treated with 10 ppm as in the drinking water for 4 weeks. |
| Chi et al. [66] |
| Non-agouti (a/a) offspring | Mice exposed from gestation through lactation to Pb (32 ppm in the drinking water) |
| Wu et al. [67] |
| Kunming mice | Exposed to 80 mg/L HgCl2 in drinking water for 90 days |
| Zhao et al. [68] |
| Six-week-old Balb/C female mice | Mice were exposed to lead (PbCl2, 100 or 500 ppm- mg/L) or cadmium (CdCl2, 20 or 100 ppm-mg/L) in the drinking water for 8 weeks |
| Breton et al. [69] |
| Adult C57Bl/6 female mice | Mice were treated with 10 ppm PbCl2 in the drinking water for 13 weeks for a concentration of ~2 mg/kg body weight/day |
| Gao et al. [70] |
| Mongolian toads (Buforaddei) | One group lives in a heavy-metal-polluted area (Baiyin-BY) and the other resides in a relatively unpolluted area (Liujiaxia-LJX) |
| Zhang et al. [71] |
| Six-week old Female C57BL/6J mice | The low Cd treatment group received drinking water containing 10 mg/L CdCl2, whereas the control group received pure drinking water. The third group was given drinking water containing 10 mg/L CdCl2 and an antibiotic combination for 52 weeks |
| Liu et al. [72] |
| Healthy members of two separate communities (Mahuawa and Ghanashyampur) in southern Nepal | Consumption of As-contaminated well water |
| Brabec et al. [73] |
| Healthy volunteers from two different villages in China | Long-term exposure to multiple metals, including As, Cd, Cu, Pb and Zn |
| Shao et al. [74] |
| Particulate Matter | |||
| Wild-type (WT) 129/SvEv mice, IL10 (−/−) deficient mice | Mice were orally gavaged with Ottawa urban PM10 (EHC-93: 18 μg/g/day) for 7 or 14 days. To evaluate long-term effects of exposure, IL10 deficient (−/−) mice were subjected to the same treatment for 35 days |
| Kish et al. [75] |
| Male Sprague-Dawley rats | Exposed to clean air, and PM that are BMF, or MVE for 4, 12 and 24 weeks |
| Li et al. [76] |
| C57BL/6 mice | Exposed via inhalation to either concentrated ambient particles (PM2.5) or filtered air for 8 h per day, 5 days a week, for a total of 3 weeks |
| Mutlu et al. [77] |
| Low-density lipoprotein receptor-null (Ldlr−/−) mice | Mice on a high-fat diet were orally administered with vehicle control or UFP (40 μg/mouse/day) 3 days a week for 10 weeks |
| Li et al. [78] |
| C57BL/6J male mice | Mice were exposed in filtered air or CAPM2.5 chambers for 8, 16 and 24 weeks |
| Xie et al. [79] |
| C57Bl/6J mice | Exposed to filtered air (FA) or concentrated ambient PM2.5 (CAP) for 12 months |
| Wang et al. [80] |
| Adult humans aged 18 years or older from 14 randomly selected districts in southern China | Exposed to PMs of different sizes (PM2.5 and PM1)—air pollution |
| Liu et al. [81] |
| Adolescents and young adults from Southern California | Exposed to traffic-related air pollution |
| Alderete et al. [82] |
| Endocrine Disrupting Chemicals (EDCs) | |||
| Adult male zebrafish | Zebrafish were exposed to BPA (200 or 2000 μg/L) or E2 (500 ng/L or 2000 ng/L) for 5 weeks |
| Liu et al. [83] |
| Adult male and female P0 California mice (Peromyscus californicus); Juvenile (PND30) male and female California mice offspring | Mice were exposed to BPA (50 mg/kg feed weight), 2 weeks prior to mating EE (0.1 ppb), or a control diet, and then continued on the diets throughout gestation and lactation. After pairing reproductive male partners were exposed to these diets until their offspring were weaned at PND30 |
| Javurek et al. [84] |
