
Antimicrobial Effects of Potential Probiotics of Bacillus spp. Isolated from Human Microbiota: In Vitro and In Silico Methods

 ,
,  ,
,  ,
,  ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microbiota Sampling Bank and Directed Culturing Approach
2.2. BPA Microbiota Tolerance Testing
2.3. Culturing- Isolation of Bacillus Catalogue
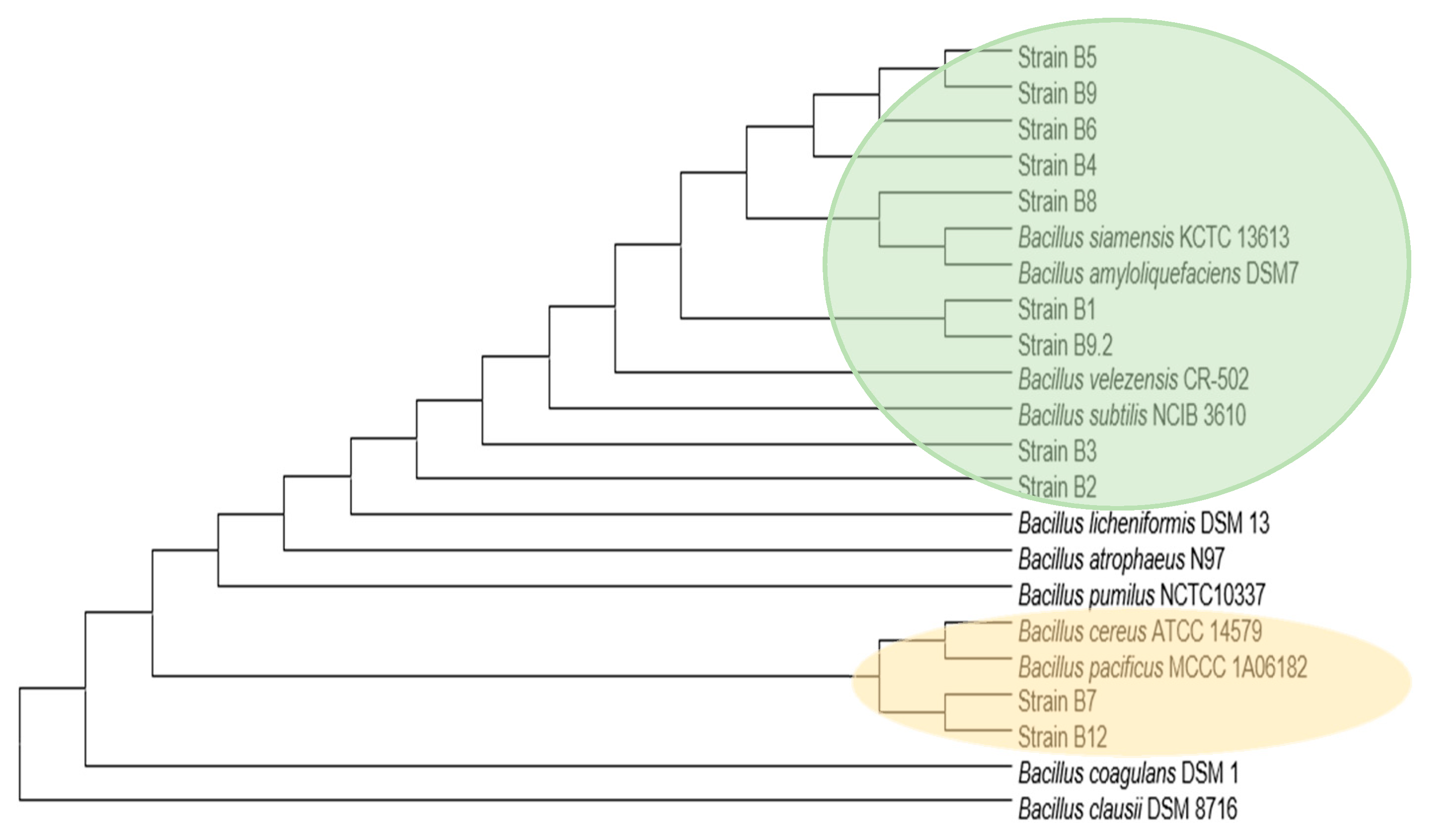
2.4. Genomic DNA Extraction, Taxonomy Identification and Phylogenetic Analysis
2.5. Enzymes Tests
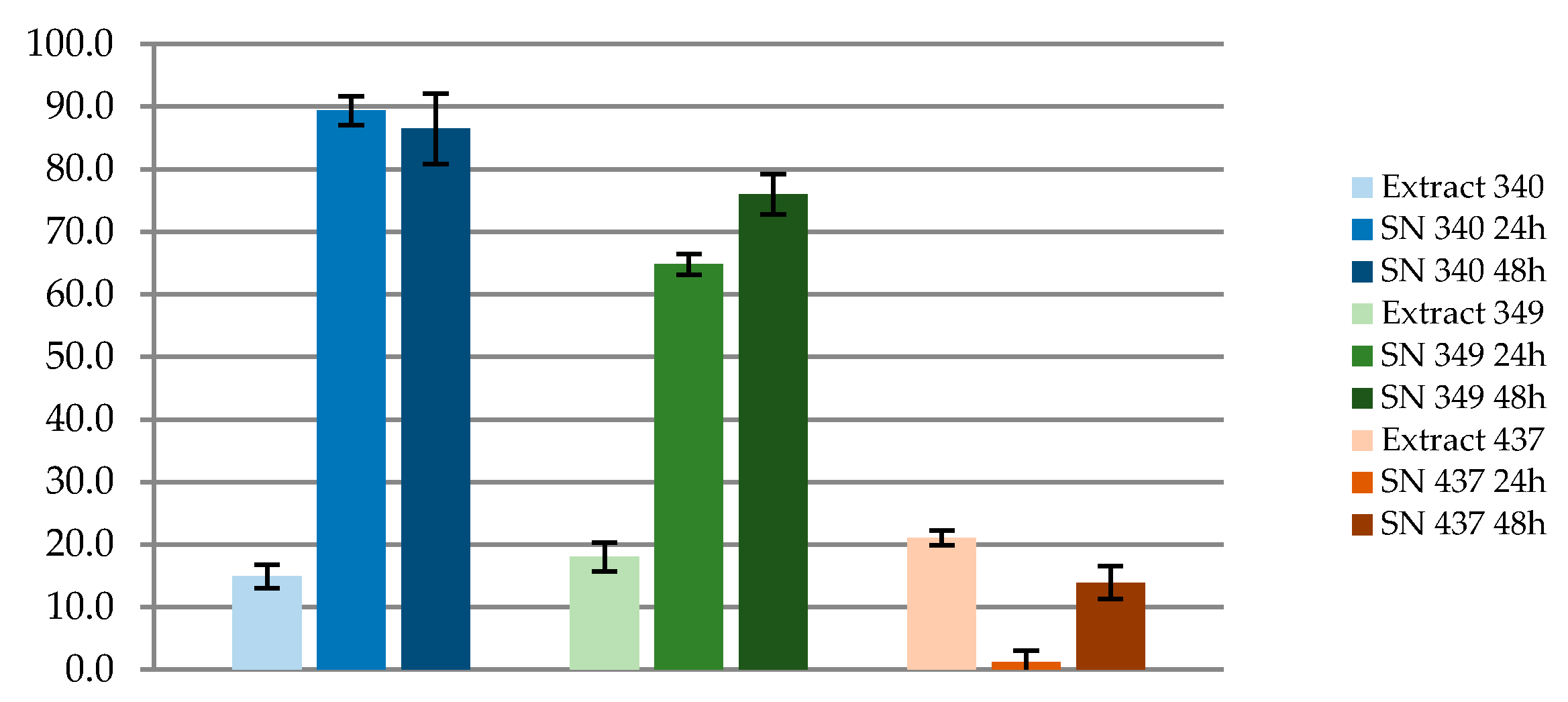
2.6. Antimicrobial In Vitro Tests
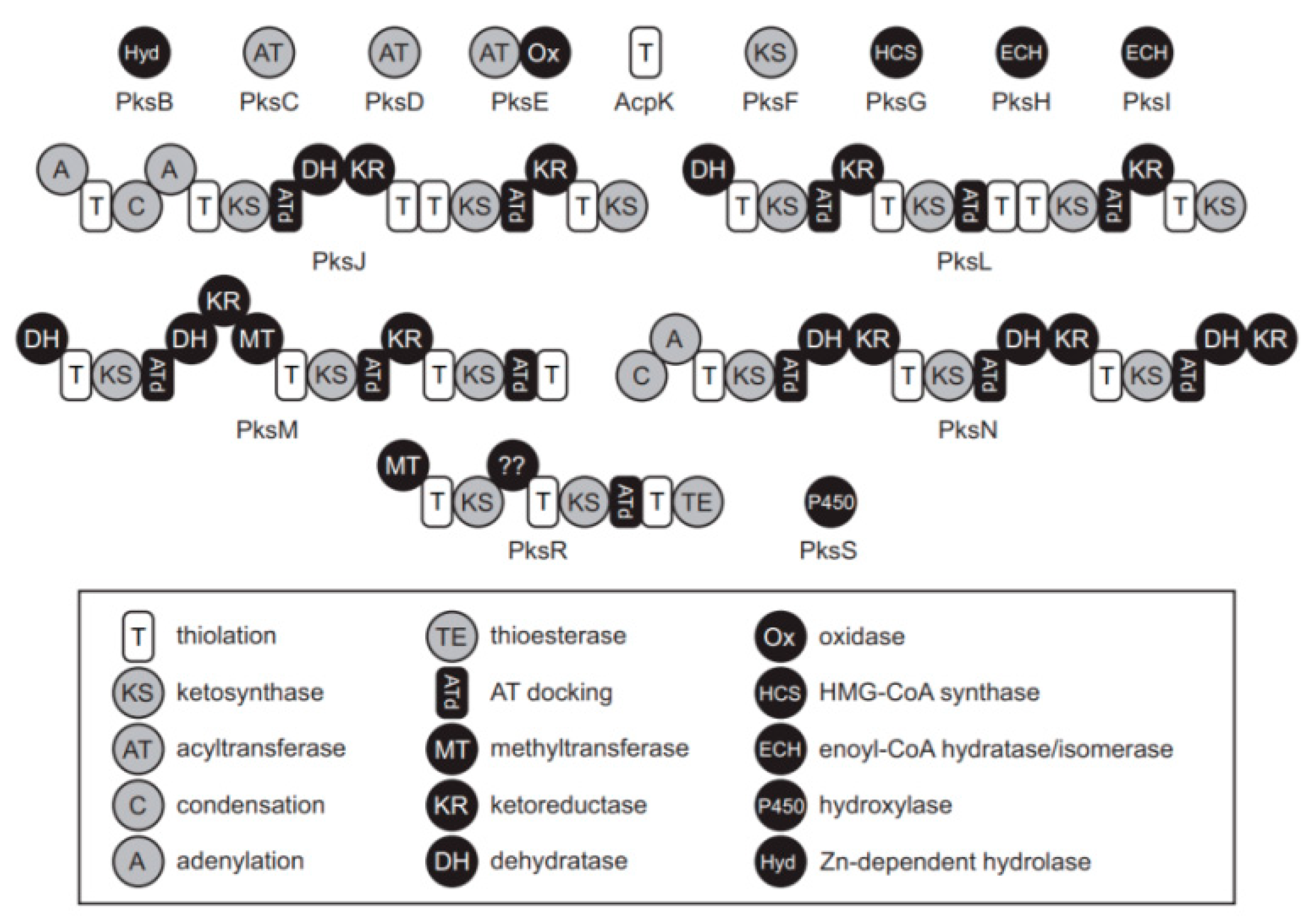
2.7. Genome Data Mining and Analysis –PKs Genes and Clusters
2.7.1. Genome Mining Tools for PKs Gene Searching
2.7.2. Prediction of Polyketides in WGS of Bacillus sp. AM1 Isolated from Microbiota
3. Results and Discussion
3.1. BPA-Tolerant Microorganisms Isolated from Human Gut Microbiota
3.1.1. BPA Microbiota Metabolization Capacities
3.1.2. Catalogue of BPA-Tolerant Bacillus spp. Isolated from Human Microbiota
3.1.3. Taxonomical and Phylogenetic Clustering
3.2. Analysis of Bioactive Compounds Production Capacities
3.2.1. Enzymatic Activity Tests
3.2.2. Antimicrobial Activity Tests
3.3. WGS Data Mining and In Silico Analysis
3.3.1. WGS Mining in Type Strains
3.3.2. WGS Representative Bacillus sp. AM1 from Microbiota: Genome Mining Data
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AN | Anorexia Nervosa |
| BHI | Brain-Hearth Infusion |
| BPA | Bisphenol A |
| CIAs | Critically Important Antimicrobials |
| CIC | Centro de Instrumentación Científica |
| EC | Enzyme Commission number |
| EFSA | European Food Safety Authority |
| GRAS | Generally Recognized as Safe |
| HC | Healthy control |
| HIAs | Highly Important Antimicrobials |
| IPBLN | Institute of Parasitology and Biomedicine “López-Neyra” |
| JECFA | Joint FAO/WHO Expert Committee on Food AdditivesLiquid |
| LC-MS/MS | Liquid Chromatography-Mass Spectrometry |
| MCL | Maximum Composite Likelihood |
| MDC | Microbiota Disrupting Chemicals |
| MetS | Metabolic syndrome |
| MIC | Minimum Inhibitory Concentration |
| MRS | Man-Rogosa-Sharpe |
| NAFLD | Nonalcoholic fatty liver disease |
| NCBI | National Center for Biotechnology Information |
| NGP | Next Generation Probiotics |
| NGS | Next Generation Sequencing |
| NMR | Nuclear Magnetic Resonance |
| NRPS | Nonribosomal Peptide-synthetase |
| OB | Obesity |
| OW | Over-weigth |
| PKs | Poliketides |
| QPS | Qualified Presumption of Safety |
| SN | Supernatant |
| T1D | Type 1 Diabetes |
| T2D | Type 2 Diabetes |
| WGS | Whole Genome Sequences |
| WHO | World Health Organization |
References
- Casado, V.; Martín, D.; Torres, C.; Reglero, G. Phospholipases in food industry: A review. Methods Mol. Biol. 2012, 861, 495–523. [Google Scholar] [PubMed]
- Moran, K.; de Lange, C.F.M.; Ferket, P.; Fellner, V.; Wilcock, P.; van Heugten, E. Enzyme supplementation to improve the nutritional value of fibrous feed ingredients in swine diets fed in dry or liquid form. J. Anim. Sci. 2016, 94, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.-C.; Armougom, F.; Million, M.; Hugon, P.; Pagnier, I.; Robert, C.; Bittar, F.; Fournous, G.; Gimenez, G.; Maraninchi, M.; et al. Microbial culturomics: Paradigm shift in the human gut microbiome study. Clin. Microbiol. Infect. 2012, 18, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Moreno, A.; Acuña, I.; Torres-Sánchez, A.; Ruiz-Moreno, Á.; Cerk, K.; Rivas, A.; Suárez, A.; Monteoliva-Sánchez, M.; Aguilera, M. Next Generation Probiotics for Neutralizing Obesogenic Effects: Taxa Culturing Searching Strategies. Nutrients 2021, 13, 1617. [Google Scholar] [CrossRef]
