
Fungal Iron Biomineralization in Río Tinto


Abstract
:1. Introduction
2. Experimental Section
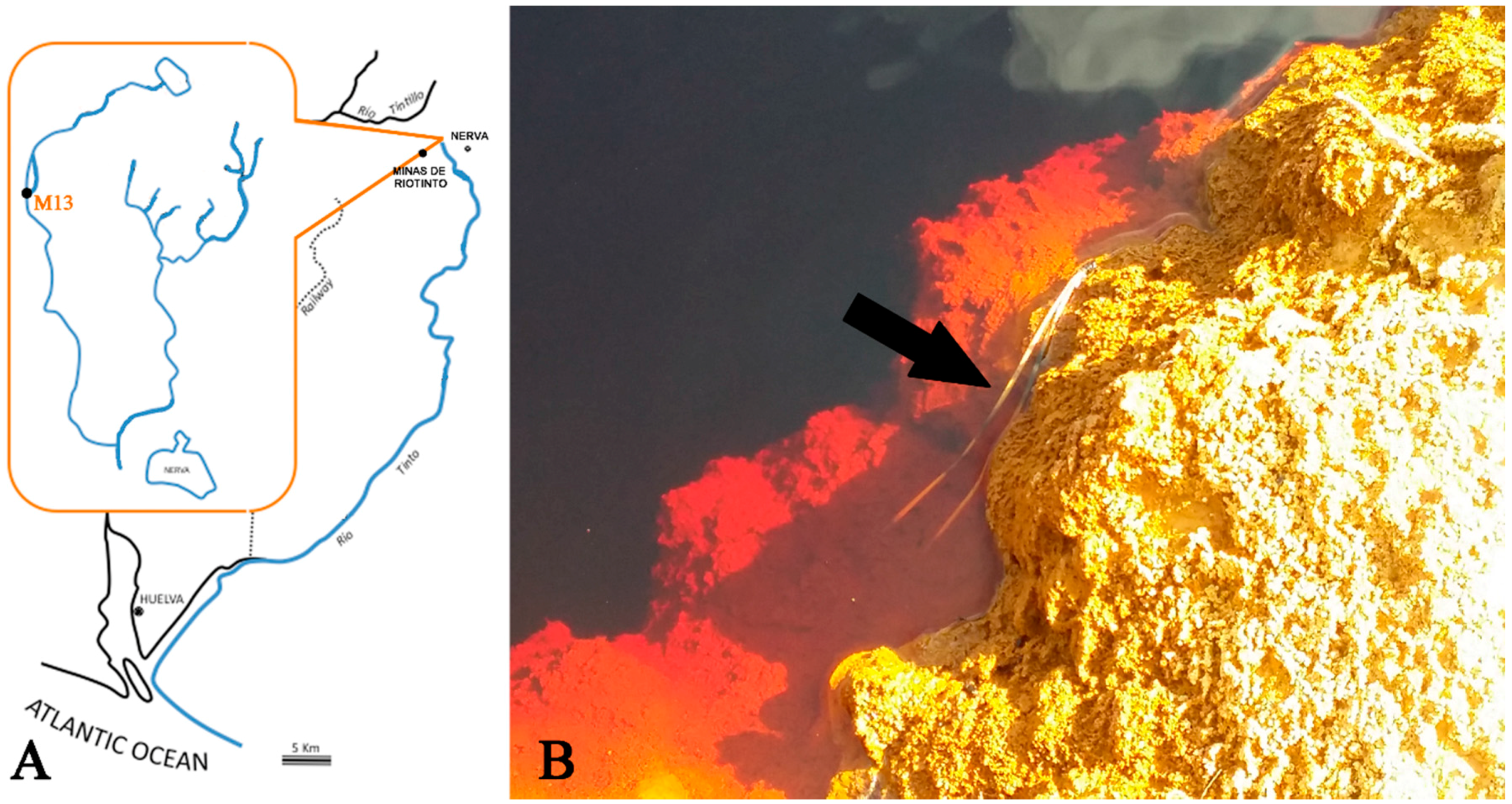
2.1. Mineral and Water Analysis
2.1.1. TXRF and Ion Chromatography Analyses
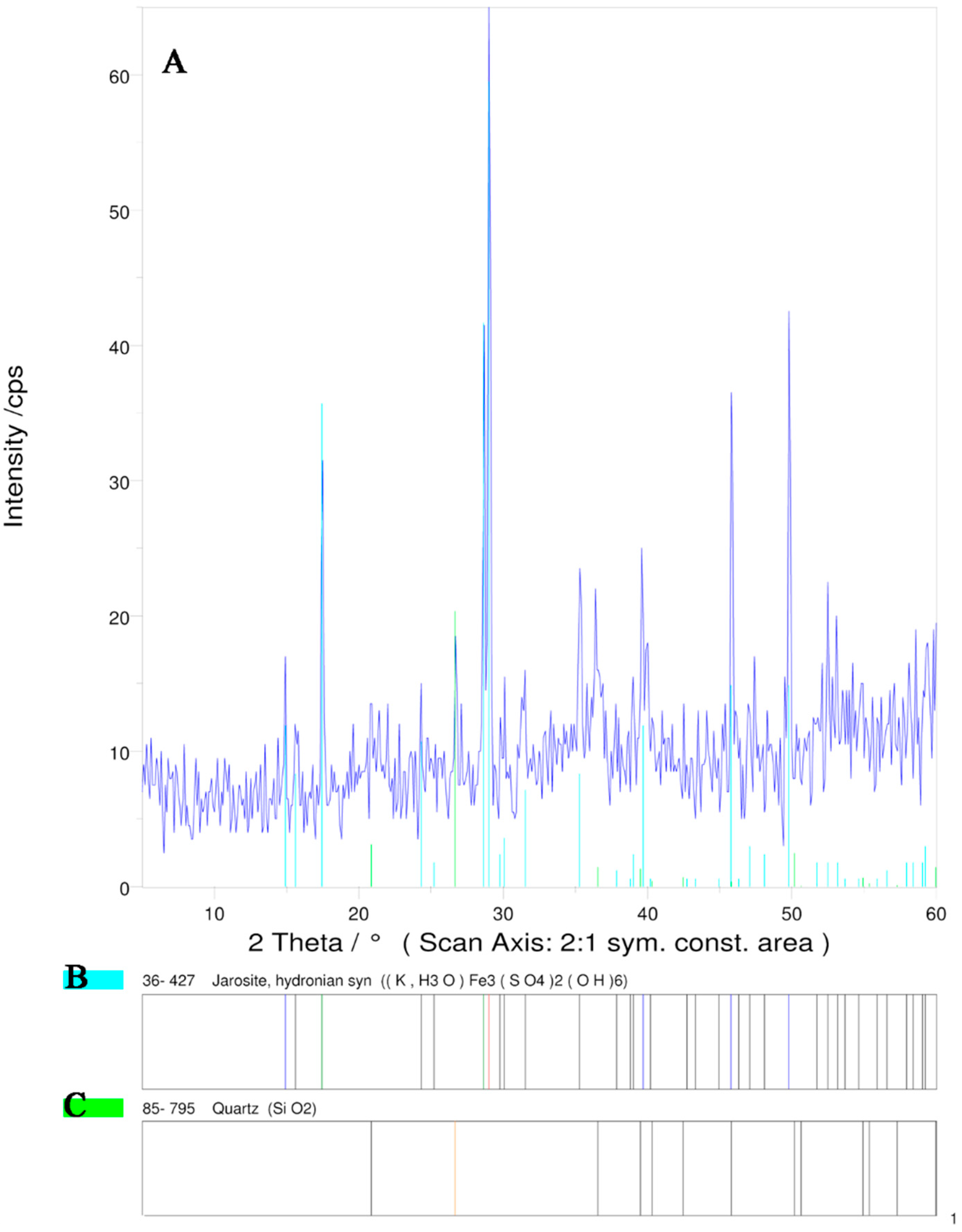
2.1.2. X-Ray Diffraction
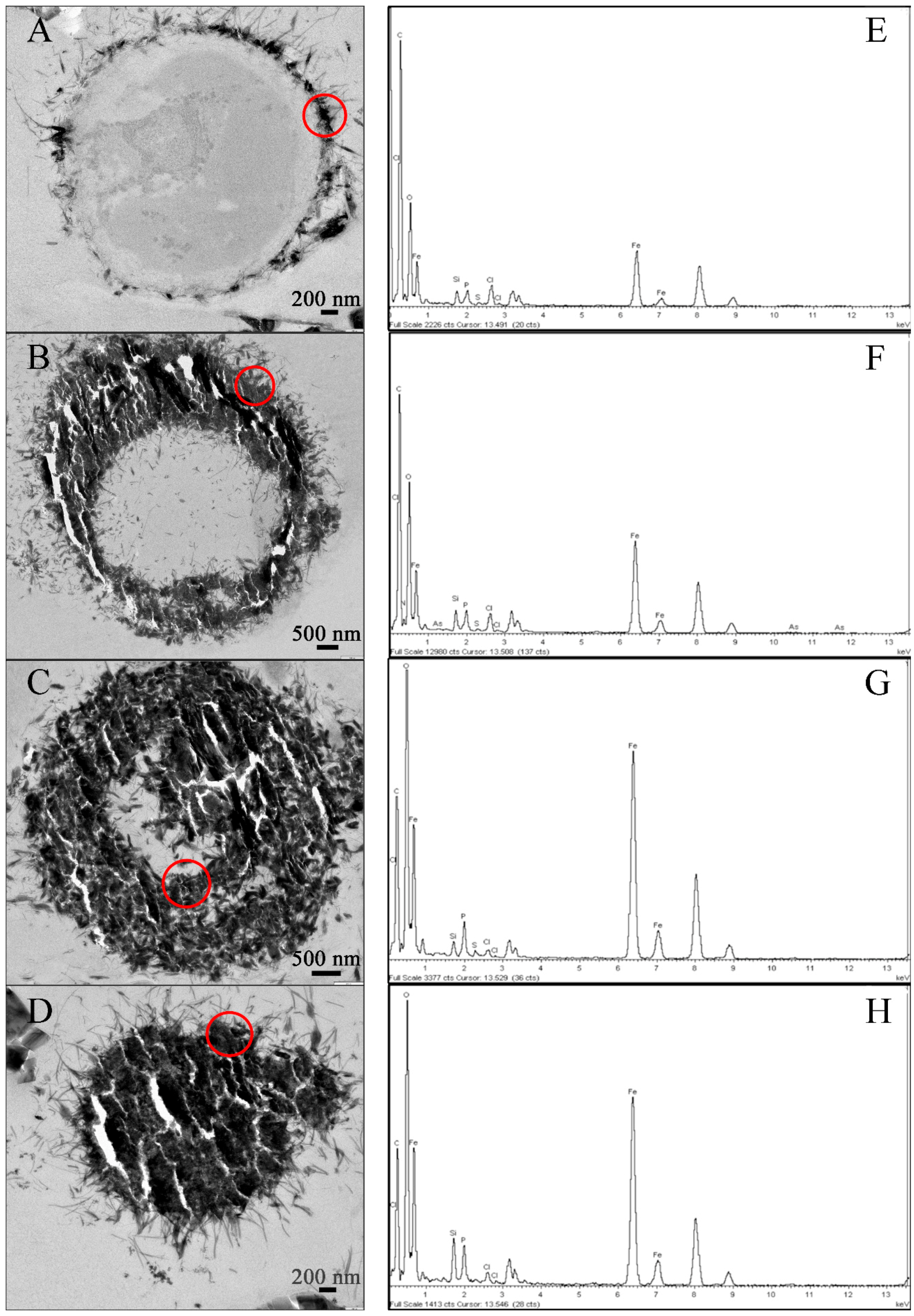
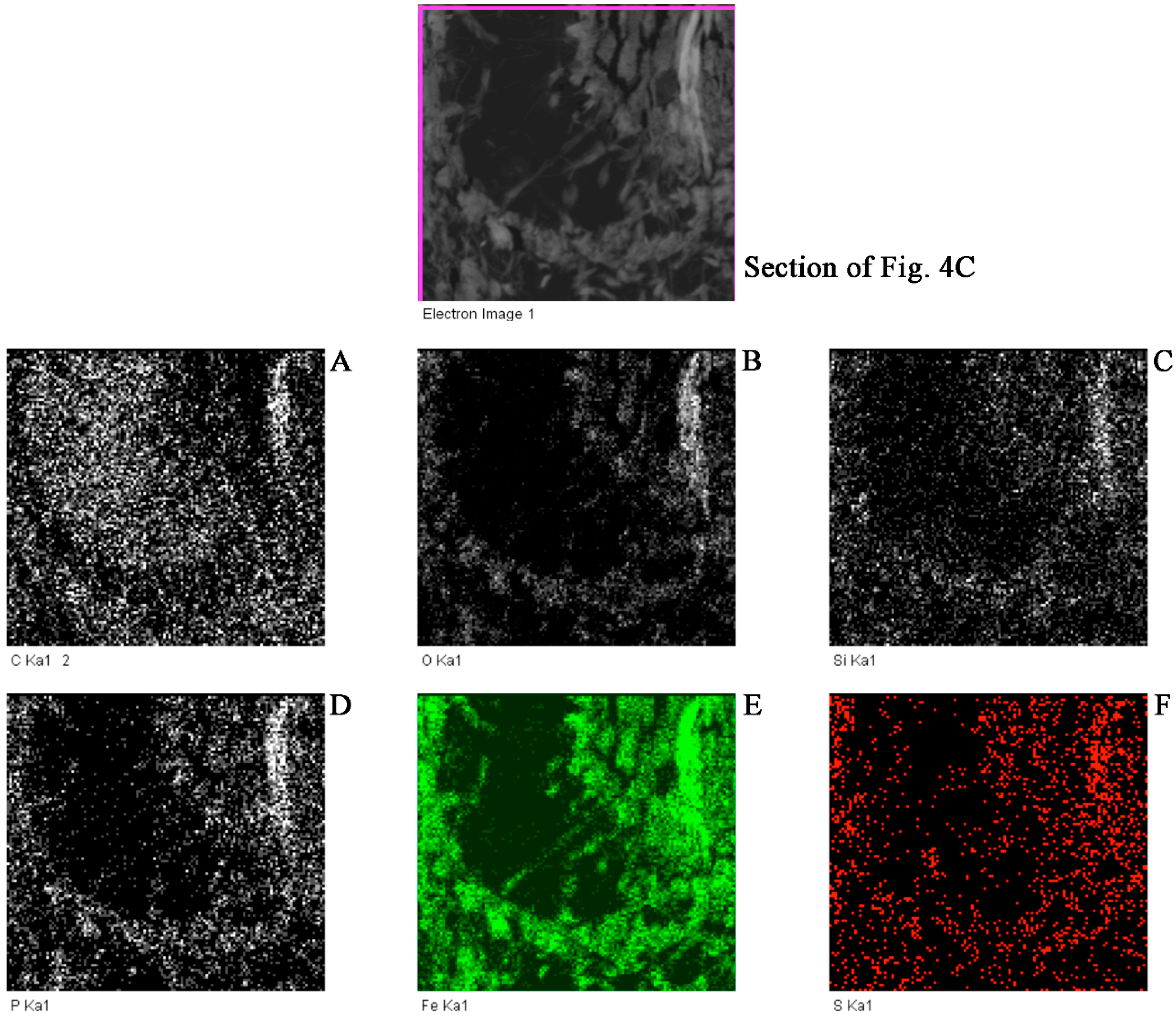
2.1.3. Transmission Electron Microscopy
2.2. Geochemical Modeling
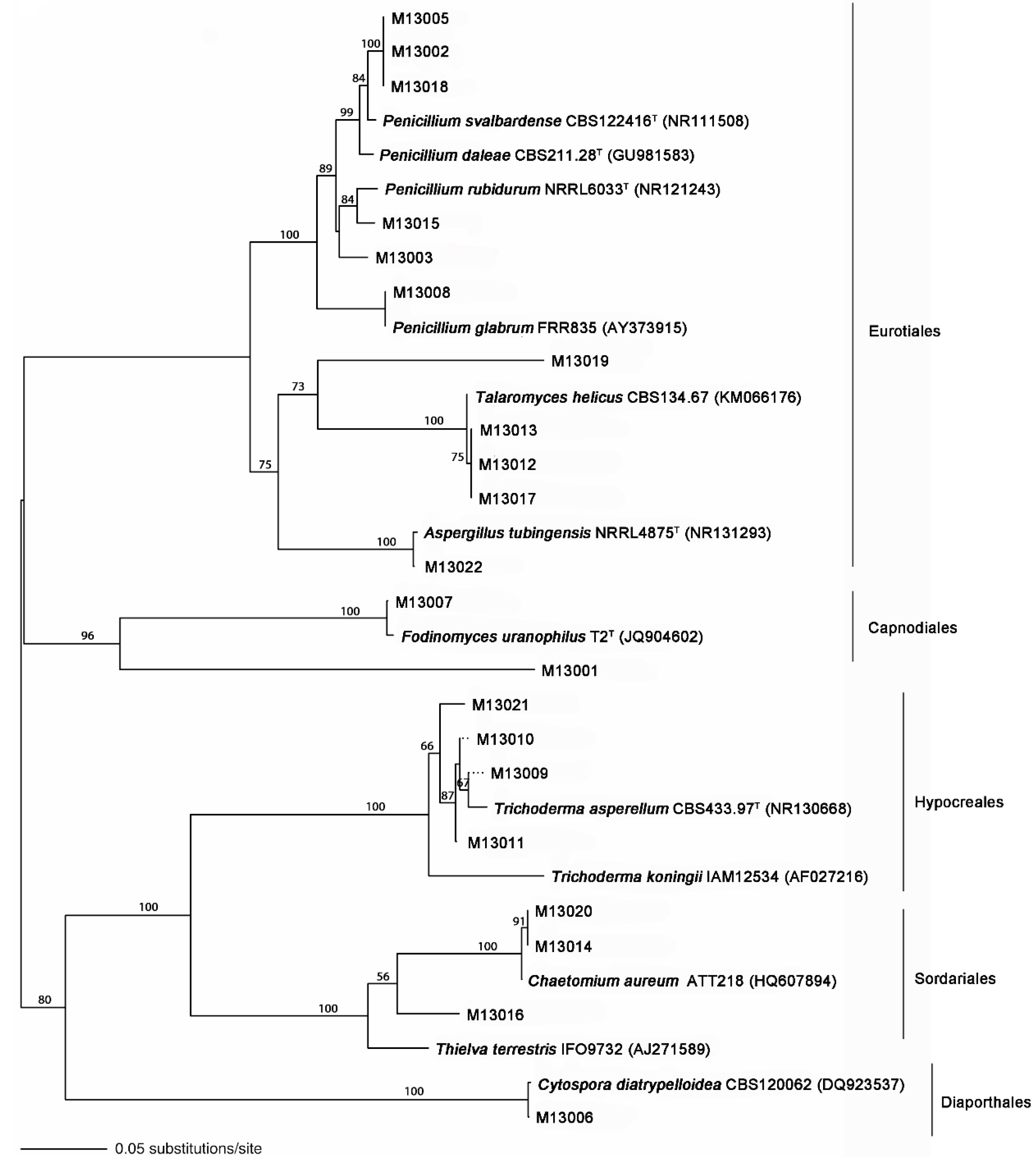
2.3. Fungal Isolation, DNA Extraction, ITS rDNA Amplification, and Sequencing
2.4. Sequence Data Analyses
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gómez-Ortiz, D.; Fernández-Remolar, C.D.; Granda, A.; Quesada, C.; Granda, T.; Prieto-Ballesteros, O.; Molina, A.; Amils, R. Identification