
Probiotic Properties of Chicken-Derived Highly Adherent Lactic Acid Bacteria and Inhibition of Enteropathogenic Bacteria in Caco-2 Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Growth Conditions
2.2. Cell Surface Properties Assay
2.2.1. Hydrophobicity
2.2.2. Auto-Aggregation
2.2.3. Co-Aggregation with Pathogenic Bacteria
2.3. Adhesion Assay to Caco-2 Cells
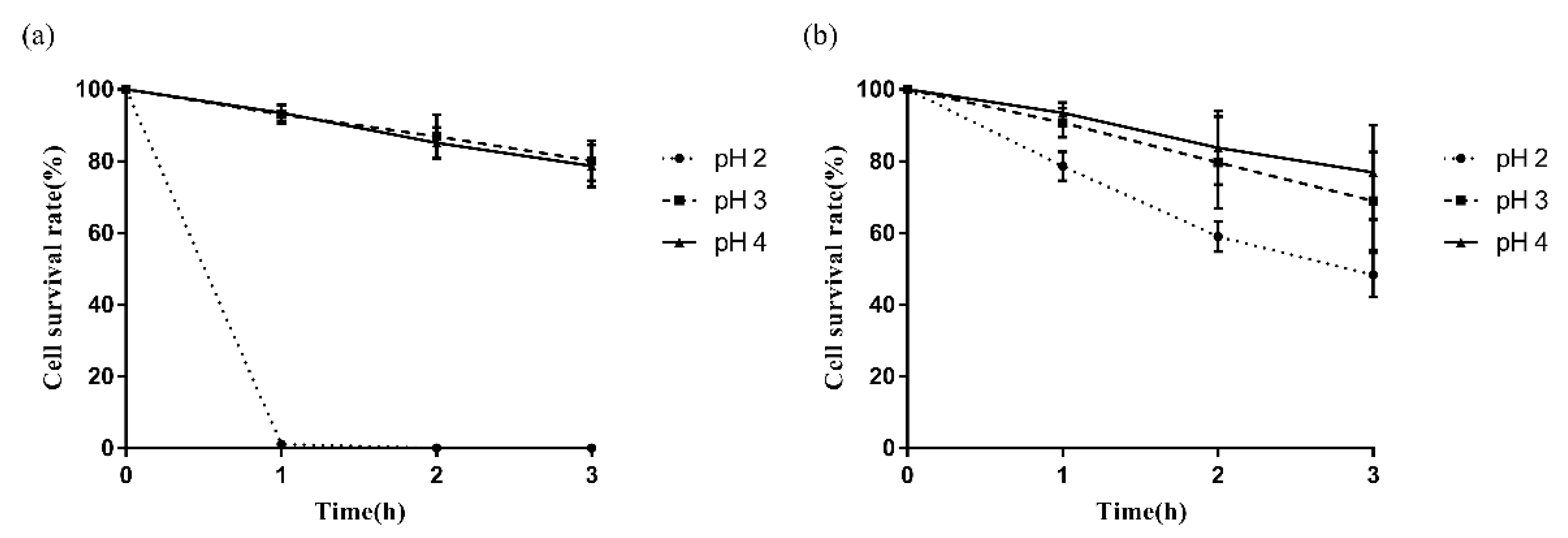
2.4. Acid Tolerance
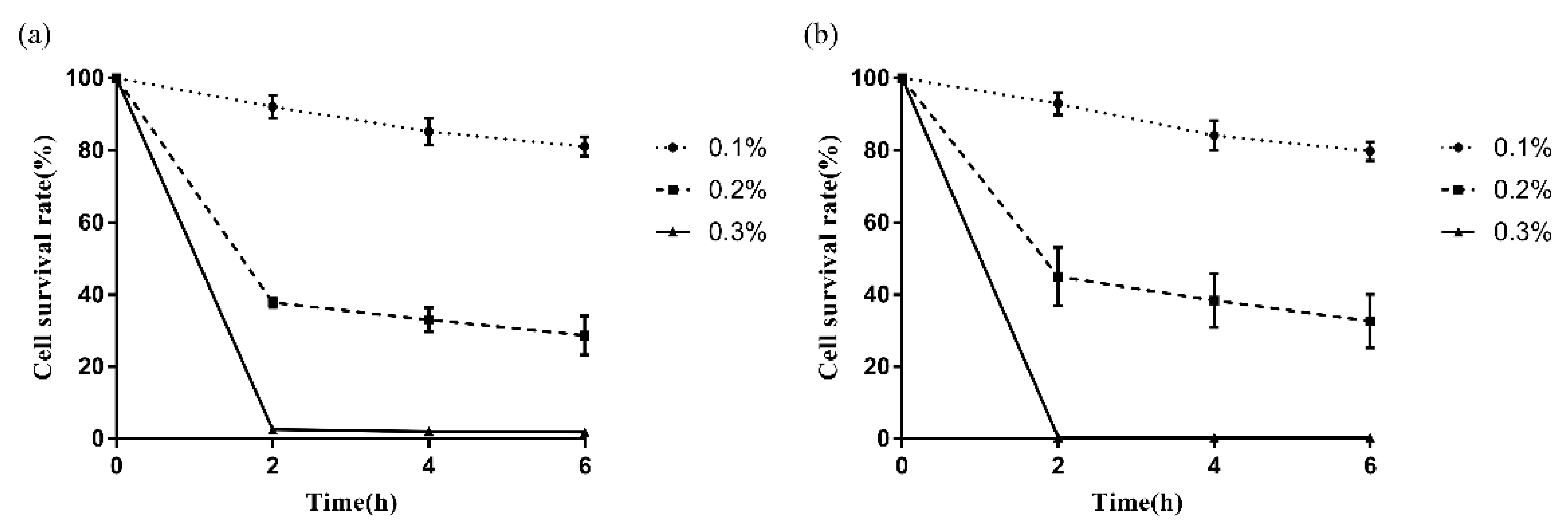
2.5. Bile Salt Tolerance
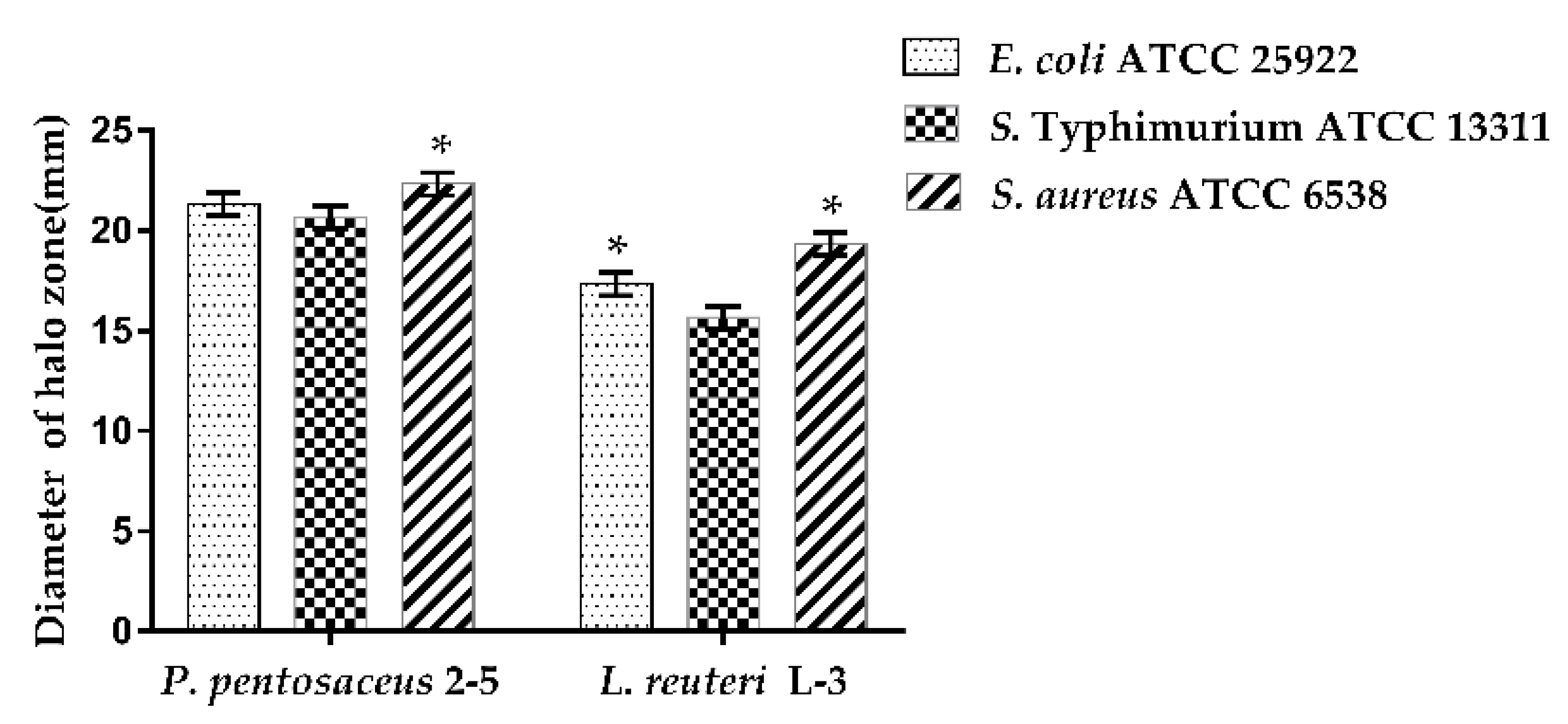
2.6. Antimicrobial Activity
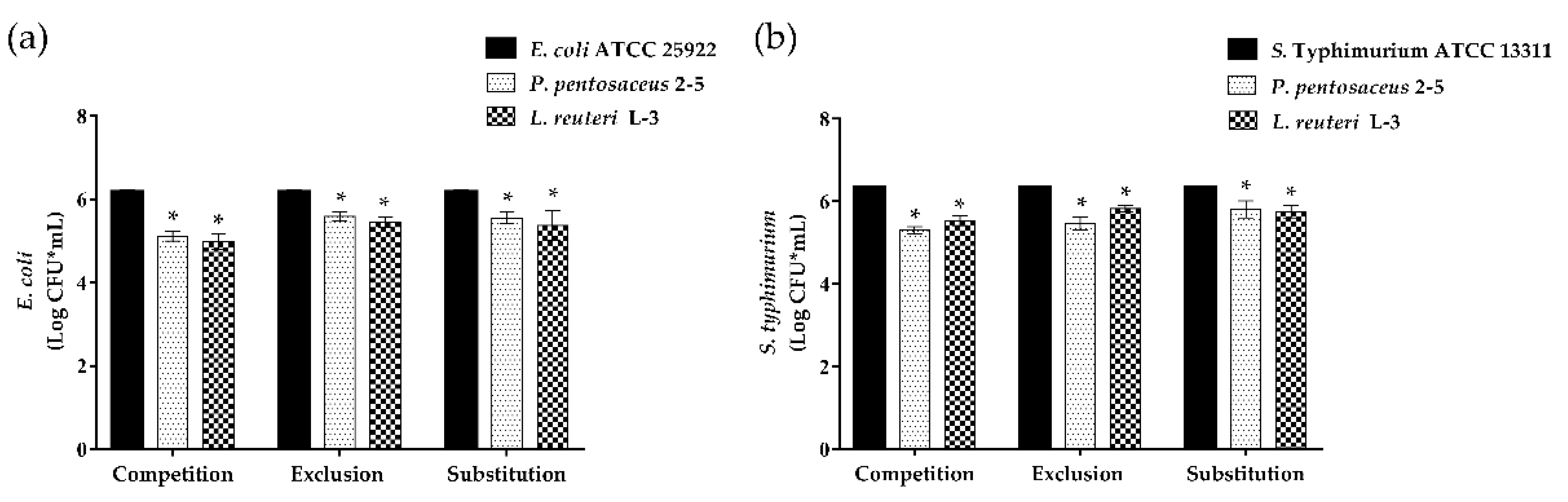
2.7. Anti-Adhesion Assay against Enteropathogenic Bacteria
2.8. Statistical Analyses
3. Results
3.1. Cell Surface Properties of LAB
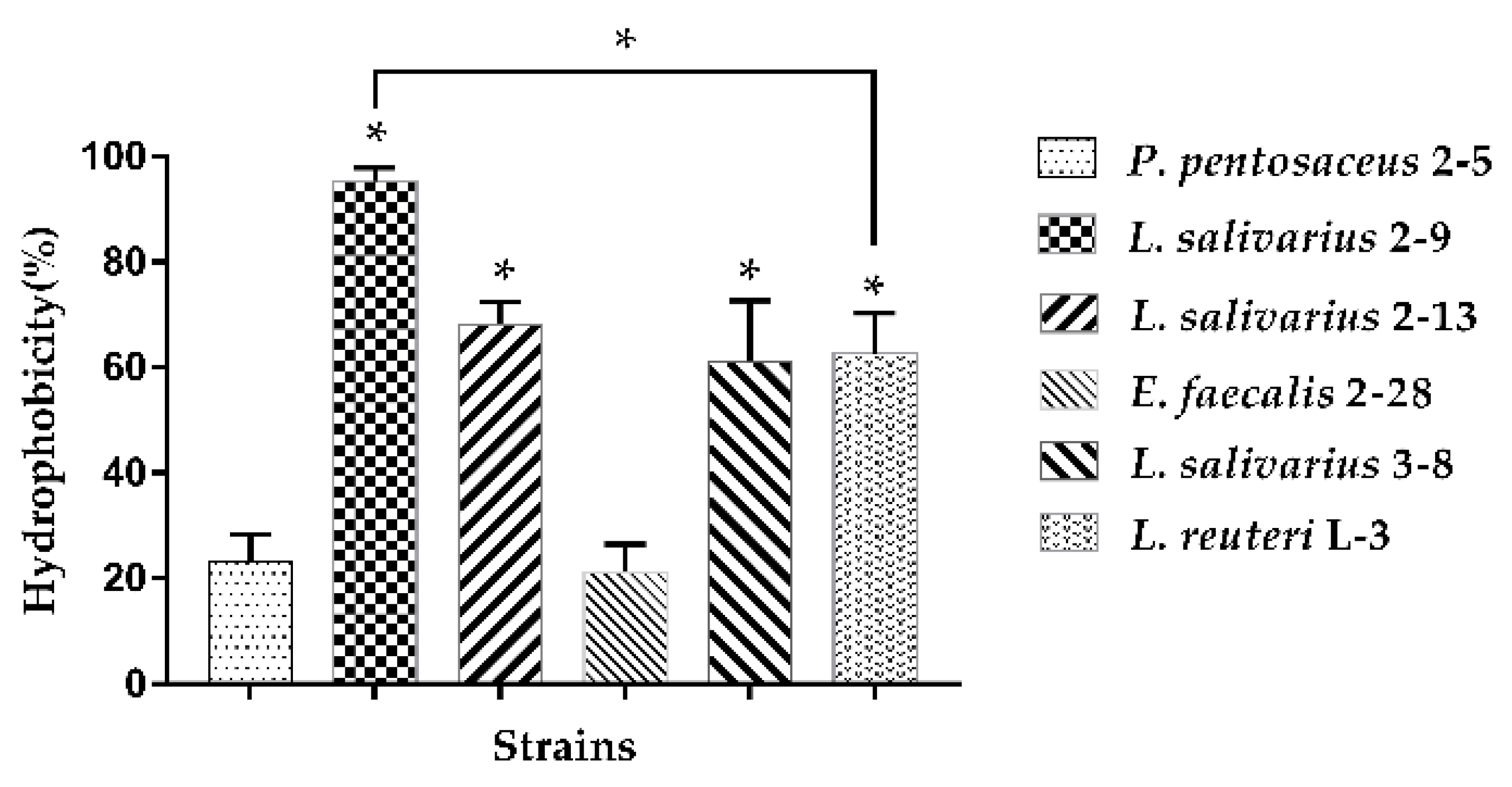
3.1.1. Hydrophobicity
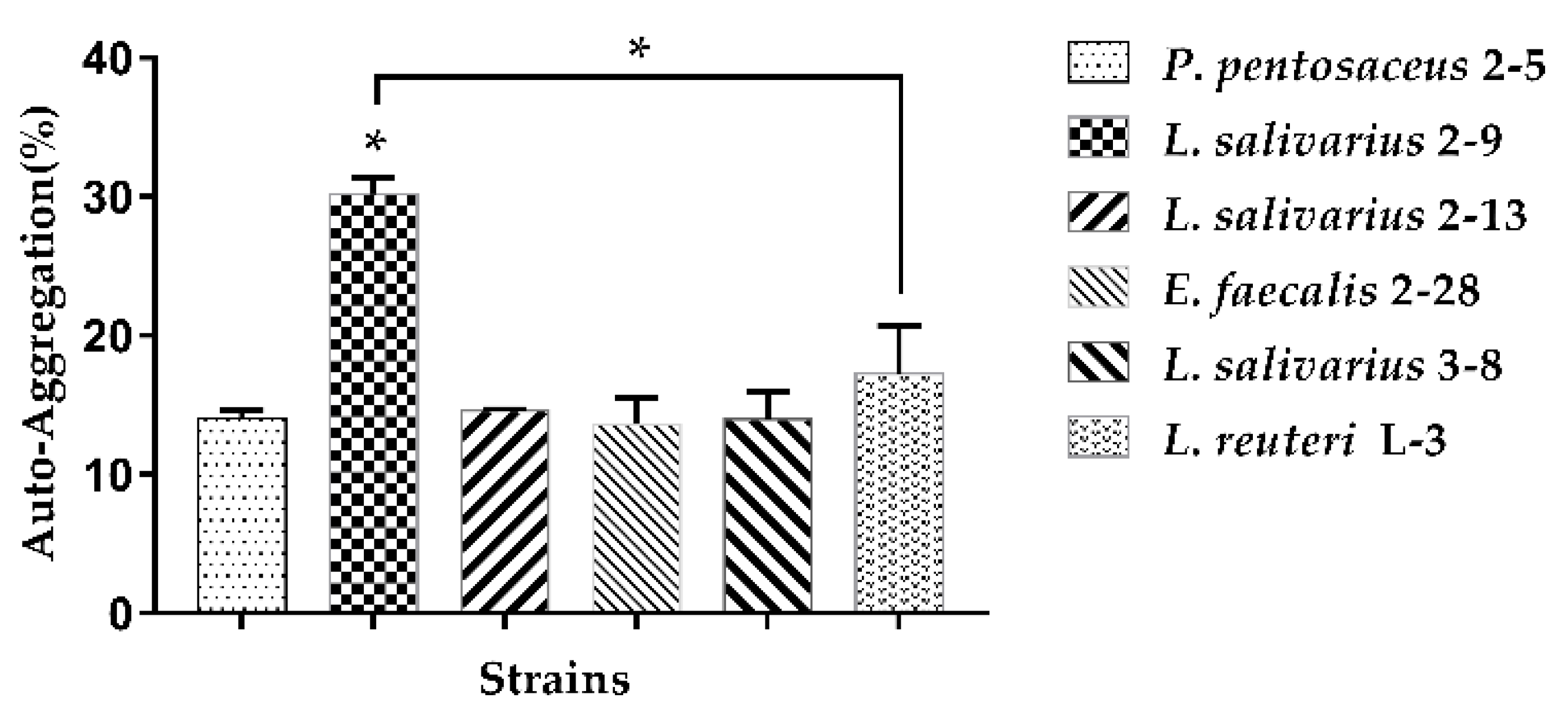
3.1.2. Auto-Aggregation
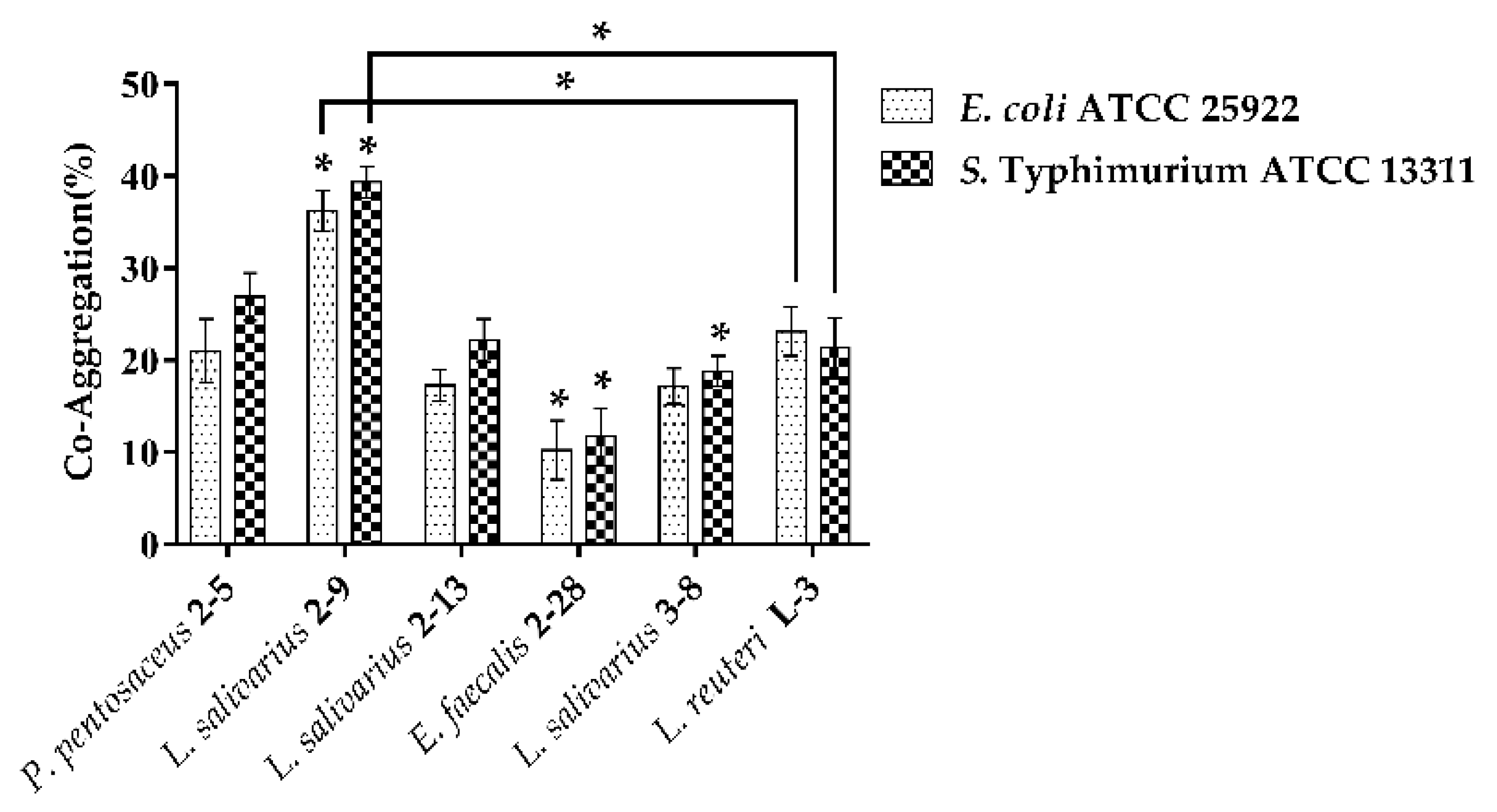
3.1.3. Co-Aggregation
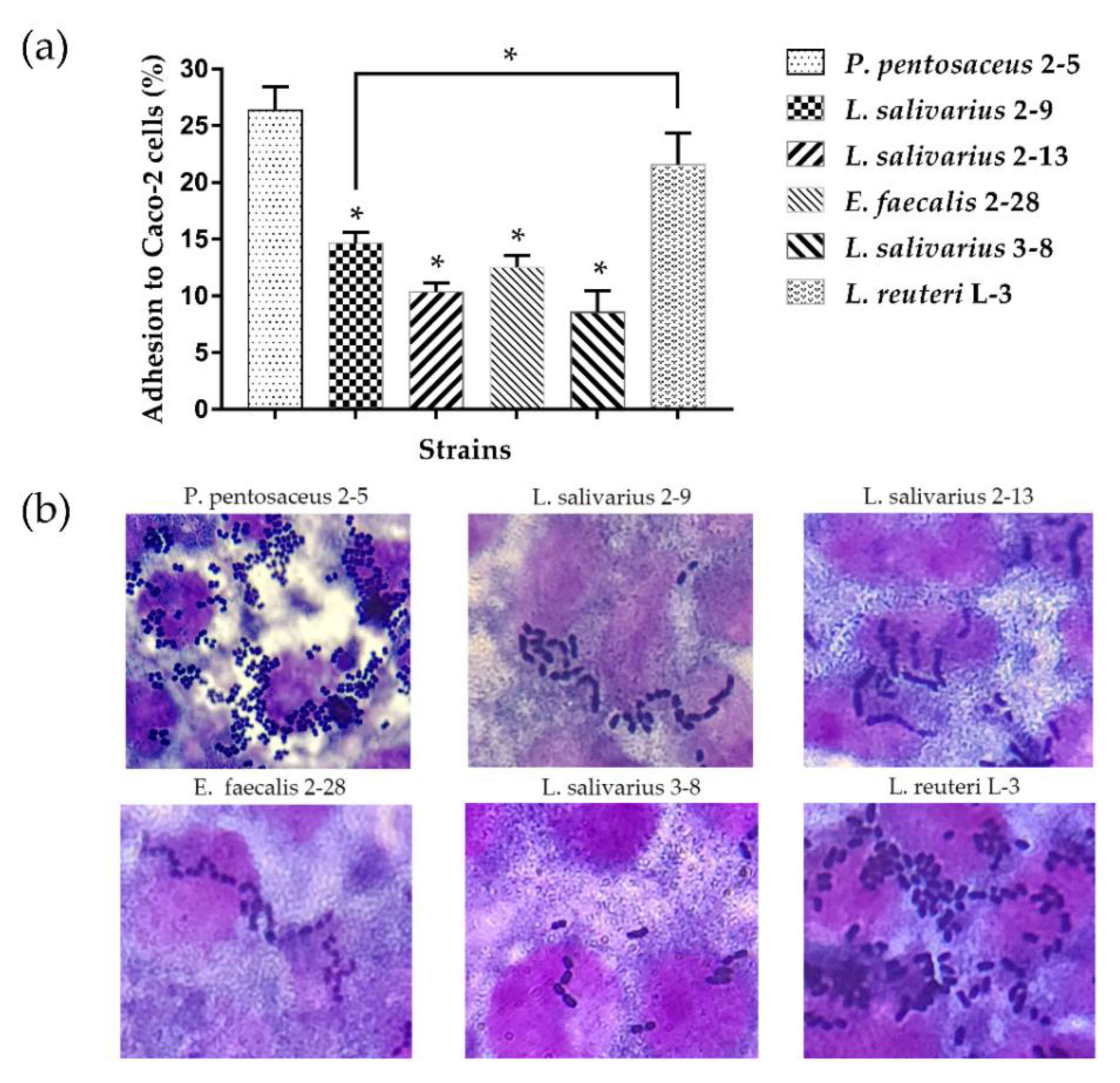
3.2. Adhesion of LAB to Caco-2 Cell Line
3.3. Survival of LAB in an Acidic Environment
3.4. Survival of LAB in Bile Salt Environment
3.5. Antimicrobial Activity of CFS of LAB
3.6. Anti-Adhesion of LAB to Enteropathogenic Bacteria on Caco-2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Diaz, J.; Munoz-Quezada, S.; Gomez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Aximujiang, K.; Kaheman, K.; Wushouer, X.; Wu, G.; Ahemaiti, A.; Yunusi, K. Lactobacillus acidophilus and HKL Suspension Alleviates Ulcerative Colitis in Rats by Regulating Gut Microbiota, Suppressing TLR9, and Promoting Metabolism. Front. Pharmacol. 2022, 13, 859628. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.; Nigam, P.S. The Gut Microbiota Influenced by the Intake of Probiotics and Functional Foods with Prebiotics Can Sustain Wellness and Alleviate Certain Ailments like Gut-Inflammation and Colon-Cancer. Microorganisms 2022, 10, 665. [Google Scholar] [CrossRef]