- Aguilera, M.; Gálvez-Ontiveros, Y.; Rivas, A. Endobolome, a New Concept for Determining the Influence of Microbiota Disrupting Chemicals (MDC) in Relation to Specific Endocrine Pathogenesis. Front. Microbiol. 2020, 11, 578007. [Google Scholar] [CrossRef] [PubMed]
- Adair, K.L.; Douglas, A.E. Making a microbiome: The many determinants of host-associated microbial community composition. Curr. Opin. Microbiol. 2017, 35, 23–29. [Google Scholar] [CrossRef]
- Diakite, A.; Dubourg, G.; Dione, N.; Afouda, P.; Bellali, S.; Ngom, I.I.; Valles, C.; Million, M.; Levasseur, A.; Cadoret, F.; et al. Extensive culturomics of 8 healthy samples enhances metagenomics efficiency. PLoS ONE 2019, 14, e0223543. [Google Scholar] [CrossRef] [Green Version]
- WoldemariamYohannes, K.; Wan, Z.; Yu, Q.; Li, H.; Wei, X.; Liu, Y.; Wang, J.; Sun, B. Prebiotic, Probiotic, Antimicrobial, and Functional Food Applications of Bacillus amyloliquefaciens. J. Agric. Food Chem. 2020, 68, 14709–14727. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus coagulans in the Food Industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut microbiota as a source of novel antimicrobials. Gut Microbes 2019, 10, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Zhu, C.; Li, H.; Yin, M.; Pan, C.; Huang, L.; Kong, C.; Wang, X.; Zhang, Y.; Qu, S.; et al. Dysbiosis Signatures of Gut Microbiota Along the Sequence from Healthy, Young Patients to Those with Overweight and Obesity. Obesity 2018, 26, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Razavi, S.; Navab-Moghadam, F.; Khamseh, M.E.; Alaei-Shahmiri, F.; Mehrtash, A.; Amirmozafari, N. Comparison of gut microbiota in adult patients with type 2 diabetes and healthy individuals. Microb. Pathog. 2017, 111, 362–369. [Google Scholar] [CrossRef]
- Ahmad, A.; Yang, W.; Chen, G.; Shafiq, M.; Javed, S.; Zaidi, S.S.A.; Shahid, R.; Liu, C.; Bokhari, H. Analysis of gut microbiota of obese individuals with type 2 diabetes and healthy individuals. PLoS ONE 2019, 14, e0226372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejtahed, H.-S.; Hoseini-Tavassol, Z.; Khatami, S.; Zangeneh, M.; Behrouzi, A.; Ahmadi Badi, S.; Moshiri, A.; Hasani-Ranjbar, S.; Soroush, A.-R.; Vaziri, F.; et al. Main gut bacterial composition differs between patients with type 1 and type 2 diabetes and non-diabetic adults. J. Diabetes Metab. Disord. 2020, 19, 265–271. [Google Scholar] [CrossRef]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered Fecal Microbiota Correlates with Liver Biochemistry in Nonobese Patients with Non-alcoholic Fatty Liver Disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef]
- Li, F.; Sun, G.; Wang, Z.; Wu, W.; Guo, H.; Peng, L.; Wu, L.; Guo, X.; Yang, Y. Characteristics of fecal microbiota in non-alcoholic fatty liver disease patients. Sci. China Life Sci. 2018, 61, 770–778. [Google Scholar] [CrossRef]
- Raman, M.; Ahmed, I.; Gillevet, P.M.; Probert, C.S.; Ratcliffe, N.M.; Smith, S.; Greenwood, R.; Sikaroodi, M.; Lam, V.; Crotty, P.; et al. Fecal Microbiome and Volatile Organic Compound Metabolome in Obese Humans With Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2013, 11, 868–875.e3. [Google Scholar] [CrossRef]
