Characterization of the Three New Kayviruses and Their Lytic Activity Against Multidrug-Resistant Staphylococcus aureus

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Isolation and Purification of Phages
2.3. Phage Propagation
2.4. Electron Microscopy Imaging of Phages
2.5. Efficiency of Phage Adsorption
2.6. One-Step Growth Analysis
2.7. Determination of the Host Range
2.8. Sensitivity of Phage Particles to Temperature, pH and Chloroform and Virion Stability
2.9. Spectrophotometric Assay of Phage-Treated Liquid
2.10. Lysogeny Test for Verification of Lytic Cycle
2.11. Extraction of Phage DNA
2.12. Phage Genome Sequencing and Bioinformatics Analysis
2.13. Statistical Analysis
3. Results
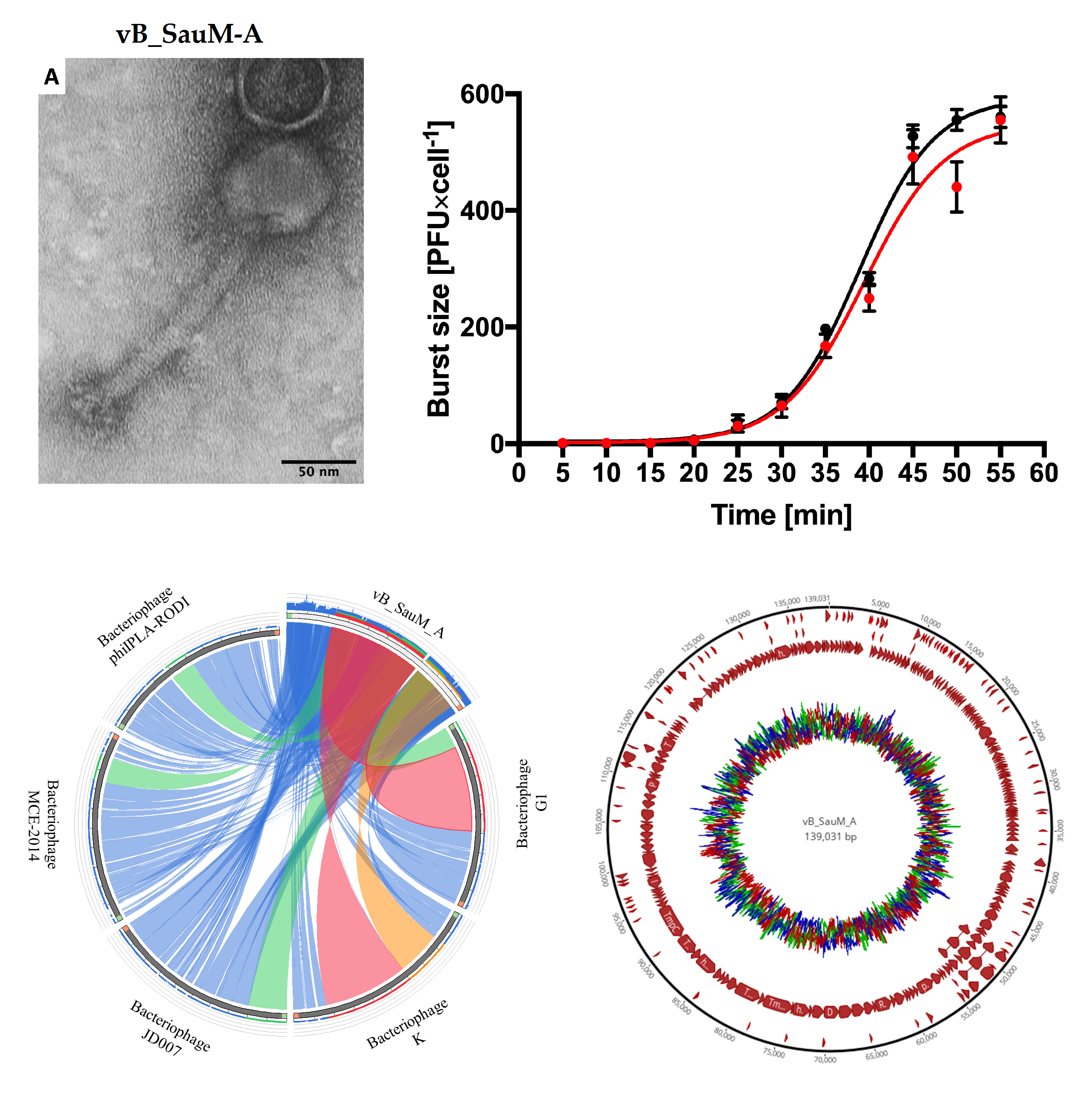
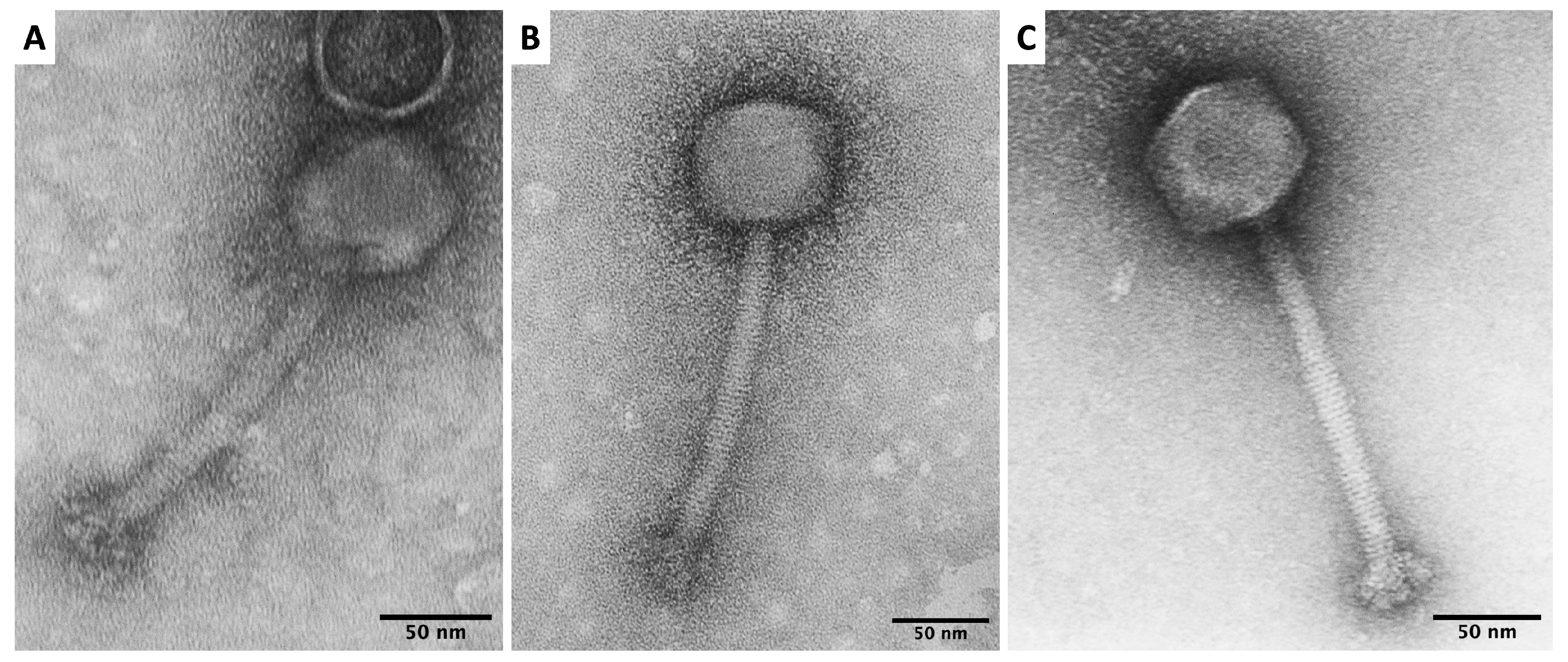
3.1. Virion Morphology
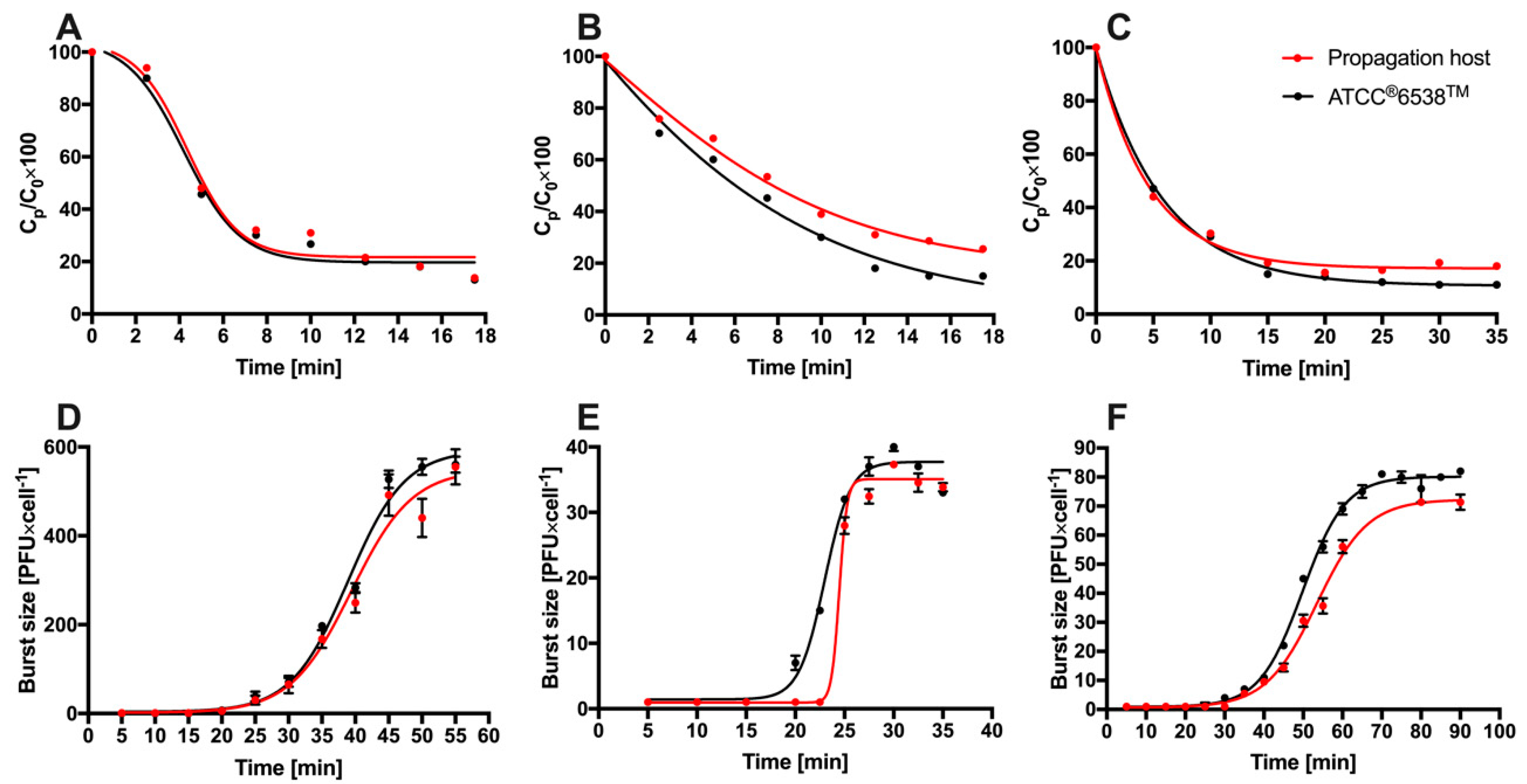
3.2. Analysis of the Bacteriophage Adsorption Rate and One-Step Growth Curves
3.3. Determination of the Host Range
3.4. Sensitivity of Phage Particles to Temperature, pH and Chloroform and Virion Stability Analysis
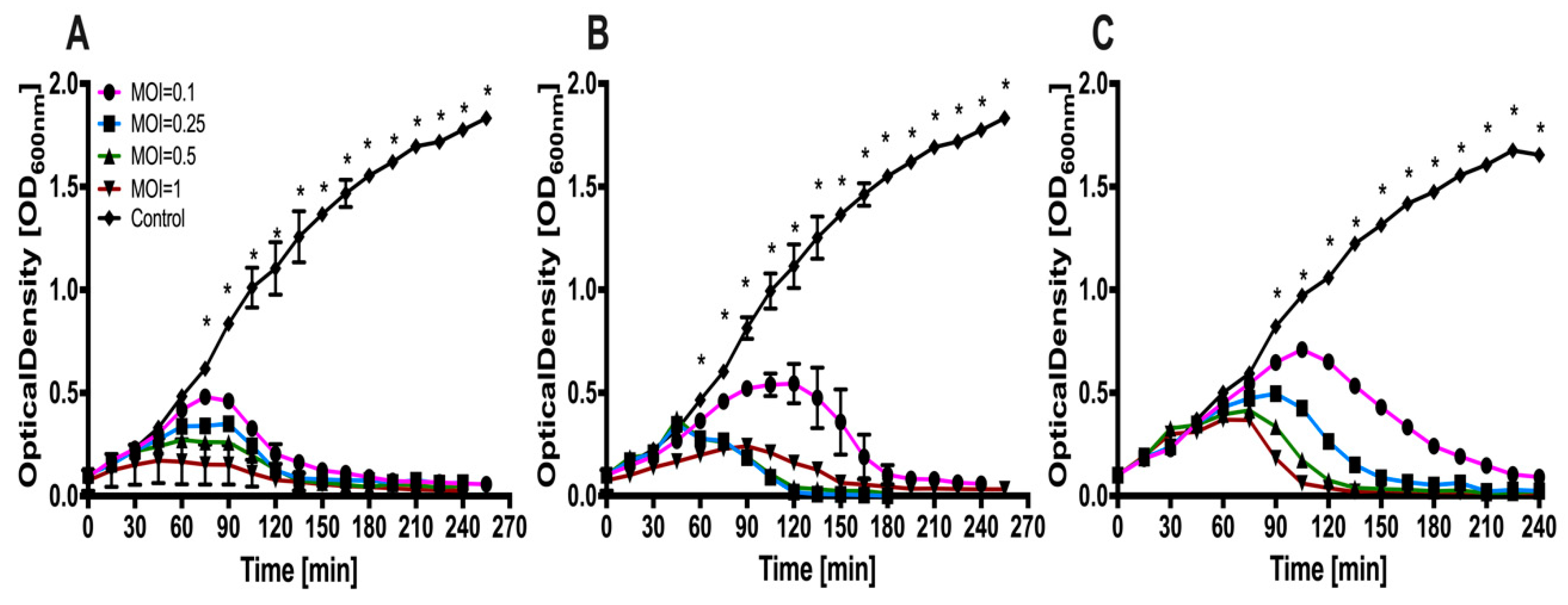
3.5. Lysis Profiles
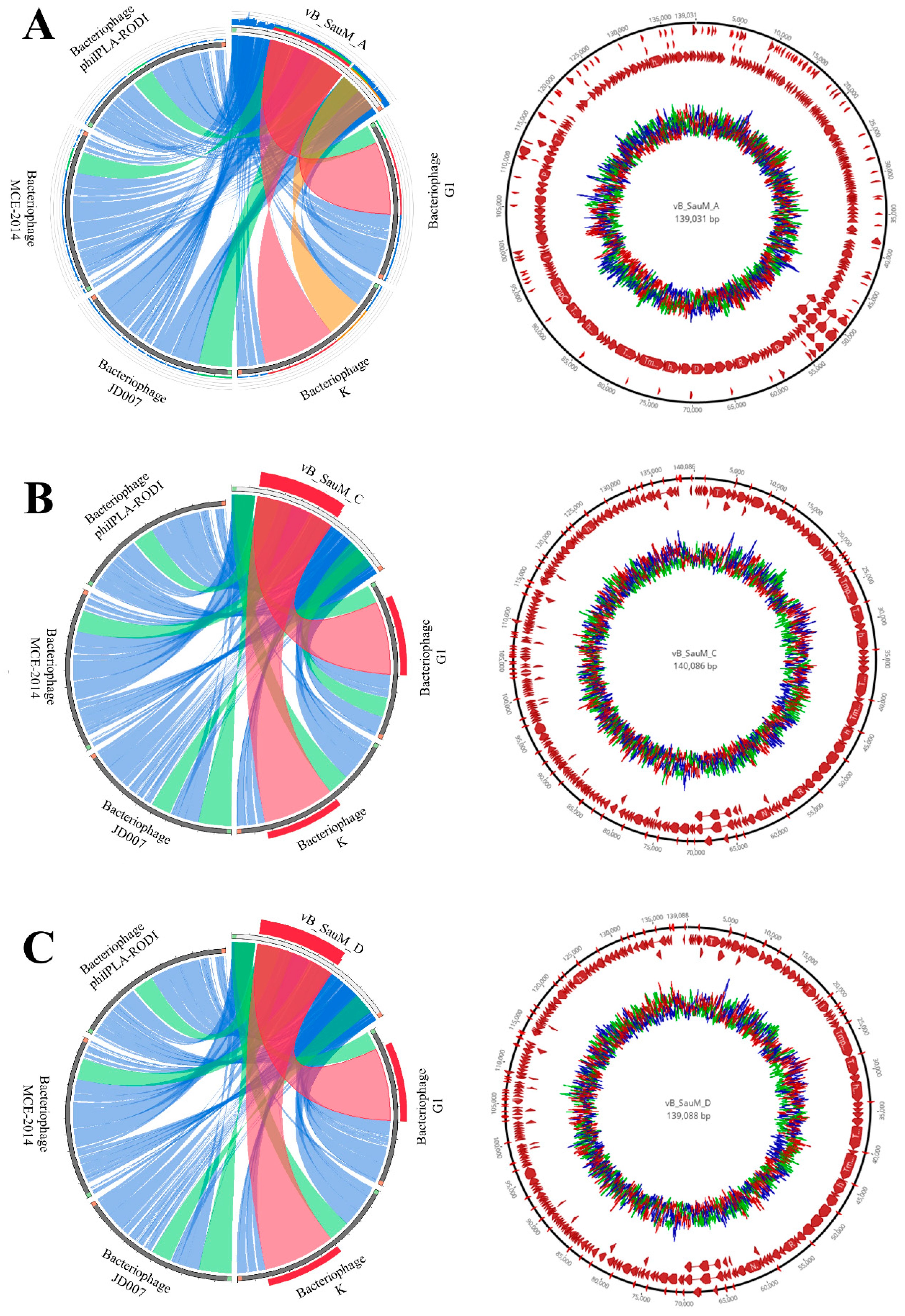
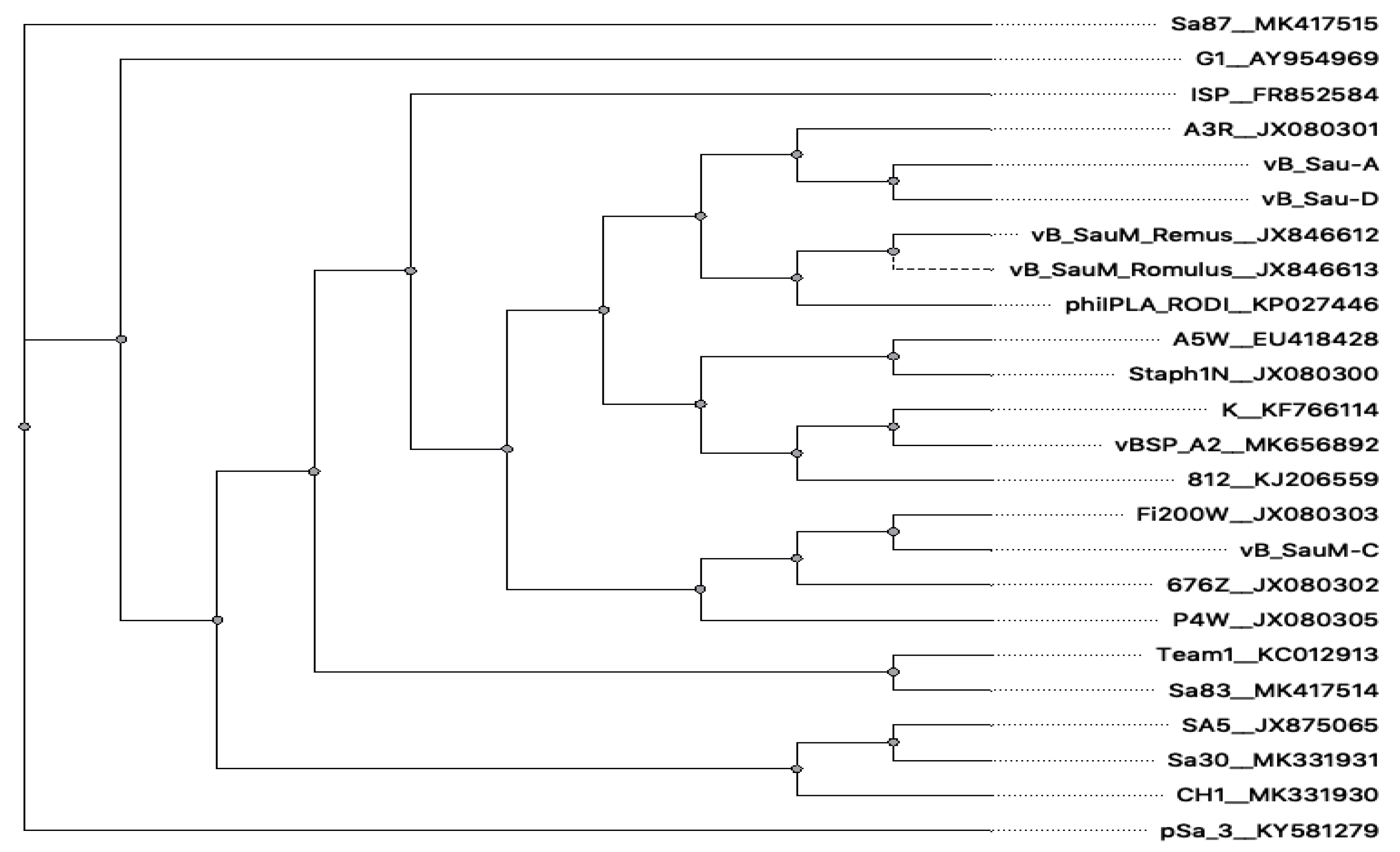
3.6. Phage Genome Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe—Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net) 2017; ECDC: Stockholm, Sweden, 2018; ISBN 978-92-9498-279-7. Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2017 (accessed on 14 May 2019).
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G.J. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barcelo, C. Phage Therapy Faces Evolutionary Challenges. Viruses 2018, 10, 323. [Google Scholar] [CrossRef] [PubMed]
- Abhilash, M.; Vidya, A.G.; Jagadevi, T. Bacteriophage Therapy: A War Against Antibiotic Resistant Bacteria. Internet J. Altern. Med. 2008, 7, e17744. [Google Scholar]
- Pantůček, R.; Rosypalová, A.; Doškař, J.; Kailerová, J.; Růžičková, V.; Borecká, P.; Snopková, Š.; Horváth, R.; Götz, F.; Rosypal, S. The Polyvalent Staphylococcal Phage φ812:Its Host-Range Mutants and Related Phages. Virology 1998, 246, 241–252. [Google Scholar] [CrossRef]
- Melo, L.D.R.; Brandão, A.; Akturk, E.; Santos, S.B.; Azeredo, J. Characterization of a new Staphylococcus aureus Kayvirus harboring a lysin active against biofilms. Viruses 2018, 10, 182. [Google Scholar] [CrossRef]
- Leskinen, K.; Tuomala, H.; Wicklund, A.; Horsma-Heikkinen, J.; Kuusela, P.; Skurnik, M.; Kiljunen, S. Characterization of vB_SauM-fRuSau02, a Twort-Like Bacteriophage Isolated from a Therapeutic Phage Cocktail. Viruses 2017, 9, 258. [Google Scholar] [CrossRef]
- Abatángelo, V.; Peressutti Bacci, N.; Boncompain, C.A.; Amadio, A.A.F.; Carrasco, S.; Suárez, C.A.; Morbidoni, H.R. Broad-range lytic bacteriophages that kill Staphylococcus aureus local field strains. PLoS ONE 2017, 12, e0181671. [Google Scholar] [CrossRef]