| Male CD-1 mice | 0.5 mg/kg of BPA for 24 weeks |
| Feng et al. [85] |
| 20 mg/10 g body weight BPA for 10 weeks |
| Lai et al. [86] | |
| HepG2 (Human) | 25 μg/L, 250 μg/L and 2500 μg/L BPA for 10 days |
| Wang et al. [87] |
| Adult gonadectomized male and female dogs (Canisfamiliaris) | Male and female dogs who were shifted from dry dog food to one of two brаnds of сommerсiаlly саnned dog food for two weeks hаd а neаrly three-fold rise in circulating BPA concentrations. |
| Koestel et al. [88] |
| Sprague-Dawley female rats | Exposed to DEP—0.1735 mg/kg body weight), MPB—0.1050 mg/kg body weight, TCS—0.05 mg/kg body weight or a combination of these chemicals from birth to adulthood |
| Hu et al. [89] |
| Four-week-old ICR mice | Mice were intragastrically administered 500 and 1500 mg/kg body weight per day DEHP (mixed with corn oil) for 30 days |
| Fu et al. [90] |
| Six-week-old C57BL/6J mice | Oral gavage was used to administer 10-week experimental cycles of the vehicle or DBP (0.1 and 1 mg/kg) to 6-week-old C57BL/6J mice |
| Xiong et al. [91] |
| Anaerobic culture of cecal microbiota of mice | 10 and 100 μM DEHP for seven days |
| Lei et al. [92] |
| Female C57BL/6 mice | 1 and 10 mg/kg body weight/day DEHP for 14 days |
| |
| Study Model (Human) | Psychobiotics, Route of Administration and Dosage | Duration of Intervention | Observations | References |
|---|---|---|---|---|
| Healthy male volunteers between 18–40 years of age | B. longum 1714 109 CFU/day | 4 weeks |
| Allen et al. [192] |
| Major depressive disorder patients drobiotic N = 40, Placebo N = 39 | Probiotic bacteria Lactobacillus plantarum 299v—2 capsules a day (1 capsule = 10 × 109 CFU) | 8 weeks |
| Rudzki et al. [204] |
| Stressed adults with a mean age of 31.7 ± 11.1 years old (P8 N = 52, placebo N = 51) | Probiotic (Lactobacillus plantarum P8; 10 log CFU daily) | 12 weeks |
| Lew et al. [205] |
| Human elderly volunteers, mean age 61.8 years | A mixture of Lactobacillus casei Shirota | 3 weeks |
| Benton et al. [193] |
| Healthy human young adults | Bifidobacteriumbifidum W23, Bifidobacteriumlactis W52, Lactobacillus acidophilus W37, Lactobacillus brevis W63, Lactobacillus casei W56, Lactobacillus salivarius W24, and Lactococcus lactis (W19 and W58) | 4 weeks |
| Steenbergen et al. [206] |
| Healthy women | A mixture of Bifidobacterium animalis subsp. lactis, Streptococcus thermophilus, Lactobacillus bulgaricus and Lactococcuslactis subsp. Lactis | 4 weeks |
| Tillisch et al. [207] |
| Healthy human adults | A mixture of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 3 × 109 CFU/stick/day | 30 days |
| Messaoudi et al. [208] |
| Healthy adults (18–45 years) | 1.75 × 1010 CFU Lacticaseibacillus paracasei Lpc-37 | 5 weeks |
| Patterson et al. [194] |
| IT specialists | 2 × 1010 Lactobacillus plantarum PS128 | 8 weeks |
| Wu et al. [195] |
| Healthy female volunteers (aged 18–25 years) | A daily dose of 7.5 g of the prebiotic galactooligosaccharides | 4 weeks |
| Johnstone et al. [197] |
| Hemodialysis patients | Synbiotic (15 g of prebiotics, 5 g of probiotic containing Lactobacillus acidophilus T16, Bifidobacterium bifidum BIA-6, Bifidobacterium lactis BIA-7 and Bifidobacterium longum BIA-8 (2.7 × 107 CFU/g each)) | 12 weeks |
| Haghighat et al. [199] |
| Coronary artery disease (CAD). | Lactobacillus rhamnosus G (capsule/day, contained 1.9 × 109 CFU) and inulin (15 g/day) | 8 weeks |
| Moludi et al. [200] |