- Nistal, E.; Sáenz de Miera, L.E.; Ballesteros Pomar, M.; Sánchez-Campos, S.; García-Mediavilla, M.V.; Álvarez-Cuenllas, B.; Linares, P.; Olcoz, J.L.; Arias-Loste, M.T.; García-Lobo, J.M.; et al. An altered fecal microbiota profile in patients with non-alcoholic fatty liver disease (NAFLD) associated with obesity. Rev. Española Enferm. Dig. 2019, 111, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.Y.; You, H.J.; Yoon, H.S.; Kwon, B.; Lee, J.Y.; Lee, S.; Song, Y.-M.; Lee, K.; Sung, J.; Ko, G. The effect of heritability and host genetics on the gut microbiota and metabolic syndrome. Gut 2017, 66, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human exposure to bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.R.; Schoenfelder, G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Cienc. Saude Coletiva 2012, 17, 407–434. [Google Scholar] [CrossRef]
- Louati, I.; Dammak, M.; Nasri, R.; Belbahri, L.; Nasri, M.; Abdelkafi, S.; Mechichi, T. Biodegradation and detoxification of bisphenol A by bacteria isolated from desert soils. 3 Biotech 2019, 9, 228. [Google Scholar] [CrossRef]
- Gramec Skledar, D.; Peterlin Mašič, L. Bisphenol A and its analogs: Do their metabolites have endocrine activity? Environ. Toxicol. Pharmacol. 2016, 47, 182–199. [Google Scholar] [CrossRef] [PubMed]
- Gálvez-Ontiveros, Y.; Moscoso-Ruiz, I.; Rodrigo, L.; Aguilera, M.; Rivas, A.; Zafra-Gómez, A. Presence of parabens and bisphenols in food commonly consumed in spain. Foods 2021, 10, 92. [Google Scholar] [CrossRef]
- Cohen, I.C.; Cohenour, E.R.; Harnett, K.G.; Schuh, S.M. BPA, BPAF and TMBPF Alter Adipogenesis and Fat Accumulation in Human Mesenchymal Stem Cells, with Implications for Obesity. Int. J. Mol. Sci. 2021, 22, 5363. [Google Scholar] [CrossRef]
- Camacho, L.; Lewis, S.M.; Vanlandingham, M.M.; Olson, G.R.; Davis, K.J.; Patton, R.E.; Twaddle, N.C.; Doerge, D.R.; Churchwell, M.I.; Bryant, M.S.; et al. A two-year toxicology study of bisphenol A (BPA) in Sprague-Dawley rats: CLARITY-BPA core study results. Food Chem. Toxicol. 2019, 132, 110728. [Google Scholar] [CrossRef]
- Gundert-Remy, U.; Bodin, J.; Bosetti, C.; FitzGerald, R.; Hanberg, A.; Hass, U.; Hooijmans, C.; Rooney, A.A.; Rousselle, C.; van Loveren, H.; et al. Bisphenol A (BPA) hazard assessment protocol. EFSA Support. Publ. 2017, 14, 1354E. [Google Scholar]
- Gómez-Gallego, C.; Pohl, S.; Salminen, S.; De Vos, W.M.; Kneifel, W. Akkermansia muciniphila: A novel functional microbe with probiotic properties. Benef. Microbes 2016, 7, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Brodmann, T.; Endo, A.; Gueimonde, M.; Vinderola, G.; Kneifel, W.; de Vos, W.M.; Salminen, S.; Gómez-Gallego, C. Safety of Novel Microbes for Human Consumption: Practical Examples of Assessment in the European Union. Front. Microbiol. 2017, 8, 1725. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; Cesare, A.D.; Hilbert, F.; Lindqvist, R.; et al. Scientific Opinion on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA (2017–2019). EFSA J. 2020, 18, e05966. [Google Scholar] [PubMed] [Green Version]
- Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Dusemund, B.; Kouba, M.; Durjava, M.F.; López-Alonso, M.; Puente, S.L.; Marcon, F.; et al. Safety and efficacy of Bacillus subtilisPB6 (Bacillus velezensisATCC PTA-6737) as a feed additive for chickens for fattening, chickens reared for laying, minor poultry species (except for laying purposes), ornamental, sporting and game birds. EFSA J. 2020, 18, e06280. [Google Scholar] [PubMed]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.d.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [PubMed]
