
Bacillales: From Taxonomy to Biotechnological and Industrial Perspectives

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Order Bacillales
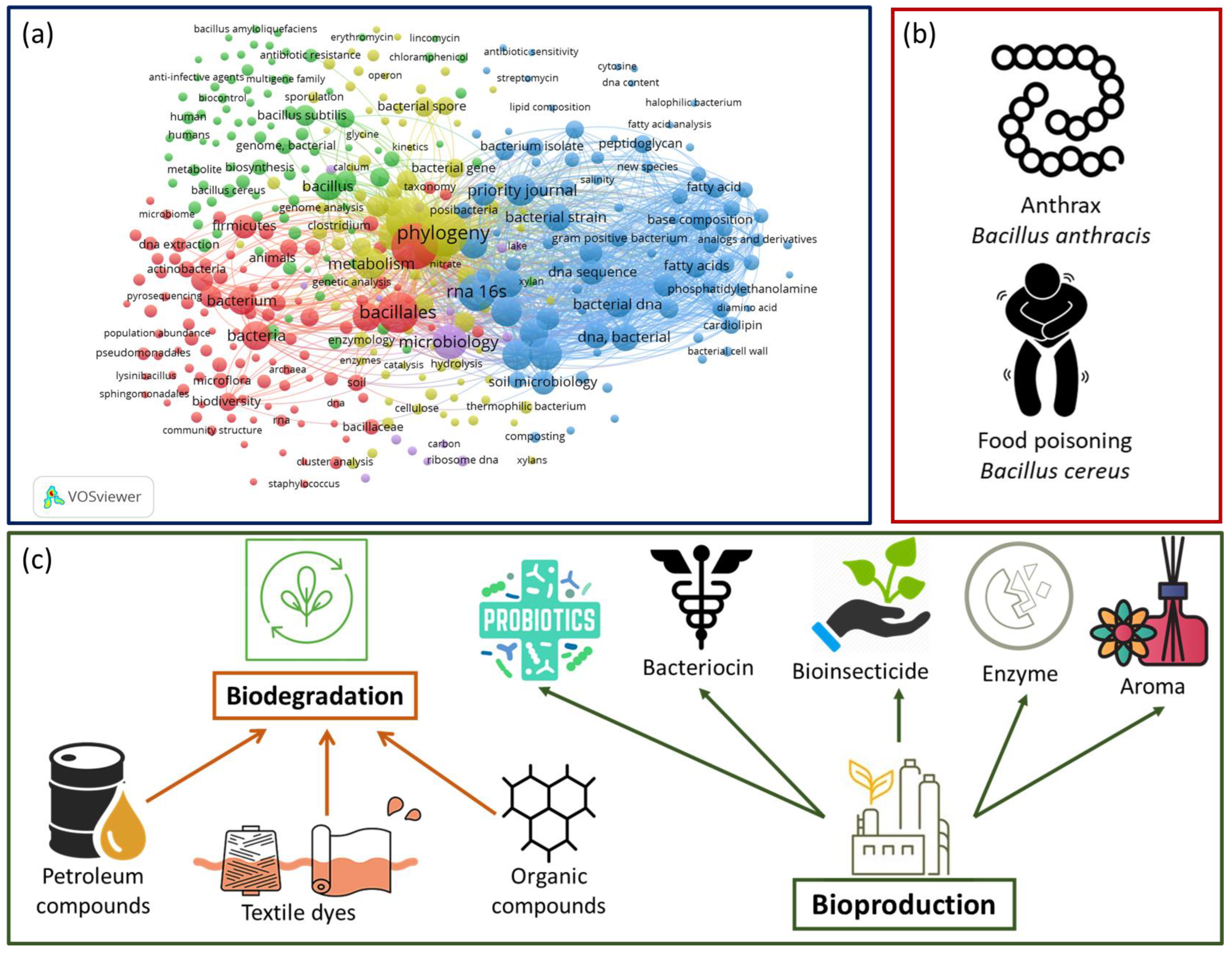
2.1. Taxonomy

{kind=link}
| Family Name | Proposed by | IJSEM 1 Validation List No. (Year) | Type Genus | Involved Valid and Invalid Genera Until 2022 in LPSN 2 | Valid Genera with Correct Name until 2022 in LPSN |
|---|---|---|---|---|---|
| Alicyclobacillaceae | da Costa and Rainey 2010 | 132 (2010) | Alicyclobacillus | 5 | 5 |
| Bacillaceae | Fischer 1895 | (1980) | Bacillus | 134 | 117 |
| Caryophanaceae | Peshkoff 1939 | (1980) | Caryophanon | 26 | 19 |
| Desulfuribacillaceae | Sorokin et al. 2021 | 200 (2021) | Desulfuribacillus | 1 | 1 |
| Listeriaceae | Ludwig et al. 2010 | 132 (2010) | Listeria | 3 | 2 |
| Paenibacillaceae | De Vos et al. 2010 | 132 (2010) | Paenibacillus | 19 | 16 |
| Pasteuriaceae | Laurent 1890 | (1980) | Pasteuria | 1 | 1 |
| Sporolactobacillaceae | Ludwig et al. 2010 | 132 (2010) | Sporolactobacillus | 5 | 5 |
| Staphylococcaceae | Schleifer and Bell 2010 | 132 (2010) | Staphylococcus | 12 | 10 |
| Thermoactinomycetaceae | Matsuo et al. 2006 | (2007) | Thermoactinomyces | 25 | 24 |
| Incertae sedis 46 | - | - | - | 1 | 1 |
2.2. General Characteristics
3. Genus Bacillus
3.1. Genotypic Characteristics
3.2. Phenotypic, Chemotaxonomic, and Morphological Characteristics
3.3. Pathological Importance
3.4. Sporulation and Spore Properties
3.5. Industrial Applications
3.5.1. Food Industry
3.5.2. Pharmaceutical Industry
| Species Name | Enzyme Class | Main Industrial Application | Reference |
|---|---|---|---|
| B. amyloliquefaciens B. subtilis B. thuringiensis | Extracellular metalloprotease | Leather industry | [107,108,109] |
| B. amyloliquefaciens B. licheniformis B. subtilis B. pumilus | Serine protease | Detergent industry Antifungal agent | [110,111,112,113] |
| B. alcalophilus B. amyloliquefaciens B. licheniformis B. subtilis B. thuringiensis | α-Amylase | Starch and food industries | [114,115,116,117,118] |
| B. cereus var. mycoides | β-Amylase | Starch industry | [119] |
| B. cereus FDTA-13 Thermophilic Bacillus sp. AN-7 | pullulanase | [120,121] | |
| Bacillus sp. CICIM 304 | Iso-amylase | [122] | |
| Bacillus sp. CSB55 B. subtilis | β-glucanase | [123,124] | |
| B. subtilis B. licheniformis | β-lactamase (Penicillinase) | Molecular Biology Antibiotic industry | [125,126] |
| B. licheniformis | Glucose iso-merase | Food industry | [127] |
| Bacillus methylotrophicus B. subtilis | Levansucrase | Food and pharmaceutical industries | [128,129] |
| B. subtilis B. pseudomycoides B. licheniformis | Cellulase | Biofuel industry | [130,131,132] |
| Bacillus australimaris Bacillus tequilensis | Xylanase | Biofuel and paper industries | [133,134] |
| B. licheniformis Bacillus sp. G-825-6 Bacillus sp. ND1 | Cyclodextrin glucanotransferase (CGTase) | Pharmaceutical, food and cosmetic industries | [135,136,137] |
| B. subtilis E9 B. mojavensis TH309 | Esterase | Biodegradation | [138,139] |
| B. thuringiensis B. licheniformis B307 | Chitinase | Biocontrol agent | [140,141] |
| B. velezensis TA3 B.cereus B. subtilis | Tannase | Delignification process | [142,143,144] |
3.5.3. Agricultural Industry
3.5.4. Other Industrial Applications of the Genus Bacillus
4. Spore-Forming Genera of Bacillales
4.1. Alicyclobacillaceae Genera
4.2. Bacillaceae Genera
4.3. Paenibacillaceae Genera
4.4. Other Spore-Forming Genera and Their Significance
5. Halophilic Genera of Bacillales
Potential and Industrial Applications
6. Thermophilic and Psychrophilic Genera of Bacillales
Industrial Applications
7. Extremophiles and Polyextremophilic Genera of Bacillales
7.1. Acidophilic Genera and Their Applications
7.2. Alkaliphilic Genera
7.3. Polyextremophilic Genera
8. Engineered Strains of Bacillales
9. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tindall, B.J. Names at the rank of class, subclass and order, their typification and current status: Supplementary information to Opinion 79. Judicial Commission of the International Committee on Systematics of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 3599–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, P.; Garrity, G.M.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.; Whitman, W.B. (Eds.) Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes; Springer: New York, NY, USA, 2010. [Google Scholar]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. Microbiol. Spectrum. 2015, 3, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherardi, G.; Di Bonaventura, G.; Savini, V. Chapter 1-Staphylococcal Taxonomy, in Pet-To-Man Travelling Staphylococci; Savini, V., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–10. [Google Scholar]
- Little, S.F.; Ivins, B.E. Molecular pathogenesis of Bacillus anthracis infection. Microbes Infect. 1999, 1, 131–139. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Planellas, M.H.; Long, K.S.; Vester, B. The Order Bacillales Hosts Functional Homologs of the Worrisome cfr Antibiotic Resistance Gene. Antimicrob. Agents Chemother. 2012, 56, 3563–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, M.; Bender, K.; Buckley, D.; Sattley, W.; Stahl, D. Brock Biology of Microorganisms; Pearson: London, UK, 2014. [Google Scholar]
- Ludwig, W.; Schleifer, K.-H.; Whitman, W.B. Revised road map to the phylum Firmicutes. Bergey’s Man. Syst. Bacteriol. 2009, 2, 1–8. [Google Scholar]
- Brenner, D.J.; Krieg, N.R.; Staley, J.T. Appendix 2. Taxonomic outline of the Archaea and Bacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Garrity, G.M., Staley, J.T., Boone, D.R., Brenner, D.J., Vos, P.D., Goodfellow, M., Krieg, N.R., Rainey, F.A., Schleifer, K., Eds.; Springer International Publishing: New York, NY, USA, 2005; pp. 207–220. [Google Scholar]
- Skerman, V.B.D.; McGowan, V.; Sneath, P.H.A. Approved lists of bacterial names. Int. J. Syst. Bacteriol. 1980, 30, 225–420. [Google Scholar] [CrossRef] [Green Version]
- Parker, C.T.; Tindall, B.J.; Garrity, G.M. International Code of Nomenclature of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2019, 69, S1–S111. [Google Scholar]
- Tindall, B.J. When treated as heterotypic synonyms the names Caryophanaceae Peshkoff 1939 (Approved Lists 1980) and Caryophanales Peshkoff 1939 (Approved Lists 1980) have priority over the names Planococcaceae Krasil’nikov 1949 (Approved Lists 1980) and Bacillales Prévot 1953 (Approved Lists 1980) and Bacillales Prévot 1953 (Approved Lists 1980), respectively. Int. J. Syst. Evol. Microbiol. 2019, 69, 2187–2195. [Google Scholar]
- NamesforLife Bacterial and Archaeal Nomenclature. Available online: https://www.namesforlife.com/search (accessed on 20 August 2022).
- Vandamme, P.; Pot, B.; Gillis, M.; de Vos, P.; Kersters, K.; Swings, J. Polyphasic taxonomy, a consensus approach to bacterial systematics. Microbiol. Rev. 1996, 60, 407–438. [Google Scholar] [CrossRef]
- De Maayer, P.; Aliyu, H.; Cowan, D.A. Reorganising the order Bacillales through phylogenomics. Syst. Appl. Microbiol. 2018, 42, 178–189. [Google Scholar] [CrossRef]
- Fritze, D. Taxonomy of the Genus Bacillus and Related Genera: The Aerobic Endospore-Forming Bacteria. Phytopathology 2004, 94, 1245–1248. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Kang, S.S.; Lee, K.C.; Lee, E.S.; Kho, Y.H.; Kang, K.H.; Park, Y.H. Planomicrobium koreense gen. nov., sp. nov., a bacterium isolated from the Korean traditional fermented seafood jeotgal, and transfer of Planococcus okeanokoites (Nakagawa et al. 1996) and Planococcus mcmeekinii (Junge et al. 1998) to the genus Planomicrobium. Int. J. Syst. Evol. Microbiol. 2001, 51, 1511–1520. [Google Scholar] [CrossRef]
- Yarza, P.; Ludwig, W.; Euzéby, J.; Amann, R.; Schleifer, K.-H.; Glöckner, F.O.; Rosselló-Móra, R. Update of the All-Species Living Tree Project based on 16S and 23S rRNA sequence analyses. Syst. Appl. Microbiol. 2010, 33, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Patel, S. Robust Demarcation of the Family Caryophanaceae (Planococcaceae) and Its Different Genera Including Three Novel Genera Based on Phylogenomics and Highly Specific Molecular Signatures. Front. Microbiol. 2020, 10, 2821. [Google Scholar] [CrossRef] [Green Version]
- Maughan, H.; van der Auwera, G. Bacillus taxonomy in the genomic era finds phenotypes to be essential though often misleading. Infect. Genet. Evol. 2011, 11, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Gupta, R.S. A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef] [PubMed]
- Gaballa, A.; Cheng, R.A.; Trmcic, A.; Kovac, J.; Kent, D.J.; Martin, N.H.; Wiedmann, M. Development of a database and standardized approach for rpoB sequence-based subtyping and identification of aerobic spore-forming Bacillales. J. Microbiol. Methods 2021, 191, 106350. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Whitman, W.B. Genome sequences as the type material for taxonomic descriptions of prokaryotes. Syst. Appl. Microbiol. 2015, 38, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust Demarcation of 17 Distinct Bacillus Species Clades, Proposed as Novel Bacillaceae Genera, by Phylogenomics and Comparative Genomic Analyses: Description of Robertmurraya kyonggiensis Sp. Nov. And Proposal for an Emended Genus Bacillus Limiting It Only to the Members of the Subtilis and Cereus Clades of Species. Int. J. Syst. Evolut. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, j.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (Lpsn) Moves to the Dsmz. Int. J. Syst. Evolut. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Tille, P.M. Bailey & Scott’s Diagnostic Microbiology; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Harwood, C.R. Bacillus; Springer: New York, NY, USA, 2013. [Google Scholar]
- Driks, A.; Eichenberger, P. The Bacterial Spore: From Molecules to Systems; ASM Press: Washington, DC, USA, 2016. [Google Scholar]
- Logan, N.A.; Berge, O.; Bishop, A.H.; Busse, H.-J.; de Vos, P.; Fritze, D.; Heyndrickx, M.; Kämpfer, P.; Rabinovitch, L.; Salkinoja-Salonen, M.S.; et al. Proposed Minimal Standards for Describing New Taxa of Aerobic, Endospore-Forming Bacteria. Int. J. Syst. Evolut. Microbiol. 2009, 59, 2114–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helgason, E.; Tourasse, N.J.; Meisal, R.; Caugant, D.A.; Kolstø, A. Multilocus Sequence Typing Scheme for Bacteria of the Bacillus Cereus Group. Appl. Environ. Microbiol. 2004, 70, 191–201. [Google Scholar] [CrossRef]
- Adelskov, J.; Patel, B.K.C. A molecular phylogenetic framework for Bacillus subtilis using genome sequences and its application to Bacillus subtilis subspecies stecoris strain D7XPN1, an isolate from a commercial food-waste degrading bioreactor. 3 Biotech 2016, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [Green Version]
- Conrad, A.I. Applications and Systematics of Bacillus and Relatives. Based on a meeting held in Bruges, Belgium, August 2000. Edited by Roger Berkeley, Marc Heyndrickx, Niall Logan, and Paul De Vos. Q. Rev. Biol. 2003, 78, 229–230. [Google Scholar]
- Harwood, C.R.; Mouillon, J.-M.; Pohl, S.; Arnau, J. Secondary metabolite production and the safety of industrially important members of the Bacillus subtilis group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, V.; Ahmod, N.Z.; Shah, H.N.; Gupta, R.S. Molecular signatures for Bacillus species: Demarcation of the Bacillus subtilis and Bacillus cereus clades in molecular terms and proposal to limit the placement of new species into the genus Bacillus. Int. J. Syst. Evol. Microbiol. 2013, 63, 2712–2726. [Google Scholar] [CrossRef] [Green Version]
- Traag, B.A.; Pugliese, A.; Eisen, J.A.; Losick, R. Gene Conservation among Endospore-Forming Bacteria Reveals Additional Sporulation Genes in Bacillus subtilis. J. Bacteriol. 2013, 195, 253. [Google Scholar] [CrossRef] [Green Version]
- Errington, J. Regulation of endospore formation in Bacillus subtilis. Nat. Rev. Genet. 2003, 1, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, T.; Littlechild, J.; Kawarabayasi, Y. Thermophilic Microbes in Environmental and Industrial Biotechnology: Biotechnology of Thermophiles; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Pichinoty, F.; Asselineau, J.; Mandel, M. Biochemical characterization of Bacillus benzoevorans sp. nov., a new filamentous, sheathed mesophilic species, degrading various aromatic acids and phenols. Ann. De Microbiol. 1984, 135, 209–217. [Google Scholar]
- Horikoshi, K. Alkaliphiles: Some applications of their products for biotechnology. Microbiol. Mol. Biol. Rev. 1999, 63, 735–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, C.; Booth, N.; Allan, E. An ELISA for the detection of Bacillus subtilis L-form bacteria confirms their symbiosis in strawberry. Lett. Appl. Microbiol. 2000, 31, 390–394. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.-M.; Pum, D. S-layers: Principles and applications. FEMS Microbiol. Rev. 2014, 38, 823–864. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.-Y.; Lövgren, A.; Landén, R.; Lögren, A. Adhesion and Cytotoxicity of Bacillus thuringiensis to Cultured Spodoptera and Drosophila Cells. J. Invertebr. Pathol. 1995, 66, 46–51. [Google Scholar] [CrossRef]
- Barnaby, W. Plague Makers: The Secret World of Biological Warfare, 3rd ed.; Bloomsbury Academic: London, UK, 2002. [Google Scholar]
- Roffey, R.; Tegnell, A.; Elgh, F. Biological warfare in a historical perspective. Clin. Microbiol. Infect. 2002, 8, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.Z.; Ali, G.M.; Nazir, F. Bacillus thuringiensis and its application in agriculture. Afr. J. Biotechnol. 2010, 9, 2022–2031. [Google Scholar]
- Feitelson, J.S.; Payne, J.; Kim, L. Bacillus thuringiensis: Insects and Beyond. Nat. Biotechnol. 1992, 10, 271–275. [Google Scholar] [CrossRef]
- Manktelow, C.J.; White, H.; Crickmore, N.; Raymond, B. Divergence in environmental adaptation between terrestrial clades of the Bacillus cereus group. FEMS Microbiol. Ecol. 2020, 97, fiaa228. [Google Scholar] [CrossRef]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus Endospores to Extreme Terrestrial and Extraterrestrial Environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piggot, P.J.; Hilbert, D.W. Sporulation of Bacillus subtilis. Curr. Opin. Microbiol. 2004, 7, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Angelini, L.L.; Chai, Y. Bacillus subtilis Cell Differentiation, Biofilm Formation and Environmental Prevalence. Microorganisms 2022, 10, 1108. [Google Scholar] [CrossRef] [PubMed]
- Henriques, A.O.; Moran, J.C.P. Structure, Assembly, and Function of the Spore Surface Layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef] [PubMed]
- Schallmey, M.; Singh, A.; Ward, O.P. Developments in the use of Bacillus species for industrial production. Can. J. Microbiol. 2004, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ma, R.; Lin, P.; Yuan, H.; Liu, H.; Jiang, Y.; Huang, D.; Ma, Y.; Wang, T. Display of Glucose Oxidase on Bacillus subtilis Spore Surface and Its Application in Electrode. World 2022, 7, 34–47. [Google Scholar]
- Nath, S.C.; Dora, K.C. Application of Bacillus sp. as a biopreservative for food preservation. Int. J. Eng. Res. Appl. 2015, 5, 85–95. [Google Scholar]
- Mohan, G.; Thangappanpillai, A.K.T.; Ramasamy, B. Antimicrobial activities of secondary metabolites and phylogenetic study of sponge endosymbiotic bacteria, Bacillus sp. at Agatti Island, Lakshadweep Archipelago. Biotechnol. Rep. 2016, 11, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Sansinenea, E.; Ortiz, A. Secondary metabolites of soil Bacillus spp. Biotechnol. Lett. 2011, 33, 1523–1538. [Google Scholar] [CrossRef]
- Mohkam, M.; Nezafat, N.; Berenjian, A.; Negahdaripour, M.; Behfar, A.; Ghasemi, Y. Role of Bacillus Genus in the Production of Value-Added Compounds. In Bacilli and Agrobiotechnology; Islam, M.T., Rahman, M., Pandey, P., Jha, C.K., Aeron, A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–33. [Google Scholar]
- Sar, T.; Harirchi, S.; Ramezani, M.; Bulkan, G.; Akbas, M.Y.; Pandey, A.; Taherzadeh, M.J. Potential utilization of dairy industries by-products and wastes through microbial processes: A critical review. Sci. Total Environ. 2022, 810, 152253. [Google Scholar] [CrossRef]
- Scawen, M.D.; Hammond, P.M.; Comer, M.J.; Atkinson, T. The application of triazine dye affinity chromatography to the large-scale purification of glycerokinase from Bacillus stearothermophilus. Anal. Biochem. 1983, 132, 413–417. [Google Scholar] [CrossRef]
- Goward, C.R.; Hartwell, R.; Atkinson, T.; Scawen, M.D. The purification and characterization of glucokinase from the thermophile Bacillus stearothermophilus. Biochem. J. 1986, 237, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, W.; Kula, M.-R. Leucine dehydrogenase from Bacillus sphaericus. Optimized production conditions and an efficient method for its large-scale purification. Appl. Microbiol. Biotechnol. 1981, 12, 22–27. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Chen, J.; Xu, M.; Yang, T.; Zheng, J.; Zhang, X.; Rao, Z. Rational Engineering of Bacillus cereus Leucine Dehydrogenase Towards α-keto Acid Reduction for Improving Unnatural Amino Acid Production. Biotechnol. J. 2018, 14, e1800253. [Google Scholar] [CrossRef] [PubMed]
- Areekit, S.; Kanjanavas, P.; Khawsak, P.; Pakpitchareon, A.; Potivejkul, K.; Chansiri, G.; Chansiri, K. Cloning, Expression, and Characterization of Thermotolerant Manganese Superoxide Dismutase from Bacillus sp. MHS47. Int. J. Mol. Sci. 2011, 12, 844–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itakorode, B.O.; Okonji, R.E.; Adedeji, O.; Torimiro, N.; Onwudiegwu, C.; Oluwaseyi, A. Studies on some physicochemical properties of Rhodanese synthesized by Bacillus cereus isolated from the effluents of iron and steel smelting industry. Afr. J. Biochem. Res. 2019, 13, 1–8. [Google Scholar]
