Recurrent Vulvovaginal Candidiasis: An Immunological Perspective


1
Department of Internal Medicine and Radboud Center for Infectious Diseases, Radboud University Medical Center, 6525 GA Nijmegen, The Netherlands
2
Department for Genomics & Immunoregulation, Life and Medical Sciences Institute (LIMES), University of Bonn, 53115 Bonn, Germany
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(2), 144; https://doi.org/10.3390/microorganisms8020144
Submission received: 19 December 2019
/
Revised: 14 January 2020
/
Accepted: 17 January 2020
/
Published: 21 January 2020
(This article belongs to the Special Issue Candida albicans Virulence Factors and Its Pathogenecity)
Abstract
:Vulvovaginal candidiasis (VVC) is a widespread vaginal infection primarily caused by Candida albicans. VVC affects up to 75% of women of childbearing age once in their life, and up to 9% of women in different populations experience more than three episodes per year, which is defined as recurrent vulvovaginal candidiasis (RVVC). RVVC results in diminished quality of life as well as increased associated healthcare costs. For a long time, VVC has been considered the outcome of inadequate host defenses against Candida colonization, as in the case of primary immunodeficiencies associated with persistent fungal infections and insufficient clearance. Intensive research in recent decades has led to a new hypothesis that points toward a local mucosal overreaction of the immune system rather than a defective host response to Candida colonization. This review provides an overview of the current understanding of the host immune response in VVC pathogenesis and suggests that a tightly regulated fungus–host–microbiota interplay might exert a protective role against recurrent Candida infections.
1. Definition and Clinical Perspective
Vulvovaginal candidiasis (VVC) is a debilitating pathological condition caused by Candida species, commonly characterized by vulvar itching, burning, pain while urinating, and vaginal discharge [1]. Candida is a dimorphic fungus from the phyla Ascomycota that inhabits the respiratory, gastrointestinal and genitourinary tracts of more than 30% of healthy individuals during their lifetime [2,3]. Alterations in the symbiotic interplay between the fungus and the mucosal ecosystem are associated with mild to moderate fungal dysbiosis, depending on the infected area as well as the patient’s health status. After anaerobic bacterial vaginosis, VVC is considered the second most common vaginal infection, affecting 75–80% of women at least once in their lifetime [1,4]. Despite the different pathogenesis, symptoms of fungal and bacterial vaginitis are often confused, thus resulting in women having an inaccurate diagnosis and reduced quality of life [5].
Up to 9% of women in various populations experience more than three or four episodes within one year, which is regarded as recurrent vulvovaginal candidiasis (RVVC) [6]. Worldwide prevalence and epidemiological data are rare and inaccurate because they are mostly carried out from self-reports and local general practitioner diagnosis. In this regard, Denning et al. systematically assessed epidemiological studies from 1985 to 2016 and, basing their study on the 6000 online surveys from five Western European countries and the United States by Foxman et al., documented a global annual prevalence of 3871 RVVC cases per 100,000 women, with the highest frequency (9%) in patients aged between 25 and 34 years old [6,7]. According to the Clinical Practice Guidelines, VVC can be treated with topical or oral antifungal formulations, among which azoles (e.g., miconazole, clotrimazole and fluconazole) are the most frequently prescribed therapeutics [8], although they do not prevent recurrent episodes after therapy cessation, necessitating antifungal prophylaxis [9]. RVVC does not correlate with mortality rates but the morbidity is dramatically increasing, and the costs associated with medical care rise accordingly. Hence, more effort needs to be made on the one hand to understand the immunopathogenesis and on the other hand to treat VVC patients efficiently and prevent recurrences.
In this review, we first provide a brief overview of the risk factors associated with increased susceptibility to VVC and then focus on RVVC immunology and pathogenesis. We hypothesize that RVVC might be due to a dysregulated immune system in response to Candida colonization rather than a defective host defense.
2. Risk Factors Associated with RVVC Susceptibility
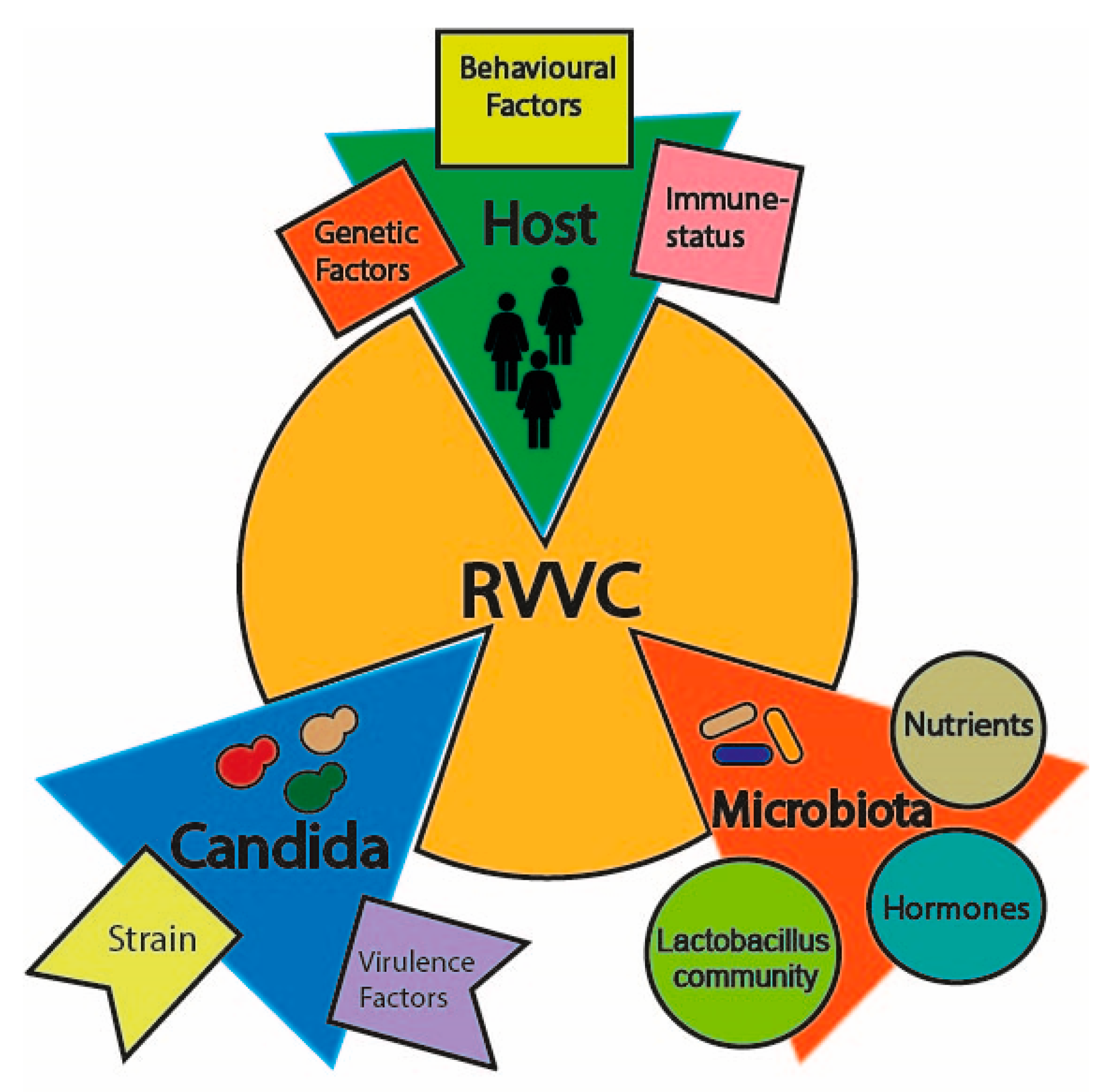
Vulvovaginal candidiasis is considered a multifactorial disorder, where an imbalanced vaginal microbiota composition, host predisposing factors and genetics as well as Candida strains are likely to favor disease onset (Figure 1). The vaginal microbiome is commonly inhabited both by bacterial communities, mainly represented by the genus Lactobacillus, such as L. iners and L. crispatus, and yeast counterparts, commonly called the mycobiome [10,11]. Candida species are the most abundant fungal organisms of the vaginal mycobiome; hence, they can be causative agents of vaginal infections under some conditions [12,13,14]. Lactobacilli species are believed to favor a healthy vaginal microbiome both by acidifying the environment through anaerobic metabolism of glycogen to D-lactic acid and through hydrogen peroxide (H2O2) production, whose antimicrobial activity is likely to inhibit Candida invasion [15,16,17,18]. Several factors can alter the vaginal microbiota in patients with RVVC: firstly, changes in the H2O2-producing Lactobacillus community (e.g., L. acidophilus, L. gasseri and L. vaginalis), and secondly, a high estrogen condition (i.e., estrogen replacement therapy, luteal phase or pregnancy) as well as diversity in carbon sources, short-chain fatty acids or eicosanoids composition (Table 1) [19,20,21,22,23,24,25]. These conditions have been shown to disrupt the balance between tolerance and invasion, leading to Candida adherence to the mucosal epithelium, abnormal yeast growth and increased risk of contracting Candida infections [26,27].