- Benmouna, Z.; Dalache, F.; Zadi-Karam, H.; Karam, N.E.; Vuotto, C. Ability of Three Lactic Acid Bacteria to Grow in Sessile Mode and to Inhibit Biofilm Formation of Pathogenic Bacteria. Adv. Exp. Med. Biol. 2020, 1282, 105–114. [Google Scholar]
- Peng, X.Q.; Ed-Dra, A.; Song, Y.; Elbediwi, M.; Nambiar, R.B.; Zhou, X.; Yue, M. Lacticaseibacillus rhamnosus alleviates intestinal inflammation and promotes microbiota-mediated protection against Salmonella fatal infections. Front. Immunol. 2022, 13, 973224. [Google Scholar] [CrossRef]
- Wang, R.; Jiang, L.; Zhang, M.; Zhao, L.; Hao, Y.L.; Guo, H.Y.; Sang, Y.; Zhang, H.; Ren, F.Z. The Adhesion of Lactobacillus salivarius REN to a Human Intestinal Epithelial Cell Line Requires S-layer Proteins. Sci Rep 2017, 7, 44029. [Google Scholar] [CrossRef] [Green Version]
- Kathayat, D.; Closs, G.; Helmy, Y.A.; Deblais, L.; Srivastava, V.; Rajashekara, G. In Vitro and In Vivo Evaluation of Lacticaseibacillus rhamnosus GG and Bifidobacterium lactis Bb12 Against Avian Pathogenic Escherichia coli and Identification of Novel Probiotic-Derived Bioactive Peptides. Probiotics Antimicrob. Proteins 2022, 14, 1012–1028. [Google Scholar] [CrossRef]
- Hai, D.; Lu, Z.X.; Huang, X.Q.; Lv, F.X.; Bie, X.M. In Vitro Screening of Chicken-Derived Lactobacillus Strains that Effectively Inhibit Salmonella Colonization and Adhesion. Foods 2021, 10, 569. [Google Scholar] [CrossRef]
- Fonseca, H.C.; Melo, D.D.; Ramos, C.L.; Dias, D.R.; Schwan, R.F. Probiotic Properties of Lactobacilli and Their Ability to Inhibit the Adhesion of Enteropathogenic Bacteria to Caco-2 and HT-29 Cells. Probiotics Antimicrob. Proteins 2021, 13, 102–112. [Google Scholar] [CrossRef]