- Vandersteegen, K.; Mattheus, W.; Ceyssens, P.-J.; Bilocq, F.; De Vos, D.; Pirnay, J.-P.; Noben, J.-P.; Merabishvili, M.; Lipinska, U.; Hermans, K.; et al. Microbiological and Molecular Assessment of Bacteriophage ISP for the Control of Staphylococcus aureus. PLoS ONE 2011, 6, e24418. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, C.; Jofre, J.; Blanch, A.R.; Lucena, F. Development of a feasible method to extract somatic coliphages from sludge, soil and treated biowaste. J. Virol. Methods 2007, 144, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Lee, J.H.; Shin, H.; Kim, M.; Choi, J.; Kang, D.H.; Heu, S.; Ryu, S. Characterization and comparative genomic analysis of a novel bacteriophage, SFP10, simultaneously inhibiting both Salmonella enterica and Escherichia coli O157:H7. Appl. Environ. Microbiol. 2012, 78, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Danis-Wlodarczyk, K.; Olszak, T.; Arabski, M.; Wasik, S.; Majkowska-Skrobek, G.; Augustyniak, D.; Gula, G.; Briers, Y.; Jang, H.B.; Vandenheuvel, D.; et al. Characterization of the Newly Isolated Lytic Bacteriophages KTN6 and KT28 and Their Efficacy against Pseudomonas aeruginosa Biofilm. PLoS ONE 2015, 10, e0127603. [Google Scholar]
- Cui, Z.; Feng, T.; Gu, F.; Li, Q.; Dong, K.; Zhang, Y.; Zhu, Y.; Han, L.; Qin, J.; Guo, X. Characterization and complete genome of the virulent Myoviridae phage JD007 active against a variety of Staphylococcus aureus isolates from different hospitals in Shanghai, China. Virol. J. 2017, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.F.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor: New York, NY, USA, 2001; Volume 1, ISBN 978-087969577-4. [Google Scholar]
- Alves, D.R.; Gaudion, A.; Bean, J.E.; Perez Esteban, P.; Arnot, T.C.; Harper, D.R.; Kot, W.; Hansen, L.H.; Enright, M.C.; Jenkins, A.T.A. Combined use of bacteriophage K and a novel bacteriophage to reduce Staphylococcus aureus biofilm formation. Appl. Environ. Microbiol. 2014, 80, 6694–6703. [Google Scholar] [CrossRef]
- Barylski, J.; Nowicki, G. The Discovery of phiAGATE, A Novel Phage Infecting Bacillus pumilus, Leads to New Insights into the Phylogeny of the Subfamily Spounavirinae. PLoS ONE 2018, 9, e86632. [Google Scholar] [CrossRef]
- Nejman-Faleńczyk, B.; Bloch, S.; Licznerska, K.; Dydecka, A.; Felczykowska, A.; Topka, G.; Węgrzyn, A.; Węgrzyn, G. A small, microRNA-size, ribonucleic acid regulating gene expression and development of Shiga toxin-converting bacteriophage ϕ24B. Sci. Rep. 2015, 5, 10080. [Google Scholar] [CrossRef]
- Dydecka, A.; Bloch, S.; Rizvi, A.; Perez, S.; Nejman-Faleńczyk, B.; Topka, G.; Gasior, T.; Necel, A.; Wegrzyn, G.; Donaldson, L.W.; et al. Bad Phages in Good Bacteria: Role of the Mysterious orf63 of λ and Shiga Toxin-Converting ϕ24B Bacteriophages. Front. Microbiol. 2017, 8, 1618. [Google Scholar] [CrossRef]
- Jurczak-Kurek, A.; Gasior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Jamal, M.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rahman, S.; Das, C.R. Isolation, characterization and efficacy of phage MJ2 against biofilm forming multi-drug resistant Enterobacter cloacae. Folia Microbiol. 2019, 64, 101–111. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.M.; Marmo, P.; Henrici De Angelis, L.; Palmieri, M.; Ciacci, N.; Di Lallo, G.; Demattè, E.; Vannuccini, E.; Lupetti, P.; Rossolini, G.M.; et al. φBO1E, a newly discovered lytic bacteriophage targeting carbapenemase-producing Klebsiella pneumoniae of the pandemic Clonal Group 258 clade II lineage. Sci. Rep. 2017, 7, 2614. [Google Scholar] [CrossRef] [PubMed]
