Candida albicans Ubiquitin and Heat Shock Factor-Type Transcriptional Factors Are Involved in 2-Dodecenoic Acid-Mediated Inhibition of Hyphal Growth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Medium
2.2. Microarray Analysis
2.3. Gene Deletion and Tagging
2.4. Microscopic Assessment of C. albicans Strains Hyphal Formation
2.5. Protein Extraction and Western Blotting
3. Results
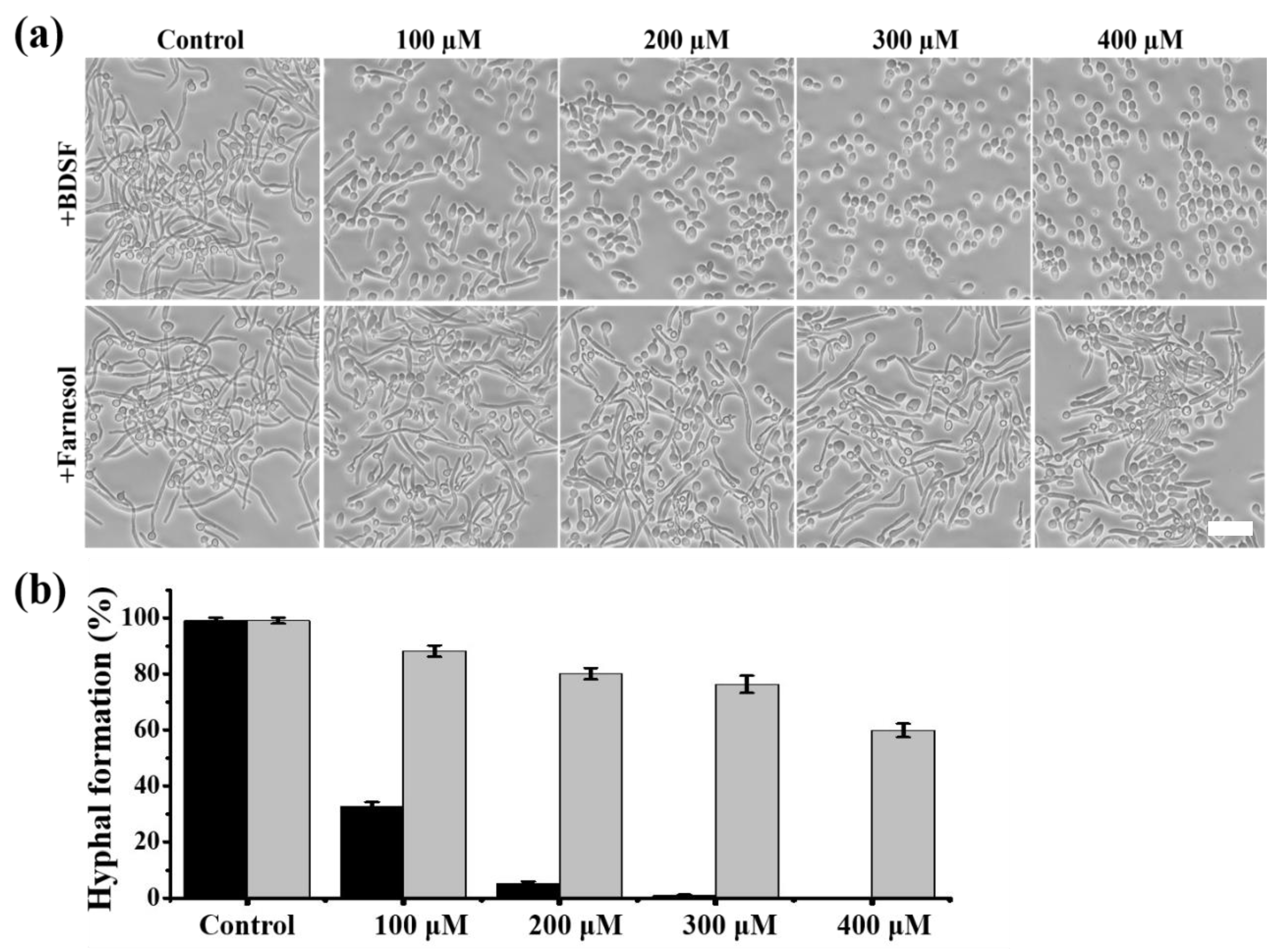
3.1. BDSF Inhibits Filament Formation in C. albicans
3.2. Differential Gene Expression in C. albicans Treated with BDSF
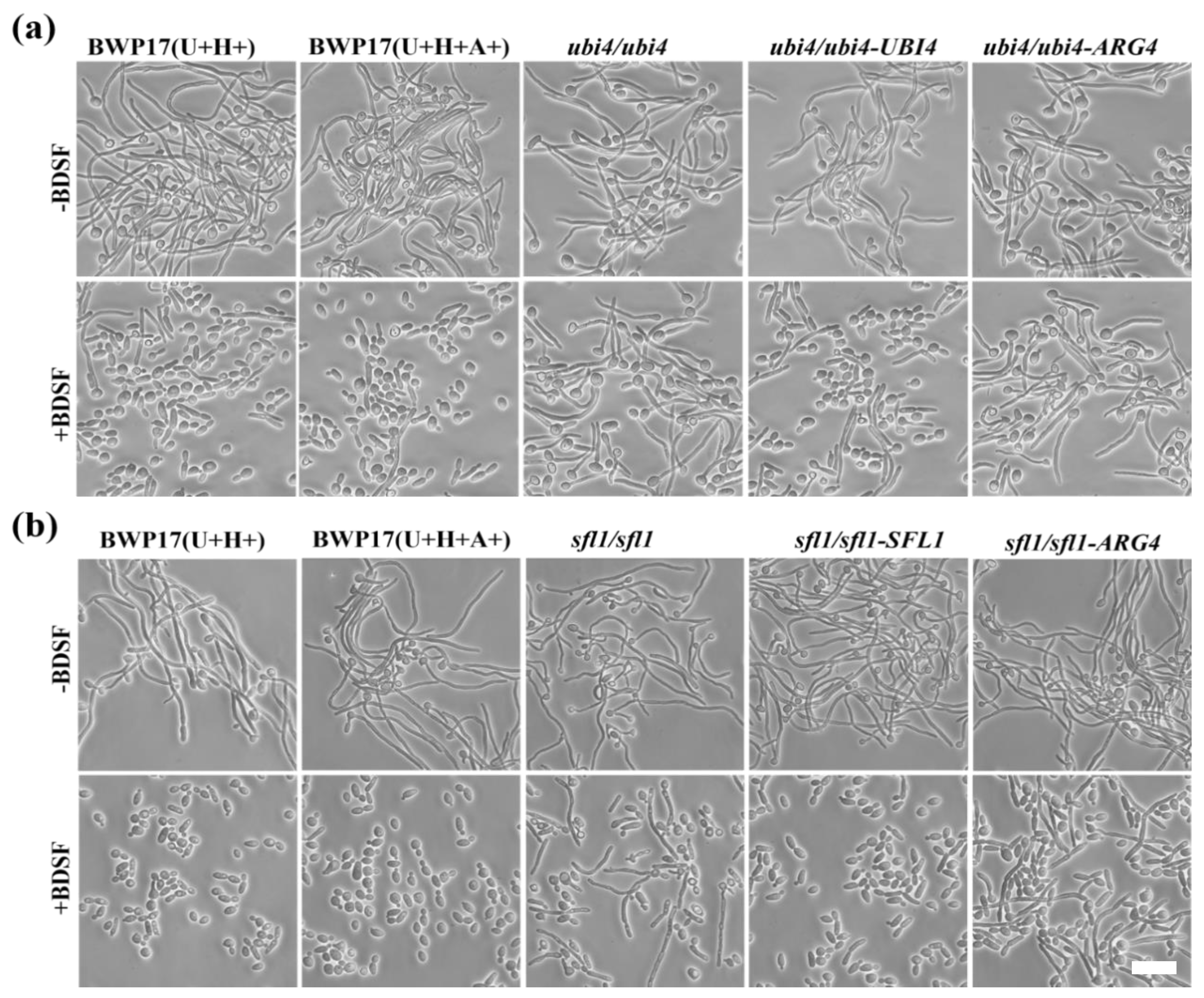
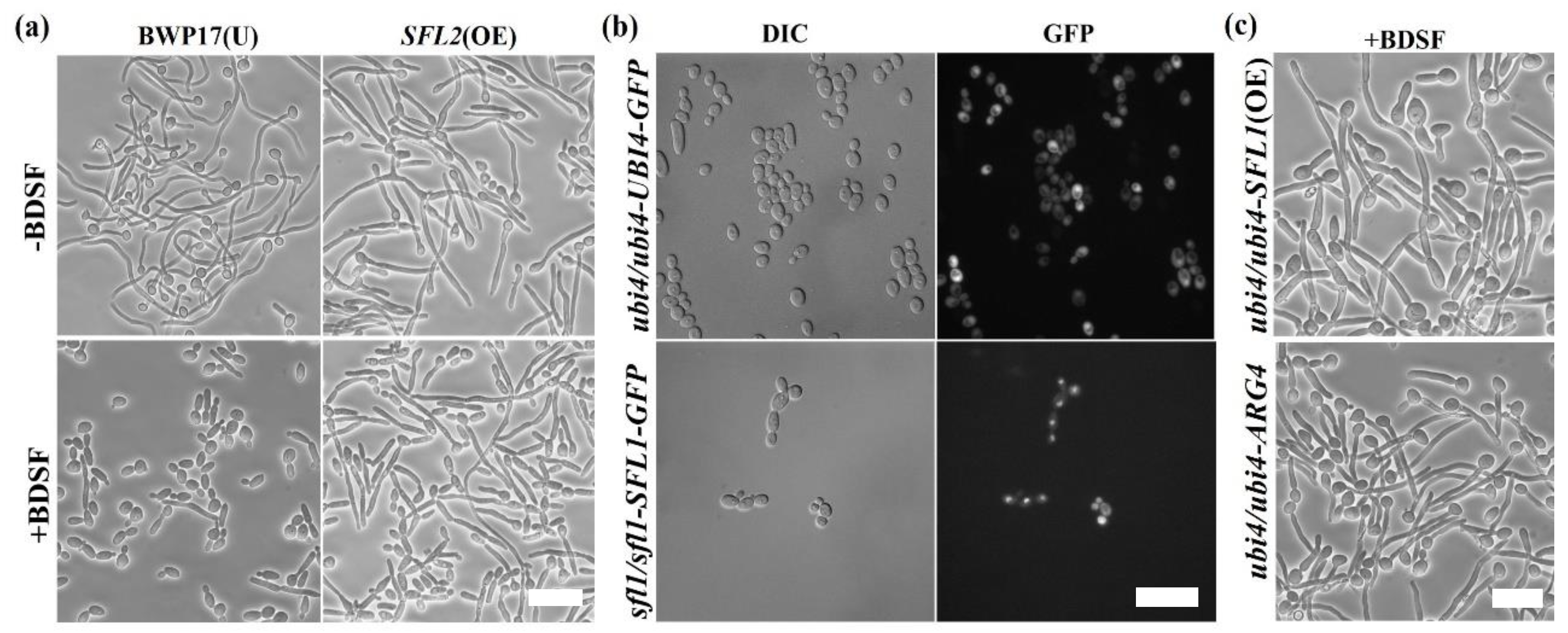
3.3. Screening for BDSF-Resistant Mutants
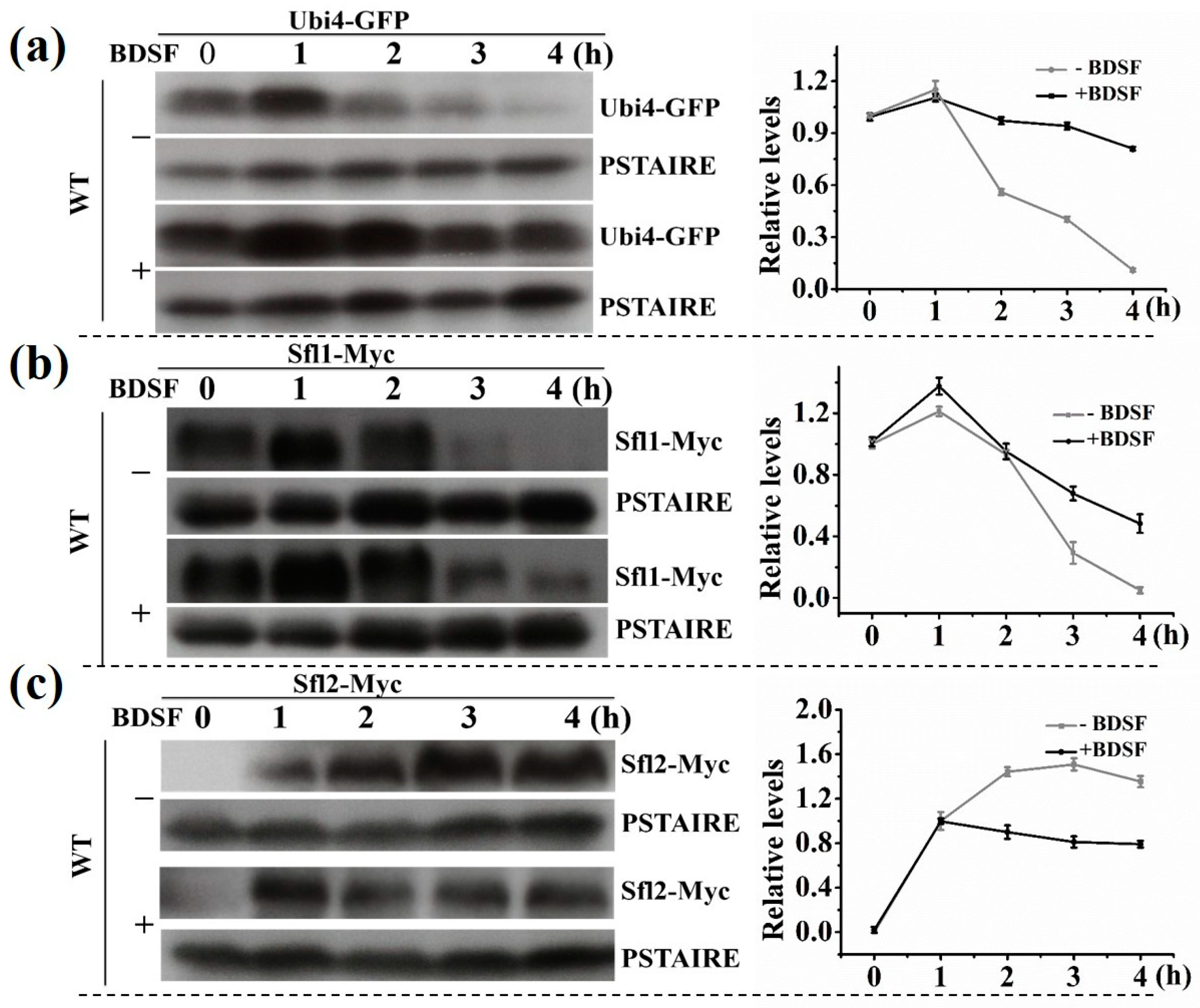
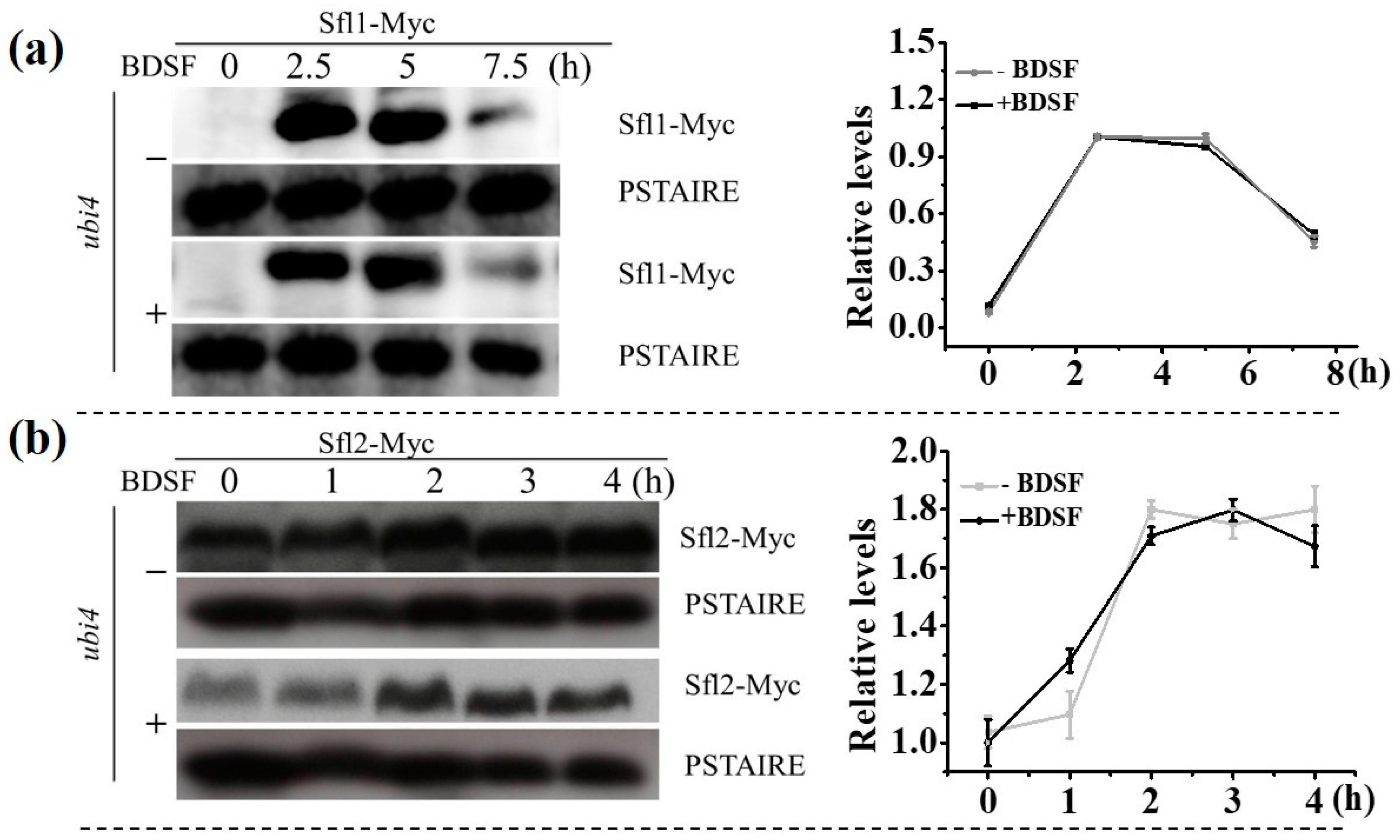
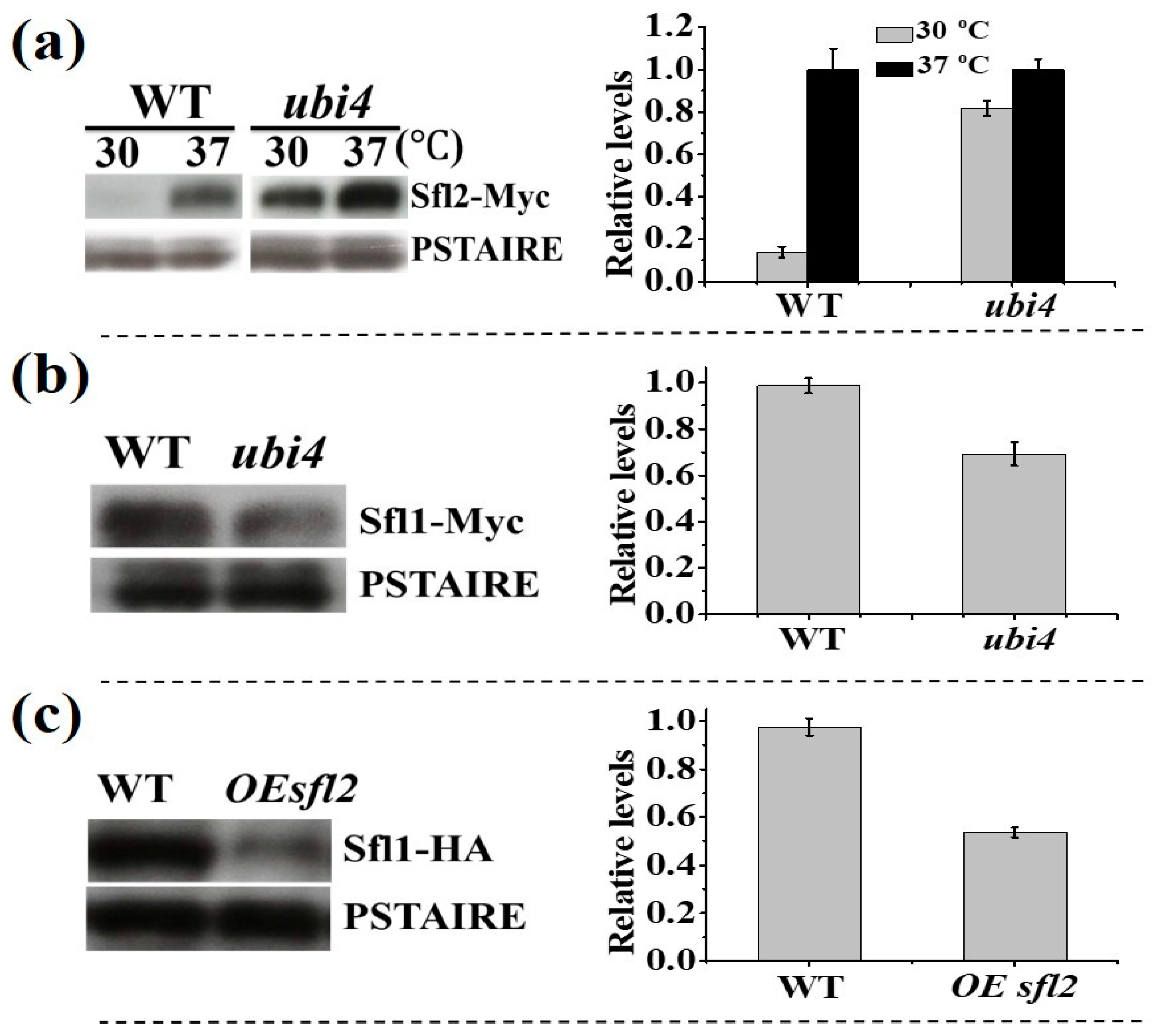
3.4. The Change in Expression Level of Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. R 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y. Fungal adenylyl cyclase acts as a signal sensor and integrator and plays a central role in interaction with bacteria. PLoS Pathog. 2013, 9, e1003612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallick, E.M.; Bennett, R.J. Sensing of the microbial neighborhood by Candida albicans. PLoS Pathog. 2013, 9, e1003661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.L.; Hu, Y.L.; Yin, Z.X.; Zeng, G.S.; Li, D.; Zhang, Y.Q.; Xu, Z.H.; Guan, X.M.; Weng, L.X.; Wang, L.H. Cis-2-dodecenoic acid mediates its synergistic effect with triazoles by interfering with efflux pumps in fluconazole-resistant Candida albicans. Biomed. Environ. Sci. 2019, 32, 199–209. [Google Scholar] [CrossRef]