In addition, a broad spectrum of host-related predisposing factors such as type-2 diabetes mellitus, immunosuppression regimens, antibiotics therapy, as well as behavioral factors such as use of contraceptives and intrauterine devices have been suggested to promote the onset of VVC [29,30,31]. However, since around 20–30% of VVC patients are healthy women without predisposing factors, it has also been suggested that inter-individual differences such as genetic background and ethnicity, as well as types of Candida strains and occurrence, might play a key role in idiopathic RVVC pathogenesis.
According to epidemiological data and multi-ethnic cohort studies, increased susceptibility to RVVC rates correlates with genetic polymorphisms as well as ethnicity. For instance, carriage of the single nucleotide polymorphism (SNP) in exon 1 codon 54 in the mannose-binding lectin 2 (MBL2) gene is reported to be more frequent in RVVC patients than healthy women, and this association has also been revealed in Brazilian, European, Chinese and Egyptian, but not Italian women [32,33,34,35,36]. In addition, clinical reports showed that African Americans are more prone to fungal infections in comparison to European or Hispanic women, thus suggesting a variation in frequency among ethnic groups [37,38].
Furthermore, it has emerged that the incidence of Candida infections is also species-related. Distribution and epidemiological studies carried out on cohorts in the United States, Europe and Australia identified C. albicans as the main occurring species, isolated in 75–90% of the positive cultures for VVC [39,40,41]. As far as the non-albicans Candida (NAC) infections are concerned, the highest frequency rates have been reported for C. glabrata—around 10–20% of cases, followed by C. parapsilosis, C. tropicalis, C. krusei and C. africana [42,43,44]. Given that species distribution and incidence vary among countries as well as the population studied, a significant increasing occurrence of NAC species in VVC patients has been revealed [45,46]. In particular, this has been reported in Tunisia, Nigeria, Middle Eastern countries and Asia, where C. glabrata is the most frequently isolated NAC (30–50%) [43,44,47,48,49]. In addition, it is believed that NAC species are more likely to favor recurrent infections in VVC patients [43,50], perhaps because of their refractoriness to the current azole drug treatment [31].
Interestingly, a correlation between genotype distributions of C. albicans and their severity has emerged from cohort studies in China, where high isolation rates of the dominant genotypes of C. albicans were more frequent in severe VVC compared to mild-to-moderate vaginal isolates [51]. Nevertheless, this correlation was not validated in Turkish women, suggesting therefore a different distribution pattern of C. albicans genotype between Chinese and non-Chinese VVC patients [52,53]. According to a microsatellite typing study in women affected by acute VVC, non-albicans species have been found to be potentially more severe than C. albicans species, the latter being mainly associated with a mild or moderate phenotype [54]. Although the aforementioned predisposing causes are believed to explain some cases of VVC worldwide, it remains to be elucidated what contributes to the shift from VVC to RVVC. Therefore, it may be hypothesized that an immunological dysregulation by the host might explain the majority of episodes, although it has yet to be clarified whether it may be due to a deficient or hypersensitive response of the host immune system.
3. Immunology: RVVC as an Immunodeficiency
Our knowledge about the immune response upon invasion of pathogens has advanced considerably during recent decades. In this regard, for more insights, comprehensive reviews have been published in the literature on this subject [55,56]. Briefly, once C. albicans impairs the first line defense, represented by epithelial cells, antimicrobial peptides and signaling molecules are released, thus leading to innate immune cell recruitment. Different fungal cell-wall components such as β-glucans and mannoproteins, commonly referred to as pathogen-associated molecular patterns (PAMPs), are sensed by several families of pattern recognition receptors (PRRs), primarily Toll-like receptors (TLRs) and C-type lectin receptors (CLRs) expressed on myeloid cells. Once recognized, Candida components are processed by phagocytic cells and, based on antigen specificity, T helper (Th) cells are differentiated in cellular subsets, leading ultimately to pathogen clearance [57].
Host genetic variants in PRRs as well as in signal transducers have long been thought to impair antifungal immune response in RVVC patients. For instance, a non-synonymous mutation in TLR2 (rs5743704, Pro631His) was reported to mediate an enhanced RVVC susceptibility [58]. Moreover, several cohort studies in different countries have investigated a SNP in the first exon of the MBL2 gene, which is commonly associated with a defective immune response in various diseases [59]. However, no central role for MBL2 polymorphism as the sole predisposing factor, either in protecting or in favoring RVVC onset, has been demonstrated, thus strengthening the concept that RVVC is a multifactorial disorder [35,60]. For its essential role in β-glucans recognition, Dectin-1 has been largely studied in antifungal immunity: knockdown of the receptor in mouse experiments resulted in higher fungal burden and a defective Th1/Th17 response, highlighting that antifungal mucosal immunity relies on host genetics [61,62]. In addition, it has been reported that a mutation in the human CLEC7A gene is associated with an impaired cytokine response and an increased susceptibility to recurrent mucosal or skin infections, but not oropharyngeal candidiasis (OPC) [63], rather than invasive candidiasis [64,65]. Hence, it has been long believed that VVC onset is the result of SNPs or other genetic alterations in key signaling proteins. Although the data suggest that the homozygous point mutation in Dectin-1 may be crucial for severe mucosal, but not systemic, infections, it is rare and limited to a small number of individuals—around 10–15% of Western women are heterozygous and less than 1% are homozygous for this polymorphism. Whether monoallelic variants that are commonly associated with a decreased immune response to fungal infection are responsible for RVVC pathogenesis remains to be elucidated.
In addition to genetic variations, a compromised antifungal host defense, such as that found in diabetes mellitus, is thought to increase the risk of vaginal fungal infections [66]. Indeed, cross-sectional studies in Brazil confirmed that Candida colonization rates are higher in diabetic women (18.8%) than in non-diabetic patients (11.8%), with the diabetic group showing more symptomatic (VVC + RVVC = 66.6%) than colonized (33.3%) patients [67]. Even though uncontrolled diabetes causes metabolic alterations that can predispose women to symptomatic vaginitis, only a minority of women are affected by RVVC, suggesting individual differences or additional factors may influence susceptibility to Candida infections.
Since a correlation between susceptibility to oral, gastrointestinal and mucocutaneous candidiasis is linked to T-cell deficiency, both human and animal studies have been carried out in recent decades to elucidate whether an impaired cell-mediated immunity (CMI) is also responsible for the lack of protection against VVC [68]. As Th1 and Th17 responses are known to be primarily involved in protection against candidiasis, clinical studies directed their attention toward a systemic immunosuppression state due to an impaired T-cell-mediated immune response. For instance, inherited genetic mutations in the Th17 pathway (e.g., STAT1, STAT3, STAT4) are known to lead to several diseases such as chronic mucocutaneous candidiasis (CMC), Hyper-IgE syndrome (HIES) and autoimmune polyendocrinopathy–candidiasis–ectodermal dystrophy (APECED). The result is an impaired IL-17 immune response, subsequent decreased neutrophil recruitment and hence a higher fungal load [69]. Interestingly, these individuals are highly prone to develop mucosal candidiasis such as recurrent OPC or onychomycosis, but not RVVC, despite some cases of vulva lesions having been described in both a Brazilian and an Iranian family [70,71,72]. Case-controlled studies in patients with acquired disorders of T-cell immunity, such as HIV/AIDS, revealed a distinct protective host defense mechanism against OPC and VVC. In fact, RVVC occurs equally among immunocompetent and immunocompromised women, while OPC is more frequent under immunocompromised states such as in HIV-positive individuals (>90%) [73,74,75]. In addition, experimental estrogen-dependent murine models reported an absence of systemic T-cell infiltration into the vaginal mucosa and normal levels of Th-1/Th-2 cytokines between RVVC patients and controls [76,77,78]. These results are in accordance with a cohort study by Talaei et al., who measured an altered T-lymphocyte proliferation in response to Candida in only 58% of RVVC patients compared to controls [79]. Since clinical observation revealed that mucosal sites are differentially susceptible to Candida infection and mice models of vaginal candidiasis failed to identify a protective role of Th-1 and Th-17 phenotypes, a local dysregulation in CMI rather than a systemic immune deficiency has been considered to be involved in RVVC susceptibility [75]. A murine model of vaginal candidiasis reported higher vaginal tissue expression and production of TGF-β1, thus suggesting its immunoregulatory activity might be responsible for a lack of CMI at the level of the vagina [76,80]. However, animal model studies failed to identify a protective role for resident vaginal T-cells upon Candida infection [76,81,82]. Moreover, evaluation of human Candida-specific antibodies (IgG and IgA) in serum and vaginal secretions both in humans and mice did not provide any significant evidence that humoral immunity (HI) mediates RVVC protection [83,84,85,86].