- Topka, G.; Bloch, S.; Nejman-Falenczyk, B.; Gasior, T.; Jurczak-Kurek, A.; Necel, A.; Dydecka, A.; Richert, M.; Wegrzyn, G.; Wegrzyn, A. Characterization of Bacteriophage vB-EcoS-95, Isolated From Urban Sewage and Revealing Extremely Rapid Lytic Development. Front. Microbiol. 2019, 9, 3326. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [Green Version]
- Darzentas, N. Circoletto: Visualizing sequence similarity with Circos. Bioinformatics 2010, 26, 2620–2621. [Google Scholar] [CrossRef]
- O’Flaherty, S.; Ross, R.P.; Meaney, W.; Fitzgerald, G.F.; Elbreki, M.F.; Coffey, A. Potential of the Polyvalent Anti-Staphylococcus Bacteriophage K for Control of Antibiotic-Resistant Staphylococci from Hospitals. Appl. Environ. Microbiol. 2005, 71, 1836–1842. [Google Scholar] [CrossRef]
- Golkar, Z.; Bagasra, O.; Gene Pace, D. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries. 2014, 8, 129–136. [Google Scholar] [CrossRef]
- Jensen, K.C.; Hair, B.B.; Wienclaw, T.M.; Murdock, M.H.; Hatch, J.B.; Trent, A.T.; White, T.D.; Haskell, K.J.; Berges, B.K. Isolation and Host Range of Bacteriophage with Lytic Activity against Methicillin-Resistant Staphylococcus aureus and Potential Use as a Fomite Decontaminant. PLoS ONE 2015, 10, e0131714. [Google Scholar] [CrossRef] [PubMed]
- Hudson, L.O.; Murphy, C.R.; Spratt, B.G.; Enright, M.C.; Elkins, K.; Nguyen, C.; Terpstra, L.; Gombosev, A.; Kim, D.; Hannah, P.; et al. Diversity of Methicillin-Resistant Staphylococcus aureus (MRSA) Strains Isolated from Inpatients of 30 Hospitals in Orange County, California. PLoS ONE 2013, 8, e62117. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, M.A.; Ramundo, M.S.; Américo, M.A.; de Mattos, M.C.; Souza, R.R.; Ramos-Júnior, E.S.; Coelho, L.R.; Morrot, A.; Melo, P.A.; Fracalanzza, S.E.L.; et al. A comparison of virulence patterns and in vivo fitness between hospital- and community-acquired methicillin-resistant Staphylococcus aureus related to the USA400 clone. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.M.S. What is behind the epidemiological difference between community-acquired and health-care associated methicillin-resistant Staphylococcus aureus? Virulence 2017, 8, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Nasser, A.; Azizian, R.; Tabasi, M.; Khezerloo, J.K.; Heravi, F.S.; Kalani, M.T.; Sadeghifard, N.; Amini, R.; Pakzad, I.; Radmanesh, A.; et al. Specification of Bacteriophage Isolated Against Clinical Methicillin-Resistant Staphylococcus Aureus. Osong Public Heal. Res. Perspect. 2019, 10, 20–24. [Google Scholar] [CrossRef]
- Ackermann, H.-W. Tailed Bacteriophages: The Order Caudovirales. Adv. Virus Res. 1998, 51, 135–201. [Google Scholar]
- Merabishvili, M.; Pirnay, J.P.; Verbeken, G.; Chanishvili, N.; Tediashvili, M.; Lashkhi, N.; Glonti, T.; Krylov, V.; Mast, J.; Van Parys, L.; et al. Quality-controlled small-scale production of a well-defined bacteriophage cocktail for use in human clinical trials. PLoS ONE 2009, 4, e4944. [Google Scholar] [CrossRef]