- Aleti, G.; Sessitsch, A.; Brader, G. Genome mining: Prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Comput. Struct. Biotechnol. J. 2015, 13, 192–203. [Google Scholar] [CrossRef]
- Straight, P.D.; Fischbach, M.A.; Walsh, C.T.; Rudner, D.Z.; Kolter, R. A singular enzymatic megacomplex from Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2007, 104, 305–310. [Google Scholar] [CrossRef] [Green Version]
- García-Córcoles, M.T.; Cipa, M.; Rodríguez-Gómez, R.; Rivas, A.; Olea-Serrano, F.; Vílchez, J.L.; Zafra-Gómez, A. Determination of bisphenols with estrogenic activity in plastic packaged baby food samples using solid-liquid extraction and clean-up with dispersive sorbents followed by gas chromatography tandem mass spectrometry analysis. Talanta 2018, 178, 441–448. [Google Scholar] [CrossRef]
- López-Moreno, A.; Torres-Sánchez, A.; Acuña, I.; Suárez, A.; Aguilera, M. Representative Bacillus sp. AM1 from Gut Microbiota Harbor Versatile Molecular Pathways for Bisphenol A Biodegradation. Int. J. Mol. Sci. 2021, 22, 4952. [Google Scholar] [CrossRef]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Menasria, T.; Aguilera, M.; Hocine, H.; Benammar, L.; Ayachi, A.; Si Bachir, A.; Dekak, A.; Monteoliva-Sánchez, M. Diversity and bioprospecting of extremely halophilic archaea isolated from Algerian arid and semi-arid wetland ecosystems for halophilic-active hydrolytic enzymes. Microbiol. Res. 2018, 207, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Montalvo-Rodríguez, R.; Vreeland, R.H.; Oren, A.; Kessel, M.; Betancourt, C.; López-Garriga, J. Halogeometricum borinquense gen. nov., sp. nov., a novel halophilic archaeon from Puerto Rico. Int. J. Syst. Bacteriol. 1998, 48 Pt 4, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Allais, J.J.; Hoyos-Lopez, G.; Kammoun, S.; Baratti, J.C. Isolation and characterization of thermophilic bacterial strains with inulinase activity. Appl. Environ. Microbiol. 1987, 53, 942–945. [Google Scholar] [CrossRef] [Green Version]
- Sierra, G. A simple method for the detection of lipolytic activity of micro-organisms and some observations on the influence of the contact between cells and fatty substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Jeffries, C.D.; Holtman, D.F.; Guse, D.G. Rapid method for determining the activity of microorganisms on nucleic acids. J. Bacteriol. 1957, 73, 590–591. [Google Scholar] [CrossRef] [Green Version]
- Combined Compendium of Food Additive Specifications. Available online: http://www.fao.org/3/a0691e/a0691e00.htm (accessed on 24 June 2021).
- Powthong, P.; Suntornthiticharoen, P. Antimicrobial and enzyme activity produced by Bacillus spp. Isolated from soil. Int. J. Pharm. Pharm. Sci. 2017, 9, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Rangwala, S.H.; Kuznetsov, A.; Ananiev, V.; Asztalos, A.; Borodin, E.; Evgeniev, V.; Joukov, V.; Lotov, V.; Pannu, R.; Rudnev, D.; et al. Accessing NCBI data using the NCBI Sequence Viewer and Genome Data Viewer (GDV). Genome Res. 2020, 31, 159–169. [Google Scholar] [CrossRef]