- Santolaya, M.E.; Thompson, L.; Benadof, D.; Tapia, C.; Legarraga, P.; Cortés, C.; Rabello, M.; Valenzuela, R.; Rojas, P.; Rabagliati, R.; et al. A prospective, multi-center study of Candida bloodstream infections in Chile. PLoS ONE 2019, 14, e0212924. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive Candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Yang, D.; Lv, X.; Xue, L.; Yang, N.; Hu, Y.; Weng, L.; Fu, N.; Wang, L.; Dong, X. A lipase-responsive antifungal nanoplatform for synergistic photodynamic/photothermal/pharmaco-therapy of azole-resistant Candida albicans infections. Chem. Commun. 2019, 55, 15145–15148. [Google Scholar] [CrossRef]
- Whiteway, M.; Bachewich, C. Morphogenesis in Candida albicans. Annu. Rev. Microbiol. 2007, 61, 529–553. [Google Scholar] [CrossRef] [Green Version]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Hogan, D.A.; Vik, Å.; Kolter, R.A. Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol. Microbiol. 2004, 54, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Su, C.; Unoje, O.; Liu, H. Quorum sensing controls hyphal initiation in Candida albicans through Ubr1-mediated protein degradation. Proc. Natl. Acad. Sci. USA 2014, 111, 1975–1980. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.H.; He, Y.; Gao, Y.; Wu, J.E.; Dong, Y.H.; He, C.; Wang, S.X.; Weng, L.X.; Xu, J.L.; Tay, L.; et al. A bacterial cell-cell communication signal with cross-kingdom structural analogues. Mol. Microbiol. 