- De Boer, A.S.; Priest, F.; Diderichsen, B. On the industrial use of Bacillus licheniformis: A review. Appl. Microbiol. Biotechnol. 1994, 40, 595–598. [Google Scholar] [CrossRef]
- de Boer Sietske, A.; Diderichsen, B. On the safety of Bacillus subtilis and B. amyloliquefaciens: A review. Appl. Microbiol. Biotechnol. 1991, 36, 1–4. [Google Scholar] [CrossRef]
- He, W.; Mu, W.; Jiang, B.; Yan, X.; Zhang, T. Construction of a Food Grade Recombinant Bacillus subtilis Based on Replicative Plasmids with an Auxotrophic Marker for Biotransformation of d-Fructose to d-Allulose. J. Agric. Food Chem. 2016, 64, 3243–3250. [Google Scholar] [CrossRef]
- Ngalimat, M.; Yahaya, R.; Baharudin, M.; Yaminudin, S.; Karim, M.; Ahmad, S.; Sabri, S. A Review on the Biotechnological Applications of the Operational Group Bacillus amyloliquefaciens. Microorganisms 2021, 9, 614. [Google Scholar] [CrossRef]
- Priest, F.G.; Goodfellow, M.; Shute, L.A.; Berkeley, R.C.W. Bacillus amyloliquefaciens sp. nov., nom. rev. Int. J. Syst. Evolut. Microbiol. 1987, 37, 69–71. [Google Scholar] [CrossRef]
- Alagöz, D.; Yildirim, D.; Güvenmez, H.K.; Sihay, D.; Tükel, S.S. Covalent Immobilization and Characterization of a Novel Pullulanase from Fontibacillus sp. Strain DSHK 107 onto Florisil® and Nano-silica for Pullulan Hydrolysis. Appl. Biochem. Biotechnol. 2016, 179, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Contact Materials; Enzymes, Flavourings and Processing Aids (CEF); Silano, V.; Bolognesi, C.; Castle, L.; Chipman, K.; Cravedi, J.P.; Fowler, P.; Franz, R.; Grob, K.; et al. Safety evaluation of the food enzyme pullulanase from Pullulanibacillus naganoensis strain AE-PL. EFSA J. 2017, 15, e05009. [Google Scholar]
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Bin Ariff, A. Pullulanase: Role in Starch Hydrolysis and Potential Industrial Applications. Enzym. Res. 2012, 2012, 921362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, R.F.; Wheals, A.E. The Microbiology of Cocoa Fermentation and its Role in Chocolate Quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Soran, H.; Beyatli, Y. Antimicrobial activities of some Bacillus spp. strains isolated from the soil. Microbiol. Res. 2006, 161, 127–131. [Google Scholar] [CrossRef]
- Sumi, C.D.; Yang, B.W.; Yeo, I.-C.; Hahm, Y.T. Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Can. J. Microbiol. 2015, 61, 93–103. [Google Scholar] [CrossRef]
- Cladera-Olivera, F.; Caron, G.; Brandelli, A. Bacteriocin-like substance production by Bacillus licheniformis strain P40. Lett. Appl. Microbiol. 2004, 38, 251–256. [Google Scholar] [CrossRef]
- Sharma, N.G.N. Antibacterial activity and characterization of bacteriocin of Bacillus mycoides isolated from whey. Indian J. Biotechnol. 2008, 7, 117–121. [Google Scholar]
- Fanaei, M.; Emtiazi, G. Microbial assisted (Bacillus mojavensis) production of bio-surfactant lipopeptide with potential pharmaceutical applications and its characterization by MALDI-TOF-MS analysis. J. Mol. Liq. 2018, 268, 707–714. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.A.P.; Omar, N.B.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, L.; Singh, G.; Choudhary, V.; Sahoo, D.K. Sonorensin: An Antimicrobial Peptide, Belonging to the Heterocycloanthracin Subfamily of Bacteriocins, from a New Marine Isolate, Bacillus sonorensis MT93. Appl. Environ. Microbiol. 2014, 80, 2981. [Google Scholar] [CrossRef] [PubMed]
- Leães, F.L.; Vanin, N.G.; Sant’Anna, V.; Brandelli, A. Use of Byproducts of Food Industry for Production of Antimicrobial Activity by Bacillus sp. P11. Food Bioprocess Technol. 2010, 4, 822–828. [Google Scholar] [CrossRef]
- Torres, N.I.; Noll, K.S.; Xu, S.; Li, J.; Huang, Q.; Sinko, P.J.; Wachsman, M.B.; Chikindas, M.L. Safety, Formulation and In Vitro Antiviral Activity of the Antimicrobial Peptide Subtilosin Against Herpes Simplex Virus Type 1. Probiotics Antimicrob. Proteins 2013, 5, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Bizani, D.; Motta, A.S.; Morrissy, J.A.C.; Terra, R.M.S.; Souto, A.; Brandelli, A. Antibacterial activity of cerein 8A, a bacteriocin-like peptide produced by Bacillus cereus. Int. Microbiol. 2005, 8, 125–131. [Google Scholar]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus coagulans in the Food Industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Shahcheraghi, S.; Ayatollahi, J.; Lotfi, M. Applications of Bacillus subtilis as an important bacterium in medical sciences and human life. Trop. J. Med. Res. 2015, 18, 1–4. [Google Scholar] [CrossRef]
- Sneader, W. Drug Discovery: A History; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Malerba, F.; Orsenigo, L. The evolution of the pharmaceutical industry. Bus. Hist. 2015, 57, 664–687. [Google Scholar] [CrossRef]
- Jozala, A.F.; Geraldes, D.C.; Tundisi, L.L.; Feitosa, V.D.A.; Breyer, C.A.; Cardoso, S.L.; Mazzola, P.; Nascimento, L.D.O.; Rangel-Yagui, C.; Magalhães, P.D.O.; et al. Biopharmaceuticals from microorganisms: From production to purification. Braz. J. Microbiol. 2016, 47, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Saengkerdsub, S. Recombinant Protein Production in Bacillus Species, Isolation and Methionine Production in Methionine-producing Microorganisms. Ph.D. Thesis, University of Arkansas, Fayetteville, NC, USA, 2012. [Google Scholar]
- Westers, L.; Westers, H.; Quax, W.J. Bacillus subtilis as cell factory for pharmaceutical proteins: A biotechnological approach to optimize the host organism. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2004, 1694, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Miralles, N.; Villaverde, A. Bacterial cell factories for recombinant protein production; expanding the catalogue. Microb. Cell Factories 2013, 12, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbás, P.; Lorence, A. Recombinant Gene Expression: Reviews and Protocols; Humana Press: Totowa, NJ, USA, 2004. [Google Scholar]
- Nakano, M.M.; Zuber, P. Molecular Biology of Antibiotic Production in Bacillus. Crit. Rev. Biotechnol. 1990, 10, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef]
- Xiang, Y.-Z.; Li, X.-Y.; Zheng, H.-L.; Chen, J.-Y.; Lin, L.-B.; Zhang, Q.-L. Purification and antibacterial properties of a novel bacteriocin against Escherichia coli from Bacillus subtilis isolated from blueberry ferments. LWT 2021, 146, 111456. [Google Scholar] [CrossRef]
- Franzetti, A.; Tamburini, E.; Banat, I.M. Applications of biological surface active compounds in remediation technologies. In Biosurfactants; Springer: Berlin/Heidelberg, Germany, 2010; pp. 121–134. [Google Scholar] [CrossRef]
- Nicolas, J. Molecular Dynamics Simulation of Surfactin Molecules at the Water-Hexane Interface. Biophys. J. 2003, 85, 1377–1391. [Google Scholar] [CrossRef] [Green Version]
- Schaller, K.D.; Fox, S.L.; Bruhn, D.F.; Noah, K.S.; Bala, G.A. Characterization of Surfactin from Bacillus subtilis for Application as an Agent for Enhanced Oil Recovery. Appl. Biochem. Biotechnol. 2004, 115, 827–836. [Google Scholar] [CrossRef]
- He, L.; Chen, W.; Liu, Y. Production and partial characterization of bacteriocin-like pepitdes by Bacillus licheniformis ZJU12. Microbiol. Res. 2006, 161, 321–326. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y.; Xu, Y.; Zhang, Y.; Tao, G.; Zhang, L.; Shi, G. Transcriptome Analysis of Bacillus licheniformis for Improving Bacitracin Production. ACS Synth. Biol. 2022, 11, 1325–1335. [Google Scholar] [CrossRef]
- Cherif, A.; Chehimi, S.; Limem, F.; Hansen, B.; Hendriksen, N.; Daffonchio, D.; Boudabous, A. Detection and characterization of the novel bacteriocin entomocin 9, and safety evaluation of its producer, Bacillus thuringiensis ssp. entomocidus HD9. J. Appl. Microbiol. 2003, 95, 990–1000. [Google Scholar] [CrossRef]
- Malanicheva, I.A.; Kozlov, D.G.; Efimenko, T.A.; Zenkova, V.A.; Katrukha, G.S.; Reznikova, M.I.; Korolev, A.M.; Borshchevskaya, L.N.; Tarasova, O.D.; Sineokii, S.P.; et al. New antibiotics produced by Bacillus subtilis strains. Microbiology 2014, 83, 352–356. [Google Scholar] [CrossRef]
- Erem, F.; Inan, M.; Karakaş Budak, B.; Certel, M. Partial Purification and Characterization of an Extracellular Metallopeptidase Produced by Bacillus Amyloliquefaciens Fe-K1. Trakya Univ. J. Nat. Sci. 2020, 21, 47–61. [Google Scholar] [CrossRef]
- Park, C.H.; Lee, S.J.; Lee, S.G.; Lee, W.S.; Byun, S.M. Hetero-and Autoprocessing of the Extracellular Metalloprotease (Mpr) in Bacillus Subtilis. J. Bacteriol. 2004, 186, 6457–6464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedhila, S.; Nel, P.; Lereclus, D. The Inha2 Metalloprotease of Bacillus Thuringiensis Strain 407 Is Required for Pathogenicity in Insects Infected Via the Oral Route. J. Bacteriol. 2002, 184, 3296–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, C.; Wei, R.; Jiang, H.; Zhao, Y.; Xu, L. Characterization of the Ammonification, the Relevant Protease Production and Activity in a High-Efficiency Ammonifier Bacillus Amyloliquefaciens Dt. Int. Biodeterior. Biodegrad. 2019, 142, 11–17. [Google Scholar] [CrossRef]
- Ji, Z.-L.; Peng, S.; Chen, L.-L.; Liu, Y.; Yan, C.; Zhu, F. Identification and Characterization of a Serine Protease from Bacillus Licheniformis W10: A Potential Antifungal Agent. Int. J. Biol. Macromol. 2020, 145, 594–603. [Google Scholar] [CrossRef]
- Thu, N.T.A.; Khue, N.T.M.; Huy, N.D.; Tien, N.Q.D.; Loc, N.H. Characterizations and Fibrinolytic Activity of Serine Protease from Bacillus Subtilis C10. Curr. Pharm. Biotechnol. 2020, 21, 110–116. [Google Scholar] [CrossRef]
- Jagadeesan, Y.; Meenakshisundaram, S.; Saravanan, V.; Balaiah, A. Sustainable Production, Biochemical and Molecular Characterization of Thermo-and-Solvent Stable Alkaline Serine Keratinase from Novel Bacillus Pumilus Ar57 for Promising Poultry Solid Waste Management. Int. J. Biol. Macromol. 2020, 163, 135–146. [Google Scholar] [CrossRef]
- Kumar, R.; Mehta, A. Isolation, Optimization and Characterization of A-Amylase from Bacillus Alcalophilus. Int. J. Sci. Res. 2013, 2, 171–174. [Google Scholar]
- Du, R.; Song, Q.; Zhang, Q.; Zhao, F.; Kim, R.; Zhou, Z.; Han, Y. Purification and Characterization of Novel Thermostable and Ca-Independent A-Amylase Produced by Bacillus Amyloliquefaciens Bh072. Int. J. Biol. Macromol. 2018, 115, 1151–1156. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, Y.; Li, Q.; Zhou, J.; Li, J.; Du, G. Growth-Coupled Evolution and High-Throughput Screening Assisted Rapid Enhancement for Amylase-Producing Bacillus Licheniformis. Bioresour. Technol. 2021, 337, 125467. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.; Nag, M.; Sarkar, T.; Dutta, B.; Ray, R.R. Antibiofilm Activity of A-Amylase from Bacillus Subtilis and Prediction of the Optimized Conditions for Biofilm Removal by Response Surface Methodology (Rsm) and Artificial Neural Network (Ann). Appl. Biochem. Biotechnol. 2021, 193, 1853–1872. [Google Scholar] [CrossRef] [PubMed]
- Maity, C.; Samanta, S.; Halder, S.K.; Mohapatra, P.K.D.; Pati, B.R.; Jana, M.; Mondal, K.C. Isozymes of A-Amylases from Newly Isolated Bacillus Thuringiensis Ckb19: Production from Immobilized Cells. Biotechnol. Bioprocess Eng. 2011, 16, 312–319. [Google Scholar] [CrossRef]
- Ye, Z.; Miyake, H.; Tatsumi, M.; Nishimura, S.; Nitta, Y. Two Additional Carbohydrate-Binding Sites of Β-Amylase from Bacillus Cereus Var. Mycoides Are Involved in Hydrolysis and Raw Starch-Binding. J. Biochem. 2004, 135, 355–363. [Google Scholar] [CrossRef]
- Ling, H.; Chuan, L.; Rosfarizan, M.; Ariff, A.B. Characterization of Pullulanase Type Ii from Bacillus Cereus H1. 5. Am. J. Biochem. Biotechnol. 2009, 5, 170–179. [Google Scholar]
- Kunamneni, A.; Singh, S. Improved High Thermal Stability of Pullulanase from a Newly Isolated Thermophilic Bacillus Sp. An-7. Enzyme Microb. Technol. 2006, 39, 1399–1404. [Google Scholar] [CrossRef]
- Li, Y.; Niu, D.; Zhang, L.; Wang, Z.; Shi, G. Purification, Characterization and Cloning of a Thermotolerant Isoamylase Produced from Bacillus Sp. Cicim 304. J. Ind. Microbiol. Biotechnol. 2013, 40, 437–446. [Google Scholar] [CrossRef]
- Dewi, R.T.K.; Mubarik, N.R.; Suhartono, M.T. Medium Optimization of Β-Glucanase Production by Bacillus Subtilis Saha 32.6 Used as Biological Control of Oil Palm Pathogen. Emir. J. Food Agric. 2016, 28, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Regmi, S.; Choi, Y.S.; Kim, Y.K.; Khan, M.M.; Lee, S.H.; Choi, Y.H.; Cho, S.S.; Jin, Y.; Yoo, J.C.; Suh, J.-W. Industrial Attributes of Β-Glucanase Produced by Bacillus Sp. Csb55 and Its Potential Application as Bio-Industrial Catalyst. Bioprocess Biosyst. Eng. 2020, 43, 249–259. [Google Scholar] [CrossRef]
- Vandevenne, M.; Filee, P.; Scarafone, N.; Cloes, B.; Gaspard, G.; Yilmaz, N.; Dumoulin, M.; François, J.; Frère, J.; Galleni, M. The Bacillus Licheniformis Blap B-Lactamase as a Model Protein Scaffold to Study the Insertion of Protein Fragments. Protein Sci. 2007, 16, 2260–2271. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Tayyab, M.; Hashmi, A.S.; Nadeem, A.; Hanif, S.; Firyal, S.; Saeed, S.; Awan, A.R.; Wasim, M. Characterization of a Novel Recombinant β-Lactamase from Bacillus subtilis R5. Pak. J. Zool. 2022, 54. [Google Scholar] [CrossRef]
- Nwokoro, O. Studies on the production of glucose isomerase by Bacillus licheniformis. Pol. J. Chem. Technol. 2015, 17, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Esawy, M.A.; Ahmed, E.F.; Helmy, W.A.; Mansour, N.M.; El-Senousy, W.M.; El-Safty, M.M. Production of Levansucrase from Novel Honey Bacillus Subtilis Isolates Capable of Producing Antiviral Levans. Carbohydr. Polym. 2011, 86, 823–830. [Google Scholar] [CrossRef]
- Zhang, T.; Li, R.; Qian, H.; Mu, W.; Miao, M.; Jiang, B. Biosynthesis of Levan by Levansucrase from Bacillus Methylotrophicus Sk 21.002. Carbohydr. Polym. 2014, 101, 975–981. [Google Scholar] [CrossRef]
- Sreena, C.P.; Sebastian, D. Augmented Cellulase Production by Bacillus Subtilis Strain Mu S1 Using Different Statistical Experimental Designs. J. Genet. Eng. Biotechnol. 2018, 16, 9–16. [Google Scholar] [CrossRef]
- Pramanik, S.K.; Mahmud, S.; Paul, G.K.; Jabin, T.; Naher, K.; Uddin, M.S.; Zaman, S.; Saleh, M.A. Fermentation Optimization of Cellulase Production from Sugarcane Bagasse by Bacillus Pseudomycoides and Molecular Modeling Study of Cellulase. Curr. Res. Microb. Sci. 2021, 2, 100013. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yang, D.; Wang, X.; Ca, W.o. A Novel Thermostable Cellulase-Producing Bacillus Licheniformis A5 Acts Synergistically with Bacillus Subtilis B2 to Improve Degradation of Chinese Distillers’ Grains. Bioresour. Technol. 2021, 325, 124729. [Google Scholar] [CrossRef]
- Dutta, P.D.; Neog, B.; Goswami, T. Xylanase Enzyme Production from Bacillus Australimaris P5 for Prebleaching of Bamboo (Bambusa Tulda) Pulp. Mater. Chem. Phys. 2020, 243, 122227. [Google Scholar] [CrossRef]
- Patel, K.; Dudhagara, P. Optimization of Xylanase Production by Bacillus Tequilensis Strain Ud-3 Using Economical Agricultural Substrate and Its Application in Rice Straw Pulp Bleaching. Biocatal. Agric. Biotechnol. 2020, 30, 101846. [Google Scholar] [CrossRef]
- Man, R.C.; Shaarani, S.M.; Arshad, Z.I.M.; Mudalip, S.K.A.; Sulaiman, S.Z.; Ramli, A.N.M. Effect of Reaction Parameters on the Production of Cyclodextrin Using Cyclodextrin Glucanotransferase from Bacillus Licheniformis. J. Chem. Eng. Ind. Biotechnol. 2020, 6, 39–45. [Google Scholar]
- Sonnendecker, C.; Melzer, S.; Zimmermann, W. Engineered Cyclodextrin Glucanotransferases from Bacillus Sp. G-825-6 Produce Large-Ring Cyclodextrins with High Specificity. MicrobiologyOpen 2019, 8, e00757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, D.; Sharma, S.; Shrivastava, D.; Kulshreshtha, N.M. Production and Characterization of B-Cyclodextrin Glucanotransferase from Bacillus Sp. Nd1. J. Basic Microbiol. 2019, 59, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Soumya, P.; Kochupurackal, J. Pineapple Peel Extract as an Effective Substrate for Esterase Production from Bacillus Subtilis E9. Curr. Microbiol. 2020, 77, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Adıgüzel, A.O. Production and Characterization of Thermo-, Halo-and Solvent-Stable Esterase from Bacillus Mojavensis Th309. Biocat. Biotransform. 2020, 38, 210–226. [Google Scholar] [CrossRef]
- Akeed, Y.; Atrash, F.; Naffaa, W. Partial Purification and Characterization of Chitinase Produced by Bacillus Licheniformis B307. Heliyon 2020, 6, e03858. [Google Scholar] [CrossRef]
- Martínez-Zavala, S.A.; Barboza-Pérez, U.E.; Hernández-Guzmán, G.; Bideshi, D.K.; Barboza-Corona, J.E. Chitinases of Bacillus Thuringiensis: Phylogeny, Modular Structure, and Applied Potentials. Front. Microbiol. 2020, 10, 3032. [Google Scholar] [CrossRef] [Green Version]
- Lekshmi, R.; Nisha, S.A.; Kaleeswaran, B.; Alfarhan, A. Pomegranate Peel Is a Low-Cost Substrate for the Production of Tannase by Bacillus Velezensis Ta3 under Solid State Fermentation. J. King Saud Univ.-Sci. 2020, 32, 1831–1837. [Google Scholar] [CrossRef]
- Mendoza, D.; Niño, L.; Gelves, G. Dynamic Modeling of Tannase Production from Bacillus Cereus: A Framework Simulation Based on Fed Batch Strategy. J. Phys. Conf. Ser. 2021, 2049, 012091. [Google Scholar] [CrossRef]
- Shakir, H.A.; Khan, M.; Irfan, M.; Ali, S.; Yousaf, M.A.; Javed, I.; Qazi, J.I.; Bukhari, S.S.I. Production and Characterization of Tannase by Bacillus Subtilis in Solid State Fermentation of Corn Leaves. J. Appl. Biotechnol. Rep. 2022, 9, 516–530. [Google Scholar]
- Chandrasekaran, R.; Revathi, K.; Senthil-Nathan, S.; Kalaivani, K.; Hunter, W.B.; Duraipandiyan, V.; Al-Dhabi, N.A.; Esmail, G.A. Eco-Friendly Formulation of Wild Bacillus Thuringiensis Secondary Metabolites through Molecular Characterization against the Lepidopteran Pest. Physiol. Mol. Plant Pathol. 2018, 101, 93–104. [Google Scholar] [CrossRef]
- Sansinenea, E. Bacillus Thuringiensis Biotechnology; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Bhalla, R.; Dalal, M.; Panguluri, S.K.; Jagadish, B.; Mandaokar, A.D.; Singh, A.; Kumar, P.A. Isolation, Characterization and Expression of a Novel Vegetative Insecticidal Protein Gene of Bacillus Thuringiensis. FEMS Microbiol. Lett. 2005, 243, 467–472. [Google Scholar] [CrossRef]