of the subsurface sulfide bodies responsible for acidity in Río Tinto source water, Spain. Earth Planet. Sci. Lett. 2014, 391, 36–41. [Google Scholar] [CrossRef]
- Amils, R.; González-Toril, E.; Fernández-Remolar, D.; Gómez, F.; Rodríguez, N.; Durán, C. Interaction of the sulfur and iron cycles in the Tinto River ecosystem. Rev. Environ. Sci. Biotechnol. 2002, 1, 299–309. [Google Scholar] [CrossRef]
- González-Toril, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar] [CrossRef] [PubMed]
- Nordstram, D.K.; Southam, G. Geomicrobiolgy of sulfide mineral oxidation. Rev. Mineral. Geochem. 1997, 35, 361–390. [Google Scholar]
- Dutrizac, J.E.; Jambor, J.L. Jarosites and their application in hydrometallurgy. Rev. Mineral. Geochem. 2000, 40, 405–453. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.; Morris, R.V.; Gruener, J.E.; Amils, R.; Knoll, A.H. The Río Tinto Basin, Spain: Mineralogy, sedimentary geobiology, and implications for interpretation of outcrop rocks at Meridiani Planum. Mars Earth Planet. Sci. Lett. 2005, 240, 149–167. [Google Scholar] [CrossRef]
- López-Archilla, A.I.; Marín, I.; Amils, R. Microbial community composition and ecology of an acidic aquatic environment: The Tinto River, Spain. Microb. Ecol. 2001, 41, 20–35. [Google Scholar] [PubMed]
- Oggerin, M.; del Moral, C.; Rodriguez, N.; Amils, R. Fungal diversity in Río Tinto (IPB) the largest natural extreme acidic ecosystem on Earth surface. In preparation.