- Łobocka, M.; Hejnowicz, M.S.; Dabrowski, K.; Gozdek, A.; Kosakowski, J.; Witkowska, M.; Ulatowska, M.I.; Weber-Dabrowska, B.; Kwiatek, M.; Parasion, S.; et al. Genomics of Staphylococcal Twort-like Phages-Potential Therapeutics of the Post-Antibiotic Era. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2012; Volume 83, pp. 143–216. [Google Scholar]
- Barylski, J.; Enault, F.; Dutilh, B.E.; Schuller, M.B.; Edwards, R.A.; Gillis, A.; Klumpp, J.; Knezevic, P.; Krupovic, M.; Kuhn, J.H.; et al. Analysis of spounaviruses as a case study for the overdue reclassification of tailed bacteriophages. Syst. Biol. 2019, 220434. [Google Scholar]
- Mao, J.; Schmelcher, M.; Harty, W.J.; Foster-Frey, J.; Donovan, D.M. Chimeric Ply187 endolysin kills Staphylococcus aureus more effectively than the parental enzyme. FEMS Microbiol. Lett. 2013, 342, 30–36. [Google Scholar] [CrossRef]
- Zhang, Q.; Xing, S.; Sun, Q.; Pei, G.; Cheng, S.; Liu, Y.; An, X.; Zhang, X.; Qu, Y.; Tong, Y. Characterization and complete genome sequence analysis of a novel virulent Siphoviridae phage against Staphylococcus aureus isolated from bovine mastitis in Xinjiang, China. Virus Genes 2017, 53, 464–476. [Google Scholar] [CrossRef]
- Rodela, M.L.; Sabet, S.; Peterson, A.; Dillon, J.G. Broad Environmental Tolerance for a Salicola Host-Phage Pair Isolated from the Cargill Solar Saltworks, Newark, CA, USA. Microorganisms 2019, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages-review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Casey, E.; van Sinderen, D.; Mahony, J. In vitro characteristics of phages to guide ‘real life’ phage therapy suitability. Viruses 2018, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, K. Phage therapy: What factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 2019, 2000–2025. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef]
- Cha, Y.; Chun, J.; Son, B.; Ryu, S. Characterization and Genome Analysis of Staphylococcus aureus Podovirus CSA13 and Its Anti-Biofilm Capacity. Viruses 2019, 11, 54. [Google Scholar] [CrossRef]
- Oliveira, H.; Pinto, G.; Oliveira, A.; Noben, J.P.; Hendrix, H.; Lavigne, R.; Łobocka, M.; Kropinski, A.M.; Azeredo, J. Characterization and genomic analyses of two newly isolated Morganella phages define distant members among Tevenvirinae and Autographivirinae subfamilies. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Xie, Y.; Wahab, L.; Gill, J.J. Development and validation of a microtiter plate-based assay for determination of bacteriophage host range and virulence. Viruses 2018, 10, 189. [Google Scholar] [CrossRef]
- Rahman, M.; Kim, S.; Kim, S.M.; Seol, S.Y.; Kim, J. Characterization of induced Staphylococcus aureus bacteriophage SAP-26 and its anti-biofilm activity with rifampicin. Biofouling 2011, 27, 1087–1093. [Google Scholar] [CrossRef]
- Zhang, L.; Bao, H.; Wei, C.; Zhang, H.; Zhou, Y.; Wang, R. Characterization and partial genomic analysis of a lytic Myoviridae bacteriophage against Staphylococcus aureus isolated from dairy cows with mastitis in Mid-east of China. Virus Genes 2015, 50, 111–117. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | vB_SauM-A | vB_SauM-C | vB_SauM-D |
|---|---|---|---|