- Mohsin, M.; Li, Y.G.; Zhang, X.; Wang, Y.L.; Huang, Z.J.; Yin, G.W.; Zhang, Z.P. Development of CRISPR-CAS9 based RNA drugs against Eimeria tenella infection. Genomics 2021, 113, 4126–4135. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Moon, A.; Huang, J.S.; Sun, Y.; Qiu, H.J. Antiviral Effects and Underlying Mechanisms of Probiotics as Promising Antivirals. Front. Cell. Infect. Microbiol. 2022, 12, 928050. [Google Scholar] [CrossRef] [PubMed]
- Schmalstig, A.A.; Freidy, S.; Hanafin, P.O.; Braunstein, M.; Rao, G.G. Reapproaching Old Treatments: Considerations for PK/PD Studies on Phage Therapy for Bacterial Respiratory Infections. Clin. Pharmacol. Ther. 2021, 109, 1443–1456. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Abbas, R.Z.; Yin, G.W.; Sindhu, Z.U.D.; Abbas, A.; Huang, Z.J.; Aleem, M.T.; Saeed, Z.; Afzal, M.Z.; Ejaz, A.; et al. Probiotics as therapeutic, antioxidant and immunomodulatory agents against poultry coccidiosis. Worlds Poult. Sci. J. 2021, 77, 331–345. [Google Scholar] [CrossRef]
- Andrade, J.C.; Kumar, S.; Kumar, A.; Cernakova, L.; Rodrigues, C.F. Application of probiotics in candidiasis management. Crit. Rev. Food Sci. Nutr. 2022, 62, 8249–8264. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [Green Version]
- Vila, B.; Esteve-Garcia, E.; Brufau, J. Probiotic micro-organisms: 100 years of innovation and efficacy; modes of action. Worlds Poult. Sci. J. 2010, 66, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Valdovska, A.; Jemeljanovs, A.; Pilmane, M.; Zitare, I.; Konosonoka, I.H.; Lazdins, M. Alternative for improving gut microbiota: Use of Jerusalem artichoke and probiotics in diet of weaned piglets. Pol. J. Vet. Sci. 2014, 17, 61–69. [Google Scholar] [CrossRef]
- Bhogoju, S.; Khwatenge, C.N.; Taylor-Bowden, T.; Akerele, G.; Kimathi, B.M.; Donkor, J.; Nahashon, S.N. Effects of Lactobacillus reuteri and Streptomyces coelicolor on Growth Performance of Broiler Chickens. Microorganisms 2021, 9, 1341. [Google Scholar] [CrossRef]