- Fedhila, S.; Gohar, M.; Slamti, L.; Nel, P.; Lereclus, D. The Bacillus thuringiensis PlcR-regulated gene inhA2 is necessary, but not sufficient, for virulence. J. Bacteriol. 2003, 185, 2820. [Google Scholar] [CrossRef] [Green Version]
- Okasov, A.; Kalimagambetov, A.; Akhmatullina, N.; Ilin, A. In Vitro Proteolytic Activation of Carcinolytic Parasporal Inclusions of Bacillus Thuringiensis Ssp. Israelensis Bacteria. Biol. Med. 2019, 5, 47–54. [Google Scholar] [CrossRef]
- de Maagd, R.A.; Bravo, A.; Crickmore, N. How Bacillus Thuringiensis Has Evolved Specific Toxins to Colonize the Insect World. Trends Genet. 2001, 17, 193–199. [Google Scholar] [CrossRef]
- Höfte, H.; Whiteley, H.R. Insecticidal Crystal Proteins of Bacillus Thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Gómez, I.; Conde, J.; Muñoz-Garay, C.; Sánchez, J.; Miranda, R.; Zhuang, M.; Gill, S.S.; Soberón, M. Oligomerization Triggers Binding of a Bacillus Thuringiensis Cry1ab Pore-Forming Toxin to Aminopeptidase N Receptor Leading to Insertion into Membrane Microdomains. Biochim. Biophys. Acta (BBA) Biomembr. 2004, 1667, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Candas, M.; Griko, N.B.; Taussig, R.; Bulla, L.A., Jr. A Mechanism of Cell Death Involving an Adenylyl Cyclase/Pka Signaling Pathway Is Induced by the Cry1ab Toxin of Bacillus Thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randhawa, G.J.; Singh, M.; Grover, M. Bioinformatic Analysis for Allergenicity Assessment of Bacillus Thuringiensis Cry Proteins Expressed in Insect-Resistant Food Crops. Food Chem. Toxicol. 2011, 49, 356–362. [Google Scholar] [CrossRef]
- Ahmad, M.; Sayyed, A.H.; Saleem, M.A.; Ahmad, M. Evidence for Field Evolved Resistance to Newer Insecticides in Spodoptera Litura (Lepidoptera: Noctuidae) from Pakistan. Crop Prot. 2008, 27, 1367–1372. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Z.; Zhang, J.; He, K.; Ferry, N.; Gatehouse, A.M.R. Cross-Resistance of Cry1ab-Selected Asian Corn Borer to Other Cry Toxins. J. Appl. Entomol. 2010, 134, 429–438. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Baxter, S.W.; Zhao, J.-Z.; Shelton, A.M.; Gould, F.; Tabashnik, B.E. The Diversity of Bt Resistance Genes in Species of Lepidoptera. J. Invertebr. Pathol. 2007, 95, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Różycki, H.; Dahm, H.; Strzelczyk, E.; Li, C.Y. Diazotrophic Bacteria in Root-Free Soil and in the Root Zone of Pine (Pinus sylvestris L.) and Oak (Quercus robur L.). Appl. Soil Ecol. 1999, 12, 239–250. [Google Scholar] [CrossRef]
- Achouak, W.; Normand, P.; Heulin, T. Comparative Phylogeny of Rrs and Nifh Genes in the Bacillaceae. Int. J. Syst. Evolut. Microbiol. 1999, 49, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Park, R.-Y.; Choi, M.-E.; Sun, H.-U.; Shin, S.-U. Production of Catechol-Siderophore and Utilization of Transferrin-Bound Iron in Bacillus Cereus. Biol. Pharm. Bull. 2005, 28, 1132–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzini, J.M.; Dunlap, C.A.; Bowman, M.J.; Chulze, S.N. Bacillus Velezensis Rc 218 as a Biocontrol Agent to Reduce Fusarium Head Blight and Deoxynivalenol Accumulation: Genome Sequencing and Secondary Metabolite Cluster Profiles. Microbiol. Res. 2016, 192, 30–36. [Google Scholar] [CrossRef]
- Handelsman, J.; Silo-Suh, L.A.; Stabb, E.V.; Raffel, S.J. Target Range of Zwittermicin a, an Aminopolyol Antibiotic from Bacillus Cereus. Curr. Microbiol. 1998, 37, 6–11. [Google Scholar]
- Wen, X.; Li, X.; Cheng, G.; Chen, S.; Luo, W.; Chen, H.; Hao, Z. Purification and Detection of Zwittermicin a from Bacillus Thuringiensis G03a. Int. J. Agric. Biol. 2018, 20, 1071–1079. [Google Scholar]
- Pérez-García, A.; Romero, D.; de Vicente, A. Plant Protection and Growth Stimulation by Microorganisms: Biotechnological Applications of Bacilli in Agriculture. Curr. Opin. Biotechnol. 2011, 22, 187–193. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Schisler, D.A. Genomic Analysis and Secondary Metabolite Production in Bacillus Amyloliquefaciens as 43.3: A Biocontrol Antagonist of Fusarium Head Blight. Biol. Control 2013, 64, 166–175. [Google Scholar] [CrossRef]
- Korenblum, E.; Sebastián, G.V.; Paiva, M.M.; Coutinho, C.M.L.M.; Magalhães, F.C.M.; Peyton, B.M.; Seldin, L. Action of Antimicrobial Substances Produced by Different Oil Reservoir Bacillus Strains against Biofilm Formation. Appl. Microbiol. Biotechnol. 2008, 79, 97–103. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J.A. Sulphate-Reducing Bacteria and Their Role in Corrosion of Ferrous Materials. In Sulphate-Reducing Bacteria Environmental and Engineered Systems; Hamilton, L.L., Barton, W.A., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 459–482. [Google Scholar]
- Djelal, H.; Chniti, S.; Jemni, M.; Weill, A.; Sayed, W.; Amrane, A. Identification of Strain Isolated from Dates (Phœnix dactylifera L.) for Enhancing Very High Gravity Ethanol Production. Environ. Sci. Pollut. Res. 2017, 24, 9886–9894. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, C.R. Microbial Polyketide Synthases: More and More Prolific. Proc. Natl. Acad. Sci. USA 1999, 96, 3336–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, T.; Wei, M.; Zhu, Z.; Liu, W.; Zhang, Z. Macrolactin a Is the Key Antibacterial Substance of Bacillus Amyloliquefaciens D2wm against the Pathogen Dickeya Chrysanthemi. Eur. J. Plant Pathol. 2019, 155, 393–404. [Google Scholar] [CrossRef]
- Im, S.M.; Yu, N.H.; Joen, H.W.; Kim, S.O.; Park, H.W.; Park, A.R.; Kim, J.-C. Biological Control of Tomato Bacterial Wilt by Oxydifficidin and Difficidin-Producing Bacillus Methylotrophicus Dr-08. Pestic. Biochem. Physiol. 2020, 163, 130–137. [Google Scholar] [CrossRef]
- Mondol, M.A.; Kim, J.H.; Lee, M.a.; Tareq, F.S.; Lee, H.; Lee, Y.; Shin, H.J. Ieodomycins a–D, Antimicrobial Fatty Acids from a Marine Bacillus sp. J. Nat. Prod. 2011, 74, 1606–1612. [Google Scholar] [CrossRef]
- Sauer, U.; Cameron, D.C.; Bailey, J.E. Metabolic Capacity of Bacillus Subtilis for the Production of Purine Nucleosides, Riboflavin, and Folic Acid. Biotechnol. Bioeng. 1998, 59, 227–238. [Google Scholar] [CrossRef]
- De Wulf, P.; Vandamme, E.J. Production of D-Ribose by Fermentation. Appl. Microbiol. Biotechnol. 1997, 48, 141–148. [Google Scholar] [CrossRef]
- Shih, I.-L.; Van, Y.-T. The Production of Poly-(Γ-Glutamic Acid) from Microorganisms and Its Various Applications. Bioresour. Technol. 2001, 79, 207–225. [Google Scholar] [CrossRef]
- Park, S.-B.; Sung, M.-H.; Uyama, H.; Han, D.K. Poly (Glutamic Acid): Production, Composites, and Medical Applications of the Next-Generation Biopolymer. Prog. Polym. Sci. 2021, 113, 101341. [Google Scholar] [CrossRef]
- Borhani, M.S.; Etemadifar, Z.; Jorjani, E. Application of Rotatable Central Composite Design for Optimization of Extracellular Alkaline Protease Production from Newly Isolated Bacillus Pseudofirmus Msb4. Biol. J. Microorg. 2017, 5, 17–31. [Google Scholar]
- Yokota, A.; Fujii, T.; Goto, K. Alicyclobacillus: Thermophilic Acidophilic Bacilli; Springer: Tokyo, Japan, 2007. [Google Scholar]
- Tianli, Y.; Jiangbo, Z.; Yahong, Y. Spoilage by Alicyclobacillus Bacteria in Juice and Beverage Products: Chemical, Physical, and Combined Control Methods. Compr. Rev. Food Sci. Food Saf. 2014, 13, 771–797. [Google Scholar] [CrossRef]
- Pillot, G.; Sunny, S.; Comes, V.; Kerzenmacher, S. Optimization of Growth and Electrosynthesis of Polyhydroxyalkanoates by the Thermophilic Bacterium Kyrpidia Spormannii. Bioresour. Technol. Rep. 2022, 17, 100949. [Google Scholar] [CrossRef]
- Bobadilla-Fazzini, R.A.; Cortés, M.P.; Maass, A.; Parada, P. Sulfobacillus Thermosulfidooxidans Strain Cutipay Enhances Chalcopyrite Bioleaching under Moderate Thermophilic Conditions in the Presence of Chloride Ion. AMB Express 2014, 4, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Z.-Y.; Gu, G.; Yang, H.; Qiu, G. Bioleaching of Pyrrhotite by Sulfobacillus Thermosulfidooxidans. J. Cent. South Univ. 2014, 21, 2638–2644. [Google Scholar] [CrossRef]
- Deng, S.; He, G.; Bichao, W.; Guohua, G. Pyrite-Promoted Dissolution of Arsenopyrite in the Presence of Sulfobacillus Thermosulfidooxidans. J. Mater. Res. Technol. 2020, 9, 9362–9371. [Google Scholar] [CrossRef]
- Ilyas, S.; Srivastava, R.R.; Kim, H.; Ilyas, N. Biotechnological Recycling of Hazardous Waste Pcbs Using Sulfobacillus Thermosulfidooxidans through Pretreatment of Toxicant Metals: Process Optimization and Kinetic Studies. Chemosphere 2022, 286, 131978. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, M.; Wang, J.; Lv, D.; Ma, Y.; Zhou, B.; Wang, B. Biocontrol Effects of Brevibacillus Laterosporus Amcc100017 on Potato Common Scab and Its Impact on Rhizosphere Bacterial Communities. Biol. Control 2017, 106, 89–98. [Google Scholar] [CrossRef]
- Joo, H.J.; Kim, H.; Kim, L.; Lee, S.; Ryu, J.-G.; Lee, T. A Brevibacillus Sp. Antagonistic to Mycotoxigenic fusarium spp. Biol. Control 2015, 87, 64–70. [Google Scholar] [CrossRef]
- Arumugam, T.; Kumar, P.S.; Gopinath, K.P. Hptlc Fingerprint Profile, In vitro Antioxidant and Evaluation of Antimicrobial Compound Produced from Brevibacillus brevis-Egs9 against Multidrug Resistant Staphylococcus Aureus. Microb. Pathog. 2017, 102, 166–172. [Google Scholar] [CrossRef]
- Yasar Yildiz, S.; Anzelmo, G.; Ozer, T.; Radchenkova, N.; Genc, S.; di Donato, P.; Nicolaus, B.; Oner, E.T.; Kambourova, M. Brevibacillus Themoruber: A Promising Microbial Cell Factory for Exopolysaccharide Production. J. Appl. Microbiol. 2014, 116, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Sankhla, I.S.; Bhati, R.; Singh, A.K.; Mallick, N. Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) Co-Polymer Production from a Local Isolate, Brevibacillus Invocatus Mtcc 9039. Bioresour. Technol. 2010, 101, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Nandanwar, H.S.; Vohra, R.M.; Hoondal, G.S. Trimeric L-N-Carbamoylase from Newly Isolated Brevibacillus Reuszeri Hsn1: A Potential Biocatalyst for Production of L-A-Amino Acids. Biotechnol. Appl. Biochem. 2013, 60, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Yin, H.; Peng, H.; Liu, Z.; Lu, G.; Dang, Z. Characteristics and Proteomic Analysis of Pyrene Degradation by Brevibacillus brevis in Liquid Medium. Chemosphere 2017, 178, 80–87. [Google Scholar] [CrossRef]
- Wei, K.; Yin, H.; Peng, H.; Lu, G.; Dang, Z. Bioremediation of Triphenyl Phosphate by Brevibacillus brevis: Degradation Characteristics and Role of Cytochrome P450 Monooxygenase. Sci. Total Environ. 2018, 627, 1389–1395. [Google Scholar] [CrossRef]
- Vivas, A.; Biro, B.; Nemeth, T.; Barea, J.M.; Azcon, R. Nickel-Tolerant Brevibacillus brevis and Arbuscular Mycorrhizal Fungus Can Reduce Metal Acquisition and Nickel Toxicity Effects in Plant Growing in Nickel Supplemented Soil. Soil Biology Biochem. 2006, 38, 2694–2704. [Google Scholar] [CrossRef]
- Shindhal, T.; Rakholiya, P.; Varjani, S.; Pandey, A.; Ngo, H.H.; Guo, W.; Ng, H.Y.; Taherzadeh, M.J. A Critical Review on Advances in the Practices and Perspectives for the Treatment of Dye Industry Wastewater. Bioengineered 2021, 12, 70–87. [Google Scholar] [CrossRef]
- Daud, N.S.; Din, A.R.J.M.; Rosli, M.A.; Azam, Z.M.; Othman, N.Z.; Sarmidi, M.R. Paenibacillus Polymyxa Bioactive Compounds for Agricultural and Biotechnological Applications. Biocatal. Agric. Biotechnol. 2019, 18, 101092. [Google Scholar] [CrossRef]
- Derikvand, P.; Etemadifar, Z.; Saber, H. Sulfur Removal from Dibenzothiophene by Newly Isolated Paenibacillus Validus Strain Pd2 and Process Optimization in Aqueous and Biphasic (Model-Oil) Systems. Pol. J. Microbiol. 2015, 64, 47–54. [Google Scholar] [CrossRef]
- Sar, T.; Chen, Y.; Bai, Y.; Liu, B.; Agarwal, P.; Stark, B.; Akbas, M. Combining Co-Culturing of Paenibacillus Strains and Vitreoscilla Hemoglobin Expression as a Strategy to Improve Biodesulfurization. Lett. Appl. Microbiol. 2021, 72, 484–494. [Google Scholar] [CrossRef]
- Sar, T.; Ozturk, M.; Stark, B.C.; Akbas, M.Y. Improvement in Desulfurization of Dibenzothiophene and Dibenzothiophene Sulfone by Paenibacillus Strains Using Immobilization or Nanoparticle Coating. J. Appl. Microbiol. 2022, 133, 1040–1051. [Google Scholar] [CrossRef]
- Davis, M.L.; Thoden, J.B.; Holden, H.M. The X-Ray Structure of Dtdp-4-Keto-6-Deoxy-D-Glucose-3,4-Ketoisomerase. J. Biol. Chem. 2007, 282, 19227–19236. [Google Scholar] [CrossRef] [Green Version]
- Berditsch, M.; Afonin, S.; Ulrich, A.S. The ability of Aneurinibacillus migulanus (Bacillus brevis) to produce the antibiotic gramicidin S is correlated with phenotype variation. Appl. Environ. Microbiol. 2007, 73, 6620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alenezi, F.N.; Rekik, I.; Bouket, A.C.; Luptakova, L.; Weitz, H.J.; Rateb, M.E.; Jaspars, M.; Woodward, S.; Belbahri, L. Increased Biological Activity of Aneurinibacillus migulanus Strains Correlates with the Production of New Gramicidin Secondary Metabolites. Front. Microbiol. 2017, 8, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowen, S.; Davies, K.G.; Pembroke, B. Potential Use of Pasteuria spp. in the Management of Plant Parasitic Nematodes. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 205–219. [Google Scholar]
- Talavera, M.; Mizukubo, T.; Ito, K.; Aiba, S. Effect of Spore Inoculum and Agricultural Practices on the Vertical Distribution of the Biocontrol Plant-Growth-Promoting Bacterium Pasteuria Penetrans and Growth of Meloidogyne Incognita-Infected Tomato. Biol. Fert. Soils 2002, 35, 435–440. [Google Scholar] [CrossRef]
- Zheng, Z.; Zheng, J.; Peng, D.; Sun, M. Complete Genome Sequence of Fictibacillus Arsenicus G25–54, a Strain with Toxicity to Nematodes. J. Biotechnol. 2017, 241, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Huang, K.; Tang, H.; Ni, J.; Liu, J.; Xu, P.; Tao, F. Steps toward High-Performance Pla: Economical Production of D-Lactate Enabled by a Newly Isolated Sporolactobacillus Terrae Strain. Biotechnol. J. 2019, 14, 1800656. [Google Scholar] [CrossRef]
- Thamacharoensuk, T.; Tolieng, V.; Thongchul, N.; Kodama, K.; Tanasupawat, S. Characterisation of Lactic Acid Producing Sporolactobacillus Strains from Tree Barks in Thailand. Ann. Microbiol. 2017, 67, 215–218. [Google Scholar] [CrossRef]
- Matsuo, Y.; Kanoh, K.; Yamori, T.; Kasai, H.; Katsuta, A.; Adachi, K.; Shin-ya, K.; Shizuri, Y. Urukthapelstatin a, a Novel Cytotoxic Substance from Marine-Derived Mechercharimyces Asporophorigenens Ym11–542. J. Antibiot. 2007, 60, 251. [Google Scholar] [CrossRef]
- Reijula, K.E. Two Bacteria Causing Farmer’s Lung: Fine Structure Ofthermoactinomyces Vulgaris Andsaccharopolyspora Rectivirgula. Mycopathologia 1993, 121, 143–147. [Google Scholar] [CrossRef]
- Steele, D.B.; Fiske, M.J.; Steele, B.P.; Kelley, V.C. Production of a Low-Molecular-Weight, Alkaline-Active, Thermostable Protease by a Novel, Spiral-Shaped Bacterium, Kurthia Spiroforme, Sp. Nov. Enzyme Microb. Technol. 1992, 14, 358–360. [Google Scholar] [CrossRef]
- Ruan, Z.; Zhou, S.; Jiang, S.; Sun, L.; Zhai, Y.; Wang, Y.; Chen, C.; Zhao, B. Isolation and Characterization of a Novel Cinosulfuron Degrading Kurthia Sp. From a Methanogenic Microbial Consortium. Bioresour. Technol. 2013, 147, 477–483. [Google Scholar] [CrossRef]
- Vreeland, R.H. Advances in Understanding the Biology of Halophilic Microorganisms; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Horikoshi, K.; Antranikian, G.; Bull, A.T.; Robb, F.T.; Stetter, K.O. Extremophiles Handbook; Springer: Tokyo, Japan, 2010. [Google Scholar]
- Seckbach, J.; Oren, A.; Stan-Lotter, H. Polyextremophiles: Life under Multiple Forms of Stress; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Chen, G.-Q.; Jiang, X.-R. Next Generation Industrial Biotechnology Based on Extremophilic Bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef]
- Singh, N.; Choudhury, B. Potential of Lentibacillus Sp. Ns12iitr for Production of Lipids with Enriched Branched-Chain Fatty Acids for Improving Biodiesel Properties Along with Hydrocarbon Co-Production. Extremophiles 2018, 22, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Sundararaman, A.; Srinivasan, S.; Lee, J.-H.; Lee, S.-S. Lentibacillus Alimentarius Sp. Nov., Isolated from Myeolchi-Jeotgal, a Traditional Korean High-Salt Fermented Anchovy. Antonie Van Leeuwenhoek 2018, 111, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Hui-Ying, Y. Purification and Characterization of an Extracellular Esterase with Organic Solvent Tolerance from a Halotolerant Isolate, Salimicrobiumsp. Ly19. BMC Biotechnol. 2013, 13, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagar, S.; Shaw, A.K.; Anand, S.; Celin, S.M.; Rai, P.K. Aerobic Biodegradation of Hmx by Planomicrobium Flavidum. 3 Biotech 2018, 8, 455. [Google Scholar] [CrossRef] [PubMed]
- Poosarla, V.G.; Chandra, T.S. Purification and Characterization of Novel Halo-Acid-Alkali-Thermo-Stable Xylanase from Gracilibacillus Sp. Tscpvg. Appl. Biochem. Biotechnol. 2014, 173, 1375–1390. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Safarpour, A.; Noghabi, K.A.; Bakhtiary, T.; Ventosa, A. Halophiles and Their Vast Potential in Biofuel Production. Front. Microbiol. 2019, 10, 1895. [Google Scholar] [CrossRef] [Green Version]
- Cosa, S.; Ugbenyen, A.M.; Mabinya, L.V.; Rumbold, K.; Okoh, A.I. Characterization and Flocculation Efficiency of a Bioflocculant Produced by a Marine Halobacillus. Environ. Technol. 2013, 34, 2671–2679. [Google Scholar] [CrossRef]
- Yang, L.; Tan, R.-X.; Wang, Q.; Huang, W.-Y.; Yin, Y.-X. Antifungal Cyclopeptides from Halobacillus Litoralis Ys3106 of Marine Origin. Tetrahedron Lett. 2002, 43, 6545–6548. [Google Scholar] [CrossRef]
- Gu, Z.J.; Wang, L.; Le Rudulier, D.; Zhang, B.; Yang, S.S. Erratum To: Characterization of the Glycine Betaine Biosynthetic Genes in the Moderately Halophilic Bacterium Halobacillus Dabanensis D-8t. Curr. Microbiol. 2009, 59, 665. [Google Scholar] [CrossRef] [Green Version]