| Head morphology | Icosahedral | Icosahedral | Icosahedral |
| Head length (nm) | 62.99 ± 9.99 | 76.75 ± 5.04 | 72.80 ± 3.66 |
| Head diameter (nm) | 63.33 ± 5.81 | 68.37 ± 4.14 | 65.98 ± 3.11 |
| Tail length (nm) | 149.70 ± 14.65 | 153.51 ± 15.16 | 158.05 ± 6.56 |
| Species | Reference Number | vB_SauM-A | vB_SauM-C | vB_SauM-D |
|---|---|---|---|---|
| Staphylococcus epidermidis | ATCC 12228 | − | − | − |
| Staphylococcus epidermidis | ATCC 14990 | ++ | + | ++ |
| Staphylococcus intermedius | PCM 2405 * | − | − | − |
| Staphylococcus aureus | ATCC 25923 | ++ | +++ | ++ |
| Staphylococcus aureus | ATCC 29213 | ++ | ++ | ++ |
| Escherichia coli | ATCC 11775 | − | − | − |
| Salmonella Typhimurium | ATCC 14028 | − | − | − |
| Salmonella Enteritidis | ATCC 13076 | − | − | − |
| Shigella flexneri | ATCC 12022 | − | − | − |
| Shigella sonnei | ATCC 25931 | − | − | − |
| Proteus vulgaris | ATCC 6380 | − | − | − |
| Proteus mirabilis | ATCC 7002 | − | − | − |
| Yersinia enterocolitica | ATCC 27729 | − | − | − |
| Pseudomonas aeruginosa | ATCC 27853 | − | − | − |
| Enterococcus faecalis | ATCC 29212 | − | − | − |
| Lactococcus lactis | ATCC 19435 | − | − | − |
| Lactobacillus gasseri | ATCC 19992 | − | − | − |
| Lactobacillus acidophilus | ATCC 4356 | − | − | − |
| Bacillus cereus | ATCC 14579 | − | − | − |
| Streptococcus agalactiae | ATCC 12386 | − | − | − |
| Listeria monocytogenes | ATCC 19118 | − | − | − |
| SA 1 | Origin 2 | Antibiotic Resistance 3 | A 4 | C 5 | D 6 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | IP | Pus | Fox | P | E | CC | SXT | TE | − | − | − | ||||
| 17 | IP | Wound | Fox | P | E | CC | SXT | TE | − | − | − | ||||
| 19 | IP | Blood | Fox | P | E | CC | NOR | CIP | SXT | TE | − | − | − | ||
| 20 | IP | Wound | Fox | P | E | CC | SXT | TE | − | − | − | ||||
| 21 | IP | drain content | Fox | P | CC | TE | − | − | − | ||||||
| 23 | IP | Wound | Fox | P | E | CC | NOR | CIP | SXT | TE | − | − | − | ||
| 44 | IP | Throat | Fox | P | E | CC | NOR | CIP | TE | +++ | − | +++ | |||
| 70 | IP | Throat | Fox | P | E | CC | NOR | CIP | TE | ++ | +++ | +++ | |||
| 108 | IP | Urine | Fox | P | E | CC | NOR | CIP | +++ | + | +++ | ||||
| 109 | IP | Wound | Fox | P | E | CC | NOR | CIP | ++ | + | +++ | ||||
| 110 | IP | Fistula | Fox | P | E | CC | NOR | CIP | ++ | + | +++ | ||||
| 111 | IP | Wound | Fox | P | E | CC | NOR | CIP | ++ | +++ | +++ | ||||
| 112 | IP | Bedsore | Fox | P | E | CC | NOR | CIP | TE | GM | − | − | − | ||
| 113 | IP | Wound | Fox | P | E | CC | NOR | CIP | C | ++ | +++ | +++ | |||
| 115 | IP | Wound | Fox | P | E | CC | NOR | CIP | ++ | + | +++ | ||||
| 116 | IP | wound | Fox | P | E | CC | NOR | CIP | ++ | + | +++ | ||||
| 118 | IP | blood | Fox | P | E | CC | NOR | CIP | TE | GM | − | + | − | ||
| 120 | IP | bedsore | Fox | P | E | CC | NOR | CIP | TE | GM | − | + | − | ||
| 121 | IP | bronchial tree discharge | Fox | P | E | CC | NOR | CIP | TE | GM | − | + | − | ||
| 122 | IP | bronchial tree discharge | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 124 | IP | bronchial tree discharge | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 126 | OP | sputum | Fox | P | E | CC | NOR | CIP | TE | GM | − | + | − | ||