- Yu, K.; Yi, S.; Li, B.; Guo, F.; Peng, X.; Wang, Z.; Wu, Y.; Alvarez-Cohen, L.; Zhang, T. An integrated meta-omics approach reveals substrates involved in synergistic interactions in a bisphenol A (BPA)-degrading microbial community. Microbiome 2019, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Pranteda, M.L.; Pérez-Davó, A.; Monteoliva-Sánchez, M.; Ramos-Cormenzana, A.; Aguilera, M. Food Omics Validation: Towards Understanding Key Features for Gut Microbiota, Probiotics and Human Health. Food Anal. Methods 2015, 8, 272–289. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Szlufman, C.; Shemesh, M. Role of Probiotic Bacilli in Developing Synbiotic Food: Challenges and Opportunities. Front. Microbiol. 2021, 12, 811. [Google Scholar] [CrossRef] [PubMed]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus velezensis as a probiotic in animal feed: A review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef]
- Sultana, O.; Lee, S.; Seo, H.; Mahmud, H.A.; Kim, S.; Seo, A.; Kim, M.; Song, H.-Y. Biodegradation and removal of PAH by Bacillus velezensis isolated from fermented food. J. Microbiol. Biotechnol. 2021, 31. [Google Scholar] [CrossRef]
- Kang, M.; Choi, H.J.; Yun, B.; Lee, J.; Yoo, J.; Yang, H.-J.; Jeong, D.-Y.; Kim, Y.; Oh, S. Bacillus amyloliquefaciens SCGB1 Alleviates Dextran Sulfate Sodium-Induced Colitis in Mice Through Immune Regulation. J. Med. Food 2021, 24, 709–719. [Google Scholar] [CrossRef]
- Harwood, C.R.; Mouillon, J.-M.; Pohl, S.; Arnau, J. Secondary metabolite production and the safety of industrially important members of the Bacillus subtilis group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef]
- Bianco, A.; Capozzi, L.; Monno, M.R.; Del Sambro, L.; Manzulli, V.; Pesole, G.; Loconsole, D.; Parisi, A. Characterization of Bacillus cereus Group Isolates From Human Bacteremia by Whole-Genome Sequencing. Front. Microbiol. 2021, 11, 599524. [Google Scholar] [CrossRef]
- Devaraj, K.; Aathika, S.; Periyasamy, K.; Periyaraman, P.M.; Palaniyandi, S.; Subramanian, S. Production of thermostable multiple enzymes from Bacillus amyloliquefaciens KUB29. Nat. Prod. Res. 2019, 33, 1674–1677. [Google Scholar] [CrossRef]
- Deb, P.; Talukdar, S.A.; Mohsina, K.; Sarker, P.K.; Sayem, S.A. Production and partial characterization of extracellular amylase enzyme from Bacillus amyloliquefaciens P-001. SpringerPlus 2013, 2, 154. [Google Scholar] [CrossRef] [Green Version]
- Latorre, J.D.; Hernandez-Velasco, X.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Menconi, A.; Bielke, L.R.; Hargis, B.M.; Tellez, G. Evaluation and Selection of Bacillus Species Based on Enzyme Production, Antimicrobial Activity, and Biofilm Synthesis as Direct-Fed Microbial Candidates for Poultry. Front. Vet. Sci. 2016, 3, 95. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, K.; Thilakan, B.; Kizhakkekalam, V.K. Antibacterial aryl-crowned polyketide from Bacillus subtilis associated with seaweed Anthophycus longifolius. J. Appl. Microbiol. 2018, 124, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Piel, J.; Butzke, D.; Fusetani, N.; Hui, D.; Platzer, M.; Wen, G.; Matsunaga, S. Exploring the Chemistry of Uncultivated Bacterial Symbionts: Antitumor Polyketides of the Pederin Family. J. Nat. Prod. 2005, 68, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Angelakis, E. Weight gain by gut microbiota manipulation in productive animals. Microb. Pathog. 2017, 106, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Hoover, D.G.; Harlander, S.K. CHAPTER 2—Screening Methods for Detecting Bacteriocin Activity. In Bacteriocins of Lactic Acid Bacteria; Hoover, D.G., Steenson, L.R., Eds.; Academic Press: Cambridge, MA, USA, 1993; pp. 23–39. ISBN 978-0-12-355510-6. [Google Scholar]