- Rivadeneyra, M.A.; Párraga, J.; Delgado, R.; Ramos-Cormenzana, A.; Delgado, G. Biomineralization of Carbonates by Halobacillus Trueperi in Solid and Liquid Media with Different Salinities. FEMS Microbiol. Ecol. 2004, 48, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Rivadeneyra, M.-A.; Delgado, G.; Soriano, M.; Ramos-Cormenzana, A.; Delgado, R. Biomineralization of Carbonates by Marinococcus Albus and Marinococcus Halophilus Isolated from the Salar De Atacama (Chile). Curr. Microbiol. 1999, 39, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.-H.; Yuan, F.-W.; Chen, W.-C.; Chen, S.-Y. Production and Characterization of Ectoine by Marinococcus Sp. Ect1 Isolated from a High-Salinity Environment. J. Biosci. Bioeng. 2011, 111, 336–342. [Google Scholar] [CrossRef]
- Mudgil, D.; Baskar, S.; Baskar, R.; Paul, D.; Shouche, Y.S. Biomineralization Potential of Bacillus Subtilis, Rummeliibacillus Stabekisii and Staphylococcus Epidermidis Strains in Vitro Isolated from Speleothems, Khasi Hill Caves, Meghalaya, India. Geomicrobiol. J. 2018, 35, 675–694. [Google Scholar] [CrossRef]
- Fariq, A.; Yasmin, A.; Jamil, M. Production, Characterization and Antimicrobial Activities of Bio-Pigments by Aquisalibacillus Elongatus Mb592, Salinicoccus Sesuvii Mb597, and Halomonas Aquamarina Mb598 Isolated from Khewra Salt Range, Pakistan. Extremophiles 2019, 23, 435–449. [Google Scholar] [CrossRef]
- Rezaei, S.; Shahverdi, A.R.; Faramarzi, M.A. Isolation, One-Step Affinity Purification, and Characterization of a Polyextremotolerant Laccase from the Halophilic Bacterium Aquisalibacillus Elongatus and Its Application in the Delignification of Sugar Beet Pulp. Bioresour. Technol. 2017, 230, 67–75. [Google Scholar] [CrossRef]
- Rezaie, R.; Rezaei, S.; Jafari, N.; Forootanfar, H.; Khoshayand, M.R.; Faramarzi, M.A. Delignification and Detoxification of Peanut Shell Bio-Waste Using an Extremely Halophilic Laccase from an Aquisalibacillus Elongatus Isolate. Extremophiles 2017, 21, 993–1004. [Google Scholar] [CrossRef]
- Åqvist, J.; Isaksen, G.V.; Brandsdal, B.O. Computation of Enzyme Cold Adaptation. Nat. Rev. Chem. 2017, 1, 0051. [Google Scholar] [CrossRef]
- Radianingtyas, H.; Wright, P.C. Alcohol Dehydrogenases from Thermophilic and Hyperthermophilic Archaea and Bacteria. FEMS Microbiol. Rev. 2003, 27, 593–616. [Google Scholar] [CrossRef]
- Urbieta, M.S.; Donati, E.R.; Chan, K.G.; Shahar, S.; Sin, L.L.; Goh, K.M. Thermophiles in the Genomic Era: Biodiversity, Science, and Applications. Biotechnol. Adv. 2015, 33 Pt 1, 633–647. [Google Scholar] [CrossRef]
- Wrighton, K.; Agbo, P.; Warnecke, F.; Weber, K.; Brodie, E.; DeSantis, T.Z.; Hugenholtz, P.; Andersen, G.; Coates, J.D. A Novel Ecological Role of the Firmicutes Identified in Thermophilic Microbial Fuel Cells. ISME J. 2008, 2, 1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cripps, R.; Eley, K.; Leak, D.; Rudd, B.; Taylor, M.; Todd, M.; Boakes, S.; Martin, S.; Atkinson, T. Metabolic Engineering of Geobacillus Thermoglucosidasius for High Yield Ethanol Production. Metab. Eng. 2009, 11, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Bischoff, K.M.; Sani, R.K. Highly Thermostable Gh39 Β-Xylosidase from a Geobacillus Sp. Strain Wsucf1. BMC Biotechnol. 2014, 14, 963. [Google Scholar] [CrossRef] [Green Version]
- Mohr, T.; Aliyu, H.; Küchlin, R.; Polliack, S.; Zwick, M.; Neumann, A.; Cowan, D.; de Maayer, P. Co-Dependent Hydrogen Production by the Facultative Anaerobe Parageobacillus Thermoglucosidasius. Microb. Cell Factories 2018, 17, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliabadi, N.; Aminzadeh, S.; Karkhane, A.A.; Haghbeen, K. Thermostable Chitinase from Cohnella Sp. A01: Isolation and Product Optimization. Braz. J. Microbiol. 2016, 47, 931–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Sun, M.; Wu, J.; Ye, M.; Ge, X.; Wei, W.; Li, H.; Hu, F. Identification and Biochemical Characterization of a Novel Endo-Type Β-Agarase Agaw from Cohnella Sp. Strain Lgh. Appl. Microbiol. Biotechnol. 2015, 99, 10019–10029. [Google Scholar] [CrossRef] [PubMed]
- Rakotoarivonina, H.; Hermant, B.; Monthe, N.; Rémond, C. The Hemicellulolytic Enzyme Arsenal of Thermobacillus Xylanilyticus Depends on the Composition of Biomass Used for Growth. Microb. Cell Factories 2012, 11, 159. [Google Scholar] [CrossRef]
- Ahmad, S.; Scopes, R.K. Isolation and Properties of a Constitutive D-Xylulokinase from a Novel Thermophilic Saccharococcus Caldoxylosilyticus Dsm 12041 (Atcc 700356). Enzyme Microb. Technol. 2002, 30, 627–632. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, J.; Jia, M.; Guang, C.; Zhang, T.; Mu, W. Characterization of a Novel D-Lyxose Isomerase from Thermoflavimicrobium Dichotomicum and Its Application for D-Mannose Production. Process Biochem. 2019, 83, 131–136. [Google Scholar] [CrossRef]
- Tang, J.; Wang, Y.; Yang, G.; Luo, H.; Zhuang, L.; Yu, Z.; Zhou, S. Complete Genome Sequence of the Dissimilatory Azo Reducing Thermophilic Bacterium Novibacillus Thermophiles Sg-1. J. Biotechnol. 2018, 284, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhou, X.; Wang, Y.; Yang, G.; Zhou, S. Dissimilatory Azoreduction of Orange I by a Newly Isolated Moderately Thermophilic Bacterium, Novibacillus Thermophilus Sg-1. Biotechnol. Bioprocess Eng. 2015, 20, 1064–1070. [Google Scholar] [CrossRef]
- Henze, J.; Randall, D.G. Microbial Induced Calcium Carbonate Precipitation at Elevated Ph Values (>11) Using Sporosarcina Pasteurii. J. Environ. Chem. Eng. 2018, 6, 5008–5013. [Google Scholar] [CrossRef]
- Omoregie, A.I.; Ngu, L.H.; Ong, D.E.L.; Nissom, P.M. Low-Cost Cultivation of Sporosarcina Pasteurii Strain in Food-Grade Yeast Extract Medium for Microbially Induced Carbonate Precipitation (Micp) Application. Biocatal. Agric. Biotechnol. 2019, 17, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Omoregie, A.I.; Khoshdelnezamiha, G.; Senian, N.; Ong, D.E.L.; Nissom, P.M. Experimental Optimisation of Various Cultural Conditions on Urease Activity for Isolated Sporosarcina Pasteurii Strains and Evaluation of Their Biocement Potentials. Ecol. Eng. 2017, 109, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Ting, A.S.Y.; Tay, H.; Peh, K.L.; Tan, W.S.; Tee, C.S. Novel Isolation of Thermophilic Ureibacillus Terrenus from Compost of Empty Fruit Bunches (Efb) of Oil Palm and Its Enzymatic Activities. Biocatal. Agric. Biotechnol. 2013, 2, 162–164. [Google Scholar] [CrossRef]
- Juibari, M.M.; Yeganeh, L.P.; Abbasalizadeh, S.; Azarbaijani, R.; Mousavi, S.H.; Tabatabaei, M.; Jouzani, G.S.; Salekdeh, G.H. Investigation of a Hot-Spring Extremophilic Ureibacillus Thermosphaericus Strain Thermo-Bf for Extracellular Biosynthesis of Functionalized Gold Nanoparticles. BioNanoScience 2015, 5, 233–241. [Google Scholar] [CrossRef]
- Okuda, N.; Soneura, M.; Ninomiya, K.; Katakura, Y.; Shioya, S. Biological Detoxification of Waste House Wood Hydrolysate Using Ureibacillus Thermosphaericus for Bioethanol Production. J. Biosci. Bioeng. 2008, 106, 128–133. [Google Scholar] [CrossRef]
- Hatayama, K.; Shoun, H.; Ueda, Y.; Nakamura, A. Tuberibacillus Calidus Gen. Nov., Sp. Nov., Isolated from a Compost Pile and Reclassification of Bacillus Naganoensis Tomimura Et Al. 1990 as Pullulanibacillus Naganoensis Gen. Nov., Comb. Nov. And Bacillus Laevolacticus Andersch Et Al. 1994 as Sporolactobacillus Laevolacticus Comb. Nov. Int. J. Syst. Evol. Microbiol. 2006, 56 Pt 11, 2545–2551. [Google Scholar]
- Hogendoorn, C.; Pol, A.; Picone, N.; Cremers, G.; Van Alen, T.A.; Gagliano, A.L.; Jetten, M.S.; D’Alessandro, W.; Quatrini, P.; Op den Camp, H.J. Hydrogen and Carbon Monoxide-Utilizing Kyrpidia Spormannii Species from Pantelleria Island, Italy. Front. Microbiol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Reiner, J.E.; Jung, T.; Lapp, C.J.; Siedler, M.; Bunk, B.; Overmann, J.; Gescher, J. Kyrpidia Spormannii Sp. Nov., a Thermophilic, Hydrogen-Oxidizing, Facultative Autotroph, Isolated from Hydrothermal Systems at São Miguel Island, and Emended Description of the Genus Kyrpidia. Int. J. Syst. Evolut. Microbiol. 2018, 68, 3735–3740. [Google Scholar] [CrossRef]
- Hackbarth, M.; Jung, T.; Reiner, J.E.; Gescher, J.; Horn, H.; Hille-Reichel, A.; Wagner, M. Monitoring and Quantification of Bioelectrochemical Kyrpidia Spormannii Biofilm Development in a Novel Flow Cell Setup. Chem. Eng. J. 2020, 390, 124604. [Google Scholar] [CrossRef]
- Niimura, Y.; Koh, E.; Yanagida, F.; Suzuki, K.; Komagata, K.; Kozaki, M. Amphibacillus Xylanus Gen. Nov., Sp. Nov., a Facultatively Anaerobic Sporeforming Xylan-Digesting Bacterium Which Lacks Cytochrome, Quinone, and Catalase. Int. J. Syst. Evolut. Microbiol. 1990, 40, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Aino, K.; Narihiro, T.; Minamida, K.; Kamagata, Y.; Yoshimune, K.; Yumoto, I. Bacterial Community Characterization and Dynamics of Indigo Fermentation. FEMS Microbiol. Ecol. 2010, 74, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Aino, K.; Yumoto, I. Fermentibacillus Polygoni Gen. Nov., Sp. Nov., an Alkaliphile That Reduces Indigo Dye. Int. J. Syst. Evolut. Microbiol. 2016, 66, 2247–2253. [Google Scholar] [CrossRef]
- Murthy, P.S.; Madhumathi, M. Polyhydroxyalkanoate (Pha) Production from Amphibacillus sp. Biotechnol. Indian J. 2009, 3, 24–29. [Google Scholar]
- Ibrahim, A.S.S.; El-Tayeb, M.A.; Elbadawi, Y.B.; Al-Salamah, A.A.; Antranikian, G. Hexavalent Chromate Reduction by Alkaliphilic Amphibacillus Sp. Ksucr3 Is Mediated by Copper-Dependent Membrane-Associated Cr(Vi) Reductase. Extremophiles 2012, 16, 659–668. [Google Scholar] [CrossRef]
- Mesbah, N.M.; Wiegel, J. Halophilic Alkali- and Thermostable Amylase from a Novel Polyextremophilic Amphibacillus Sp. Nm-Ra2. Int. J. Biol. Macromol. 2014, 70, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Brininger, C.; Spradlin, S.; Cobani, L.; Evilia, C. The More Adaptive to Change, the More Likely You Are to Survive: Protein Adaptation in Extremophiles. Semin. Cell Dev. Biol. 2018, 84, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Kasana, R.C.; Pandey, C.B. Exiguobacterium: An Overview of a Versatile Genus with Potential in Industry and Agriculture. Crit. Rev. Biotechnol. 2018, 38, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Emtiazi, G.; Abari, A.H.; Rahehagh, F. Removal of Mtbe by Novel Exiguobacterium in Seawater. Desalination Water Treat. 2011, 28, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Weichler, M.T.; Kurteva-Yaneva, N.; Przybylski, D.; Schuster, J.; Müller, R.H.; Harms, H.; Rohwerder, T. Thermophilic coenzyme B12-dependent acyl coenzyme A (CoA) mutase from Kyrpidia tusciae DSM 2912 preferentially catalyzes isomerization of (R)-3-hydroxybutyryl-CoA and 2-hydroxyisobutyryl-CoA. Appl. Environ. Microbiol. 2015, 81, 4564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harirchi, S.; Etemadifar, Z.; Yazdian, F.; Taherzadeh, M.J. Efficacy of Polyextremophilic Aeribacillus Pallidus on Bioprocessing of Beet Vinasse Derived from Ethanol Industries. Bioresour. Technol. 2020, 313, 123662. [Google Scholar] [CrossRef]
- Harirchi, S.; Etemadifar, Z.; Mahboubi, A.; Yazdian, F.; Taherzadeh, M.J. The Effect of Calcium/Magnesium Ratio on the Biomass Production of a Novel Thermoalkaliphilic Aeribacillus Pallidus Strain with Highly Heat-Resistant Spores. Curr. Microbiol. 2020, 77, 2565–2574. [Google Scholar] [CrossRef]
- Radchenkova, N.; Vassilev, S.; Martinov, M.; Kuncheva, M.; Panchev, I.; Vlaev, S.; Kambourova, M. Optimization of the Aeration and Agitation Speed of Aeribacillus Palidus 418 Exopolysaccharide Production and the Emulsifying Properties of the Product. Process Biochem. 2014, 49, 576–582. [Google Scholar] [CrossRef]
- Zheng, C.; Li, Z.; Su, J.; Zhang, R.; Liu, C.; Zhao, M. Characterization and Emulsifying Property of a Novel Bioemulsifier by Aeribacillus Pallidus Ym-1. J. Appl. Microbiol. 2012, 113, 44–51. [Google Scholar] [CrossRef]
- Radchenkova, N.; Panchev, I.; Vassilev, S.; Kuncheva, M.; Dobreva, S.; Kambourova, M. Continuous Cultivation of a Thermophilic Bacterium Aeribacillus Pallidus 418 for Production of an Exopolysaccharide Applicable in Cosmetic Creams. J. Appl. Microbiol. 2015, 119, 1301–1309. [Google Scholar] [CrossRef] [Green Version]
- Başbülbül, G.; Ozteber, M.; Bıyık, H.H.; Erman, O.; Bozdoğan, B. Presence of Enta and Entb Genes among Thermophilic Bacteria Isolated from Hot Springs in Turkey. Acta Microbiol. Bulg. 2018, 34, 25–31. [Google Scholar]
- Yasawong, M.; Areekit, S.; Pakpitchareon, A.; Santiwatanakul, S.; Chansiri, K. Characterization of Thermophilic Halotolerant Aeribacillus Pallidus Td1 from Tao Dam Hot Spring, Thailand. Int. J. Mol. Sci. 2011, 12, 5294–5303. [Google Scholar] [CrossRef] [Green Version]
- Bose, H.; Satyanarayana, T. Suitability of the Alkalistable Carbonic Anhydrase from a Polyextremophilic Bacterium Aeribacillus Pallidus Tshb1 in Biomimetic Carbon Sequestration. Bioprocess Biosyst. Eng. 2016, 39, 1515–1525. [Google Scholar] [CrossRef]
- Tourova, T.P. Detection of N-Alkane Biodegradation Genes Alkb and Lada in Thermophilic Hydrocarbon-Oxidizing Bacteria of the Genera Aeribacillus and Geobacillus. Microbiology 2016, 85, 693–707. [Google Scholar] [CrossRef]
- Mehetre, G.T.; Dastager, S.G.; Dharne, M.S. Biodegradation of Mixed Polycyclic Aromatic Hydrocarbons by Pure and Mixed Cultures of Biosurfactant Producing Thermophilic and Thermo-Tolerant Bacteria. Sci. Total Environ. 2019, 679, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, V.; Baltaci, M.O.; Ozgencli, I.; Sisecioglu, M.; Adiguzel, A.; Adiguzel, G. Purification and Biochemical Characterization of a Novel Thermostable Serine Alkaline Protease from Aeribacillus Pallidus C10: A Potential Additive for Detergents. J. Enzyme Inhib. Med. Chem. 2017, 32, 468–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechri, S.; Berrouina, M.B.E.; Benmrad, M.O.; Jaouadi, N.Z.; Rekik, H.; Moujehed, E.; Chebbi, A.; Sayadi, S.; Chamkha, M.; Bejar, S.; et al. Characterization of a Novel Protease from Aeribacillus Pallidus Strain Vp3 with Potential Biotechnological Interest. Int. J. Biol. Macromol. 2017, 94, 221–232. [Google Scholar] [CrossRef]
- Dongdong, Z.; Yi, W.; Chunfang, Z.; Dan, Z.; Peng, G.; Zongjun, C. Characterization of a Thermophilic Lignocellulose-Degrading Microbial Consortium with High Extracellular Xylanase Activity. J. Microbiol. Biotechnol. 2018, 28, 305–313. [Google Scholar]
- EFSA Panel on Food Contact Materials, Enzymes, Processing Aids; Silano, V.; Baviera, J.M.B.; Bolognesi, C.; Brüschweiler, B.J.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; et al. Safety Evaluation of the Food Enzyme 4-A-Glucanotransferase from Aeribacillus pallidus (Strain ae-Sas). EFSA J. 2019, 17, e05628. [Google Scholar]
- Ma, Y.; Zhong, H.; He, Z. Cr(Vi) Reductase Activity Locates in the Cytoplasm of Aeribacillus Pallidus Bk1, a Novel Cr(Vi)-Reducing Thermophile Isolated from Tengchong Geothermal Region, China. Chem. Eng. J. 2019, 371, 524–534. [Google Scholar] [CrossRef]
- Samaei-Nouroozi, A.; Rezaei, S.; Khoshnevis, N.; Doosti, M.; Hajihoseini, R.; Khoshayand, M.R.; Faramarzi, M.A. Medium-Based Optimization of an Organic Solvent-Tolerant Extracellular Lipase from the Isolated Halophilic Alkalibacillus Salilacus. Extremophiles 2015, 19, 933–947. [Google Scholar] [CrossRef]
- Abdel-Hamed, A.R.; Abo-Elmatty, D.M.; Wiegel, J.; Mesbah, N.M. Biochemical Characterization of a Halophilic, Alkalithermophilic Protease from Alkalibacillus Sp. Nm-Da2. Extremophiles 2016, 20, 885–894. [Google Scholar] [CrossRef]
- Bergmann, S.; David, F.; Franco-Lara, E.; Wittmann, C.; Krull, R. Ectoine Production by Alkalibacillus Haloalkaliphilus—Bioprocess Development Using Response Surface Methodology and Model-Driven Strategies. Eng. Life Sci. 2013, 13, 399–407. [Google Scholar] [CrossRef]
- Goh, K.M.; Kahar, U.M.; Chai, Y.Y.; Chong, C.S.; Chai, K.P.; Ranjani, V.; Illias, R.; Chan, K.G. Recent Discoveries and Applications of Anoxybacillus. Appl. Microbiol. Biotechnol. 2013, 97, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Cabrol, L.; Marone, A.; Tapia-Venegas, E.; Steyer, J.-P.; Ruiz-Filippi, G.; Trably, E. Microbial Ecology of Fermentative Hydrogen Producing Bioprocesses: Useful Insights for Driving the Ecosystem Function. FEMS Microbiol. Rev. 2017, 41, 158–181. [Google Scholar] [CrossRef] [PubMed]
- Stolz, J.F.; Oremland, R.S. Bacterial Respiration of Arsenic and Selenium. FEMS Microbiol. Rev. 1999, 23, 615–627. [Google Scholar] [CrossRef]
- Cui, K.; Wu, Q.; Xu, Y. Biodegradation of Ethyl Carbamate and Urea with Lysinibacillus Sphaericus Mt33 in Chinese Liquor Fermentation. J. Agric. Food Chem. 2018, 66, 1583–1590. [Google Scholar] [CrossRef]
- Mohamed, E.A.H.; Farag, A.G. Arsenic Removal from Aqueous Solutions by Different Bacillus and Lysinibacillus Species. Bioremed. J. 2015, 19, 269–276. [Google Scholar] [CrossRef]
- Marilena Stancu, M. Investigation of the Organic Solvent Resistance Mechanisms in Rhodococcus and Lysinibacillus Using Several Molecular Forensic Tools. Environ. Forensics 2015, 16, 242–256. [Google Scholar] [CrossRef]
- Mokashe, N.; Chaudhari, B.; Patil, U. Detergent-Compatible Robust Alkaline Protease from Newly Isolated Halotolerant Salinicoccus Sp. Un-12. J. Surfactants Deterg. 2017, 20, 1377–1393. [Google Scholar] [CrossRef]
- Mokashe, N.; Chaudhari, A.; Patil, U. Optimal Production and Characterization of Alkaline Protease from Newly Isolated Halotolerant Jeotgalicoccus sp. Biocatal. Agric. Biotechnol. 2015, 4, 235–243. [Google Scholar] [CrossRef]
- Mazhar, S.; Hill, C.; McAuliffe, O. Chapter One—The Genus Macrococcus: An Insight into Its Biology, Evolution, and Relationship with Staphylococcus. In Advances in Applied Microbiology; Gadd, G.M., Sariaslani, S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–50. [Google Scholar]