- Jiang, S.M.; Cai, L.Z.; Lv, L.X.; Li, L.J. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell. Fact. 2021, 20, 45. [Google Scholar] [CrossRef]
- Benbara, T.; Lalouche, S.; Drider, D.; Bendali, F. Lactobacillus plantarum S27 from chicken faeces as a potential probiotic to replace antibiotics: In vivo evidence. Benef. Microbes 2020, 11, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Hai, D.; Kong, L.Y.; Lu, Z.X.; Huang, X.Q.; Bie, X.M. Inhibitory effect of different chicken-derived lactic acid bacteria isolates on drug resistant Salmonella SE47 isolated from eggs. Lett. Appl. Microbiol. 2021, 73, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.P.; Xin, Y.P.; Kong, J.; Guo, T.T. Genetic tools for the development of recombinant lactic acid bacteria. Microb. Cell. Fact. 2021, 20, 118. [Google Scholar] [CrossRef] [PubMed]
- Jacob, B.; Makarewicz, O.; Hartung, A.; Brodt, S.; Roehner, E.; Matziolis, G. In vitro additive effects of dalbavancin and rifampicin against biofilm of Staphylococcus aureus. Sci Rep 2021, 11, 23425. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Behbahani, B.A.; Noshad, M.; Falah, F. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microb. Pathog. 2019, 136, 103677. [Google Scholar] [CrossRef]
- Talib, N.; Mohamad, N.E.; Yeap, S.K.; Hussin, Y.; Aziz, M.N.M.; Masarudin, M.J.; Sharifuddin, S.A.; Hui, Y.W.; Ho, C.L.; Alitheen, N.B. Isolation and Characterization of Lactobacillus spp. from Kefir Samples in Malaysia. Molecules 2019, 24, 2606. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic Activity of Lactobacillus reuteri Strains on the Adhesion Characteristics of Selected Pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Brownlie, E.J.E.; Chaharlangi, D.; Wong, E.O.Y.; Kim, D.; Navarre, W.W. Acids produced by lactobacilli inhibit the growth of commensal Lachnospiraceae and S24-7 bacteria. Gut Microbes 2022, 14, 2046452. [Google Scholar] [CrossRef]