- Tang, G.-L.; Cheng, Y.-Q.; Shen, B. Leinamycin biosynthesis revealing unprecedented architectural complexity for a hybrid polyketide synthase and nonribosomal peptide synthetase. Chem. Biol. 2004, 11, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-W.; Niu, Q.-H.; Zhou, W.; Zhang, K.-Q. Bacillus nematocida sp. nov., a novel bacterial strain with nematotoxic activity isolated from soil in Yunnan, China. Syst. Appl. Microbiol. 2005, 28, 323–327. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Statement on the Requirements for Whole Genome Sequence Analysis of Microorganisms Intentionally Used in the Food Chain, 2021. Available online: https://www.efsa.europa.eu/sites/default/files/2021-03/EFSA-statement-EFSA-Q-2019-00434.pdf (accessed on 30 May 2021).
- Swann, J.R.; Rajilic-Stojanovic, M.; Salonen, A.; Sakwinska, O.; Gill, C.; Meynier, A.; Fança-Berthon, P.; Schelkle, B.; Segata, N.; Shortt, C.; et al. Considerations for the design and conduct of human gut microbiota intervention studies relating to foods. Eur. J. Nutr. 2020, 59, 3347–3368. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.R.; Sardi, J.d.C.O.; Pitangui, N.d.S.; Roque, S.M.; Silva, A.C.B.d.; Rosalen, P.L. Probiotics as an alternative antimicrobial therapy: Current reality and future directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- World Health Organization. WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance. In Critically Important Antimicrobials for Human Medicine: Ranking of Antimicrobial Agents for Risk Management of Antimicrobial Resistance Due to Non-Human Use; World Health Organization: Geneva, Switzerland, 2017; ISBN 978-92-4-151222-0. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Clinical Trials—Disease /Sample Size and Clinical Traits | Taxa Modifications |
|---|---|---|
| [13] | OB; n = 192; HC n = 25; OW n = 22; OB n = 145 | ↑ Bacillus in OW and OB |
| [14] | OB, AN; n = 49; HC n = 20; OB n = 20; AN n = 9 | ↑ Lactobacillus in OB |
| [15] | T2D; n = 36; HC n = 18; T2D n = 18 | ↑ Lactobacillus in T2D |
| [16] | T2D, OB; n = 60; HC n = 20; Obese-T2D n = 40 | ↑ Bacillus sporothermodurans in OB-T2D |
| [17] | T1D, T2D; n = 110; HC n = 40; T2D n = 49; T1D n = 21 | ↑ Lactobacillus in T1D and T2D |
| [18] | NAFLD; n = 126; HC n = 83; NAFLD n = 43 | ↓ Lactobacillus in NAFLD |
| [19] | NAFLD; n = 67; HC n = 37; NAFLD n = 30 | ↑ Lactobacillaceae in NAFLD |
| [20] | NAFLD; n = 60; HC n = 30; NAFLD n = 30 | ↑ Lactobacillus in NAFLD |
| [21] | NAFLD, OB; n = 73; HC n = 20; OB-NAFLD n = 36; OB-non-NAFLD n = 17 | ↑ Bacilli in OB-NAFLD ↑ Lactobacillus in non-NAFLD |
| [22] | MetS; n = 655; Monozygotic twins n = 306; Dizygotic twins n = 74; Siblings n = 275 | ↑ Lactobacillus in MetS |
| Microbiota Isolates | Closest Taxa—[Strain] Best Hit | bp Position 16S rRNA | Query Cover (%) | Identity (%) | Accession Number |
|---|---|---|---|---|---|
| B1 | Bacillus siamensis [LRM10-3D] | 15,030 | 100 | 100 | MT645306.1 |
| Bacillus velezensis [XC1] | 100 | 100 | MT649755.1 | ||
| B2 | Bacillus velezensis [CR-502] | 1483 | 95.4 | 99.14 | AY603658 |
| B3 | Bacillus siamensis [KCTC 13613] | 1490 | 100 | 98.00 | AJVF01000043 |
| B4 | Bacillus siamensis [KCTC 13613] | 1515 | 100 | 99.66 | AJVF01000043 |
| Bacillus nematocida [B-16] | 100 | 99.73 | AY820954 | ||
| Bacillus amyloliquefaciens [DSM7] | 100 | 99.52 | FN597644 | ||
| B5 | Bacillus siamensis [KCTC 13613] | 1516 | 100 | 98.91 | AJVF01000043 |
| Bacillus nematocida [B-16] | 100 | 98.98 | AY820954 | ||
| Bacillus velezensis [CR-502] | 95.4 | 99.22 | AY603658 FN597644 | ||
| Bacillus amyloliquefaciens [DSM7] | 100 | 98.78 | |||