- Abdel-Aziz, M.; Abou-El-Sherbini, K.S.; Hamzawy, E.; Amr, M.H.A.; El-Dafrawy, S. Green Synthesis of Silver Nano-Particles by Macrococcus Bovicus and Its Immobilization onto Montmorillonite Clay for Antimicrobial Functionality. Appl. Biochem. Biotechnol. 2015, 176, 2225–2241. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Han, Y.; Sheng, S.; Meng, Q.; Wang, Z.; Liu, W.; Di, L. Evaluation of a Novel Thermophilic Nitrifying Bacillaceae Species Aliibacillus Thermotolerans Bm62t Promoting Nitrogen Retention in Livestock Manure Compost. Environ. Progress Sustain. Energy 2020, 39, e13357. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.-H.; Men, M.-Q.; Meng, Q.-X.; Xu, B.-S. Aliibacillus Thermotolerans Gen. Nov., Sp. Nov.: A Thermophilic and Heterotrophic Ammonia-Oxidizing Bacterium from Compost. Arch. Microbiol. 2018, 200, 1357–1363. [Google Scholar] [CrossRef]
- Schmidt, M.; Stougaard, P. Identification, Cloning and Expression of a Cold-Active Β-Galactosidase from a Novel Arctic Bacterium, Alkalilactibacillus Ikkense. Environ. Technol. 2010, 31, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Priemé, A.; Johansen, A.; Stougaard, P. Alkalilactibacillus Ikkensis, Gen. Nov., Sp. Nov., a Novel Enzyme-Producing Bacterium from a Cold and Alkaline Environment in Greenland. Extremophiles 2012, 16, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Chuprom, J.; Bovornreungroj, P.; Ahmad, M.; Kantachote, D.; Enomoto, T. Statistical Optimization for the Improved Production of an Extracellular Alkaline Nuclease by Halotolerant Allobacillus Halotolerans Msp69: Scale-up Approach and Its Potential as Flavor Enhancer of Fish Sauce. Biocatal. Agric. Biotechnol. 2016, 8, 236–247. [Google Scholar] [CrossRef]
- Ghatge, S.; Yang, Y.; Song, W.-Y.; Kim, T.-Y.; Hur, H.-G. A Novel Laccase from Thermoalkaliphilic Bacterium Caldalkalibacillus Thermarum Strain Ta2.A1 Able to Catalyze Dimerization of a Lignin Model Compound. Appl. Microbiol. Biotechnol. 2018, 102, 4075–4086. [Google Scholar] [CrossRef]
- de Jong, S.I.; Broek, M.A.V.D.; Merkel, A.Y.; Cortes, P.D.L.T.; Kalamorz, F.; Cook, G.M.; van Loosdrecht, M.C.M.; McMillan, D.G.G. Genomic Analysis of Caldalkalibacillus Thermarum Ta2.A1 Reveals Aerobic Alkaliphilic Metabolism and Evolutionary Hallmarks Linking Alkaliphilic Bacteria and Plant Life. Extremophiles 2020, 24, 923–935. [Google Scholar] [CrossRef]
- Hiraga, K.; Nishikata, Y.; Namwong, S.; Tanasupawat, S.; Takada, K.; Oda, K. Purification and Characterization of Serine Proteinase from a Halophilic Bacterium, Filobacillus Sp. Rf2–5. Biosci. Biotechnol. Biochem. 2005, 69, 38–44. [Google Scholar] [CrossRef]
- Calandrelli, V.; Gambacorta, A.; Romano, I.; Carratore, V.; Lama, L. A Novel Thermo-Alkali Stable Catalase–Peroxidase from Oceanobacillus Oncorhynchi Subsp. Incaldaniensis: Purification and Characterization. World J. Microbiol. Biotechnol. 2008, 24, 2269. [Google Scholar] [CrossRef]
- Romano, I.; Lama, L.; Nicolaus, B.; Poli, A.; Gambacorta, A.; Giordano, A. Oceanobacillus Oncorhynchi Subsp. Incaldanensis Subsp. Nov., an Alkalitolerant Halophile Isolated from an Algal Mat Collected from a Sulfurous Spring in Campania (Italy), and Emended Description of Oceanobacillus Oncorhynchi. Int. J. Syst. Evolut. Microbiol. 2006, 56, 805–810. [Google Scholar] [CrossRef]
- Seghal Kiran, G.; Lipton, A.N.; Kennedy, J.; Dobson, A.D.W.; Selvin, J. A Halotolerant Thermostable Lipase from the Marine Bacterium Oceanobacillus Sp. Pumb02 with an Ability to Disrupt Bacterial Biofilms. Bioengineered 2014, 5, 305–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Nishita, M.; Matsuyama, H.; Yumoto, I. Paralkalibacillus Indicireducens Gen., Nov., Sp. Nov., an Indigo-Reducing Obligate Alkaliphile Isolated from Indigo Fermentation Liquor Used for Dyeing. Int. J. Syst. Evol. Microbiol. 2017, 67, 4050–4056. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lu, K.; Dong, X.; Xie, W. Virgibacillus Sp. Sk37 and Staphylococcus Nepalensis Js11 as Potential Starters to Improve Taste of Shrimp Paste. LWT 2022, 154, 112657. [Google Scholar] [CrossRef]
- Udomsil, N.; Chen, S.; Rodtong, S.; Yongsawatdigul, J. Improvement of Fish Sauce Quality by Combined Inoculation of Tetragenococcus Halophilus Ms33 and Virgibacillus Sp. Sk37. Food Control 2017, 73, 930–938. [Google Scholar] [CrossRef]
- Lapsongphon, N.; Yongsawatdigul, J. Production and Purification of Antioxidant Peptides from a Mungbean Meal Hydrolysate by Virgibacillus Sp. Sk37 Proteinase. Food Chem. 2013, 141, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Lomthong, T.; Lertwattanasakul, N.; Kitpreechavanich, V. Production of Raw Starch Degrading Enzyme by the Thermophilic Filamentous Bacterium Laceyella Sacchari Lp175 and Its Application for Ethanol Production from Dried Cassava Chips. Starch Stärke 2016, 68, 1264–1274. [Google Scholar] [CrossRef]
- Shibasaki, H.; Uchimura, K.; Miura, T.; Kobayashi, T.; Usami, R.; Horikoshi, K. Highly Thermostable and Surfactant-Activated Chitinase from a Subseafloor Bacterium, Laceyella Putida. Appl. Microbiol. Biotechnol. 2014, 98, 7845–7853. [Google Scholar] [CrossRef]
- Suresh, M.; Renugadevi, B.; Brammavidhya, S.; Iyapparaj, P.; Anantharaman, P. Antibacterial Activity of Red Pigment Produced by Halolactibacillus Alkaliphilus Msrd1—An Isolate from Seaweed. Appl. Biochem. Biotechnol. 2015, 176, 185–195. [Google Scholar] [CrossRef]
- Arun, J.; Selvakumar, S.; Sathishkumar, R.; Moovendhan, M.; Ananthan, G.; Maruthiah, T.; Palavesam, A. In Vitro Antioxidant Activities of an Exopolysaccharide from a Salt Pan Bacterium Halolactibacillus Miurensis. Carbohydr. Polym. 2017, 155, 400–406. [Google Scholar] [CrossRef]
- Zhang, K.; Su, L.; Wu, J. Recent Advances in Recombinant Protein Production by Bacillus Subtilis. Annu. Rev. Food Sci. Technol. 2020, 11, 295–318. [Google Scholar] [CrossRef] [Green Version]
- van Tilburg, A.Y.; Cao, H.; van der Meulen, S.B.; Solopova, A.; Kuipers, O.P. Metabolic Engineering and Synthetic Biology Employing Lactococcus lactis and Bacillus subtilis Cell Factories. Curr. Opin. Biotechnol. 2019, 59, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, K.; Yokoyama, S. Trends in the Application of Bacillus in Fermented Foods. Curr. Opin. Biotechnol. 2019, 56, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-Q.; Liu, D.-Y.; Chen, T.; Wang, Z.-W. Recent Advances in Crispr/Cas9 Mediated Genome Editing in Bacillus Subtilis. World J. Microbiol. Biotechnol. 2018, 34, 153. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Bhatia, S.K.; Park, H.A.; Kim, S.Y.; Sudheer, P.D.V.N.; Yang, Y.-H.; Choi, K.-Y. Bacillus Subtilis as a Robust Host for Biochemical Production Utilizing Biomass. Crit. Rev. Biotechnol. 2021, 41, 827–848. [Google Scholar] [CrossRef] [PubMed]
- Aristidou, A.; Penttilä, M. Metabolic Engineering Applications to Renewable Resource Utilization. Curr. Opin. Biotechnol. 2000, 11, 187–198. [Google Scholar] [CrossRef]
- Bhavsar, A.P.; Zhao, X.; Brown, E.D. Development and Characterization of a Xylose-Dependent System for Expression of Cloned Genes in Bacillus Subtilis: Conditional Complementation of a Teichoic Acid Mutant. Appl. Environ. Microbiol. 2001, 67, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, M.; Hollenberg, C.P. Nucleotide Sequence of the Bacillus Subtilis Xylose Isomerase Gene: Extensive Homology between the Bacillus and Escherichia Coli Enzyme. Nucleic Acids Res. 1985, 13, 5717–5722. [Google Scholar] [CrossRef] [Green Version]
- Krispin, O.; Allmansberger, R. The Bacillus Subtilis Arae Protein Displays a Broad Substrate Specificity for Several Different Sugars. J. Bacteriol. 1998, 180, 3250–3252. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-C.; Jun, S.Y.; Seo, J.-H. Construction and Characterization of Recombinant Bacillus Subtilis Jy123 Able to Transport Xylose Efficiently. J. Bacteriol. 2012, 161, 402–406. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.; Fu, J.; Li, N.; Wang, Z.; Tang, Y.; Chen, T. Production of Acetoin through Simultaneous Utilization of Glucose, Xylose, and Arabinose by Engineered Bacillus Subtilis. PLoS ONE 2016, 11, e0159298. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huang, C.; Yu, X.; Liu, S.; Wang, Z.; Tang, Y.-J.; Chen, T. Highly Efficient Hemicellulose Utilization for Acetoin Production by an Engineered Bacillus Subtilis. J. Chem. Technol. Biotechnol. 2018, 93, 3428–3435. [Google Scholar] [CrossRef]
- Chen, T.; Liu, W.; Fu, J.; Zhang, B.; Tang, Y. Engineering Bacillus Subtilis for Acetoin Production from Glucose and Xylose Mixtures. J. Bacteriol. 2013, 168, 499–505. [Google Scholar] [CrossRef]
- Sun, J.-D.; Tang, C.; Zhou, J.; Wei, P.; Wang, Y.-J.; An, W.; Yan, Z.-Y.; Yong, X.-Y. Production of Poly-Γ-Glutamic Acid (Γ-Pga) from Xylose-Glucose Mixtures by Bacillus Amyloliquefaciens C1. 3 Biotech 2021, 11, 100. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, H.; Chen, S.; Qi, G. Production of Optically Pure 2,3-Butanediol from Miscanthus Floridulus Hydrolysate Using Engineered Bacillus Licheniformis Strains. World J. Microbiol. Biotechnol. 2018, 34, 66. [Google Scholar] [CrossRef] [PubMed]
- Cal, A.J.; Kibblewhite, R.E.; Sikkema, W.D.; Torres, L.F.; Hart-Cooper, W.M.; Orts, W.J.; Lee, C.C. Production of Polyhydroxyalkanoate Copolymers Containing 4-Hydroxybutyrate in Engineered Bacillus Megaterium. Int. J. Biol. Macromol. 2021, 168, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Ashiuchi, M. Microbial Production and Chemical Transformation of Poly-Γ-Glutamate. Microb. Biotechnol. 2013, 6, 664–674. [Google Scholar] [CrossRef]
- Halmschlag, B.; Putri, S.P.; Fukusaki, E.; Blank, L.M. Poly-Γ-Glutamic Acid Production by Bacillus Subtilis 168 Using Glucose as the Sole Carbon Source: A Metabolomic Analysis. J. Biosci. Bioeng. 2020, 130, 272–282. [Google Scholar] [CrossRef]
- Cai, D.; Chen, Y.; He, P.; Wang, S.; Mo, F.; Li, X.; Wang, Q.; Nomura, C.T.; Wen, Z.; Ma, X.; et al. Enhanced Production of Poly-Γ-Glutamic Acid by Improving Atp Supply in Metabolically Engineered Bacillus Licheniformis. Biotechnol. Bioeng. 2018, 115, 2541–2553. [Google Scholar] [CrossRef]
- Stark, B.C.; Dikshit, K.L.; Pagilla, K.R. The Biochemistry of Vitreoscilla Hemoglobin. Computational and structural. Biotechnol. J. 2012, 3, e201210002. [Google Scholar]
- Wang, Q.; Yu, H.; Wang, M.; Yang, H.; Shen, Z. Enhanced Biosynthesis and Characterization of Surfactin Isoforms with Engineered Bacillus Subtilis through Promoter Replacement and Vitreoscilla Hemoglobin Co-Expression. Process Biochem. 2018, 70, 36–44. [Google Scholar] [CrossRef]
- Jung, J.; Yu, K.O.; Ramzi, A.B.; Choe, S.H.; Kim, S.W.; Han, S.O. Improvement of Surfactin Production in Bacillus Subtilis Using Synthetic Wastewater by Overexpression of Specific Extracellular Signaling Peptides, Comx and Phrc. Biotechnol. Bioeng. 2012, 109, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Akdoğan, M.; Çelik, E. Enhanced Production of Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) Biopolymer by Recombinant Bacillus Megaterium in Fed-Batch Bioreactors. Bioprocess Biosyst. Eng. 2021, 44, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Biswas, R.; Yamaoka, M.; Nakayama, H.; Kondo, T.; Yoshida, K.-I.; Bisaria, V.S.; Kondo, A. Enhanced Production of 2,3-Butanediol by Engineered Bacillus Subtilis. Appl. Microbiol. Biotechnol. 2012, 94, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.Y.; Park, J.M.; Hong, W.-K.; Kim, Y.S.; Jung, Y.R.; Kim, S.-B.; Heo, S.-Y.; Lee, S.-M.; Kang, J.Y.; Oh, B.-R.; et al. Enhanced Production of 2,3-Butanediol by a Genetically Engineered Bacillus Sp. Brc1 Using a Hydrolysate of Empty Palm Fruit Bunches. Bioprocess Biosyst. Eng. 2015, 38, 299–305. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhang, J.; Li, L.; Wen, Z.; Nomura, C.T.; Wu, S.; Chen, S. Engineering Bacillus Licheniformis for the Production of Meso-2,3-Butanediol. Biotechnol. Biofuels 2016, 9, 117. [Google Scholar] [CrossRef]
- Fu, J.; Huo, G.; Feng, L.; Mao, Y.; Wang, Z.; Ma, H.; Chen, T.; Zhao, X. Metabolic Engineering of Bacillus Subtilis for Chiral Pure Meso-2,3-Butanediol Production. Biotechnol. Biofuels 2016, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Drejer, E.B.; Chan, D.T.C.; Haupka, C.; Wendisch, V.F.; Brautaset, T.; Irla, M. Methanol-Based Acetoin Production by Genetically Engineered Bacillus Methanolicus. Green Chem. 2020, 22, 788–802. [Google Scholar] [CrossRef]
- Kim, E.-J.; Seo, D.; Choi, K.-Y. Bioalcohol Production from Spent Coffee Grounds and Okara Waste Biomass by Engineered Bacillus Subtilis. Biomass Convers. Biorefin. 2020, 10, 167–173. [Google Scholar] [CrossRef]
- Kogan, G.; Šoltés, L.; Stern, R.; Gemeiner, P. Hyaluronic Acid: A Natural Biopolymer with a Broad Range of Biomedical and Industrial Applications. Biotechnol. Lett. 2007, 29, 17–25. [Google Scholar] [CrossRef]
- Li, Y.; Shi, Z.; Shao, Y.; Wu, M.; Li, G.; Ma, T. Temperature-Controlled Molecular Weight of Hyaluronic Acid Produced by Engineered Bacillus Subtilis. Biotechnol. Lett. 2021, 43, 271–277. [Google Scholar] [CrossRef]
- Li, Y.; Li, G.; Zhao, X.; Shao, Y.; Wu, M.; Ma, T. Regulation of Hyaluronic Acid Molecular Weight and Titer by Temperature in Engineered Bacillus Subtilis. 3 Biotech 2019, 9, 225. [Google Scholar] [CrossRef] [PubMed]
- Widner, B.; Behr, R.; Von Dollen, S.; Tang, M.; Heu, T.; Sloma, A.; Sternberg, D.; DeAngelis, P.L.; Weigel, P.H.; Brown, S. Hyaluronic Acid Production in Bacillus Subtilis. Appl. Environ. Microbiol. 2005, 71, 3747–3752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, L.-J.; Lee, C. Enhanced Hyaluronic Acid Production in Bacillus subtilis by Coexpressing Bacterial Hemoglobin. Biotechnol. Prog. 2007, 23, 1017–1022. [Google Scholar] [PubMed]
- Westbrook, A.W.; Ren, X.; Oh, J.; Moo-Young, M.; Chou, C.P. Metabolic Engineering to Enhance Heterologous Production of Hyaluronic Acid in Bacillus Subtilis. Metab. Eng. 2018, 47, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Kang, Z.; Yuan, P.; Du, G.; Chen, J. Production of Specific-Molecular-Weight Hyaluronan by Metabolically Engineered Bacillus Subtilis 168. Metab. Eng. 2016, 35, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Liang, Z.; Li, H.; Chen, C.; Xue, Y.; Du, Q. Biosynthesis of Low-Molecular-Weight Mannan Using Metabolically Engineered Bacillus Subtilis 168. Carbohydr. Polym. 2021, 251, 117115. [Google Scholar] [CrossRef]
- Jin, P.; Zhang, L.; Yuan, P.; Kang, Z.; Du, G.; Chen, J. Efficient Biosynthesis of Polysaccharides Chondroitin and Heparosan by Metabolically Engineered Bacillus Subtilis. Carbohydr. Polym. 2016, 140, 424–432. [Google Scholar] [CrossRef]
- Yang, X.-M.; Wang, S.S. Phase-Specific Optimization of Multiple Endotoxin-Protein Production with Genetically Engineered Bacillus Thuringiensis. Biotechnol. Appl. Biochem. 2000, 31, 71–76. [Google Scholar] [CrossRef]
- Huang, D.-F.; Zhang, J.; Song, F.-P.; Lang, Z.-H. Microbial Control and Biotechnology Research on Bacillus Thuringiensis in China. J. Invertebr. Pathol. 2007, 95, 175–180. [Google Scholar] [CrossRef]
- Sanchis, V.; Agaisse, H.; Chaufaux, J.; Lereclus, D. A Recombinase-Mediated System for Elimination of Antibiotic Resistance Gene Markers from Genetically Engineered Bacillus Thuringiensis Strains. Appl. Environ. Microbiol. 1997, 63, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Song, F.; Shu, C.; Liu, J.; Liu, C.; Huang, D.; Feng, S.; Zhang, J. An Engineered Bacillus Thuringiensis Strain with Insecticidal Activity against Scarabaeidae (Anomala Corpulenta) and Chrysomelidae (Leptinotarsa Decemlineata and Colaphellus Bowringi). Biotechnol. Lett. 2009, 31, 697. [Google Scholar] [CrossRef] [PubMed]
- Lertcanawanichakul, M.; Wiwat, C.; Bhumiratana, A.; Dean, D.H. Expression of Chitinase-Encoding Genes in Bacillus Thuringiensis and Toxicity of Engineered B. Thuringiensis Subsp. Aizawai toward Lymantria Dispar Larvae. Curr. Microbiol. 2004, 48, 175–181. [Google Scholar] [CrossRef] [PubMed]
- El-Menofy, W.; Osman, G.; Assaeedi, A.; Salama, M. A Novel Recombinant Baculovirus Overexpressing a Bacillus Thuringiensis Cry1ab Toxin Enhances Insecticidal Activity. Biol. Proced. Online 2014, 16, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Pang, Y.; Tang, M.; Xie, R.; Tan, L.; Zeng, S.; Yuan, M.; Liu, J. Highly Toxic and Broad-Spectrum Insecticidal Bacillus Thuringiensis Engineered by Using the Transposon Tn917 and Protoplast Fusion. Curr. Microbiol. 2001, 43, 112–119. [Google Scholar] [CrossRef]
- Doruk, T.; Avican, U.; Camci, I.Y.; Gedik, S.T. Overexpression of Polyphosphate Kinase Gene (Ppk) Increases Bioinsecticide Production by Bacillus Thuringiensis. Microbiol. Res. 2013, 168, 199–203. [Google Scholar] [CrossRef]
- Donker, H.J.L. Bijdrage Tot De Kennis Der Boterzuur-, Butylalcohol-En Acetongistingen; Meinema: Delft, The Netherlands, 1926. [Google Scholar]
- Oshima, T.; Moriya, T. A Preliminary Analysis of Microbial and Biochemical Properties of High-Temperature Compost. Ann. N. Y. Acad. Sci. 2008, 1125, 338–344. [Google Scholar] [CrossRef]
- Kolinko, S.; Wu, Y.-W.; Tachea, F.; Denzel, E.; Hiras, J.; Gabriel, R.; Bäcker, N.; Chan, L.J.G.; Eichorst, S.A.; Frey, D.; et al. A Bacterial Pioneer Produces Cellulase Complexes That Persist through Community Succession. Nat. Microbiol. 2018, 3, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Lal, D.; Khan, F.; Gupta, S.K.; Schumann, P.; Lal, R. Edaphobacillus Lindanitolerans Gen. Nov., Sp. Nov., Isolated from Hexachlorocyclohexane (Hch) Contaminated Soil. J. Basic Microbiol. 2013, 53, 758–765. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Li, M.; Du, Q.; Li, Q.; Ma, P. Jilinibacillus Soli Gen. Nov., Sp. Nov., a Novel Member of the Family Bacillaceae. Arch. Microbiol. 2015, 197, 11–16. [Google Scholar] [CrossRef]