Overall, although our knowledge has advanced considerably during recent years, the role of cell-mediated immunity remains limited. While some sub-groups of VVC patients, such as those with CLEC7/dectin-1 deficiency, may suffer from RVVC, classical immune deficiencies characterized by defects in antifungal host defense (e.g., HIV-infected patients, patients with CMC) fail to show an increased susceptibility to VVC, which suggests that there are additional pathways for the pathogenesis of the disease.
4. Immunology: RVVC as an Autoinflammatory Disease
Given the unconfirmed protective role of T-cells in modulating the host response to vaginal candidiasis, the idea of a dysregulated innate immune response to Candida colonization has gained a foothold in recent years.
Even though animal models are indispensable for the study of RVVC immunopathogenesis, they are not completely able to reproduce the human condition: in fact, high exogenous estrogen administration, as well as a large Candida inoculum, are required to establish persistent infections in mice in which Candida is absent from the normal microbiota [87]. To shed light on the immune regulation of Candida infection, Fidel and colleagues challenged healthy women intravaginally with live Candida and found that the protection against infection is non-inflammatory; in contrast, symptomatic infection correlates with elevated vaginal polymorphonuclear neutrophil (PMN) infiltration and fungal burden, thus suggesting that symptoms are PMN-mediated [88]. In accordance, high levels of neutrophils have been reported in vaginal lavages of VVC patients [89]. Vaginal fluids from mice inoculated with Candida exhibited high levels of epithelial-derived S100A8 calcium-binding protein (also known as calgranulin A). S100A8 is an endogenous danger signal involved in the amplification of inflammation in autoimmunity as well as in response to infections [90,91]. Although alarmins are potent chemoattractants of PMNs in the vaginal lumen, they are sufficient but dispensable for initiating the acute neutrophil response upon interaction with small numbers of Candida in symptomatic individuals [92]. However, since activated phagocytes fail to reduce the fungal burden and vaginal epithelial cell anti-Candida activity is merely fungistatic [93], other mediators are thought to be involved in PMN dysfunction once recruited in the vaginal mucosa. Interestingly, the proteoglycan heparan sulfate (HS) has been recently indicated as a novel candidate involved in the lack of PMNs’ killing activity, placing them in an anergic state once recruited in the vaginal epithelium [94,95].
Transcriptomic analysis of vaginal tissue from a murine model of VVC demonstrated an increased expression of host genes involved in myeloid cell mobilization, phagocyte infiltration, as well as Th17 cytokine secretion (IL-22, IL-17A and IL-17F). It has also been demonstrated that NLRC4, IFN-γ, Nrh1 and miR-21 mediate NLRP3 inflammasome expression, subsequently leading to interleukin-1β (IL-1β) cleavage and infiltration of PMNs into vaginal tissue, as also reported in vaginal lavage fluid [96,97,98]. Indeed, RVVC patients carrying a polymorphism in the NLRP3 gene had a higher production of IL-1β but low IL-1Ra levels, strengthening the central role of NLRP3 in favoring a persistent hyperinflammatory state in VVC [99]. Alteration of the vaginal microbiota, and diet-induced changes in composition as well as concentration (e.g., impaired tryptophan or short-chain fatty acid levels) have been shown to impact host immune response to Candida colonization by driving pathogenic NLRP3 inflammation in the absence of IL-22. A mouse model of VVC showed that stimulation of the aryl hydrocarbon receptor (AhR) on group 3 innate lymphoid cells (ILC3) by its ligand indole-3-aldehyde (3-IAld) promotes IL-22 expression, which in turn induces NLRC4 phosphorylation and subsequent NLRP3 restrained bioactivity via IL-1Ra production [100,101,102]. Disruption of the crucial IL-22/NLRC4/IL-1Ra axis has been addressed as being responsible for the lack of antifungal resistance, thus suggesting IL-22 deficiency might be a risk factor for RVVC susceptibility [103]. In addition to NLRP3, the inflammasome component NLPR6 has been found to trigger IL-18 production during the early stages of infection. Stimulation of the IL-22/NRLC4 axis by IL-18 production results in IL-18 release during a later phase, subsequently mounting a positive feedback loop mechanism. As deficiency in IL-18/IL-22 crosstalk is associated with impaired Th/Treg cell development and increased VVC susceptibility, IL-18 appears to act as a bridge between innate and adaptive immunity during vaginal candidiasis [104].
A dual role of IL-9 has been reported in gastrointestinal candidiasis [105]. Studies in mice showed that IL-9 is likely to modulate vaginal response to Candida in a time-dependent manner, initially favoring inflammation via NLRP3 activation and then tolerance to the fungus by stimulating IL-1Ra production, mast cell (MC) engagement and macrophage crawling [106,107]. Interestingly, as the IL-9/MC axis contributes to chronic allergic inflammation [108] and is associated in a sex-specific manner with IgE levels in cystic fibrosis [109], it may lead to the hypothesis that VVC is an allergic reaction to the ability of Candida-specific IgE and prostaglandin E2 (PGE2) to inhibit vaginal cell-mediated immune response [110]. These findings are consistent with a positive correlation between sensitization to atopic reactions and non-responders to RVVC maintenance treatment [111]. However, this phenomenon has been observed only in a minority of women, and further investigations are needed [112].
In accordance with transcriptomic results from the mouse model, a comprehensive genome-wide study in RVVC patients found that cell morphogenesis, adhesion and cellular metabolism pathways are dysregulated in RVVC patients. Furthermore, the combination with immunological data revealed that T-cell cytokines IL-17, IL-22 and IFN-γ were higher in individuals bearing a variant in the SIGLEC15 (sialic acid-binding immunoglobulin-like lectin 15) gene, resulting then in a hyperinflammatory state in response to Candida. Given the fact that sialic structures are also expressed on the surface of Candida, SIGLEC15 has been suggested as a novel susceptibility factor in RVVC [113]. Even though IL-23p19−/−, IL-17RA−/− and IL-22−/− mice intravaginally inoculated with Candida suggested little-to-no evidence of Th17 cytokines being primary effectors during acute inflammation [114], the role of IL-22 has been long investigated recently for its ability to provide antifungal resistance during infection at the mucosal sites by preventing epithelial cell damage [115]. Interestingly, a mouse model of VVC revealed that the role of the IL-17/IL-22 axis is independent from estrogen administration [116].