2004, 51, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Weng, L.X.; Zhang, Y.Q.; Wang, L.H. BDSF inhibits Candida albicans adherence to urinary catheters. Microb. Pathog. 2013, 64, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Zhang, Y.Q.; Hu, Y.L.; Weng, L.X.; Zeng, G.S.; Wang, L.H. Protective effects of cis-2-dodecenoic acid in an experimental mouse model of vaginal candidiasis. Biomed. Environ. Sci. 2018, 31, 816–828. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, H.; Chen, J. Candida albicans Sfl2, a temperature-induced transcriptional regulator, is required for virulence in a murine gastrointestinal infection model. FEMS Yeast Res. 2011, 11, 209–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Su, C.; Liu, H. Candida albicans hyphal initiation and elongation. Trends Microbiol. 2014, 22, 707–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Li, D.; Li, W.; Wang, Y.; Sang, J. CaSfl1 plays a dual role in transcriptional regulation in Candida albicans. Chin. Sci. Bull. 2008, 53, 2624–2631. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Su, C.; Mao, X.; Cao, F.; Chen, J. Roles of Candida albicans Sfl1 in hyphal development. Eukaryot. Cell 2007, 6, 2112–2121. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Wendland, J. Candida albicans Sfl1 suppresses flocculation and filamentation. Eukaryot. Cell 2007, 6, 1736–1744. [Google Scholar] [CrossRef] [Green Version]