- Wang, H.; Huang, X.J.; Tan, H.Z.; Chen, X.M.; Chen, C.H.; Nie, S.P. Interaction between dietary fiber and bifidobacteria in promoting intestinal health. Food Chem. 2022, 393, 133407. [Google Scholar] [CrossRef]
- Mohsin, M.; Zhang, Z.P.; Yin, G.W. Effect of Probiotics on the Performance and Intestinal Health of Broiler Chickens Infected with Eimeria tenella. Vaccines 2022, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Huang, L.; Zeng, Y.; Li, W.; Zhou, D.; Xie, J.; Xie, J.; Tu, Q.; Deng, D.; Yin, J. Pediococcus pentosaceus: Screening and Application as Probiotics in Food Processing. Front. Microbiol. 2021, 12, 762467. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Li, L.N.; Huang, X.H.; Aleem, M.T.; Habib, Y.J.; Shehata, A.I.; Afzal, M.Z.; Abbas, R.Z.; Abbas, A.; Yin, G.W. Immunogenicity and Protective Efficacy of Probiotics with EtIMP1C against Eimeria tenella Challenge. Pak. Vet. J. 2021, 41, 274–278. [Google Scholar]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus Adhesion to Mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, C.R.; Chen, X.; Jiang, X.R.; Zhao, R.F.; Yuan, Y.; Chen, D.W.; Zhang, C.C.; Lu, M.L.; Lu, Z.; Gu, R.X. In vitrostudies of adhesion properties of six lactic acid bacteria isolated from the longevous population of China. RSC Adv. 2020, 10, 24234–24240. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.M.; Hu, X.Y.; Chen, Y.; Xie, J.H.; Ying, M.X.; Wang, Y.D.; Yu, Q. Differentiated Caco-2 cell models in food-intestine interaction study: Current applications and future trends. Trends Food Sci. Technol. 2021, 107, 455–465. [Google Scholar] [CrossRef]
- Damodharan, K.; Lee, Y.S.; Palaniyandi, S.A.; Yang, S.H.; Suh, J.W. Preliminary probiotic and technological characterization of Pediococcus pentosaceus strain KID7 and in vivo assessment of its cholesterol-lowering activity. Front. Microbiol. 2015, 6, 768. [Google Scholar] [CrossRef] [Green Version]