- Amils, R.; Fernández-Remolar, D.; The IPBSL Team. Río Tinto: A geochemical and mineralogical terrestrial analogue of Mars. Life 2014, 4, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.T.; Hall, I.R. Metal tolerance in fungi. In Heavy Metal Tolerance in Plants: Evolutionary Aspects; Shaw, J., Ed.; CRC Press: Boca Raton, FL, USA, 1990; pp. 95–104. [Google Scholar]
- Gadd, G.M. Bacterial and fungal geomicrobiology: A problem with communities? Geobiology 2008, 6, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Lowestam, H.A. Mineral formed by organisms. Science 1981, 211, 1126–1131. [Google Scholar] [CrossRef]
- Beveridge, T.J. Role of cellular design in bacterial metal accumulation and ineralization. Annu. Rev. Microbiol. 1989, 43, 147–171. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A. Bacterial mineralization. In Encyclopedia of Materials: Science and Technology; Buschow, K.H.J., Cahn, R.W., Flemings, M.C., Ilschner, B., Kramer, E.J., Mahajan, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 441–448. [Google Scholar]
- Konhauser, K.O. Diversity of bacterial iron mineralization. Earth Sci. Rev. 1998, 43, 91–121. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Murray, R.G.E. Uptake and retention of metals by cell walls of Bacillus subtilis. J. Bacterial. 1976, 127, 1502–1518. [Google Scholar]
- Urrutia, M.M.; Kemper, M.; Doyle, R.; Beveridge, T.J. The membrane-induced proton motive force influences the metal binding ability of Bacillus subtilis cell walls. Appl. Environ. Microbiol. 1992, 58, 3837–3844. [Google Scholar]
- Gadd, G.M.; Rhee, Y.J.; Stephenson, K.; Wei, Z. Geomycology: Metals, actinides and biominerals. Environ. Microbiol. Rep. 2012, 4, 270–296. [Google Scholar] [CrossRef] [PubMed]
- Schwertmann, U.; Cornell, R.M. General preparative techniques. In Iron Oxides in the Laboratory: Preparation and Characterization, 2nd ed.; Wiley VCH Verlag: Weinheim, Germany, 2007; pp. 19–25. [Google Scholar]
- Martín, J.D. Using XPowder—A Sofware Package for Powder X-Ray Diffraction Analysis, 2004; D.L.GR-1001/04, ISBN: 84-609-1497-6, 105p. Spain. Available online: http:// www.xpowder.com/ (accessed on 1 January 2010).