- Znaidi, S.; Nesseir, A.; Chauvel, M.; Rossignol, T.; d’Enfert, C. A comprehensive functional portrait of two heat shock factor-type transcriptional regulators involved in Candida albicans morphogenesis and virulence. PLoS Pathog. 2013, 9, e1003519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weathington, N.M.; Mallampalli, R.K. Emerging therapies targeting the ubiquitin proteasome system in cancer. J. Clin. Investig. 2014, 124, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig, P.; Gozalbo, D. Depletion of polyubiquitin encoded by the UBI4 gene confers pleiotropic phenotype to Candida albicans cells. Fungal Genet. Biol. 2003, 39, 70–81. [Google Scholar] [CrossRef]
- Atir-Lande, A.; Gildor, T.; Kornitzer, D. Role for the SCF(CDC4) ubiquitin ligase in Candida albicans morphogenesis. Mol. Biol. Cell 2005, 16, 2772–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cao, Y.Y.; Jia, X.M.; Cao, Y.B.; Gao, P.H.; Fu, X.P.; Ying, K.; Chen, W.S.; Jiang, Y.Y. Cap1p is involved in multiple pathways of oxidative stress response in Candida albicans. Free Radic. Biol. Med. 2006, 40, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.K.; All, O.; Goffena, J.; Loveless, T.; Wilson, T.; Toenjes, K.A. The GRR1 gene of Candida albicans is involved in the negative control of pseudohyphal morphogenesis. Fungal Genet. Biol. 2006, 43, 573–582. [Google Scholar] [CrossRef]
- Wang, L.H.; Zhang, L.H. Inhibitors of Yeast Filamentous Growth and Method of Their Manufacture. U.S. Patent 7915313B2, 29 March 2011. [Google Scholar]
- Zeng, G.; Wang, Y.M.; Chan, F.Y.; Wang, Y. One-step targeted gene deletion in Candida albicans haploids. Nat. Protoc. 2014, 9, 464–473. [Google Scholar] [CrossRef]