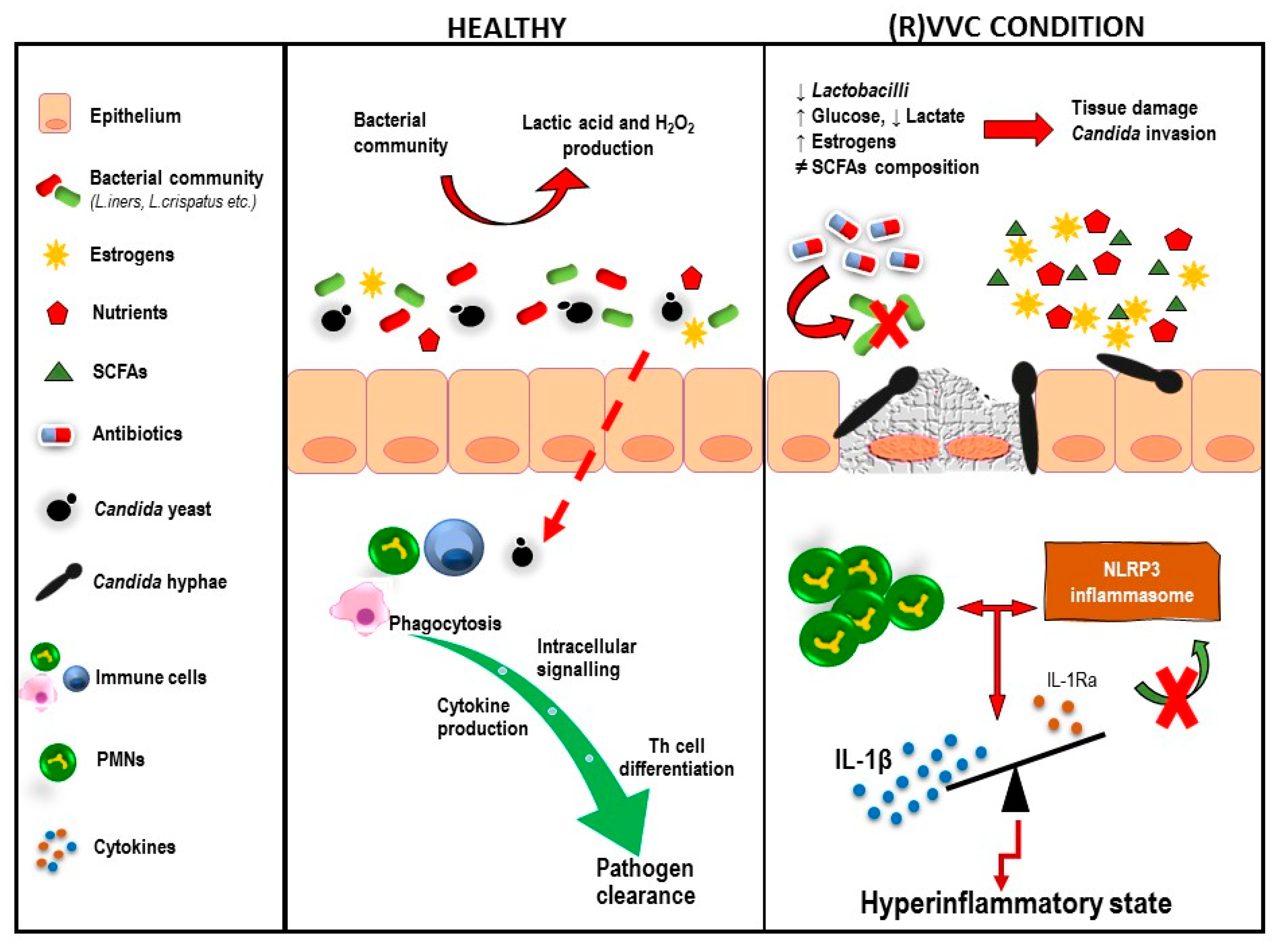
Overall, on the basis of mouse and human studies, a PMN-mediated immunopathology in symptomatic VVC/RVVC has been proposed: under certain vaginal environmental conditions, Candida switches from the commensal to opportunistic state by expressing a broad range of hypha-associated morphogenetic and virulence factors (nicely reviewed in [117])—mostly secreted aspartyl proteases (SAPs) and candidalysin. Inflammatory mediators such as alarmins and IL-1β produced by vaginal epithelial cells lead to PMN chemotaxis and recruitment to the vaginal mucosa. This event, in turn, induces elevated infiltration levels of the same mediators, establishing a positive feedback mechanism loop that results in a hyperinflammatory state [95]. Notably, the same pattern of inflammatory mediators has been reported in VVC patients, with little-to-no expression in asymptomatic colonized carriers and healthy women, providing evidence that NLRP3 is a hallmark of symptomatic immunopathology in humans [118]. Hence, VVC/RVVC might result from an overly aggressive innate reaction by PMNs, rather than an impaired adaptive immune response (Figure 2) [88,119]. Further studies are needed to elucidate the mechanisms that contribute to (R)VVC pathogenesis.
5. Perspectives and Future Directions
To date, data both from murine and human studies suggest that the immunological profile differs among RVVC patient subgroups: whereas deficiencies in the immune response to fungal infections characterize a small number of patients, the majority display a hyperinflammatory response towards Candida colonization and invasion, which might be due to the host’s genetic background. Of note, on the contrary to what one would expect if RVVC were an immunodeficiency, an acute response has been revealed in patients infected by very low fungal loads [120]. For these reasons, Candida burden alone cannot be considered predictive of disease. In this regard, a combination of genetic studies in RVVC patients and carefully matched controls combined with immunological data is needed to elucidate the role of the innate/adaptive immune response and decipher the factors involved in vaginal mucosal defense mechanisms upon Candida infection.
Even though antifungal agents are effective in patients, they provide static effects and do not prevent recurrences in roughly half of patients. Clarification of resistance mechanisms could improve the development of long-lasting and efficient drugs that target the inflammatory reaction. Adjuvant immunotherapy approaches have been developed over the years: humanized neutralizing antibodies against IL-9 [106], IL-1Ra [103] or molecular agents able to promote protection signaling to Candida infection, such as AhR agonists [104]. They could represent an option to ameliorate inflammation and alleviate symptoms in acute VVC patients, but future studies to demonstrate their efficacy are needed. Alongside immunotherapies, vaccination is a possible approach to boost the immune system and provide long-lasting memory of Candida. As an example, recombinant vaccines such as NVD-3, based on the virulence factor Alsp3 (agglutinin-like sequence proteins), and PEV-7 based on Sap2p [121,122] have progressed in the first trial steps. However, no vaccine is currently available in the clinical setting [123]. In addition, it has been proposed to use exogenous Lactobacilli administration—such as probiotics—as an adjuvant therapy, which might restore the microenvironmental balance and prevent VVC recurrences, since an abnormal vaginal microbiota is considered to favor reproductive tract infections. A recent study of vaginal microbiome transplantation in bacterial vaginosis may provide a template for such future studies [124]. Only a comprehensive approach to investigate all these potential treatments will be able to improve the outcome of this disease with high morbidity among women.
Author Contributions
Conceptualization, M.G.N. and M.J.; Methodology, D.R.; M.B.; M.J.; J.t.O.; M.G.N.; Writing—Original Draft Preparation, D.R. and M.B.; Writing—Review & Editing, D.R.; M.B.; M.J.; J.t.O.; M.G.N.; Visualization D.R.; Supervision, M.B., M.J. and M.G.N.; Project Administration, M.G.N.; Funding Acquisition, M.G.N. All authors have read and agreed to the published version of the manuscript.