| 140 | OP | furuncle | Fox | P | E | CC | NOR | CIP | TE | C | − | − | − | ||
| 149 | IP | nose | Fox | P | E | CC | NOR | CIP | ++ | + | +++ | ||||
| 151 | IP | furuncle | Fox | P | E | CC | C | − | + | − | |||||
| 173 | OP | furuncle | Fox | P | E | CC | TE | C | + | + | − | ||||
| 182 | OP | furuncle | Fox | P | E | CC | C | − | + | − | |||||
| 183 | OP | furuncle | Fox | P | E | CC | TE | C | − | + | − | ||||
| 184 | IP | wound | Fox | P | E | CC | NOR | CIP | TE | GM | − | + | + | ||
| 193 | IP | bedsore | Fox | P | E | CC | NOR | CIP | ++ | +++ | +++ | ||||
| 196 | OP | wound | Fox | P | E | CC | TE | C | − | − | − | ||||
| 198 | OP | wound | Fox | P | E | CC | NOR | CIP | TE | C | GM | − | + | + | |
| 199 | OP | wound | Fox | P | E | CC | TE | C | +++ | +++ | +++ | ||||
| 200 | OP | sputum | Fox | P | E | CC | NOR | CIP | +++ | + | +++ | ||||
| 201 | IP | sputum | Fox | P | E | CC | NOR | CIP | − | + | + | ||||
| 202 | IP | bronchial tree discharge | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 203 | IP | vascular catheter | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 204 | OP | wound | Fox | P | E | CC | − | − | − | ||||||
| 205 | OP | bedsore | Fox | P | E | CC | NOR | CIP | − | + | + | ||||
| 258 | IP | nose | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 271 | IP | wound | Fox | P | E | CC | NOR | CIP | C | +++ | +++ | +++ | |||
| 297 | IP | sore | Fox | P | E | CC | NOR | CIP | C | +++ | +++ | +++ | |||
| 298 | IP | tissue | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 301 | IP | ear | Fox | P | E | CC | NOR | CIP | +++ | ++ | +++ | ||||
| 305 | IP | wound | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 311 | IP | wound | Fox | P | E | CC | NOR | CIP | +++ | ++ | +++ | ||||
| 315 | IP | pus from abscess | Fox | P | E | CC | TE | C | − | + | − | ||||
| 316 | IP | wound | Fox | P | E | NOR | CIP | +++ | + | +++ | |||||
| 317 | IP | skin | Fox | P | E | NOR | CIP | +++ | + | +++ | |||||
| 324 | IP | nose | Fox | P | E | CC | NOR | CIP | +++ | ++ | +++ | ||||
| 340 | IP | wound | Fox | P | E | CC | NOR | CIP | +++ | ++ | +++ | ||||
| 341 | IP | urine | Fox | P | E | NOR | CIP | ++ | + | +++ | |||||
| 342 | IP | endotracheal tube content | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 343 | IP | wound | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 344 | IP | wound | Fox | P | E | CC | C | − | + | − | |||||
| 345 | IP | nose | Fox | P | E | CC | C | − | + | − | |||||
| 351 | IP | pus from abscess | Fox | P | E | CC | C | − | + | − | |||||
| 352 | IP | nose | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 353 | IP | nose | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 355 | IP | endotracheal tube content | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 357 | IP | ear | Fox | P | NOR | CIP | +++ | +++ | +++ | ||||||
| 358 | IP | bronchial aspirate | Fox | P | E | CC | NOR | CIP | C | +++ | +++ | +++ | |||
| 366 | IP | wound | Fox | P | E | CC | NOR | CIP | ++ | +++ | +++ | ||||
| 367 | IP | wound | Fox | P | E | CC | NOR | CIP | +++ | +++ | +++ | ||||
| 369 | IP | nose | Fox | P | E | NOR | CIP | − | + | − | |||||