- Gao, J.; Wang, H.; Wong, A.H.; Zeng, G.; Huang, Z.; Wang, Y.; Sang, J.; Wang, Y. Regulation of Rfa2 phosphorylation in response to genotoxic stress in Candida albicans. Mol. Microbiol. 2014, 94, 141–155. [Google Scholar] [CrossRef]
- Liu, Q.; Han, Q.; Wang, N.; Yao, G.; Zeng, G.; Wang, Y.; Huang, Z.; Sang, J.; Wang, Y. Tpd3-Pph21 phosphatase plays a direct role in Sep7 dephosphorylation in Candida albicans. Mol. Microbiol. 2016, 101, 109–121. [Google Scholar] [CrossRef]
- Zeng, G.; Wang, Y.M.; Wang, Y. Cdc28-Cln3 phosphorylation of Sla1 regulates actin patch dynamics in different modes of fungal growth. Mol. Biol. Cell 2012, 23, 3485–3497. [Google Scholar] [CrossRef]
- Boon, C.; Deng, Y.; Wang, L.; He, Y.; Xu, J.; Fan, Y.; Pan, S.Q.; Zhang, L. A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition. ISME J. 2007, 2, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cai, C.; Yang, Y.; Weng, L.; Wang, L. Blocking of Candida albicans biofilm formation by cis-2-dodecenoic acid and trans-2-dodecenoic acid. J. Med. Microbiol. 2011, 60, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.; Grussemeyer, C.A.; Spalding, J.R.; Benjamin, D.K.; Reed, S.D. Candida albicans and non-albicans bloodstream infections in adult and pediatric patients: Comparison of mortality and costs. Pediatr. Infect. Dis. J. 2009, 28, 433–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada-Okabe, T.; Mio, T.; Ono, N.; Kashima, Y.; Matsui, M.; Arisawa, M.; Yamada-Okabe, H. Roles of three histidine kinase genes in hyphal development and virulence of the pathogenic fungus Candida albicans. J. Bacteriol. 