- Pringsheim, E.G.; Robinow, C.F.; Ainsworth, G.C.; Brown, A.M.; Marsden, P.S.S.F.; Smith, P.A.; Spilsbury, J.F. Observations on Two Very Large Bacteria, Caryophanon Latum Peshkoff and Lineola Longa (Nomen Provisorium). Microbiology 1947, 1, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Sefrji, F.O.; Marasco, R.; Michoud, G.; Seferji, K.A.; Merlino, G.; Daffonchio, D. Insights into the Cultivable Bacterial Fraction of Sediments from the Red Sea Mangroves and Physiological, Chemotaxonomic, and Genomic Characterization of Mangrovibacillus Cuniculi Gen. Nov., Sp. Nov., a Novel Member of the Bacillaceae Family. Front. Microbiol. 2022, 13, 77798. [Google Scholar] [CrossRef] [PubMed]
- Alou, M.T.; Rathored, J.; Lagier, J.-C.; Khelaifia, S.; Labas, N.; Sokhna, C.; Diallo, A.; Raoult, D.; Dubourg, G. Massilibacterium Senegalense Gen. Nov., Sp. Nov., a New Bacterial Genus Isolated from the Human Gut. New Microbes New Infect. 2016, 10, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsi, R.H.; Wiedmann, M. Characteristics and Distribution of Listeria Spp., Including Listeria Species Newly Described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alou, M.T.; Nguyen, T.-T.; Armstrong, N.; Rathored, J.; Khelaifia, S.; Raoult, D.; Fournier, P.-E.; Lagier, J.-C. Numidum Massiliense Gen. Nov., Sp. Nov., a New Member of the Bacillaceae Family Isolated from the Human Gut. New Microbes New Infect. 2016, 12, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alou, M.T.; Rathored, J.; Lagier, J.-C.; Khelaifia, S.; Michelle, C.; Sokhna, C.; Diallo, A.; Fournier, P.-E.; Raoult, D.; Edouard, S. Rubeoparvulum Massiliense Gen. Nov., Sp. Nov., a New Bacterial Genus Isolated from the Human Gut of a Senegalese Infant with Severe Acute Malnutrition. New Microbes New Infect. 2017, 15, 49–60. [Google Scholar] [CrossRef]
- Ping, W.; Zhou, D.; Sun, J.; Fan, C.; Ding, Y. A New Genus of Oral Bacteria in Human. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 1998, 38, 146–151. [Google Scholar]
- Tetz, G.; Tetz, V. Draft Genome Sequence of Tetzosporium hominis Vt-49 Gen. Nov., Sp. Nov., Isolated from the Dental Decay Plaque of a Patient with Periodontitis. Genome Announc. 2018, 6, e01541-17. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Yuan, C.; Xiao, M.; Tian, X.; Khan, I.; Kim, C.; Zhi, X.; Li, W. Abyssicoccus Albus Gen. Nov., Sp. Nov., a Novel Member of the Family Staphylococcaceae Isolated from Marine Sediment of the Indian Ocean. Antonie Van Leeuwenhoek 2016, 109, 1153–1160. [Google Scholar] [CrossRef]
- Miñana-Galbis, D.; Pinzón, D.L.; Lorén, J.G.; Manresa, À.; Oliart-Ros, R.M. Reclassification of Geobacillus Pallidus (Scholz Et Al. 1988) Banat Et Al. 2004 as Aeribacillus Pallidus Gen. Nov., Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2010, 60, 1600–1604. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chunyu, W.-X.; Jiang, G.-Q.; Huang, L.; Zhang, B.; Ji, Y.; Ding, Z.-G.; Yin, M.; Tang, S.-K. Aidingibacillus Halophilus Gen. Nov., Sp. Nov., a Novel Member of the Family Bacillaceae. Antonie Van Leeuwenhoek 2018, 111, 601–608. [Google Scholar] [CrossRef]
- Wisotzkey, J.D.; Jurtshuk, P.; Fox, G.E.; Deinhard, G.; Poralla, K. Comparative Sequence Analyses on the 16s Rrna (Rdna) of Bacillus Acidocaldarius, Bacillus Acidoterrestris, and Bacillus Cycloheptanicus and Proposal for Creation of a New Genus, Alicyclobacillus Gen. Nov. Int. J. Syst. Evolut. Microbiol. 1992, 42, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Amoozegar, M.A.; Bagheri, M.; Makhdoumi-Kakhki, A.; Didari, M.; Schumann, P.; Nikou, M.M.; Sánchez-Porro, C.; Ventosa, A. Aliicoccus Persicus Gen. Nov., Sp. Nov., a Halophilic Member of the Firmicutes Isolated from a Hypersaline Lake. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 6, 1964–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, C.O.; Lim, J.-M.; Lee, J.-M.; Xu, L.-H.; Jiang, C.-L.; Kim, C.-J. Reclassification of Bacillus Haloalkaliphilus Fritze 1996 as Alkalibacillus Haloalkaliphilus Gen. Nov., Comb. Nov. And the Description of Alkalibacillus Salilacus Sp. Nov., a Novel Halophilic Bacterium Isolated from a Salt Lake in China. Int. J. Syst. Evolut. Microbiol. 2005, 55, 1891–1896. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Thite, S.; Karodi, P.; Joseph, N.; Lodha, T. Alkalihalobacterium Elongatum Gen. Nov. Sp. Nov.: An Antibiotic-Producing Bacterium Isolated from Lonar Lake and Reclassification of the Genus Alkalihalobacillus into Seven Novel Genera. Front. Microbiol. 2021, 12, 722369. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lu, W.; Zhang, S.; Liu, K.; Yan, Y.; Li, J. Reclassification of Bacillus Saliphilus as Alkalicoccus Saliphilus Gen. Nov., Comb. Nov., and Description of Alkalicoccus Halolimnae Sp. Nov., a Moderately Halophilic Bacterium Isolated from a Salt Lake. Int. J. Syst. Evolut. Microbiol. 2017, 67, 1557–1563. [Google Scholar] [CrossRef]
- Sheu, S.-Y.; Arun, A.B.; Jiang, S.-R.; Young, C.-C.; Chen, W.-M. Allobacillus Halotolerans Gen. Nov., Sp. Nov. Isolated from Shrimp Paste. Int. J. Syst. Evolut. Microbiol. 2011, 61, 1023–1027. [Google Scholar] [CrossRef]
- Didari, M.; Amoozegar, M.A.; Bagheri, M.; Schumann, P.; Spröer, C.; Sánchez-Porro, C.; Ventosa, A. Alteribacillus Bidgolensis Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium from a Hypersaline Lake, and Reclassification of Bacillus Persepolensis as Alteribacillus Persepolensis Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 11, 2691–2697. [Google Scholar] [CrossRef] [Green Version]
- Sakai, M.; Deguchi, D.; Hosoda, A.; Kawauchi, T.; Ikenaga, M. Ammoniibacillus Agariperforans Gen. Nov., Sp. Nov., a Thermophilic, Agar-Degrading Bacterium Isolated from Compost. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 2, 570–577. [Google Scholar] [CrossRef]
- Zaitsev, G.M.; Tsitko, I.V.; Rainey, F.A.; Trotsenko, Y.A.; Uotila, J.S.; Stackebrandt, E.; Salkinoja-Salonen, M.S. New Aerobic Ammonium-Dependent Obligately Oxalotrophic Bacteria: Description of Ammoniphilus Oxalaticus Gen. Nov., Sp. Nov. And Ammoniphilus Oxalivorans Gen. Nov., Sp. Nov. Int. J. Syst. Evolut. Microbiol. 1998, 48, 151–163. [Google Scholar] [CrossRef]
- Zavarzina, D.G.; Tourova, T.P.; Kolganova, T.V.; Boulygina, E.S.; Zhilina, T.N. Description of Anaerobacillus Alkalilacustre Gen. Nov., Sp. Nov.—Strictly Anaerobic Diazotrophic Bacillus Isolated from Soda Lake and Transfer of Bacillus Arseniciselenatis, Bacillus Macyae, and Bacillus Alkalidiazotrophicus to Anaerobacillus as the New Combinations A. Arseniciselenatis Comb. Nov., A. Macyae Comb. Nov., and A. Alkalidiazotrophicus Comb. Nov. Microbiology 2009, 78, 723–731. [Google Scholar]
- Shida, O.; Takagi, H.; Kadowaki, K.; Komagata, K. Proposal for Two New Genera, Brevibacillus Gen. Nov. And Aneurinibacillus Gen. Nov. Int. J. Syst. Evolut. Microbiol. 1996, 46, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikuta, E.; Lysenko, A.; Chuvilskaya, N.; Mendrock, U.; Hippe, H.; Suzina, N.; Nikitin, D.; Osipov, G.; Laurinavichius, K. Anoxybacillus Pushchinensis Gen. Nov., Sp. Nov., a Novel Anaerobic, Alkaliphilic, Moderately Thermophilic Bacterium from Manure, and Description of Anoxybacillus Flavitherms Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2000, 50, 2109–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amoozegar, M.A.; Bagheri, M.; Didari, M.; Mehrshad, M.; Schumann, P.; Spröer, C.; Sánchez-Porro, C.; Ventosa, A. Aquibacillus Halophilus Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium from a Hypersaline Lake, and Reclassification of Virgibacillus Koreensis as Aquibacillus Koreensis Comb. Nov. And Virgibacillus Albus as Aquibacillus Albus Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 11, 3616–3623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez, M.C.; Carrasco, I.J.; Xue, Y.; Ma, Y.; Cowan, D.; Jones, B.; Grant, W.D.; Ventosa, A. Aquisalibacillus Elongatus Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium of the Family Bacillaceae Isolated from a Saline Lake. Int. J. Syst. Evolut. Microbiol. 2008, 58, 1922–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liang, J.; Zhang, Z.; Yu, M.; Wang, M.; Zhang, X.-H. Aureibacillus Halotolerans Gen. Nov., Sp. Nov., Isolated from Marine Sediment. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 11, 3950–3958. [Google Scholar] [CrossRef] [PubMed]
- Cohn, F. Untersuchungen Über Bacterien: I; JU Kern: Toky, Japan, 1875. [Google Scholar]
- Guan, X.; Liu, C.; Fang, B.; Zhao, J.; Jin, P.; Li, J.; Jia, F.; Wang, X.; Xiang, W. Baia Soyae Gen. Nov., Sp. Nov., a Mesophilic Representative of the Family Thermoactinomycetaceae, Isolated from Soybean Root [Glycine Max (L.) Merr]. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 10, 3754–3760. [Google Scholar] [CrossRef]
- Manorama, R.; Pindi, P.K.; Reddy, G.S.N.; Shivaji, S. Bhargavaea Cecembensis Gen. Nov., Sp. Nov., Isolated from the Chagos–Laccadive Ridge System in the Indian Ocean. Int. J. Syst. Evolut. Microbiol. 2009, 59, 2618–2623. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Jones, D. Brochothrix, a New Genus Tentatively Placed in the Family Lactobacillaceae. Int. J. Syst. Evolut. Microbiol. 1976, 26, 102–104. [Google Scholar]
- Tsujimoto, Y.; Saito, R.; Furuya, H.; Ishihara, D.; Sahara, T.; Kimura, N.; Nishino, T.; Tsuruoka, N.; Shigeri, Y.; Watanabe, K. Caenibacillus Caldisaponilyticus Gen. Nov., Sp. Nov., a Thermophilic, Spore-Forming and Phospholipid-Degrading Bacterium Isolated from Acidulocompost. Int. J. Syst. Evolut. Microbiol. 2016, 66, 2684–2690. [Google Scholar] [CrossRef] [Green Version]
- Min, U.-G.; Kim, S.-J.; Hong, H.; Kim, S.-G.; Gwak, J.-H.; Jung, M.-Y.; Kim, J.-G.; Na, J.-G.; Rhee, S.-K. Calculibacillus Koreensis Gen. Nov., Sp. Nov., an Anaerobic Fe(Iii)-Reducing Bacterium Isolated from Sediment of Mine Tailings. J. Microbiol. 2016, 54, 413–419. [Google Scholar] [CrossRef]
- Xue, Y.; Zhang, X.; Zhou, C.; Zhao, Y.; Cowan, D.; Heaphy, S.; Grant, W.D.; Jones, B.; Ventosa, A.; Ma, Y. Caldalkalibacillus Thermarum Gen. Nov., Sp. Nov., a Novel Alkalithermophilic Bacterium from a Hot Spring in China. Int. J. Syst. Evolut. Microbiol. 2006, 56, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Coorevits, A.; Dinsdale, A.E.; Halket, G.; Lebbe, L.; de Vos, P.; van Landschoot, A.; Logan, N.A. Taxonomic Revision of the Genus Geobacillus: Emendation of Geobacillus, G. Stearothermophilus, G. Jurassicus, G. Toebii, G. Thermodenitrificans and G. Thermoglucosidans (Nom. Corrig., Formerly ‘Thermoglucosidasius’); Transfer of Bacillus Thermantarcticus to the Genus as G. Thermantarcticus Comb. Nov.; Proposal of Caldibacillus Debilis Gen. Nov., Comb. Nov.; Transfer of G. Tepidamans to Anoxybacillus as A. Tepidamans Comb. Nov.; and Proposal of Anoxybacillus Caldiproteolyticus Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 7, 1470–1485. [Google Scholar]
- Moriya, T.; Hikota, T.; Yumoto, I.; Ito, T.; Terui, Y.; Yamagishi, A.; Oshima, T. Calditerricola Satsumensis Gen. Nov., Sp. Nov. And Calditerricola Yamamurae Sp. Nov., Extreme Thermophiles Isolated from a High-Temperature Compost. Int. J. Syst. Evolut. Microbiol. 2011, 61, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Adiguzel, A.; Ay, H.; Baltaci, M.O.; Akbulut, S.; Albayrak, S.; Omeroglu, M.A. Genome-Based Classification of Calidifontibacillus Erzurumensis Gen. Nov., Sp. Nov., Isolated from a Hot Spring in Turkey, with Reclassification of Bacillus Azotoformans as Calidifontibacillus Azotoformans Comb. Nov. And Bacillus Oryziterrae as Calidifontibacillus Oryziterrae Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2020, 70, 6418–6427. [Google Scholar]
- Lin, P.; Yan, Z.; Yi, T.-H. Camelliibacillus Cellulosilyticus Gen. Nov., Sp. Nov., a Cellulose-Degrading Bacterium Isolated from Tea. Int. J. Syst. Evolut. Microbiol. 2018, 68, 1867–1873. [Google Scholar] [CrossRef] [PubMed]
- Peshkoff, M.A. Cytology, Karyology and Cycle of Development of New Microbes, Caryophanon Latum and Caryophanon Tenue. Compt. Rend.(Doklady) Acad. Sci. l’URRS 1939, 25, 244–247. [Google Scholar]
- Nakamura, K.; Haruta, S.; Ueno, S.; Ishii, M.; Yokota, A.; Igarashi, Y. Cerasibacillus Quisquiliarum Gen. Nov., Sp. Nov., Isolated from a Semi-Continuous Decomposing System of Kitchen Refuse. Int. J. Syst. Evolut. Microbiol. 2004, 54, 1063–1069. [Google Scholar] [CrossRef]
- Cao, W.-R.; Guo, L.-Y.; Du, Z.-J.; Das, A.; Saren, G.; Jiang, M.-Y.; Dunlap, C.A.; Rooney, A.P.; Yu, X.-K.; Li, T.-G. Chengkuizengella Sediminis Gen. Nov. Sp. Nov., Isolated from Sediment. Int. J. Syst. Evolut. Microbiol. 2017, 67, 2672–2678. [Google Scholar] [CrossRef]
- Arora, P.K.; Chauhan, A.; Pant, B.; Korpole, S.; Mayilraj, S.; Jain, R.K. Chryseomicrobium Imtechense Gen. Nov., Sp. Nov., a New Member of the Family Planococcaceae. Int. J. Syst. Evolut. Microbiol. 2011, 61, 1859–1864. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Traiwan, J.; Park, M.; Jung, M.Y.; Oh, S.; Yoon, J.; Sukhoom, A. Chungangia Koreensis Gen. Nov., Sp. Nov., Isolated from Marine Sediment. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 8, 1914–1920. [Google Scholar] [CrossRef] [Green Version]
- Kämpfer, P.; Rosselló-Mora, R.; Falsen, E.; Busse, H.; Tindall, B.J. Cohnella Thermotolerans Gen. Nov., Sp. Nov., and Classification of ‘Paenibacillus Hongkongensis’ as Cohnella Hongkongensis Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2006, 56, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wen, J.; Yang, G.; Liu, J.; Zhou, S. Compostibacillus Humi Gen. Nov., Sp. Nov., a Member of the Family Bacillaceae, Isolated from Sludge Compost. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 2, 346–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, S.; Xue, H.; Chang, J.; Guo, L.; Yang, X. Corticicoccus Populi Gen. Nov., Sp. Nov., a Member of the Family Staphylococcaceae, Isolated from Symptomatic Bark of Populus × Euramericana Canker. Int. J. Syst. Evolut. Microbiol. 2017, 67, 789–794. [Google Scholar] [CrossRef]
- Hatayama, K.; Kuno, T. Croceifilum Oryzae Gen. Nov., Sp. Nov., Isolated from Rice Paddy Soil. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 11, 4061–4065. [Google Scholar] [CrossRef]
- Bhatt, H.; Begum, M.A.; Chintalapati, S.; Chintalapati, V.R.; Singh, S.P. Desertibacillus Haloalkaliphilus Gen. Nov., Sp. Nov., Isolated from a Saline Desert. Int. J. Syst. Evolut. Microbiol. 2017, 67, 4435–4442. [Google Scholar] [CrossRef] [PubMed]
- Yassin, A.F.; Hupfer, H.; Klenk, H.-P.; Siering, C. Desmospora Activa Gen. Nov., Sp. Nov., a Thermoactinomycete Isolated from Sputum of a Patient with Suspected Pulmonary Tuberculosis, and Emended Description of the Family Thermoactinomycetaceae Matsuo Et Al. 2006. Int. J. Syst. Evolut. Microbiol. 2009, 59, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Tourova, T.P.; Sukhacheva, M.V.; Muyzer, G. Desulfuribacillus Alkaliarsenatis Gen. Nov. Sp. Nov., a Deep-Lineage, Obligately Anaerobic, Dissimilatory Sulfur and Arsenate-Reducing, Haloalkaliphilic Representative of the Order Bacillales from Soda Lakes. Extremophiles 2012, 16, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Seiler, H.; Wenning, M.; Scherer, S. Domibacillus Robiginosus Gen. Nov., Sp. Nov., Isolated from a Pharmaceutical Clean Room. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 6, 2054–2061. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Kojima, H.; Fukui, M. Proposal of Effusibacillus Lacus Gen. Nov., Sp. Nov., and Reclassification of Alicyclobacillus Pohliae as Effusibacillus Pohliae Comb. Nov. And Alicyclobacillus Consociatus as Effusibacillus Consociatus Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 8, 2770–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.D.; Lund, B.M.; Farrow, J.A.E.; Schleifer, K.H. Chemotaxonomic Study of an Alkalophilic Bacterium, Exiguobacterium Aurantiacum Gen. Nov., Sp. Nov. Microbiology 1983, 129, 2037–2042. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xu, J.; Xu, L.; Tindall, B.J. Tindall. Falsibacillus Pallidus to Replace the Homonym Bacillus Pallidus Zhou Et Al. 2008. Int. J. Syst. Evolut. Microbiol. 2009, 59, 3176–3180. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.P.; Dott, W.; Busse, H.; Kämpfer, P. Fictibacillus Phosphorivorans Gen. Nov., Sp. Nov. And Proposal to Reclassify Bacillus Arsenicus, Bacillus Barbaricus, Bacillus Macauensis, Bacillus Nanhaiensis, Bacillus Rigui, Bacillus Solisalsi and Bacillus Gelatini in the Genus Fictibacillus. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 8, 2934–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesner, H.; A Lawson, P.; Collins, M.D.; Weiss, N.; Wehmeyer, U.; Völker, H.; Thomm, M. Filobacillus Milensis Gen. Nov., Sp. Nov., a New Halophilic Spore-Forming Bacterium with Orn-D-Glu-Type Peptidoglycan. Int. J. Syst. Evolut. Microbiol. 2001, 51, 425–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, P.; Krishnamurthi, S.; Bhattacharya, A.; Sharma, R.; Chakrabarti, T. Fontibacillus Aquaticus Gen. Nov., Sp. Nov., Isolated from a Warm Spring. Int. J. Syst. Evolut. Microbiol. 2010, 60, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Berger, U. Neisseria Haemolysans (Thjotta and Boe, 1938): Studies on Its Place in the System. Z. Fur Hyg. Und Infekt. Med. Mikrobiol. Immunol. Und Virol. 1960, 146, 253–259. [Google Scholar]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.; E Ivanova, A.; Lysenko, A.M.; Petrunyaka, V.V.; A Osipov, G.; Belyaev, S.S.; et al. Taxonomic Study of Aerobic Thermophilic Bacilli: Descriptions of Geobacillus Subterraneus Gen. Nov., Sp. Nov. And Geobacillus Uzenensis Sp. Nov. From Petroleum Reservoirs and Transfer of Bacillus Stearothermophilus, Bacillus Thermocatenulatus, Bacillus Thermoleovorans, Bacillus Kaustophilus, Bacillus Thermodenitrificans to Geobacillus as the New Combinations G. Stearothermophilus, G. Th. Int. J. Syst. Evolut. Microbiol. 2001, 51, 433–446. [Google Scholar]
- Echigo, A.; Minegishi, H.; Mizuki, T.; Kamekura, M.; Usami, R. Geomicrobium Halophilum Gen. Nov., Sp. Nov., a Moderately Halophilic and Alkaliphilic Bacterium Isolated from Soil. Int. J. Syst. Evolut. Microbiol. 2010, 60, 990–995. [Google Scholar] [CrossRef] [Green Version]