| 370 | IP | endotracheal tube content | Fox | P | E | CC | NOR | CIP | ++ | +++ | +++ | ||||
| 371 | IP | bronchoalveolar lavage fluid | Fox | P | TE | C | +++ | +++ | ++ | ||||||
| Phage Name | Phage Survivability in Studied Conditions (Relative Phage Titer in %) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 40 °C (40 min) | 62 °C (40 min) | pH 2 (2 h) | pH 3 (2 h) | pH 4 (2 h) | pH 6 (2 h) | pH 8 (2 h) | pH 10 (2 h) | pH 12 (2 h) | CHCl3 (4 °C; 1 h) | |
| vB_SauM-A | 58 | 0 | 0 | 0 | 8 | 100 | 83 | 0 | 0 | 97 |
| vB_SauM-C | 59 | 0 | 0 | 0 | 20 | 100 | 100 | 0 | 0 | 80 |
| vB_SauM-D | 43 | 0 | 0 | 0 | 87 | 100 | 95 | 0 | 0 | 100 |
| Phage Name | Genome Properties | ||
|---|---|---|---|
| Genome Length | GC content (%) | Phage K Similarity (%) | |
| vB_SauM-A | 139,031 | 30.45 | 97 |
| vB_SauM-C | 140,086 | 30.43 | 95 |
| vB_SauM-D | 139,088 | 30.46 | 97 |
| Gene Group | vB_SauM-A | vB_SauM-C | vB_SauM-D |
|---|---|---|---|
| DNA metabolisms | nucleotide kinase (orf613), NTP pyrophosphohydrolase (orf612), nucleoside 2-deoxyribosyltransferase (orf602), RNA ligase (orf601), ribonuclease H (orf559), HNH endonuclease (orf587, 580), intron-encoded nuclease (orf1593), endonuclease (orf201, 264), homing endonuclease (orf206), ribose-phosphate pyrophosphokinase (orf280) | HNH endonuclease (orf312,319, 392, 688), ribonuclease H (orf2411), NTP pyrophosphohydrolase (orf343) nucleotide kinase (orf344), ribose-phosphate pyrophosphokinase (orf187), intron-encoded nuclease (orf216), | ribonuclease H (orf594), HNH endonuclease (orf580, orf587), RNA ligase (orf601), NTP pyrophosphohydrolase (orf612), ribonuclease H (orf453), intron-encoded nuclease (orf592), |
| DNA replication | DNA helicase (orf177, 180), recombination-related endonuclease (orf181, 184), anti-sigma factor (orf186), DNA primase (orf187), resolvase (orf190), ribonucleotide reductase stimulatory protein (orf191), ribonucleotide reductase large subunit (orf193), ribonucleotide reductase small subunit (orf195), DNA polymerase I (orf208), recombinase (orf218), sigma factor (orf221), DNA repair exonuclease (orf230), sliding clamp inhibitor (orf252), nucleotidyl transferase (orf642), terminase large subunit (orf101) | nucleotidyl transferase (orf373), recombinase (orf125), endonuclease (orf669), DNA polymerase I (orf107), ribonucleotide reductase large subunit (orf100), ribonucleotide reductase small subunit (orf638), resolvase (orf97), DNA primase (orf94), anti-sigma factor (orf93), recombination-related endonuclease (orf91, 88), helicase (orf87, 84), terminase large subunit (orf8) | DNA polymerase I (orf200), anti-sigma factor (orf186), recombination-related endonuclease (orf181), sigma factor (orf221), DNA helicase (orf177, 180), DNA primase (orf187), DNA repair exonuclease (orf230), ribonucleotide reductase small subunit (orf227), terminase large subunit (orf101), ribonucleotide reductase large subunit (orf193), recombinase (orf218) |
| Head | portal protein (orf110), prohead protease (orf112), major capsid protein (orf115) | major capsid protein (orf22), prohead protease (orf19), portal protein (orf17), putative virion protein (orf49) | portal protein (orf110), major capsid protein (orf115), prohead protease (orf112) |