- El Hage, R.; El Hage, J.; Snini, S.P.; Ammoun, I.; Touma, J.; Rachid, R.; Mathieu, F.; Sabatier, J.-M.; Abi Khattar, Z.; El Rayess, Y. The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin. Antibiotics (Basel, Switz.) 2022, 11, 1147. [Google Scholar] [CrossRef]
- Nwoko, E.; Okeke, I.N. Bacteria autoaggregation: How and why bacteria stick together. Biochem. Soc. Trans. 2021, 49, 1147–1157. [Google Scholar] [CrossRef]
- Stevens, M.R.E.; Luo, T.L.; Vornhagen, J.; Jakubovics, N.S.; Gilsdorf, J.R.; Marrs, C.F.; Moretro, T.; Rickard, A.H. Coaggregation occurs between microorganisms isolated from different environments. FEMS Microbiol. Ecol. 2015, 91, fiv123. [Google Scholar] [CrossRef]
- Wang, J.J.; Wu, Z.C.; Wang, S.; Wang, X.; Zhang, D.M.; Wang, Q.J.; Lin, L.L.; Wang, G.Q.; Guo, Z.X.; Chen, Y.K. Inhibitory effect of probiotic Bacillus spp. isolated from the digestive tract of Rhynchocypris Lagowskii on the adhesion of common pathogenic bacteria in the intestinal model. Microb. Pathog. 2022, 169, 105623. [Google Scholar] [CrossRef]
- Seddik, H.A.; Bendali, F.; Cudennec, B.; Drider, D. Anti-pathogenic and probiotic attributes of Lactobacillus salivarius and Lactobacillus plantarum strains isolated from feces of Algerian infants and adults. Res. Microbiol. 2017, 168, 244–254. [Google Scholar] [CrossRef]
- Liu, C.; Xue, W.J.; Ding, H.; An, C.; Ma, S.J.; Liu, Y. Probiotic Potential of Lactobacillus Strains Isolated From Fermented Vegetables in Shaanxi, China. Front. Microbiol. 2022, 12, 774903. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Ramirez, L.M.; Hernandez-Chinas, U.; Moreno-Guerrero, S.S.; Ramirez-Pacheco, A.; Eslava, C.A. Probiotic Properties and Immunomodulatory Activity of Lactobacillus Strains Isolated from Dairy Products. Microorganisms 2021, 9, 825. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.G.; El Sohaimy, S.A.; El-Sahn, M.A.; Youssef, M.M. Screening of isolated potential probiotic lactic acid bacteria for cholesterol lowering property and bile salt hydrolase activity. Ann. Agric. Sci. 2016, 61, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453, table of contents. [Google Scholar] [CrossRef] [Green Version]
- Bezkorovainy, A. Probiotics: Determinants of survival and growth in the gut. Am. J. Clin. Nutr. 2001, 73, 399S–405S. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.W.; Ma, H.; Yu, H.J.; Qin, G.Y.; Tan, Z.F.; Wang, Y.P.; Pang, H.L. Screening of Lactobacillus plantarum subsp. plantarum with Potential Probiotic Activities for Inhibiting ETEC K88 in Weaned Piglets. Molecules 2020, 25, 4481. [Google Scholar] [CrossRef]
- Gilliland, S.E. Health and nutritional benefits from lactic acid bacteria. FEMS Microbiol. Rev. 1990, 7, 175–188. [Google Scholar] [CrossRef]
- Jin, L.Z.; Ho, Y.W.; Abdullah, N.; Jalaludin, S. Acid and bile tolerance of Lactobacillus isolated from chicken intestine. Lett. Appl. Microbiol. 1998, 27, 183–185. [Google Scholar] [CrossRef]
- Noriega, L.; Gueimonde, M.; Sanchez, B.; Margolles, A.; de los Reyes-Gavilan, C.G. Effect of the adaptation to high bile salts concentrations on glycosidic activity, survival at low PH and cross-resistance to bile salts in Bifidobacterium. Int. J. Food Microbiol. 2004, 94, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, D.L.; O’Neill, B.L.; Hone, D.M.; Metcalf, E.S. Differential early interactions between Salmonella enterica serovar Typhi and two other pathogenic Salmonella serovars with intestinal epithelial cells. Infect. Immun. 1998, 66, 2310–2318. [Google Scholar] [CrossRef] [PubMed]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Vasiee, A.; Falah, F.; Mortazavi, S.A. Evaluation of probiotic potential of autochthonous lactobacilli strains isolated from Zabuli yellow kashk, an Iranian dairy product. J. Appl. Microbiol. 2022, 133, 3201–3214. [Google Scholar] [CrossRef]