| B6 | Bacillus velezensis [CR-502] | 1504 | 95.4 | 99.93 | AY603658 |
| B7 | Bacillus cereus [AFS039342] | 1510 | 100 | 99.39 | NUMR01000072 |
| Bacillus pacificus [NCCP 15909] | 100 | 99.34 | CP041979.1 | ||
| B8 | Bacillus velezensis [CR-502] | 1520 | 95.4 | 99.93 | AY603658 |
| B9 | Bacillus velezensis [CR-502] | 1499 | 95.4 | 99.22 | AY603658 |
| B9.2 | Bacillus siamensis [KCTC 13613] | 1499 | 100 | 99.52 | AJVF01000043 |
| Bacillus nematocida [B-16] | 100 | 99.59 | AY820954 | ||
| Bacillus amyloliquefaciens [DSM 7] | 100 | 99.39 | FN597644 | ||
| B12 | Bacillus cereus [AFS039342] | 1543 | 100 | 99.39 | JMQC01000008 |
| Bacillus pacificus [NCCP 15909] | 99.0 | 99.35 | CP041979.1 |
| Enzyme Test | Microbiota Isolates | ||
|---|---|---|---|
| rB1 | rB3 | rB7 | |
| Starch | + | ++ | ++ |
| Carboxymethylcellulose | - | - | - |
| Inulin | + | - | + |
| Tween 80 | - | - | - |
| DNase | ++ | - | - |
| Target Indicator Bacteria | Strains rB1 | Strains rB3 | Strains rB7 |
|---|---|---|---|
| Diameter of inhibitory zone (mm) ± SD 1 | |||
| Bacillus cereus | 15 ± 0 | 17 ± 0 | - |
| Bacillus circulans | 13 ± 0 | 14.3 ± 1.2 | - |
| Staphylococcus aureus | 11.7 ± 0.6 | 10 ± 0 | - |
| Streptococcus pyogenes | 15 ± 0 | 13.3 ± 0.6 | - |
| Serratia marcescens | 17 ± 0 | 15.3 ± 1.5 | - |
| E. coli | 15 ± 0 | 13.3 ± 0.6 | - |
| Salmonella | 11 ± 0 | 10 ± 0 | - |
| Klebsiella | 20 ± 0 * | 15 ± 0 * | - |
| Pseudomonas | - | - | - |
| Enzyme | Enzyme description EC number | B. amyloliquefaciens WF02T NZ_CP053376 | B. siamenensis SCSIO 05746T NZ_CP025001 | B. velezensis CBMB205T NZ_CP011937 | B. subtilis 168T NC_000964 | B. atrophaeus BSST NZ_CP007640 | B. sp-AM1 B1T CP047644.1) |
|---|---|---|---|---|---|---|---|
| PksA | Hypothetical protein/EC:3.1.2.6 | WP_024085315.1 174131..1741526 | WP_060962748.1 2494188..2494397 | WP_032874955.1 2222103..2222312 | NP_000389590.1 1782713..1783390 | WP_013390522.1 1165636..1167084 | 1787442..1787651 QHJ03379.1 |
| - | Hypothetical protein/EC:3.1.2.6 | WP_024085326.1 1816193..1816555 | WP_016936035.1 2419160..2419522 | WP_007410383.1 2146808..2147170 | YP_0009513956.1 1783500..1783766 | WP_003328852.1 1167393..1167932 | - |
| Regulator | TetR family transcriptional regulator C terminal | - | - | - | NP_000389589.1 1781906..1782523 | WP_003328851.1 1168054..1168644 | - |
| PksB | MBL fold metallo hydrolase/ EC: 2.3.1.39 | WP_024085316.1 1742160..1742837 | WP_060962747.1 2492787..2493464 | WP_032874957.1 2220496..2221173 | YP_0009513956.1 1783500..1783766 | WP_003328850.1 1168942..1169619 | 1788295..1788972 QHJ03380.1 |
| PksC | ACP S malonyltransferase/ EC:2.3.1.51 | WP_014305029.1 1743152..1744021 | WP_060962746.1 2491603..2492472 | WP_032874959.1 2219312..2220181 | NP_000389591.1 1783763..1784629 | WP_003328849.1 1170013..1170879 | 1789287..1790156 QHJ03381.1 |
| PksD | Acyltransferase domain containing protein/EC: 2.3.1.39 | WP_003154101.1 1744158..1745132 | WP_060962745.1 2490494..2491468 | WP_032874961.1 2218201..2219175 | NP_000389592.2 1785133..1786107 | WP_003328847.1 1171417..1172382 | 1790293..1791267 QHJ03382.1 |
| PksE | ACP S malonyltransferase/ EC:1.3.1.9 and 1.3.1.10 | WP_003154100.1 1745134..1747374 | ID Not found 2488250..2490492 | WP_032874963.1 2215959..2218199 | NP_000389593.3 1786104..1788407 | WP_003328846.1 1172389..1174752 | 1791269..1793509 QHJ03383.1 |
| AcpK | Acyl carrier protein/EC:2.3.3.10 | WP_003154099.1 1747440..1747688 | WP_060962743.1 2487934..2488182 | WP_012117592.1 2215645..2215893 | NP_00570904.1 1788469..1788717 | WP_003328845.1 1174891..1175139 | 1793575..1793823 QHJ03384.1 |