Funding
D.R. was supported by the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement [No 812969].M.G.N. was supported by an ERC Advanced Grant [No 833247] and a Spinoza Grant from the Netherlands Organization for Scientific Research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anderson, M.R.; Klink, K.; Cohrssen, A. Evaluation of vaginal complaints. JAMA 2004, 291, 1368–1379. [Google Scholar] [CrossRef]
- Netea, M.G.; Marodi, L. Innate immune mechanisms for recognition and uptake of Candida species. Trends Immunol. 2010, 31, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Diezmann, S.; Cox, C.J.; Schonian, G.; Vilgalys, R.J.; Mitchell, T.G. Phylogeny and evolution of medical species of Candida and related taxa: A multigenic analysis. J. Clin. Microbiol. 2004, 42, 5624–5635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, A.O.; Heidrich, F.E.; Fihn, S.D.; Bergman, J.J.; Wood, R.W.; Stamm, W.E.; Holmes, K.K. Establishing the cause of genitourinary symptoms in women in a family practice. Comparison of clinical examination and comprehensive microbiology. JAMA 1984, 251, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D. Epidemiology and pathogenesis of recurrent vulvovaginal candidiasis. Am. J. Obstet. Gynecol. 1985, 152, 924–935. [Google Scholar] [CrossRef]
- Foxman, B.; Muraglia, R.; Dietz, J.P.; Sobel, J.D.; Wagner, J. Prevalence of recurrent vulvovaginal candidiasis in 5 European countries and the United States: Results from an internet panel survey. J. Low Genit. Tract Dis. 2013, 17, 340–345. [Google Scholar] [CrossRef]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, e1–e50. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D.; Wiesenfeld, H.C.; Martens, M.; Danna, P.; Hooton, T.M.; Rompalo, A.; Sperling, M.; Livengood, C., 3rd; Horowitz, B.; Von Thron, J.; et al. Maintenance fluconazole therapy for recurrent vulvovaginal candidiasis. N. Engl. J. Med. 2004, 351, 876–883. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef] [Green Version]
- Drell, T.; Lillsaar, T.; Tummeleht, L.; Simm, J.; Aaspollu, A.; Vain, E.; Saarma, I.; Salumets, A.; Donders, G.G.; Metsis, M. Characterization of the vaginal micro- and mycobiome in asymptomatic reproductive-age Estonian women. PLoS ONE 2013, 8, e54379. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Bent, S.J.; Schneider, M.G.; Davis, C.C.; Islam, M.R.; Forney, L.J. Characterization of vaginal microbial communities in adult healthy women using cultivation-independent methods. Microbiology 2004, 150, 2565–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wijgert, J.H.; Borgdorff, H.; Verhelst, R.; Crucitti, T.; Francis, S.; Verstraelen, H.; Jespers, V. The vaginal microbiota: What have we learned after a decade of molecular characterization? PLoS ONE 2014, 9, e105998. [Google Scholar] [CrossRef] [Green Version]
- Bradford, L.L.; Ravel, J. The vaginal mycobiome: A contemporary perspective on fungi in women’s health and diseases. Virulence 2017, 8, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Boskey, E.R.; Cone, R.A.; Whaley, K.J.; Moench, T.R. Origins of vaginal acidity: High D/L lactate ratio is consistent with bacteria being the primary source. Hum. Reprod. 2001, 16, 1809–1813. [Google Scholar] [CrossRef]
- Cadieux, P.A.; Burton, J.; Devillard, E.; Reid, G. Lactobacillus by-products inhibit the growth and virulence of uropathogenic Escherichia coli. J. Physiol. Pharmacol. 2009, 60, 13–18. [Google Scholar] [PubMed]
- Parolin, C.; Marangoni, A.; Laghi, L.; Foschi, C.; Nahui Palomino, R.A.; Calonghi, N.; Cevenini, R.; Vitali, B. Isolation of Vaginal Lactobacilli and Characterization of Anti-Candida Activity. PLoS ONE 2015, 10, e0131220. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.J.; Lee, K.; Kwon, B.; You, H.J.; Ko, G. Vaginal lactobacilli inhibit growth and hyphae formation of Candida albicans. Sci. Rep. 2019, 9, 8121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceccarani, C.; Foschi, C.; Parolin, C.; D’Antuono, A.; Gaspari, V.; Consolandi, C.; Laghi, L.; Camboni, T.; Vitali, B.; Severgnini, M.; et al. Diversity of vaginal microbiome and metabolome during genital infections. Sci. Rep. 2019, 9, 14095. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Essmann, M.; Burt, E.T.; Larsen, B. Estrogen effects on Candida albicans: A potential virulence-regulating mechanism. J. Infect. Dis. 2000, 181, 1441–1446. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, A.; Pedro, N.A.; Salazar, S.B.; Mira, N.P. Effect of Acetic Acid and Lactic Acid at Low pH in Growth and Azole Resistance of Candida albicans and Candida glabrata. Front. Microbiol. 2018, 9, 3265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinan, J.; Wang, S.; Hazbun, T.R.; Yadav, H.; Thangamani, S. Antibiotic-induced decreases in the levels of microbial-derived short-chain fatty acids correlate with increased gastrointestinal colonization of Candida albicans. Sci. Rep. 2019, 9, 8872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noverr, M.C.; Huffnagle, G.B. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect. Immun. 2004, 72, 6206–6210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maidan, M.M.; Thevelein, J.M.; Van Dijck, P. Carbon source induced yeast-to-hypha transition in Candida albicans is dependent on the presence of amino acids and on the G-protein-coupled receptor Gpr1. Biochem. Soc. Trans. 2005, 33, 291–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ende, M.; Wijnants, S.; Van Dijck, P. Sugar Sensing and Signaling in Candida albicans and Candida glabrata. Front. Microbiol. 2019, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Vitali, B.; Pugliese, C.; Biagi, E.; Candela, M.; Turroni, S.; Bellen, G.; Donders, G.G.; Brigidi, P. Dynamics of vaginal bacterial communities in women developing bacterial vaginosis, candidiasis, or no infection, analyzed by PCR-denaturing gradient gel electrophoresis and real-time PCR. Appl. Environ. Microbiol. 2007, 73, 5731–5741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarry, W.; Fisher, M.; Shen, S.; Mawhinney, M. Candida albicans: The estrogen target for vaginal colonization. J. Surg. Res. 2005, 129, 278–282. [Google Scholar] [CrossRef]
- Konno, N.; Ishii, M.; Nagai, A.; Watanabe, T.; Ogasawara, A.; Mikami, T.; Matsumoto, T. Mechanism of Candida albicans transformation in response to changes of pH. Biol. Pharm. Bull. 2006, 29, 923–926. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D. Vulvovaginal candidosis. Lancet 2007, 369, 1961–1971. [Google Scholar] [CrossRef]
- Veer, C.V.D.; Bruisten, S.; Houdt, R.V.; Rutten, J.; Matser, A.; Wijgert, J.V.D.; Vries, H.D.; Helm, J.V.D. O04.2 Effects of over-the-counter lactic acid-containing vaginal douching products on the vaginal microbiota. Sex. Transm. Infect. 2017, 93, A8. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammad, N.M.; El Badawy, N.E.; Nasr, A.M.; Ghramh, H.A.; Al Kady, L.M. Mannose-Binding Lectin Gene Polymorphism and Its Association with Susceptibility to Recurrent Vulvovaginal Candidiasis. Biomed. Res. Int. 2018, 2018, 7648152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babula, O.; Lazdane, G.; Kroica, J.; Ledger, W.J.; Witkin, S.S. Relation between recurrent vulvovaginal candidiasis, vaginal concentrations of mannose-binding lectin, and a mannose-binding lectin gene polymorphism in Latvian women. Clin. Infect. Dis. 2003, 37, 733–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Liao, Q.; Liu, Z. Mannose-binding lectin and vulvovaginal candidiasis. Int. J. Gynaecol. Obstet. 2006, 92, 43–47. [Google Scholar] [CrossRef]
- Giraldo, P.C.; Babula, O.; Goncalves, A.K.; Linhares, I.M.; Amaral, R.L.; Ledger, W.J.; Witkin, S.S. Mannose-binding lectin gene polymorphism, vulvovaginal candidiasis, and bacterial vaginosis. Obstet. Gynecol. 2007, 109, 1123–1128. [Google Scholar] [CrossRef]
- Donders, G.G.; Babula, O.; Bellen, G.; Linhares, I.M.; Witkin, S.S. Mannose-binding lectin gene polymorphism and resistance to therapy in women with recurrent vulvovaginal candidiasis. BJOG 2008, 115, 1225–1231. [Google Scholar] [CrossRef]