- Zhou, E.-M.; Yu, T.-T.; Liu, L.; Ming, H.; Yin, Y.-R.; Dong, L.; Tseng, M.; Nie, G.-X.; Li, W.-J. Geothermomicrobium Terrae Gen. Nov., Sp. Nov., a Novel Member of the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 9, 2998–3004. [Google Scholar] [CrossRef] [Green Version]
- Keita, M.B.; Padhmanabhan, R.; Caputo, A.; Robert, C.; Delaporte, E.; Raoult, D.; Fournier, P.-E.; Bittar, F. Non-Contiguous Finished Genome Sequence and Description of Gorillibacterium Massiliense Gen. Nov, Sp. Nov., a New Member of the Family Paenibacillaceae. Stand. Genomic Sci. 2014, 9, 807–820. [Google Scholar] [CrossRef]
- Wainø, M.; Tindall, B.J.; Schumann, P.; Ingvorsen, K. Gracilibacillus Gen. Nov., with Description of Gracilibacillus Halotolerans Gen. Nov., Sp. Nov.; Transfer of Bacillus Dipsosauri to Gracilibacillus Dipsosauri Comb. Nov., and Bacillus Salexigens to the Genus Salibacillus Gen. Nov., as Salibacillus Salexigens Comb. Nov. Int. J. Syst. Evolut. Microbiol. 1999, 49, 821–831. [Google Scholar]
- Echigo, A.; Fukushima, T.; Mizuki, T.; Kamekura, M.; Usami, R. Halalkalibacillus Halophilus Gen. Nov., Sp. Nov., a Novel Moderately Halophilic and Alkaliphilic Bacterium Isolated from a Non-Saline Soil Sample in Japan. Int. J. Syst. Evolut. Microbiol. 2007, 57, 1081–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spring, S.; Ludwig, W.; Marquez, M.C.; Ventosa, A.; Schleifer, K.-H. Halobacillus Gen. Nov., with Descriptions of Halobacillus Litoralis Sp. Nov. And Halobacillus Trueperi Sp. Nov., and Transfer of Sporosarcina Halophila to Halobacillus Halophilus Comb. Nov. Int. J. Syst. Evolut. Microbiol. 1996, 46, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Nakajima, K.; Itamiya, Y.; Furukawa, S.; Yamamoto, Y.; Yamasato, K. Halolactibacillus Halophilus Gen. Nov., Sp. Nov. And Halolactibacillus Miurensis Sp. Nov., Halophilic and Alkaliphilic Marine Lactic Acid Bacteria Constituting a Phylogenetic Lineage in Bacillus Rrna Group 1. Int. J. Syst. Evolut. Microbiol. 2005, 55, 2427–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buss, S.N.; Cole, J.A.; Hannett, G.E.; Nazarian, E.J.; Nazarian, L.; Coorevits, A.; van Landschoot, A.; de Vos, P.; Schumann, P.; Musser, K.A.; et al. Hazenella Coriacea Gen. Nov., Sp. Nov., Isolated from Clinical Specimens. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 11, 4087–4093. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Glaeser, S.P.; Busse, H.-J. Transfer of Bacillus Schlegelii to a Novel Genus and Proposal of Hydrogenibacillus Schlegelii Gen. Nov., Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 5, 1723–1727. [Google Scholar] [CrossRef] [Green Version]
- Pal, Y.; Mayilraj, S.; Paul, M.; Schumann, P.; Krishnamurthi, S. Indiicoccus Explosivorum Gen. Nov., Sp. Nov., Isolated from an Explosives Waste Contaminated Site. Int. J. Syst. Evolut. Microbiol. 2019, 69, 2555–2564. [Google Scholar] [CrossRef]
- Yoon, J.H.; Weiss, N.; Lee, K.C.; Lee, I.S.; Kang, K.H.; Park, Y.H. Jeotgalibacillus Alimentarius Gen. Nov., Sp. Nov., a Novel Bacterium Isolated from Jeotgal with L-Lysine in the Cell Wall, and Reclassification of Bacillus Marinus Rüger 1983. As Mmrinibacillus Marinus Gen Nov., Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2001, 51, 2087–2093. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Lee, K.; Weiss, N.; Kang, K.H.; Park, Y. Jeotgalicoccus Halotolerans Gen. Nov., Sp. Nov. And Jeotgalicoccus Psychrophilus Sp. Nov., Isolated from the Traditional Korean Fermented Seafood Jeotgal. Int. J. Syst. Evolut. Microbiol. 2003, 53, 595–602. [Google Scholar] [CrossRef] [Green Version]
- Von Jan, M.; Riegger, N.; Pötter, G.; Schumann, P.; Verbarg, S.; Spröer, C.; Rohde, M.; Lauer, B.; Labeda, D.P.; Klenk, H.-P. Kroppenstedtia Eburnea Gen. Nov., Sp. Nov., a Thermoactinomycete Isolated by Environmental Screening, and Emended Description of the Family Thermoactinomycetaceae Matsuo Et Al. 2006 Emend. Yassin Et Al. 2009. Int. J. Syst. Evolut. Microbiol. 2011, 61, 2304–2310. [Google Scholar] [CrossRef]
- Trevisan, V. Caratteri Di Alcuni Nuovi Generi Di Batteriacee. Atti Della Accad. Fisio-Med.-Stat. Milano Ser. 1885, 4, 92–107. [Google Scholar]
- Klenk, H.-P.; Lapidus, A.; Chertkov, O.; Copeland, A.; Del Rio, T.G.; Nolan, M.; Lucas, S.; Chen, F.; Tice, H.; Cheng, J.-F.; et al. Complete Genome Sequence of the Thermophilic, Hydrogen-Oxidizing Bacillus Tusciae Type Strain (T2t) and Reclassification in the New Genus, Kyrpidia Gen. Nov. As Kyrpidia Tusciae Comb. Nov. And Emendation of the Family Alicyclobacillaceae Da Costa and Rainey, 2010. Stand. Genomic Sci. 2011, 5, 121–134. [Google Scholar] [PubMed] [Green Version]
- Yoon, J.-H.; Kim, I.-G.; Shin, Y.-K.; Park, Y.-H. Proposal of the Genus Thermoactinomyces Sensu Stricto and Three New Genera, Laceyella, Thermoflavimicrobium and Seinonella, on the Basis of Phenotypic, Phylogenetic and Chemotaxonomic Analyses. Int. J. Syst. Evolut. Microbiol. 2005, 55, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.-H.; Kang, K.H.; Park, Y. Lentibacillus Salicampi Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium Isolated from a Salt Field in Korea. Int. J. Syst. Evolut. Microbiol. 2002, 52, 2043–2048. [Google Scholar]
- Yu, T.-T.; Zhang, B.-H.; Yao, J.-C.; Tang, S.-K.; Zhou, E.-M.; Yin, Y.-R.; Wei, D.-Q.; Ming, H.; Li, W.-J. Lihuaxuella Thermophila Gen. Nov., Sp. Nov., Isolated from a Geothermal Soil Sample in Tengchong, Yunnan, South-West China. Antonie Van Leeuwenhoek 2012, 102, 711–718. [Google Scholar] [CrossRef]
- Pirie, J. The Genus Listerella. Pirie Sci. 1940, 91, 383. [Google Scholar] [CrossRef]
- Mahmoud, H.; Eapen, S.; Al-Bajjali, F.; Al-Qattan, A.; Jose, L. Litoribacterium Kuwaitense Gen. Nov., Sp. Nov., Isolated from a Kuwait Tidal Flat. Int. J. Syst. Evolut. Microbiol. 2021, 71, 4. [Google Scholar] [CrossRef]
- Chen, R.-W.; Zhang, J.; He, Y.-Q.; Wang, K.-X.; Li, C.; Long, L.-J. Longirhabdus Pacifica Gen. Nov., Sp. Nov., Isolated from a Deep-Sea Hydrothermal Sediment in the West Pacific Ocean. Int. J. Syst. Evolut. Microbiol. 2019, 69, 3362–3367. [Google Scholar] [CrossRef]
- Liu, R.; Huang, Z.; Dong, C.; Shao, Z. Lottiidibacillus Patelloidae Gen. Nov., Sp. Nov., Isolated from the Intestinal Tract of a Marine Limpet and Reclassification of Bacillus Taeanensis as Maribacillus Taeanensis Gen. Nov., Comb. Nov. Antonie Van Leeuwenhoek 2019, 112, 797–807. [Google Scholar] [CrossRef]
- Ahmed, I.; Yokota, A.; Yamazoe, A.; Fujiwara, T. Proposal of Lysinibacillus Boronitolerans Gen. Nov. Sp. Nov., and Transfer of Bacillus Fusiformis to Lysinibacillus Fusiformis Comb. Nov. And Bacillus Sphaericus to Lysinibacillus Sphaericus Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2007, 57, 1117–1125. [Google Scholar] [CrossRef] [Green Version]
- Kloos, W.E.; Ballard, D.N.; George, C.G.; Webster, J.A.; Hubner, R.J.; Ludwig, W.; Schleifer, K.H.; Fiedler, F.; Schubert, K. Delimiting the Genus Staphylococcus through Description of Macrococcus Caseolyticus Gen. Nov., Comb. Nov. And Macrococcus Equipercicus Sp. Nov., Macrococcus Bovicus Sp. Nov. And Macrococcus Carouselicus Sp. Nov. Int. J. Syst. Evolut. Microbiol. 1998, 48, 859–877. [Google Scholar] [CrossRef] [Green Version]
- Madhaiyan, M.; Wirth, J.S.; Saravanan, V.S. Phylogenomic Analyses of the Staphylococcaceae Family Suggest the Reclassification of Five Species within the Genus Staphylococcus as Heterotypic Synonyms, the Promotion of Five Subspecies to Novel Species, the Taxonomic Reassignment of Five Staphylococcus Species to Mammaliicoccus Gen. Nov., and the Formal Assignment of Nosocomiicoccus to the Family Staphylococcaceae. Int. J. Syst. Evolut. Microbiol. 2020, 70, 5926–5936. [Google Scholar]
- Guo, L.-Y.; Xia, J.; Ling, S.; Chen, G.; Du, Z. Marinicrinis Sediminis Gen. Nov., Sp. Nov., Isolated from Marine Sediment. Int. J. Syst. Evolut. Microbiol. 2016, 66, 3725–3730. [Google Scholar] [CrossRef]
- Li, J.; Zhang, G.-T.; Yang, J.; Tian, X.-P.; Wang, F.-Z.; Zhang, C.S.; Zhang, S.; Li, W.-J. Marininema Mesophilum Gen. Nov., Sp. Nov., a Thermoactinomycete Isolated from Deep Sea Sediment, and Emended Description of the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 6, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Tian, X.; Zhang, S. Marinithermofilum Abyssi Gen. Nov., Sp. Nov. And Desmospora Profundinema Sp. Nov., Isolated from a Deep-Sea Sediment, and Emended Description of the Genus Desmosporayassin Et Al. 2009. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 8, 2622–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, M.V.; Kocur, M.; Komagata, K. Marinococcus Gen. Nov., a New Genus for Motile Cocci with Meso-Diaminopimelic Acid in the Cell Wall; and Marinococcus Albus Sp. Nov. And Marinococcus Halophilus (Novitsky and Kushner) Comb. Nov. J. Gener. Appl. Microbiol. 1984, 30, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Katsuta, A.; Matsuda, S.; Shizuri, Y.; Yokota, A.; Kasai, H. Mechercharimyces Mesophilus Gen. Nov., Sp. Nov. And Mechercharimyces Asporophorigenens Sp. Nov., Antitumour Substance-Producing Marine Bacteria, and Description of Thermoactinomycetaceae Fam. Nov. Int. J. Syst. Evolut. Microbiol. 2006, 56, 2837–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addou, N.A.; Schumann, P.; Spröer, C.; Ben Hania, W.; Hacene, H.; Fauque, G.; Cayol, J.-L.; Fardeau, M.-L. Melghiribacillus Thermohalophilus Gen. Nov., Sp. Nov., a Novel Filamentous, Endospore-Forming, Thermophilic and Halophilic Bacterium. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 4, 1172–1179. [Google Scholar] [CrossRef]
- Addou, A.N.; Schumann, P.; Spröer, C.; Hacene, H.; Cayol, J.-L.; Fardeau, M.-L. Melghirimyces Algeriensis Gen. Nov., Sp. Nov., a Member of the Family Thermoactinomycetaceae, Isolated from a Salt Lake. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 7, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Khelifi, N.; Romdhane, E.B.; Hedi, A.; Postec, A.; Fardeau, M.; Hamdi, M.; Tholozan, J.; Ollivier, B.; Hirschler-Réa, A. Characterization of Microaerobacter Geothermalis Gen. Nov., Sp. Nov., a Novel Microaerophilic, Nitrate- and Nitrite-Reducing Thermophilic Bacterium Isolated from a Terrestrial Hot Spring in Tunisia. Extremophiles 2010, 14, 297–304. [Google Scholar] [CrossRef]
- Echigo, A.; Minegishi, H.; Shimane, Y.; Kamekura, M.; Usami, R. Natribacillus Halophilus Gen. Nov., Sp. Nov., a Moderately Halophilic and Alkalitolerant Bacterium Isolated from Soil. Int. J. Syst. Evolut. Microbiol. 2012, 62, 289–294. [Google Scholar] [CrossRef]
- Sorokin, I.D.; Zadorina, E.V.; Kravchenko, I.K.; Boulygina, E.S.; Tourova, T.P.; Sorokin, D.Y. Natronobacillus Azotifigens Gen. Nov., Sp. Nov., an Anaerobic Diazotrophic Haloalkaliphile from Soda-Rich Habitats. Extremophiles 2008, 12, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.; Nogueira, C.; de Magalhaes-Sant’Ana, A.; Chung, A.P.; Morais, P.V.; da Costa, M.S. Nosocomiicoccus Ampullae Gen. Nov., Sp. Nov., Isolated from the Surface of Bottles of Saline Solution Used in Wound Cleansing. Int. J. Syst. Evolut. Microbiol. 2008, 58, 2939–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Chen, J.; Zhou, S. Novibacillus Thermophilus Gen. Nov., Sp. Nov., a Gram-Staining-Negative and Moderately Thermophilic Member of the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 8, 2591–2597. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Nogi, Y.; Takami, H. Oceanobacillus Iheyensis Gen. Nov., Sp. Nov., a Deep-Sea Extremely Halotolerant and Alkaliphilic Species Isolated from a Depth of 1050 M on the Iheya Ridge. FEMS Microbiol. Lett. 2001, 205, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Mayr, R.; Busse, H.-J.; Worliczek, H.L.; Ehling-Schulz, M.; Scherer, S. Ornithinibacillus Gen. Nov., with the Species Ornithinibacillus Bavariensis Sp. Nov. And Ornithinibacillus Californiensis Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2006, 56, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Córdoba, J.J.; Fernandez-Garayzabal, J.F.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A.E. The Phylogeny of the Genus Clostridium: Proposal of Five New Genera and Eleven New Species Combinations. Int. J. Syst. Evolut. Microbiol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Xiao, M.; Yang, L.-L.; Zhi, X.-Y.; Li, W.-J. Genome-Based Taxonomic Classification within the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2019, 69, 2028–2036. [Google Scholar] [CrossRef]
- Xu, L.; Huang, X.; Wang, H.; Tang, S.; Shen, B.; Sun, J. Description and Characterization of Three Endophytic Bacillaceae from the Halophyte Suaeda Salsa: Paenalkalicoccus Suaedae Gen. Nov., Sp. Nov., Cytobacillus Suaedae Sp. Nov., and Bacillus Suaedae Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2022, 72, 5. [Google Scholar]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular Identification of Rrna Group 3 Bacilli (Ash, Farrow, Wallbanks and Collins) Using a Pcr Probe Test. Antonie Van Leeuwenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef]
- Krishnamurthi, S.; Bhattacharya, A.; Mayilraj, S.; Saha, P.; Schumann, P.; Chakrabarti, T. Description of Paenisporosarcina Quisquiliarum Gen. Nov., Sp. Nov., and Reclassification of Sporosarcina Macmurdoensis Reddy Et Al. 2003 as Paenisporosarcina Macmurdoensis Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2009, 59, 1364–1370. [Google Scholar] [CrossRef]
- Frikha-Dammak, D.; Fardeau, M.-L.; Cayol, J.-L.; Ben Fguira-Fourati, L.; Najeh, S.; Ollivier, B.; Maalej, S. Paludifilum Halophilum Gen. Nov., Sp. Nov., a Thermoactinomycete Isolated from Superficial Sediment of a Solar Saltern. Int. J. Syst. Evolut. Microbiol. 2016, 66, 5371–5378. [Google Scholar] [CrossRef]
- Hwang, W.M.; Ko, Y.; Kang, K.; Ahn, T.-Y. Paludirhabdus Telluriireducens Gen. Nov., Sp. Nov. And Paludirhabdus Pumila Sp. Nov., Isolated from Soil of a Mountain Wetland and Emended Description of Gorillibacterium Massiliense. Int. J. Syst. Evolut. Microbiol. 2018, 68, 3040–3046. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; de Maayer, P. Corrigendum to “Phylogenomic Re-Assessment of the Thermophilic Genus Geobacillus” [Syst. Appl. Microbiol. 39 (2016) 527–533]. Syst. Appl. Microbiol. 2018, 41, 529–530. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Ishizaki, S.; Yamamoto, Y.; Yamasato, K. Paraliobacillus Ryukyuensis Gen. Nov., Sp. Nov., a New Gram-Positive, Slightly Halophilic, Extremely Halotolerant, Facultative Anaerobe Isolated from a Decomposing Marine Alga. J. Gener. Appl. Microbiol. 2002, 48, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Metchnikoff, M.E. Pasteuria Ramosa Un Representant Des Bacteries a Division Longitudinale. Ann. Inst. Pasteur 1888, 2, 165–170. [Google Scholar]
- Nunes, I.; Tiago, I.; Pires, A.L.; da Costa, M.S.; Veríssimo, A. Paucisalibacillus Globulus Gen. Nov., Sp. Nov., a Gram-Positive Bacterium Isolated from Potting Soil. Int. J. Syst. Evolut. Microbiol. 2006, 56, 1841–1845. [Google Scholar] [CrossRef]
- Sultanpuram, V.R.; Mothe, T.; Chintalapati, S.; Chintalapati, V.R. Pelagirhabdus Alkalitolerans Gen. Nov., Sp. Nov., an Alkali-Tolerant and Thermotolerant Bacterium Isolated from Beach Sediment, and Reclassification of Amphibacillus Fermentum as Pelagirhabdus Fermentum Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2016, 66, 84–90. [Google Scholar] [CrossRef]
- Tanasupawat, S.; Namwong, S.; Kudo, T.; Itoh, T. Piscibacillus Salipiscarius Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium from Fermented Fish (Pla-Ra) in Thailand. Int. J. Syst. Evolut. Microbiol. 2007, 57, 1413–1417. [Google Scholar] [CrossRef] [Green Version]
- Hatayama, K.; Shoun, H.; Ueda, Y.; Nakamura, A. Planifilum Fimeticola Gen. Nov., Sp. Nov. And Planifilum Fulgidum Sp. Nov., Novel Members of the Family ‘Thermoactinomycetaceae’ Isolated from Compost. Int. J. Syst. Evolut. Microbiol. 2005, 55, 2101–2104. [Google Scholar] [CrossRef]
- Migula, N. Arbeiten Aus Dem Bakteriologischen Institut Der Technischen Hochschule Zu. Karlsruhe 1894, 1, 235–238. [Google Scholar]
- Tsubouchi, T.; Shimane, Y.; Mori, K.; Usui, K.; Hiraki, T.; Tame, A.; Uematsu, K.; Maruyama, T.; Hatada, Y. Polycladomyces Abyssicola Gen. Nov., Sp. Nov., a Thermophilic Filamentous Bacterium Isolated from Hemipelagic Sediment. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 6, 1972–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Okamoto, T.; Matsuyama, H.; Yumoto, I. Polygonibacillus Indicireducens Gen. Nov., Sp. Nov., an Indigo-Reducing and Obligate Alkaliphile Isolated from Indigo Fermentation Liquor for Dyeing. Int. J. Syst. Evolut. Microbiol. 2016, 66, 4650–4656. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-M.; Jeon, C.O.; Song, S.M.; Kim, C.-J. Pontibacillus Chungwhensis Gen. Nov., Sp. Nov., a Moderately Halophilic Gram-Positive Bacterium from a Solar Saltern in Korea. Int. J. Syst. Evolut. Microbiol. 2005, 55, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, T.; Ranjan, V.K.; Ganguli, S.; Thakur, S.; Chakraborty, B.; Barman, P.; Ghosh, W.; Chakraborty, R. Pradoshia Eiseniae Gen. Nov., Sp. Nov., a Spore-Forming Member of the Family Bacillaceae Capable of Assimilating 3-Nitropropionic Acid, Isolated from the Anterior Gut of the Earthworm Eisenia Fetida. Int. J. Syst. Evolut. Microbiol. 2019, 69, 1265–1273. [Google Scholar] [CrossRef]
- Glaeser, S.P.; McInroy, J.A.; Busse, H.; Kämpfer, P. Pseudogracilibacillus Auburnensis Gen. Nov., Sp. Nov., Isolated from the Rhizosphere of Zea Mays. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 7, 2442–2448. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Glaeser, S.P.; Busse, H.-J.; McInroy, J.A.; Clermont, D.; Criscuolo, A. Pseudoneobacillus Rhizosphaerae Gen. Nov., Sp. Nov., Isolated from Maize Root Rhizosphere. Int. J. Syst. Evolut. Microbiol. 2022, 72, 5. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthi, S.; Ruckmani, A.; Pukall, R.; Chakrabarti, T. Psychrobacillus Gen. Nov. And Proposal for Reclassification of Bacillus Insolitus Larkin & Stokes, 1967, B. Psychrotolerans Abd-El Rahman Et Al., 2002 and B. Psychrodurans Abd-El Rahman Et Al., 2002 as Psychrobacillus Insolitus Comb. Nov., Psychrobacillus Psychrotolerans Comb. Nov. And Psychrobacillus Psychrodurans Comb. Nov. Syst. Appl. Microbiol. 2010, 33, 367–373. [Google Scholar] [PubMed]