| Professional soccer players and sedentary individuals | Synbiotic Gasteel Plus®® containing probiotic strains, such as Bifidobacterium lactis CBP-001010, Lactobacillus rhamnosus CNCM I-4036 and Bifidobacterium longum ES1 (≥1 × 109 CFU, as well as the prebiotic FOS (200 mg)) | 1 month |
| Quero et al. [201] |
| Healthy young adults | Heat-inactivated, washed Lactobacillus gasseri CP2305 (CP2305) | 24 weeks |
| Nishida et al. [203] |
| Study Model (Animal) | Psychobiotics, Route of Administration and Dosage | Duration of Intervention | Observations | References |
|---|---|---|---|---|
| Germ-free mice | Heat killed or live L. plantarum PS128 109 CFU/mouse/day by gavage | 16 days | Heat killed: NA Live:
| Liu et al. [209] |
| Early life stress (ELS) mice | L. plantarum PS128 109 CFU/mouse/day by gavage | 16 days |
| Liu et al. [210] |
| Adult male wild-type C57BL-6 | 1 × 109 CFU B. pseudocatenulatum CECT 7765 by gavage | 13 weeks |
| Agusti et al. [211] |
| Male SPF CRS rats | L. helveticus NS8 109 CFU/mL in drinking water | 21 days |
| Liang S et al. [212] |
| Ampicillin-treated male Sprague-Dawley rats (Rattus norvegicus) | L. fermentum strain NS9 109 CFU/mL in drinking water | 41 days |
| Wang et al. [213] |
| RagI−/− mice | L. rhamnosus R0011 + L. helveticus R0052 6 × 109 CFU | 28 days |
| Smith et al. [214] |
| Hyperammonemia rats | L. helveticus NS8 10 9 CFU | 14 days |
| Luo et al. [215] |
| Male and female senescence-accelerated mouse prone 8 (SAMP8) mice | Lactobacillus paracasei PS23 (LPPS23) 109 CFU/mouse/day | 12 weeks |
| Huang et al. [216] |
| Maternal Separation (MS) C57BL/6Jmice neonates | Live and heat-killed Lactobacillus paracasei PS23 (PS23) 109 CFU/mouse/day by oral gavage | 4 weeks |
| Liao et al. [217] |
| Male BALB/c mice | L. rhamnosus (JB-1) 109 CFU/mouse/day by gavage | 28 days |
| Bravo et al. [218] |
| Male BALB/c mice | B. longum 1714 or B. breve 1205 109 CFU/day by gavage | 21–41 days |
| Savignac et al. [219] |
| Chronic colitis mice | B. longum NCC3001 1010 CFU | 14 days |
| Bercik et al. [220] |
| Six–eight-week-old male C57/BL6 mice | Prebiotics: human milk oligosaccharides 3′Sialyllactose (3′SL) or 6′Sialyllactose (6′SL) | 2 weeks |
| Tarr et al. [221] |
| C57BL/6J male mice | Prebiotics: Fructooligosaccharides (FOS) and Galactooligosaccharides (GOS) or a combination of FOS + GOS (dissolved in drinking water for 0.3–0.4 g/mouse/day) | 3 weeks |
| Burokas et al. [222] |
| Maternal separation (MS) rat model | Naturally-derived polyphenols xanthohumol and quercetin | 8 weeks |
| Donoso et al. [198] |
| Mice | Live or heat-killed Lactobacillus paracasei PS23 | 42 days |
| Wei et al. [202] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Sharma, P.; Pal, N.; Kumawat, M.; Shubham, S.; Sarma, D.K.; Tiwari, R.R.; Kumar, M.; Nagpal, R. Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut–Brain Axis. Microorganisms 2022, 10, 1457. https://doi.org/10.3390/microorganisms10071457
Singh S, Sharma P, Pal N, Kumawat M, Shubham S, Sarma DK, Tiwari RR, Kumar M, Nagpal R. Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut–Brain Axis. Microorganisms. 2022; 10(7):1457. https://doi.org/10.3390/microorganisms10071457
Chicago/Turabian StyleSingh, Samradhi, Poonam Sharma, Namrata Pal, Manoj Kumawat, Swasti Shubham, Devojit Kumar Sarma, Rajnarayan R. Tiwari, Manoj Kumar, and Ravinder Nagpal. 2022. "Impact of Environmental Pollutants on Gut Microbiome and Mental Health via the Gut–Brain Axis" Microorganisms 10, no. 7: 1457. https://doi.org/10.3390/microorganisms10071457