- Barzegari, A.; Kheyrolahzadeh, K.; Khatibi, S.M.H.; Sharifi, S.; Memar, M.Y.; Vahed, S.Z. The Battle of Probiotics and Their Derivatives Against Biofilms. Infect. Drug Resistance 2020, 13, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Gou, H.-Z.; Zhang, Y.-L.; Ren, L.-F.; Li, Z.-J.; Zhang, L. How do intestinal probiotics restore the intestinal barrier? Front. Microbiol. 2022, 13, 929346. [Google Scholar] [CrossRef]
- Vasiee, A.; Falah, F.; Behbahani, B.A.; Tabatabaee-yazdi, F. Probiotic characterization of Pediococcus strains isolated from Iranian cereal-dairy fermented product: Interaction with pathogenic bacteria and the enteric cell line Caco-2. J. Biosci. Bioeng. 2020, 130, 471–479. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Chen, S.; Liao, C.; Jia, Y.; Li, J.; Shang, K.; Chen, J.; Cao, P.; Li, W.; Li, Y.; et al. Probiotic Properties of Chicken-Derived Highly Adherent Lactic Acid Bacteria and Inhibition of Enteropathogenic Bacteria in Caco-2 Cells. Microorganisms 2022, 10, 2515. https://doi.org/10.3390/microorganisms10122515
Wang P, Chen S, Liao C, Jia Y, Li J, Shang K, Chen J, Cao P, Li W, Li Y, et al. Probiotic Properties of Chicken-Derived Highly Adherent Lactic Acid Bacteria and Inhibition of Enteropathogenic Bacteria in Caco-2 Cells. Microorganisms. 2022; 10(12):2515. https://doi.org/10.3390/microorganisms10122515
Chicago/Turabian StyleWang, Pudi, Songbiao Chen, Chengshui Liao, Yanyan Jia, Jing Li, Ke Shang, Jian Chen, Pinghua Cao, Wang Li, Yuanxiao Li, and et al. 2022. "Probiotic Properties of Chicken-Derived Highly Adherent Lactic Acid Bacteria and Inhibition of Enteropathogenic Bacteria in Caco-2 Cells" Microorganisms 10, no. 12: 2515. https://doi.org/10.3390/microorganisms10122515