- Jaffe, S.; Normand, N.; Jayaram, A.; Orfanelli, T.; Doulaveris, G.; Passos, M.; Kanninen, T.T.; Bongiovanni, A.M.; Linhares, I.M.; Witkin, S.S. Unique variation in genetic selection among Black North American women and its potential influence on pregnancy outcome. Med. Hypotheses 2013, 81, 919–922. [Google Scholar] [CrossRef]
- Foxman, B.; Barlow, R.; D’Arcy, H.; Gillespie, B.; Sobel, J.D. Candida vaginitis: Self-reported incidence and associated costs. Sex. Transm. Dis. 2000, 27, 230–235. [Google Scholar] [CrossRef]
- Vermitsky, J.P.; Self, M.J.; Chadwick, S.G.; Trama, J.P.; Adelson, M.E.; Mordechai, E.; Gygax, S.E. Survey of vaginal-flora Candida species isolates from women of different age groups by use of species-specific PCR detection. J. Clin. Microbiol. 2008, 46, 1501–1503. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.; Young, M.L.; Lee, O.; C-A Chen, S. Vulvovaginal carriage of yeasts other than Candida albicans. Sex. Transm. Infect. 2003, 79, 249–250. [Google Scholar] [CrossRef] [Green Version]
- Corsello, S.; Spinillo, A.; Osnengo, G.; Penna, C.; Guaschino, S.; Beltrame, A.; Blasi, N.; Festa, A. An epidemiological survey of vulvovaginal candidiasis in Italy. Eur J. Obstet. Gynecol. Reprod. Biol. 2003, 110, 66–72. [Google Scholar] [CrossRef]
- Linhares, L.M.; Witkin, S.S.; Miranda, S.D.; Fonseca, A.M.; Pinotti, J.A.; Ledger, W.J. Differentiation between women with vulvovaginal symptoms who are positive or negative for Candida species by culture. Infect. Dis. Obstet. Gynecol. 2001, 9, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Cetin, M.; Ocak, S.; Gungoren, A.; Hakverdi, A.U. Distribution of Candida species in women with vulvovaginal symptoms and their association with different ages and contraceptive methods. Scand. J. Infect. Dis. 2007, 39, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Okungbowa, F.I.; Isikhuemhen, O.S.; Dede, A.P. The distribution frequency of Candida species in the genitourinary tract among symptomatic individuals in Nigerian cities. Rev. Iberoam. Micol. 2003, 20, 60–63. [Google Scholar] [PubMed]
- Spinillo, A.; Capuzzo, E.; Gulminetti, R.; Marone, P.; Colonna, L.; Piazzi, G. Prevalence of and risk factors for fungal vaginitis caused by non-albicans species. Am. J. Obstet. Gynecol. 1997, 176, 138–141. [Google Scholar] [CrossRef]
- Achkar, J.M.; Fries, B.C. Candida Infections of the Genitourinary Tract. Clin. Microbiol. Rev. 2010, 23, 253–273. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J.D. Recurrent vulvovaginal candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef]
- Mohanty, S.; Xess, I.; Hasan, F.; Kapil, A.; Mittal, S.; Tolosa, J.E. Prevalence & susceptibility to fluconazole of Candida species causing vulvovaginitis. Indian J. Med. Res. 2007, 126, 216–219. [Google Scholar]
- Amouri, I.; Sellami, H.; Borji, N.; Abbes, S.; Sellami, A.; Cheikhrouhou, F.; Maazoun, L.; Khaled, S.; Khrouf, S.; Boujelben, Y.; et al. Epidemiological survey of vulvovaginal candidosis in Sfax, Tunisia. Mycoses 2011, 54, e499–e505. [Google Scholar] [CrossRef]
- De Vos, M.M.; Cuenca-Estrella, M.; Boekhout, T.; Theelen, B.; Matthijs, N.; Bauters, T.; Nailis, H.; Dhont, M.A.; Rodriguez-Tudela, J.L.; Nelis, H.J. Vulvovaginal candidiasis in a Flemish patient population. Clin. Microbiol. Infect. 2005, 11, 1005–1011. [Google Scholar] [CrossRef]
- Fan, S.R.; Bai, F.Y.; Liao, Q.P.; Liu, Z.H.; Li, J.; Liu, X.P. Genotype distribution of Candida albicans strains associated with different conditions of vulvovaginal candidiasis, as revealed by microsatellite typing. Sex. Transm. Infect. 2008, 84, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, S.H.; Xie, J.; Xu, J.; Li, J.; Li, D.M.; Zong, L.L.; Zheng, Y.C.; Bai, F.Y. Prevalence of specific and phylogenetically closely related genotypes in the population of Candida albicans associated with genital candidiasis in China. Fungal Genet. Biol. 2012, 49, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Guzel, A.B.; Dogen, A.; Aydin, M.; Serin, A.; Serin, M.S.; Kalkanci, A.; Ilkit, M. Genotyping reveals no link between Candida albicans genotype and vaginitis severity in Turkish women. Mycopathologia 2013, 175, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zong, L.L.; Mao, T.; Huang, Y.X.; Xu, Z.M. Distribution of Candida albican genotype and Candida species is associated with the severity of vulvovagianl candidiasis. Nan Fang Yi Ke Da Xue Xue Bao 2011, 31, 1649–1653. [Google Scholar]
- Netea, M.G.; Joosten, L.A.; van der Meer, J.W.; Kullberg, B.J.; van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Richardson, J.P.; Moyes, D.L.; Ho, J.; Naglik, J.R. Candida innate immunity at the mucosa. Semin. Cell Dev. Biol. 2019, 89, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Hofs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: Virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef]
- Rosentul, D.C.; Delsing, C.E.; Jaeger, M.; Plantinga, T.S.; Oosting, M.; Costantini, I.; Venselaar, H.; Joosten, L.A.; van der Meer, J.W.; Dupont, B.; et al. Gene polymorphisms in pattern recognition receptors and susceptibility to idiopathic recurrent vulvovaginal candidiasis. Front. Microbiol. 2014, 5, 483. [Google Scholar] [CrossRef]
- Eisen, D.P.; Minchinton, R.M. Impact of mannose-binding lectin on susceptibility to infectious diseases. Clin. Infect. Dis. 2003, 37, 1496–1505. [Google Scholar] [CrossRef] [Green Version]
- Milanese, M.; Segat, L.; De Seta, F.; Pirulli, D.; Fabris, A.; Morgutti, M.; Crovella, S. MBL2 genetic screening in patients with recurrent vaginal infections. Am. J. Reprod. Immunol. 2008, 59, 146–151. [Google Scholar] [CrossRef]
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for beta-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Giovannini, G.; De Luca, A.; D’Angelo, C.; Casagrande, A.; Iannitti, R.G.; Ricci, G.; Cunha, C.; Romani, L. Dectin-1 isoforms contribute to distinct Th1/Th17 cell activation in mucosal candidiasis. Cell Mol. Immunol. 2012, 9, 276–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobel, J.D. Pathogenesis and treatment of recurrent vulvovaginal candidiasis. Clin. Infect. Dis. 1992, 14, S148–S153. [Google Scholar] [CrossRef] [PubMed]
- Ferwerda, B.; Ferwerda, G.; Plantinga, T.S.; Willment, J.A.; van Spriel, A.B.; Venselaar, H.; Elbers, C.C.; Johnson, M.D.; Cambi, A.; Huysamen, C.; et al. Human dectin-1 deficiency and mucocutaneous fungal infections. N. Engl. J. Med. 2009, 361, 1760–1767. [Google Scholar] [CrossRef] [Green Version]
- Glocker, E.O.; Hennigs, A.; Nabavi, M.; Schaffer, A.A.; Woellner, C.; Salzer, U.; Pfeifer, D.; Veelken, H.; Warnatz, K.; Tahami, F.; et al. A homozygous CARD9 mutation in a family with susceptibility to fungal infections. N. Engl. J. Med. 2009, 361, 1727–1735. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.F.; Rodrigues, M.E.; Henriques, M. Candida sp. Infections in Patients with Diabetes Mellitus. J. Clin. Med. 2019, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Gunther, L.S.; Martins, H.P.; Gimenes, F.; Abreu, A.L.; Consolaro, M.E.; Svidzinski, T.I. Prevalence of Candida albicans and non-albicans isolates from vaginal secretions: Comparative evaluation of colonization, vaginal candidiasis and recurrent vaginal candidiasis in diabetic and non-diabetic women. Sao Paulo Med. J. 2014, 132, 116–120. [Google Scholar] [CrossRef]
- Peters, B.M.; Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Candida vaginitis: When opportunism knocks, the host responds. PLoS Pathog. 2014, 10, e1003965. [Google Scholar] [CrossRef] [Green Version]