1999, 181, 7243–7247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhl, M.A.; Biery, M.; Craig, N.; Johnson, A.D. Haploinsufficiency-based large-scale forward genetic analysis of filamentous growth in the diploid human fungal pathogen C. albicans. EMBO J. 2003, 22, 2668–2678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig, P.; Gozalbo, D. Candida albicans UBI3 and UBI4 promoter regions confer differential regulation of invertase production to Saccharomyces cerevisiae cells in response to stress. Int. Microbiol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, M.D.; Stead, D.A.; Argo, E.; MacCallum, D.M.; Brown, A.J.P. Molecular and proteomic analyses highlight the importance of ubiquitination for the stress resistance, metabolic adaptation, morphogenetic regulation and virulence of Candida albicans. Mol. Microbiol. 2011, 79, 1574–1593. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.G.; Lopez-Ribot, J.L. Candidiasis drug discovery and development: New approaches targeting virulence for discovering and identifying new drugs. Expert Opin. Drug. Discov. 2013, 8, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Onyewu, C.; Blankenship, J.R.; Del Poeta, M.; Heitman, J. Ergosterol biosynthesis inhibitors become fungicidal when combined with calcineurin inhibitors against Candida albicans, Candida glabrata, and Candida krusei. Antimicrob. Agents Chemother. 2003, 47, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Silver, P.M.; Oliver, B.G.; White, T.C. Role of Candida albicans transcription factor Upc2p in drug resistance and sterol metabolism. Eukaryot. Cell 2004, 3, 1391–1397. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.W.; Konopka, J.B. Lipid raft polarization contributes to hyphal growth in Candida albicans. Eukaryot. Cell 2004, 3, 675–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.Y.; Cao, Y.B.; Xu, Z.; Ying, K.; Li, Y.; Xie, Y.; Zhu, Z.Y.; Chen, W.S.; Jiang, Y.Y. cDNA microarray analysis of differential gene expression in Candida albicans biofilm exposed to farnesol. Antimicrob. Agents Chemother. 2005, 49, 584–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.A.; Turner, K.J.; Chaloupka, J.; Cottier, F.; De Sordi, L.; Sanglard, D.; Levin, L.R.; Buck, J.; Mühlschlegel, F.A. The quorum-sensing molecules farnesol/homoserine lactone and dodecanol operate via distinct modes of action in Candida albicans. Eukaryot. Cell 2011, 10, 1034–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornitzer, D.; Ciechanover, A. Modes of regulation of ubiquitin-mediated protein degradation. J. Cell Physiol. 2000, 182, 1–11. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Genotype | Source |
|---|---|---|
| BWP17 | ura3::imm434/ura3::imm434 his1::hisG/his1::hisG arg4::hisG/arg4::hisG | WY lab |
| BWP17(A+) | ARG4 | WY lab |
| BWP17(U+) | URA3 | WY lab |
| BWP17(U+H+) | URA3, HIS1 | WY lab |
| BWP17(U+A+) | URA3, ARG4 | WY lab |
| BWP17(H+A+) | HIS1, ARG4 | WY lab |
| BWP17(U+H+A+) | HIS1, ARG4, URA3 | WY lab |
| WLX 11 | ubi4Δ::HIS1/ubi4Δ::URA3 | This study |
| WLX 12 | ubi4Δ::HIS1/ubi4Δ::URA3, ARG4 | This study |
| WLX 13 | ubi4Δ::HIS1/ubi4Δ::URA3, UBI4-GFP-ARG4 | This study |