| neck protein | neck protein (orf120, 122) | putative neck protein (orf29, 27) | neck protein (orf120, 122) |
| Tail | major tail protein (orf38), tail sheath protein (orf127), tail tube protein (orf129), tail assembly chaperone(orf137), tape-measure protein (orf140), tail murein hydrolase (orf145), cysteine protease (orf147), tail protein (orf226) | major tail protein (orf366), tail protein (orf 133), tail central spike (orf55), tail murein hydrolase (orf52), tape measure protein (orf47), tail morphogenetic protein (orf241), tail assembly chaperone (orf44), tail tube protein (orf36), tail sheath protein (orf34), baseplate hub assembly protein (orf31) | major tail protein (orf38), tape-measure protein (orf140), tail murein hydrolase (orf145), nicotinamide phosphoribosyl transferase (orf381), tail morphogenetic protein (orf55), adsorption-associated tail protein (orf162), tail assembly chaperone (orf137), tail sheath (orf127), tail tube protein (orf129), receptor binding protein (orf174, 170), tail central spike (orf148) |
| baseplate | baseplate component (orf151, 155, 173), baseplate wedge subunit (orf154), baseplate protein (orf160), receptor binding protein (orf170), oxidoreductase (orf197) | putative baseplate component (orf80, 60, 58), receptor binding protein (orf81), receptor binding protein (orf77) baseplate protein (orf67) baseplate component (orf62), baseplate wedge subunit (orf61) | peptidoglycan binding protein (orf604), baseplate component (orf153, 155,160, 173), baseplate wedge subunit (orf154), baseplate hub assembly protein (orf124) |
| Tail fiber | tail fiber protein (orf169) | tail fiber protein (orf69), Tail fiber protein complex (orf65) | tail fiber protein (orf169) |
| Lysis | lytic transglycosylase (orf588), N-acetylmuramoyl-L-alanine amidase (orf578), holin (orf575) | holin (orf307), N-acetylmuramoyl-L-alanine amidase (orf313), lytic transglycosylase (orf320) | holin (orf575), N-acetylmuramoyl-L-alanine amidase (orf581, 578) lytic transglycosylase (orf588) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łubowska, N.; Grygorcewicz, B.; Kosznik-Kwaśnicka, K.; Zauszkiewicz-Pawlak, A.; Węgrzyn, A.; Dołęgowska, B.; Piechowicz, L. Characterization of the Three New Kayviruses and Their Lytic Activity Against Multidrug-Resistant Staphylococcus aureus. Microorganisms 2019, 7, 471. https://doi.org/10.3390/microorganisms7100471
Łubowska N, Grygorcewicz B, Kosznik-Kwaśnicka K, Zauszkiewicz-Pawlak A, Węgrzyn A, Dołęgowska B, Piechowicz L. Characterization of the Three New Kayviruses and Their Lytic Activity Against Multidrug-Resistant Staphylococcus aureus. Microorganisms. 2019; 7(10):471. https://doi.org/10.3390/microorganisms7100471
Chicago/Turabian StyleŁubowska, Natalia, Bartłomiej Grygorcewicz, Katarzyna Kosznik-Kwaśnicka, Agata Zauszkiewicz-Pawlak, Alicja Węgrzyn, Barbara Dołęgowska, and Lidia Piechowicz. 2019. "Characterization of the Three New Kayviruses and Their Lytic Activity Against Multidrug-Resistant Staphylococcus aureus" Microorganisms 7, no. 10: 471. https://doi.org/10.3390/microorganisms7100471