- Wang, D.; Xiang, Y.; Jiang, C.; Zhang, J.; Hua, Z.; Niu, L.; Luo, L. Pueribacillus Theae Gen. Nov., Sp. Nov., Isolated from Pu’er Tea. Int. J. Syst. Evolut. Microbiol. 2018, 68, 2878–2882. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, B.; Liu, G.; Liu, Y.; Yang, H.; Yang, R.; Huang, Y.; Li, S.; Chen, T.; Zhang, W.; et al. Radiobacillus Deserti Gen. Nov., Sp. Nov., a Uv-Resistant Bacterium Isolated from Desert Soil. Int. J. Syst. Evolut. Microbiol. 2020, 70, 6338–6347. [Google Scholar] [CrossRef]
- Kim, M.; Kim, T.; Ri, S.; Jiang, F.; Chang, X.; Liu, Y.; Ren, L.; Qiu, X.; Deng, S.; Fang, C.; et al. Risungbinella Pyongyangensis Gen. Nov., Sp. Nov., a Mesophilic Member of the Family Thermoactinomycetaceae Isolated from an Agricultural Soil Sample. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 8, 2726–2733. [Google Scholar] [CrossRef]
- Vaishampayan, P.; Miyashita, M.; Ohnishi, A.; Satomi, M.; Rooney, A.; La Duc, M.T.; Venkateswaran, K. Description of Rummeliibacillus Stabekisii Gen. Nov., Sp. Nov. And Reclassification of Bacillus Pycnus Nakamura Et Al. 2002 as Rummeliibacillus Pycnus Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2009, 59, 1094–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, R.; García-Fraile, P.; Zurdo-Piñeiro, J.L.; Mateos, P.F.; Martínez-Molina, E.; Bedmar, E.J.; Sánchez-Raya, J.; Velázquez, E. Saccharibacillus Sacchari Gen. Nov., Sp. Nov., Isolated from Sugar Cane. Int. J. Syst. Evolut. Microbiol. 2008, 58, 1850–1854. [Google Scholar] [CrossRef] [PubMed]
- Nystrand, R. Saccharococcus Thermophilus Gen. Nov., Sp. Nov. Isolated from Beet Sugar Extraction. Syst. Appl. Microbiol. 1984, 5, 204–219. [Google Scholar] [CrossRef]
- Vishnuvardhan Reddy, S.; Thirumala, M.; Sasikala, C.; Ramana, C.V. Salibacterium Halotolerans Gen. Nov., Sp. Nov., a Bacterium Isolated from a Salt Pan, Reclassification of Bacillus Qingdaonensis as Salibacterium Qingdaonense Comb. Nov. And Bacillus Halochares as Salibacterium Halochares Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2015, 65 Pt 11, 4270–4275. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.-Y.; Oh, Y.J.; Lim, S.K.; Park, H.K.; Lee, C.; Kim, J.Y.; Lee, M.-A.; Choi, H.-J. Salicibibacter Kimchii Gen. Nov., Sp. Nov., a Moderately Halophilic and Alkalitolerant Bacterium in the Family Bacillaceae, Isolated from Kimchi. J. Microbiol. 2018, 56, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Kang, S.; Oh, T. Reclassification of Marinococcus Albus Hao Et Al. 1985 as Salimicrobium Album Gen. Nov., Comb. Nov. And Bacillus Halophilus Ventosa Et Al. 1990 as Salimicrobium Halophilum Comb. Nov., and Description of Salimicrobium Luteum Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2007, 57, 2406–2411. [Google Scholar] [CrossRef]
- Ren, P.-G.; Zhou, P.-J. Salinibacillus Aidingensis Gen. Nov., Sp. Nov. And Salinibacillus Kushneri Sp. Nov., Moderately Halophilic Bacteria Isolated from a Neutral Saline Lake in Xin-Jiang, China. Int. J. Syst. Evolut. Microbiol. 2005, 55, 949–953. [Google Scholar] [CrossRef] [Green Version]
- Ventosa, A.; Márquez, M.C.; Ruiz-Berraquero, F.; Kocur, M. Salinicoccus Roseus Gen. Nov., Sp. Nov., a New Moderately Halophilic Gram-Positive Coccus. Syst. Appl. Microbiol. 1990, 13, 29–33. [Google Scholar] [CrossRef]
- Zarparvar, P.; Amoozegar, M.A.; Nikou, M.M.; Schumann, P.; Ventosa, A. Salinithrix Halophila Gen. Nov., Sp. Nov., a Halophilic Bacterium in the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 12, 4115–4119. [Google Scholar] [CrossRef] [Green Version]
- Sultanpuram, V.R.; Mothe, T. Salipaludibacillus Aurantiacus Gen. Nov., Sp. Nov. A Novel Alkali Tolerant Bacterium, Reclassification of Bacillus Agaradhaerens as Salipaludibacillus Agaradhaerens Comb. Nov. And Bacillus Neizhouensis as Salipaludibacillus Neizhouensis Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2016, 66, 2747–2753. [Google Scholar] [CrossRef]
- Albuquerque, L.; Tiago, I.; Rainey, F.A.; Taborda, M.; Nobre, M.F.; Verissimo, A.; Da Costa, M.S. Salirhabdus Euzebyi Gen. Nov., Sp. Nov., a Gram-Positive, Halotolerant Bacterium Isolated from a Sea Salt Evaporation Pond. Int. J. Syst. Evolut. Microbiol. 2007, 57, 1566–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Cao, S.-J.; Li, Z.-H.; Fan, H.; Li, H.-F.; Liu, W.-J.; Yuan, H.-L. Salisediminibacterium Halotolerans Gen. Nov., Sp. Nov., a Halophilic Bacterium from Soda Lake Sediment. Int. J. Syst. Evolut. Microbiol. 2012, 62 Pt 9, 2127–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amoozegar, M.A.; Bagheri, M.; Didari, M.; Fazeli, S.A.S.; Schumann, P.; Sánchez-Porro, C.; Ventosa, A. Saliterribacillus Persicus Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium Isolated from a Hypersaline Lake. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 1, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, I.J.; Márquez, M.C.; Xue, Y.; Ma, Y.; Cowan, D.A.; Jones, B.E.; Grant, W.D.; Ventosa, A. Salsuginibacillus Kocurii Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium from Soda-Lake Sediment. Int. J. Syst. Evolut. Microbiol. 2007, 57, 2381–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, T.R.; Johnson, C.N.; Patel, N.; Cotta, M.A.; Moore, E.; Lawson, P.A. Savagea Faecisuis Gen. Nov., Sp. Nov., a Tylosin- and Tetracycline-Resistant Bacterium Isolated from a Swine-Manure Storage Pit. Antonie Van Leeuwenhoek 2015, 108, 151–161. [Google Scholar] [CrossRef]
- Lee, S.D.; Lee, D.W. Scopulibacillus Darangshiensis Gen. Nov., Sp. Nov., Isolated from Rock. J. Microbiol. 2009, 47, 710–715. [Google Scholar] [CrossRef]
- Carrasco, I.J.; Márquez, M.C.; Xue, Y.; Ma, Y.; Cowan, D.A.; Jones, B.E.; Grant, W.D.; Ventosa, A. Sediminibacillus Halophilus Gen. Nov., Sp. Nov., a Moderately Halophilic, Gram-Positive Bacterium from a Hypersaline Lake. Int. J. Syst. Evolut. Microbiol. 2008, 58, 1961–1967. [Google Scholar] [CrossRef] [Green Version]
- Park, D.-J.; Dastager, S.; Lee, J.-C.; Yeo, S.-H.; Yoon, J.-H.; Kim, C.-J. Shimazuella Kribbensis Gen. Nov., Sp. Nov., a Mesophilic Representative of the Family Thermoactinomycetaceae. Int. J. Syst. Evolut. Microbiol. 2007, 57, 2660–2664. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Zhou, S. Sinibacillus Soli Gen. Nov., Sp. Nov., a Moderately Thermotolerant Member of the Family Bacillaceae. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 5, 1647–1653. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-J.; Zhi, X.; Euzéby, J.P. Proposal of Yaniellaceae Fam. Nov., Yaniella Gen. Nov. And Sinobaca Gen. Nov. As Replacements for the Illegitimate Prokaryotic Names Yaniaceae Li Et Al. 2005, Yania Li Et Al. 2004, Emend Li Et Al. 2005, and Sinococcus Li Et Al. 2006, Respectively. Int. J. Syst. Evolut. Microbiol. 2008, 58, 525–527. [Google Scholar] [CrossRef]
- Krishnamurthi, S.; Chakrabarti, T.; Stackebrandt, E. Re-Examination of the Taxonomic Position of Bacillus Silvestris Rheims Et Al. 1999 and Proposal to Transfer It to Solibacillus Gen. Nov. As Solibacillus Silvestris Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2009, 59, 1054–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitahara, K.; Suzuki, J. Sporolactobacillus Nov. Subgen. J. Gener. Appl. Microbiol. 1963, 9, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Kluyver, A.J.; van Niel, C.B. Prospects for a Natural System of Classification of Bacteria. Zent. Fur Bakteriol. Parasitenkd. Infekt. Und Hyg. 1936, 94, 369–403. [Google Scholar]
- Rosenbach, A.J.F. Mikro-Organismen Bei Den Wund-Infections-Krankheiten Des Menschen; JF Bergmann: Paris, France, 1884. [Google Scholar]
- Wang, K.-X.; He, Y.-Q.; Chen, R.-W.; Li, C.; Tian, X.-P.; Long, L.-J. Staphylospora Marina Gen. Nov., Sp. Nov., a Novel Member of the Family Thermoactinomycetaceae, Isolated from a Deep-Sea Hydrothermal Vent in the Pacific Ocean. Int. J. Syst. Evolut. Microbiol. 2019, 69, 1452–1458. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xue, Y.; Ma, Y. Streptohalobacillus Salinus Gen. Nov., Sp. Nov., a Moderately Halophilic, Gram-Positive, Facultative Anaerobe Isolated from Subsurface Saline Soil. Int. J. Syst. Evolut. Microbiol. 2011, 61, 1127–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golovacheva, R.S.; Karavaĭko, G.I. Sulfobacillus, a New Genus of Thermophilic Sporulating Bacteria. Mikrobiologiia 1978, 47, 815–822. [Google Scholar] [PubMed]
- Li, J.; Liu, J.; Shen, Y.; Ni, J. Swionibacillus Sediminis Gen. Nov., Sp. Nov., a Member of the Family Bacillaceae Isolated from Ocean Sediment. Int. J. Syst. Evolut. Microbiol. 2017, 67, 3440–3445. [Google Scholar] [CrossRef]
- Ren, P.-G.; Zhou, P.-J. Tenuibacillus Multivorans Gen. Nov., Sp. Nov., a Moderately Halophilic Bacterium Isolated from Saline Soil in Xin-Jiang, China. Int. J. Syst. Evolut. Microbiol. 2005, 55, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Slobodkina, G.B.; Panteleeva, A.N.; Kostrikina, N.A.; Kopitsyn, D.S.; Bonch-Osmolovskaya, E.A.; Slobodkin, A.I. Tepidibacillus Fermentans Gen. Nov., Sp. Nov.: A Moderately Thermophilic Anaerobic and Microaerophilic Bacterium from an Underground Gas Storage. Extremophiles 2013, 17, 833–839. [Google Scholar] [CrossRef]
- An, S.-Y.; Asahara, M.; Goto, K.; Kasai, H.; Yokota, A.; Nam, Y.-D.; Chang, H.-W.; Park, J.R.; Kwon, H.-Y.; Quan, Z.-X.; et al. Terribacillus Saccharophilus Gen. Nov., Sp. Nov. And Terribacillus Halophilus Sp. Nov., Spore-Forming Bacteria Isolated from Field Soil in Japan. Int. J. Syst. Evolut. Microbiol. 2007, 57, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Prasirtsak, B.; Thongchul, N.; Tolieng, V.; Tanasupawat, S. Terrilactibacillus Laevilacticus Gen. Nov., Sp. Nov., Isolated from Soil. Int. J. Syst. Evolut. Microbiol. 2016, 66, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Romero, E.; Coutiño-Coutiño, M.d.; Valenzuela-Encinas, C.; López-Ramírez, M.P.; Marsch, R.; Dendooven, L. Texcoconibacillus Texcoconensis Gen. Nov., Sp. Nov., Alkalophilic and Halotolerant Bacteria Isolated from Soil of the Former Lake Texcoco (Mexico). Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 9, 3336–3341. [Google Scholar] [CrossRef] [PubMed]
- García, M.T.; Gallego, V.; Ventosa, A.; Mellado, E. Thalassobacillus Devorans Gen. Nov., Sp. Nov., a Moderately Halophilic, Phenol-Degrading, Gram-Positive Bacterium. Int. J. Syst. Evolut. Microbiol. 2005, 55, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Sultanpuram, V.R.; Mothe, T. Thalassorhabdus Alkalitolerans Gen. Nov., Sp. Nov., a Novel Bacillaceae Member Isolated from Marine Sediment. Int. J. Syst. Evolut. Microbiol. 2018, 68, 2969–2976. [Google Scholar] [CrossRef] [PubMed]
- Gössner, A.S.; Devereux, R.; Ohnemüller, N.; Acker, G.; Stackebrandt, E.; Drake, H.L. Thermicanus Aegyptius Gen. Nov., Sp. Nov., Isolated from Oxic Soil, a Fermentative Microaerophile That Grows Commensally with the Thermophilic Acetogen Moorella Thermoacetica. Appl. Environ. Microbiol. 1999, 65, 5124–5133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiklinsky, P. Sur Les Mucédinées Thermophiles. Ann. Inst. Pasteur 1899, 13, 500–505. [Google Scholar]
- Touzel, J.P.; O’Donohue, M.; Debeire, P.; Samain, E.; Breton, C. Thermobacillus Xylanilyticus Gen. Nov., Sp. Nov., a New Aerobic Thermophilic Xylan-Degrading Bacterium Isolated from Farm Soil. Int. J. Syst. Evolut. Microbiol. 2000, 50, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Cihan, A.C.; Koc, M.; Ozcan, B.; Tekin, N.; Cokmus, C. Thermolongibacillus Altinsuensis Gen. Nov., Sp. Nov. And Thermolongibacillus Kozakliensis Sp. Nov., Aerobic, Thermophilic, Long Bacilli Isolated from Hot Springs. Int. J. Syst. Evolut. Microbiol. 2014, 64 Pt 1, 187–197. [Google Scholar] [CrossRef]
- Steven, B.; Chen, M.Q.; Greer, C.W.; Whyte, L.G.; Niederberger, T.D. Tumebacillus Permanentifrigoris Gen. Nov., Sp. Nov., an Aerobic, Spore-Forming Bacterium Isolated from Canadian High Arctic Permafrost. Int. J. Syst. Evolut. Microbiol. 2008, 58, 1497–1501. [Google Scholar] [CrossRef] [Green Version]
- Fortina, M.G.; Pukall, R.; Schumann, P.; Mora, D.; Parini, C.; Manachini, P.L.; Stackebrandt, E. Ureibacillus Gen. Nov., a New Genus to Accommodate Bacillus Thermosphaericus (Andersson Et Al. 1995), Emendation of Ureibacillus Thermosphaericus and Description of Ureibacillus Terrenus Sp. Nov. Int. J. Syst. Evolut. Microbiol. 2001, 51, 447–455. [Google Scholar] [CrossRef]
- Heyndrickx, M.; Lebbe, L.; Kersters, K.; De Vos, P.; Forsyth, G.; Logan, N.A. Virgibacillus: A New Genus to Accommodate Bacillus Pantothenticus (Proom and Knight 1950). Emended Description of Virgibacillus Pantothenticus. Int. J. Syst. Evolut. Microbiol. 1998, 48, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Albert, R.A.; Archambault, J.; Lempa, M.; Hurst, B.; Richardson, C.; Gruenloh, S.; Duran, M.; Worliczek, H.L.; Huber, B.E.; Rossello-Mora, R.; et al. Proposal of Viridibacillus Gen. Nov. And Reclassification of Bacillus Arvi, Bacillus Arenosi and Bacillus Neidei as Viridibacillus Arvi Gen. Nov., Comb. Nov., Viridibacillus Arenosi Comb. Nov. And Viridibacillus Neidei Comb. Nov. Int. J. Syst. Evolut. Microbiol. 2007, 57, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Haridon, S.L.; Miroshnichenko, M.L.; Kostrikina, N.A.; Tindall, B.J.; Spring, S.; Schumann, P.; Stackebrandt, E.; Bonch-Osmolovskaya, E.A.; Jeanthon, C. Vulcanibacillus Modesticaldus Gen. Nov., Sp. Nov., a Strictly Anaerobic, Nitrate-Reducing Bacterium from Deep-Sea Hydrothermal Vents. Int. J. Syst. Evolut. Microbiol. 2006, 56, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Kukolya, J.; Bata-Vidács, I.; Luzics, S.; Tóth, E.; Kéki, Z.; Schumann, P.; Táncsics, A.; Nagy, I.; Olasz, F.; Tóth, Á. Xylanibacillus Composti Gen. Nov., Sp. Nov., Isolated from Compost. Int. J. Syst. Evolut. Microbiol. 2018, 68, 698–702. [Google Scholar] [CrossRef] [PubMed]

| Genus Name | Strain Name | Type Strain | Important Properties | Potential Applications | (Poly)Extremophilic Features | Reference | |
|---|---|---|---|---|---|---|---|
| Aliibacillus | A. thermotolerans strain BM62 | Yes | Ammonia oxidation | Reduction of the air pollution caused by ammonia volatilization; Increase nitrate content in the compost | Moderately thermophile | [294,295] | |
| Alkalilactibacillus | A. ikkensis strain GCM68 | Yes | Production of a cold-active β-galactosidase | Heat-labile enzymatic digestions in genetic engineering; An affordable choice for laundry detergents; Food processing at low temperature (lactose hydrolyzing of milk) | Alkaliphile Halotolerant Psychrophile | [296,297] | |
| Allobacillus | A. halotolerans strain MSP69 | No | Production of an extracellular alkaline nuclease | Biocatalyst and flavor enhancer of fish sauce | Halotolerant | [298] | |
| Caldalkalibacillus | C. thermarum strain TA2.A1 | No | Production of a thermostable laccase causing dimerization of a dimeric lignin model compound GGGE | Biocatalyst for delignification and detoxification of lignocellulosic biomass | Thermoalkaliphile | [299,300] | |
| Filobacillus | Filobacillus sp. RF2–5 | No | Production of a halo- and thermostable serine proteinase | Useful for the degradation of fish protein during fermentation at high salt concentrations and might be useful for reduction of the fermentation period | Moderately halophile | [301] | |
| Oceanobacillus | Oceanobacillus oncorhynchi subsp. incaldaniensis strain 20AG | Yes | Production of a novel thermo-alkali stable catalase–peroxidase | Useful for phenol resin synthesis; Bioremediation of anilines and phenols; Lignin degradation in paper industry | Halophile Alkalitolerant | [302,303] | |
| Oceanobacillus sp. PUMB02 | No | Production of a halotolerant thermostable lipase | Potential utility in inhibiting biofilm formation in a food processing environment | [304] | |||
| Paralkalibacillus | Paralkalibacillus indicireducens strain Bps-1 | Yes | Indigo reduction | Potential use in indigo dying industry | Obligately alkaliphile Halotolerant | [305] | |
| Virgibacillus | Virgibacillus sp. SK37 | No | NaCl-activated Proteinase | Potential starter to improve fish sauce quality | Moderately halophile | [306,307,308] | |
| Laceyella | Laceyella sacchari strain LP175 | No | Thermostable amylase | Raw starch industry; Bioethanol production | Thermophile | [309] | |
| Laceyella putida strain JAM FM3001 | No | Highly thermostable and surfactant-activated chitinase | Potential applications in waste management; Biocontrol agent; Pharmaceutical industry | [310] | |||
| Halolactibacillus | Halolactibacillus alkaliphilus MSRD1 | No | Antibacterial activity of red pigment | Pharmaceutical industry | Halotolerant | [311] | |
| Halolactibacillus miurensis strain SEEN MKU3 | No | Production of exopolysaccharides with antioxidant activities | Potential applications in functional foods; Cosmetic industry; Pharmaceutical industry | [312] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harirchi, S.; Sar, T.; Ramezani, M.; Aliyu, H.; Etemadifar, Z.; Nojoumi, S.A.; Yazdian, F.; Awasthi, M.K.; Taherzadeh, M.J. Bacillales: From Taxonomy to Biotechnological and Industrial Perspectives. Microorganisms 2022, 10, 2355. https://doi.org/10.3390/microorganisms10122355
Harirchi S, Sar T, Ramezani M, Aliyu H, Etemadifar Z, Nojoumi SA, Yazdian F, Awasthi MK, Taherzadeh MJ. Bacillales: From Taxonomy to Biotechnological and Industrial Perspectives. Microorganisms. 2022; 10(12):2355. https://doi.org/10.3390/microorganisms10122355
Chicago/Turabian StyleHarirchi, Sharareh, Taner Sar, Mohaddaseh Ramezani, Habibu Aliyu, Zahra Etemadifar, Seyed Ali Nojoumi, Fatemeh Yazdian, Mukesh Kumar Awasthi, and Mohammad J. Taherzadeh. 2022. "Bacillales: From Taxonomy to Biotechnological and Industrial Perspectives" Microorganisms 10, no. 12: 2355. https://doi.org/10.3390/microorganisms10122355