| PksF | Polyketide beta ketoacyl:ACP synthase/EC: 4.2.1.17 | - | - | - | NP_000389594.2 1788695..1789942 | WP_003328844.1 1175117..1176364 | - |
| PksG | Hydroxymethylglutaryl CoA synthase family/EC: 4.2.1.17 | WP_003154098.1 1747740..1749002 | WP_060962742.1 2486620..2487882 | WP_032874965.1 2214331..2215593 | NP_000389595.2 1789943..1791205 | WP_010788667.1 1176364..1177626 | 1793875..1795137 QHJ03385.1 |
| PksH | Enoyl CoA hydratase/isomerase | WP_024085319.1 1748999..1749772 | WP_060962741.1 2485850..2486623 | WP_032874967.1 2213561..2214334 | NP_000389596.1 1791193..1791972 | WP_087941777.1 1177614..1178390 | 1795134..1795907 QHJ03386.1 |
| PksI | enoyl CoA hydratase/isomerase family protein | WP_003154094.1 1749782..1750531 | WP_060962740.1 2485091..2485840 | WP_003154094.1 2212802..2213551 | NP_000389597.2 1792012..1792761 | WP_003328841.1 1178438..1179184 | 1795917..1796666 QHJ03387.1 |
| PksJ | Non ribosomal peptide synthetase | WP_024085320.1 1750571..1765525 | WP_060962739.1 2470129..2485062 | WP_032874969.1 2197814..2212762 | NP_000389598.3 1792806..1807937 | WP_013390525.1 1179247..1194429 | 1796706..1811657 QHJ03388.1 |
| PksM | SDR family NAD(P) dependent oxidoreductase EC:1.6.5.2 | WP_165869029.1 1765509..1778951 | WP_167388675.1 2456724..2470145 | WP_162859398.1 2184400..2197830 | NP_000389601.3 1821553..1834341 | WP_013390526.1 1194431..1208248 | 1811659..1825086 QHJ03389.1 |
| PksM | SDR family NAD(P) dependent oxidoreductase/EC:1.6.5.2 | WP_024085322.1 1778969..1789513 | WP_101605493.1 2446202..2456707 | WP_032874973.1 2173847..2184382 | NP_000389602.3 1834409..1850875 | WP_013390527.1 1208267..1221238 | 1825104..1835639 QHJ03390.1 |
| PksN | Non ribosomal peptide synthetase | - | WP_101605492.1 2429908 2446212 | WP_032874975.1 2157559 2173857 | NP_000389604.2 1850890 1858521 | WP_087941783.1 1221318..1237793 | 1835629..1851930 QHJ03391.1 |
| PksR | Polyketide synthase dehydratase domain/EC:2.1.1.- | WP_024085324.1 1805818..1813275 | WP_060962735.1 2422440..2429894 | WP_032874977.1 2150088..2157545 | NP_000389600.3 1807921..1821537 | WP_003328830.1 1237809..1245533 | 1851944..1859401 QHJ03392.1 |
| PksS | Cytochrome P450/EC:1.14.14.- | WP_024085325.1 1813410..1814621 | WP_060962734.1 2421090..2422301 | WP_032875233.1 2148742..2149953 | NP_000389605.2 1858566..1859783 | WP_003328829.1 1245647..1246888 | 1859536..1860747 QHJ03393.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Sánchez, A.; Pardo-Cacho, J.; López-Moreno, A.; Ruiz-Moreno, Á.; Cerk, K.; Aguilera, M. Antimicrobial Effects of Potential Probiotics of Bacillus spp. Isolated from Human Microbiota: In Vitro and In Silico Methods. Microorganisms 2021, 9, 1615. https://doi.org/10.3390/microorganisms9081615
Torres-Sánchez A, Pardo-Cacho J, López-Moreno A, Ruiz-Moreno Á, Cerk K, Aguilera M. Antimicrobial Effects of Potential Probiotics of Bacillus spp. Isolated from Human Microbiota: In Vitro and In Silico Methods. Microorganisms. 2021; 9(8):1615. https://doi.org/10.3390/microorganisms9081615
Chicago/Turabian StyleTorres-Sánchez, Alfonso, Jesús Pardo-Cacho, Ana López-Moreno, Ángel Ruiz-Moreno, Klara Cerk, and Margarita Aguilera. 2021. "Antimicrobial Effects of Potential Probiotics of Bacillus spp. Isolated from Human Microbiota: In Vitro and In Silico Methods" Microorganisms 9, no. 8: 1615. https://doi.org/10.3390/microorganisms9081615