- Gaffen, S.L.; Hernandez-Santos, N.; Peterson, A.C. IL-17 signaling in host defense against Candida albicans. Immunol. Res. 2011, 50, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Van de Veerdonk, F.L.; Plantinga, T.S.; Hoischen, A.; Smeekens, S.P.; Joosten, L.A.; Gilissen, C.; Arts, P.; Rosentul, D.C.; Carmichael, A.J.; Smits-van der Graaf, C.A.; et al. STAT1 mutations in autosomal dominant chronic mucocutaneous candidiasis. N. Engl. J. Med. 2011, 365, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Eyerich, K.; Foerster, S.; Rombold, S.; Seidl, H.P.; Behrendt, H.; Hofmann, H.; Ring, J.; Traidl-Hoffmann, C. Patients with chronic mucocutaneous candidiasis exhibit reduced production of Th17-associated cytokines IL-17 and IL-22. J. Investig. Dermatol. 2008, 128, 2640–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambo Filho, A.; Souza Filho, J.B.; Pignaton, C.C.; Zon, I.; Fernandes, A.S.; Cardoso, L.Q. Chronic mucocutaneous candidiasis: A case with exuberant cutaneous horns in nipples. An. Bras. Dermatol. 2014, 89, 641–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, J.E.; Barousse, M.; Swoboda, R.K.; Myers, T.; Hager, S.; Wolf, N.A.; Cutright, J.L.; Thompson, J.; Sobel, J.D.; Fidel, P.L., Jr. Candida-specific systemic cell-mediated immune reactivities in human immunodeficiency virus-positive persons with mucosal candidiasis. J. Infect. Dis. 2001, 183, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.H. Is vulvovaginal candidiasis an AIDS-related illness? Clin. Infect. Dis. 1996, 22, S124–S127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidel, P.L., Jr. History and update on host defense against vaginal candidiasis. Am. J. Reprod. Immunol. 2007, 57, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.N.; Saavedra, M.; Fidel, P.L., Jr. Local Th1/Th2 cytokine production during experimental vaginal candidiasis: Potential importance of transforming growth factor-beta. Med. Mycol. 2000, 38, 419–431. [Google Scholar] [CrossRef]
- Black, C.A.; Eyers, F.M.; Russell, A.; Dunkley, M.L.; Clancy, R.L.; Beagley, K.W. Increased severity of Candida vaginitis in BALB/c nu/nu mice versus the parent strain is not abrogated by adoptive transfer of T cell enriched lymphocytes. J. Reprod. Immunol. 1999, 45, 1–18. [Google Scholar] [CrossRef]
- Richardson, J.P.; Moyes, D.L. Adaptive immune responses to Candida albicans infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Talaei, Z.; Sheikhbahaei, S.; Ostadi, V.; Ganjalikhani Hakemi, M.; Meidani, M.; Naghshineh, E.; Yaran, M.; Emami Naeini, A.; Sherkat, R. Recurrent Vulvovaginal Candidiasis: Could It Be Related to Cell-Mediated Immunity Defect in Response to Candida Antigen? Int. J. Fertil. Steril. 2017, 11, 134–141. [Google Scholar] [CrossRef]
- Ouyang, W.; Chen, S.; Liu, Z.; Wu, Y.; Li, J. Local Th1/Th2 cytokine expression in experimental murine vaginal candidiasis. J. Huazhong Univ. Sci. Technol. Med. Sci 2008, 28, 352–355. [Google Scholar] [CrossRef]
- Fidel, P.L., Jr.; Luo, W.; Steele, C.; Chabain, J.; Baker, M.; Wormley, F., Jr. Analysis of vaginal cell populations during experimental vaginal candidiasis. Infect. Immun. 1999, 67, 3135–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saavedra, M.; Taylor, B.; Lukacs, N.; Fidel, P.L., Jr. Local production of chemokines during experimental vaginal candidiasis. Infect. Immun. 1999, 67, 5820–5826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, C.H.; Rich, R.R.; Bennett, J.E. Chronic mucocutaneous candidiasis: Model-building in cellular immunity. Ann. Intern. Med. 1971, 74, 955–978. [Google Scholar] [CrossRef] [PubMed]
- Fidel, P.L., Jr.; Ginsburg, K.A.; Cutright, J.L.; Wolf, N.A.; Leaman, D.; Dunlap, K.; Sobel, J.D. Vaginal-associated immunity in women with recurrent vulvovaginal candidiasis: Evidence for vaginal Th1-type responses following intravaginal challenge with Candida antigen. J. Infect. Dis. 1997, 176, 728–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, S.; Goust, J.M.; Horger, E.O., 3rd; Fudenberg, H.H. Immunoglobulin E anti-Candida antibodies and candidiasis. Infect. Immun. 1977, 18, 257–259. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, K.L.; Wormley, F.L., Jr.; Fidel, P.L., Jr. Candida-specific antibodies during experimental vaginal candidiasis in mice. Infect. Immun. 2002, 70, 5790–5799. [Google Scholar] [CrossRef] [Green Version]
- Cassone, A.; Sobel, J.D. Experimental Models of Vaginal Candidiasis and Their Relevance to Human Candidiasis. Infect. Immun. 2016, 84, 1255–1261. [Google Scholar] [CrossRef] [Green Version]
- Fidel, P.L., Jr.; Barousse, M.; Espinosa, T.; Ficarra, M.; Sturtevant, J.; Martin, D.H.; Quayle, A.J.; Dunlap, K. An intravaginal live Candida challenge in humans leads to new hypotheses for the immunopathogenesis of vulvovaginal candidiasis. Infect. Immun. 2004, 72, 2939–2946. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, P.C.; de Carvalho, J.B.; do Amaral, R.L.; da Silveira Goncalves, A.K.; Eleuterio, J., Jr.; Guimaraes, F. Identification of immune cells by flow cytometry in vaginal lavages from women with vulvovaginitis and normal microflora. Am. J. Reprod. Immunol. 2012, 67, 198–205. [Google Scholar] [CrossRef]
- Yano, J.; Lilly, E.; Barousse, M.; Fidel, P.L., Jr. Epithelial cell-derived S100 calcium-binding proteins as key mediators in the hallmark acute neutrophil response during Candida vaginitis. Infect. Immun. 2010, 78, 5126–5137. [Google Scholar] [CrossRef] [Green Version]
- Ehrchen, J.M.; Sunderkotter, C.; Foell, D.; Vogl, T.; Roth, J. The endogenous Toll-like receptor 4 agonist S100A8/S100A9 (calprotectin) as innate amplifier of infection, autoimmunity, and cancer. J. Leukoc. Biol. 2009, 86, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Palmer, G.E.; Eberle, K.E.; Peters, B.M.; Vogl, T.; McKenzie, A.N.; Fidel, P.L., Jr. Vaginal epithelial cell-derived S100 alarmins induced by Candida albicans via pattern recognition receptor interactions are sufficient but not necessary for the acute neutrophil response during experimental vaginal candidiasis. Infect. Immun. 2014, 82, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomanbhoy, F.; Steele, C.; Yano, J.; Fidel, P.L., Jr. Vaginal and oral epithelial cell anti-Candida activity. Infect. Immun. 2002, 70, 7081–7088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Vaginal Heparan Sulfate Linked to Neutrophil Dysfunction in the Acute Inflammatory Response Associated with Experimental Vulvovaginal Candidiasis. MBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Yano, J.; Peters, B.M.; Noverr, M.C.; Fidel, P.L., Jr. Novel Mechanism behind the Immunopathogenesis of Vulvovaginal Candidiasis: “Neutrophil Anergy”. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Bruno, V.M.; Shetty, A.C.; Yano, J.; Fidel, P.L., Jr.; Noverr, M.C.; Peters, B.M. Transcriptomic analysis of vulvovaginal candidiasis identifies a role for the NLRP3 inflammasome. MBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Palmer, G.E.; Nash, A.K.; Lilly, E.A.; Fidel, P.L., Jr.; Noverr, M.C. Fungal morphogenetic pathways are required for the hallmark inflammatory response during Candida albicans vaginitis. Infect. Immun. 2014, 82, 532–543. [Google Scholar] [CrossRef] [Green Version]
- Yano, J.; Noverr, M.C.; Fidel, P.L., Jr. Cytokines in the host response to Candida vaginitis: Identifying a role for non-classical immune mediators, S100 alarmins. Cytokine 2012, 58, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, M.; Carvalho, A.; Cunha, C.; Plantinga, T.S.; van de Veerdonk, F.; Puccetti, M.; Galosi, C.; Joosten, L.A.; Dupont, B.; Kullberg, B.J.; et al. Association of a variable number tandem repeat in the NLRP3 gene in women with susceptibility to RVVC. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- De Luca, A.; Carvalho, A.; Cunha, C.; Iannitti, R.G.; Pitzurra, L.; Giovannini, G.; Mencacci, A.; Bartolommei, L.; Moretti, S.; Massi-Benedetti, C.; et al. IL-22 and IDO1 affect immunity and tolerance to murine and human vaginal candidiasis. PLoS Pathog. 2013, 9, e1003486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szilagyi, M.; Laky, G.; Suri, A.; Guba, F. Activities of some serum enzymes in halothane reacted and non-reacted pigs. Acta Vet. Hung. 1989, 37, 117–121. [Google Scholar] [PubMed]