| WLX 19 | sfl1Δ::HIS1/sfl1Δ::URA3 | This study |
| WLX 20 | sfl1Δ::HIS1/sfl1Δ::URA3, ARG4 | This study |
| WLX 21 | sfl1Δ::HIS1/sfl1Δ::URA3, SFL1-GFP-ARG4 | This study |
| WLX 60 | SFL1-HA-ARG4, SFL2-Myc (overexpression)-URA3 | This study |
| WLX 67 | SFL1-HA-ARG4 | This study |
| WLX 68 | sfl1Δ::HIS1/SFL1-Myc-ARG4 | This study |
| WLX 70 | ubi4Δ::HIS1/ubi41Δ::URA3, SFL1-Myc (overexpression)-ARG4 | This study |
| WLX 71 | ubi4Δ::HIS1/ubi41Δ::URA3, SFL2-Myc-ARG4 | This study |
| WLX 72 | SFL2-Myc::ARG4 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Hu, Y.; Yin, Z.; Gao, Q.; Zhang, Y.; Chan, F.Y.; Zeng, G.; Weng, L.; Wang, L.; Wang, Y. Candida albicans Ubiquitin and Heat Shock Factor-Type Transcriptional Factors Are Involved in 2-Dodecenoic Acid-Mediated Inhibition of Hyphal Growth. Microorganisms 2020, 8, 75. https://doi.org/10.3390/microorganisms8010075
Yang D, Hu Y, Yin Z, Gao Q, Zhang Y, Chan FY, Zeng G, Weng L, Wang L, Wang Y. Candida albicans Ubiquitin and Heat Shock Factor-Type Transcriptional Factors Are Involved in 2-Dodecenoic Acid-Mediated Inhibition of Hyphal Growth. Microorganisms. 2020; 8(1):75. https://doi.org/10.3390/microorganisms8010075
Chicago/Turabian StyleYang, Dongliang, Yanling Hu, Zixin Yin, Qianru Gao, Yuqian Zhang, Fong Yee Chan, Guisheng Zeng, Lixing Weng, Lianhui Wang, and Yue Wang. 2020. "Candida albicans Ubiquitin and Heat Shock Factor-Type Transcriptional Factors Are Involved in 2-Dodecenoic Acid-Mediated Inhibition of Hyphal Growth" Microorganisms 8, no. 1: 75. https://doi.org/10.3390/microorganisms8010075