- Borghi, M.; De Luca, A.; Puccetti, M.; Jaeger, M.; Mencacci, A.; Oikonomou, V.; Pariano, M.; Garlanda, C.; Moretti, S.; Bartoli, A.; et al. Pathogenic NLRP3 Inflammasome Activity during Candida Infection Is Negatively Regulated by IL-22 via Activation of NLRC4 and IL-1Ra. Cell Host Microbe 2015, 18, 198–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, M.; Pariano, M.; Solito, V.; Puccetti, M.; Bellet, M.M.; Stincardini, C.; Renga, G.; Vacca, C.; Sellitto, F.; Mosci, P.; et al. Targeting the Aryl Hydrocarbon Receptor With Indole-3-Aldehyde Protects From Vulvovaginal Candidiasis via the IL-22-IL-18 Cross-Talk. Front. Immunol. 2019, 10, 2364. [Google Scholar] [CrossRef] [PubMed]
- Renga, G.; Moretti, S.; Oikonomou, V.; Borghi, M.; Zelante, T.; Paolicelli, G.; Costantini, C.; De Zuani, M.; Villella, V.R.; Raia, V.; et al. IL-9 and Mast Cells Are Key Players of Candida albicans Commensalism and Pathogenesis in the Gut. Cell Rep. 2018, 23, 1767–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renga, G.; Borghi, M.; Oikonomou, V.; Mosci, P.; Bartoli, A.; Renauld, J.C.; Romani, L.; Costantini, C. IL-9 Integrates the Host-Candida Cross-Talk in Vulvovaginal Candidiasis to Balance Inflammation and Tolerance. Front. Immunol. 2018, 9, 2702. [Google Scholar] [CrossRef] [Green Version]
- De Zuani, M.; Paolicelli, G.; Zelante, T.; Renga, G.; Romani, L.; Arzese, A.; Pucillo, C.E.M.; Frossi, B. Mast Cells Respond to Candida albicans Infections and Modulate Macrophages Phagocytosis of the Fungus. Front. Immunol. 2018, 9, 2829. [Google Scholar] [CrossRef]
- Kearley, J.; Erjefalt, J.S.; Andersson, C.; Benjamin, E.; Jones, C.P.; Robichaud, A.; Pegorier, S.; Brewah, Y.; Burwell, T.J.; Bjermer, L.; et al. IL-9 governs allergen-induced mast cell numbers in the lung and chronic remodeling of the airways. Am. J. Respir. Crit. Care Med. 2011, 183, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Renga, G.; Oikonomou, V.; Galosi, C.; Pariano, M.; Iannitti, R.G.; Borghi, M.; Puccetti, M.; De Zuani, M.; Pucillo, C.E.; et al. A mast cell-ILC2-Th9 pathway promotes lung inflammation in cystic fibrosis. Nat. Commun. 2017, 8, 14017. [Google Scholar] [CrossRef]
- Bernstein, J.A.; Seidu, L. Chronic vulvovaginal Candida hypersensitivity: An underrecognized and undertreated disorder by allergists. Allergy Rhinol. 2015, 6, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Donders, G.G.G.; Grinceviciene, S.; Bellen, G.; Jaeger, M.; Ten Oever, J.; Netea, M.G. Is non-response to fluconazole maintenance therapy for recurrent Candida vaginitis related to sensitization to atopic reactions? Am. J. Reprod. Immunol. 2018, 79, e12811. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S. Immunologic factors influencing susceptibility to recurrent candidal vaginitis. Clin. Obstet. Gynecol. 1991, 34, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, M.; Pinelli, M.; Borghi, M.; Constantini, C.; Dindo, M.; van Emst, L.; Puccetti, M.; Pariano, M.; Ricano-Ponce, I.; Bull, C.; et al. A systems genomics approach identifies SIGLEC15 as a susceptibility factor in recurrent vulvovaginal candidiasis. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.; Kolls, J.K.; Happel, K.I.; Wormley, F.; Wozniak, K.L.; Fidel, P.L., Jr. The acute neutrophil response mediated by S100 alarmins during vaginal Candida infections is independent of the Th17-pathway. PLoS ONE 2012, 7, e46311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, A.; Zelante, T.; D’Angelo, C.; Zagarella, S.; Fallarino, F.; Spreca, A.; Iannitti, R.G.; Bonifazi, P.; Renauld, J.C.; Bistoni, F.; et al. IL-22 defines a novel immune pathway of antifungal resistance. Mucosal Immunol. 2010, 3, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Coleman, B.M.; Willems, H.M.E.; Barker, K.S.; Aggor, F.E.Y.; Cipolla, E.; Verma, A.H.; Bishu, S.; Huppler, A.H.; Bruno, V.M.; et al. The IL-17R/IL-22R signaling axis is dispensable for vulvovaginal candidiasis regardless of estrogen status. J. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Moyes, D.L.; Richardson, J.P.; Naglik, J.R. Candida albicans-epithelial interactions and pathogenicity mechanisms: Scratching the surface. Virulence 2015, 6, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Roselletti, E.; Perito, S.; Gabrielli, E.; Mencacci, A.; Pericolini, E.; Sabbatini, S.; Cassone, A.; Vecchiarelli, A. NLRP3 inflammasome is a key player in human vulvovaginal disease caused by Candida albicans. Sci. Rep. 2017, 7, 17877. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G. Chronic vulvovaginal candidiasis: What we know and what we have yet to learn. Australas. J. Dermatol. 2012, 53, 247–254. [Google Scholar] [CrossRef]
- Black, C.A.; Eyers, F.M.; Russell, A.; Dunkley, M.L.; Clancy, R.L.; Beagley, K.W. Acute neutropenia decreases inflammation associated with murine vaginal candidiasis but has no effect on the course of infection. Infect. Immun. 1998, 66, 1273–1275. [Google Scholar] [CrossRef] [Green Version]
- De Bernardis, F.; Boccanera, M.; Adriani, D.; Girolamo, A.; Cassone, A. Intravaginal and intranasal immunizations are equally effective in inducing vaginal antibodies and conferring protection against vaginal candidiasis. Infect. Immun. 2002, 70, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.S.; White, C.J.; Ibrahim, A.S.; Filler, S.G.; Fu, Y.; Yeaman, M.R.; Edwards, J.E., Jr.; Hennessey, J.P., Jr. NDV-3, a recombinant alum-adjuvanted vaccine for Candida and Staphylococcus aureus, is safe and immunogenic in healthy adults. Vaccine 2012, 30, 7594–7600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tso, G.H.W.; Reales-Calderon, J.A.; Pavelka, N. The Elusive Anti-Candida Vaccine: Lessons From the Past and Opportunities for the Future. Front. Immunol. 2018, 9, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lev-Sagie, A.; Goldman-Wohl, D.; Cohen, Y.; Dori-Bachash, M.; Leshem, A.; Mor, U.; Strahilevitz, J.; Moses, A.E.; Shapiro, H.; Yagel, S.; et al. Vaginal microbiome transplantation in women with intractable bacterial vaginosis. Nat. Med. 2019, 25, 1500–1504. [Google Scholar] [CrossRef]
Figure 1.
The factors contributing to recurrent vulvovaginal candidiasis (RVVC) onset.

Figure 2.
Environmental and immunological components that contribute to the host overreaction during RVVC.
Figure 2.
Environmental and immunological components that contribute to the host overreaction during RVVC.


{kind=link}
{kind=link}
Table 1.
Summary of the microbiological factors that act in quorum sensing of vaginal microbiota with potential stimulatory or inhibitory effects on Candida growth/morphological switch.
Table 1.
Summary of the microbiological factors that act in quorum sensing of vaginal microbiota with potential stimulatory or inhibitory effects on Candida growth/morphological switch.
| Factors | Potential Effects on Candida Growth | Reference |
|---|---|---|
| Lactobacilli community | Inhibitory | [17,18,19] |
| Carbon sources: | ||
| Glucose | Stimulatory | [24,25] |
| Lactate | Potentially inhibitory | [21] |
| Short-chain fatty acids (SCFAs): | ||
| Acetate | Inhibitory | [21,22] |
| Butyrate | Inhibitory | [22] |
| Propionate | Inhibitory | [22] |
| Eicosanoids: | ||
| Prostaglandin E2 | Stimulatory | [23] |
| Thromboxane B2 | Stimulatory | [23] |
| Estrogens concentrations | Stimulatory | [20] |
| Low pH (pH: 4–4.5) | Potentially inhibitory | [28] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rosati, D.; Bruno, M.; Jaeger, M.; ten Oever, J.; Netea, M.G. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms 2020, 8, 144. https://doi.org/10.3390/microorganisms8020144
AMA Style
Rosati D, Bruno M, Jaeger M, ten Oever J, Netea MG. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms. 2020; 8(2):144. https://doi.org/10.3390/microorganisms8020144
Chicago/Turabian StyleRosati, Diletta, Mariolina Bruno, Martin Jaeger, Jaap ten Oever, and Mihai G. Netea. 2020. "Recurrent Vulvovaginal Candidiasis: An Immunological Perspective" Microorganisms 8, no. 2: 144. https://doi.org/10.3390/microorganisms8020144
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.