- Oggerin, M.; Tornos, F.; Rodríguez, N.; del Moral, C.; Sánchez-Román, M.; Amils, R. Specific jarosite biomineralization by Purpureocillium lilacinum, an acidophilic fungi isolated from Río Tinto. Environ. Microbiol. 2013, 15, 2228–2237. [Google Scholar] [CrossRef] [PubMed]
- Bethke, C.M. Geochemical and Biogeochemical Modeling; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Wolery, T.J. EQ3/6, a Software Package for Geochemical Modeling of Aqueous Systems: Package Overview and Installation Guide (Version 7.0); Lawrence Livermore National Laboratory: Livermore, CA, USA, 1992; p. 66. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Higgins, D.G. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Se-Al Carbon Software, 1996. Available online: http://tree.bio.ed.ac.uk/software/seal/ (accessed on 1 January 2005).
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (* and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tornos, F. Environment of formation and styles of volcanogenic massive sulfides: The Iberian Pyrite Belt. Ore Geol. Rev. 2006, 28, 259–307. [Google Scholar] [CrossRef]
- Wei, Z.; Liang, X.; Pendlowski, H.; Hillier, S.; Suntornvongsagul, K.; Sihanonth, P.; Gadd, G.M. Fungal biotransformation of zinc silicate and sulfide mineral ores. Environ. Microbiol. 2013, 15, 2173–2186. [Google Scholar] [CrossRef] [PubMed]
- Sterflinger, K. Fungi as geologic agents. Geomicrobiol. J. 2000, 17, 97–124. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomycology: Biogeochemical transfor-mations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef] [PubMed]
- Gueidan, C.; Ruibal, C.; de Hoog, G.S.; Schneider, H. Rock-inhabiting fungi originated during periods of dry climate in the late Devonian and middle Triassic. Fungal Biol. 2011, 115, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Gorbushina, A.A.; Kotlova, E.R.; Sherstneva, O.A. Cellular responses of microcolonial rock fungi to long-term desiccation and subsequent rehydration. Stud. Mycol. 2008, 61, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Burford, E.P.; Fomina, M.; Gadd, G.M. Fungal involvement in bioweathering and biotransformation of rocks and minerals. Mineral. Mag. 2003, 67, 1127–1155. [Google Scholar] [CrossRef]
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediaton. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Gomez, F.; Zettler, E.; Keenan, B.G.; Amils, R.; Sogin, M.L. Eukaryotic diversity in Spain’s River of Fire. Nature 2002, 417, 137. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.J.; Lutz, M.A.; Dawson, S.C.; Bond, P.L.; Banfield, J.F. Metabolically active eukaryotes in extremely acidic mine drainage. Appl. Environ. Microbiol. 2004, 70, 6264–6271. [Google Scholar] [CrossRef] [PubMed]
- Chaput, D.L.; Hansel, C.M.; Burgos, W.D.; Santelli, C.M. Profiling microbial communities in manganese remediation systems treating coal mine drainage. Appl. Environ. Microbiol. 2015, 81, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Cosmidis, J.; Benzerara, K.; Morin, G.; Busigny, V.; Lebeau, O.; Jézéquel, D.; Noël, V.; Dublet, G.; Othmane, G. Biomineralization of iron-phosphates in the water column of Lake Pavin (Massif Central, France). Geochim. Coschim. Acta 2014, 126, 78–96. [Google Scholar] [CrossRef]
- Schwertmann, U.; Cornell, R.M. Goethite. In Iron Oxides in the Laboratory: Preparation and Characterization, 2nd ed.; Wiley VCH Verlag: Weinheim, Germany, 2007; pp. 67–92. [Google Scholar]
- Schwertmann, U.; Cornell, R.M. Lepidocrocite. In Iron Oxides in the Laboratory: Preparation and Characterization, 2nd ed.; Wiley VCH Verlag: Weinheim, Germany, 2007; pp. 93–97. [Google Scholar]
- Bigham, J.M.; Nordstroma, D.K. Iron and aluminium hydroxysulfates from acid sulfate waters. Rev. Mineral. Geochem. 2000, 40, 351–403. [Google Scholar] [CrossRef]
- Fortin, D.; Ferris, F.G.; Beveridge, T.J. Surface mediated mineral development by bacteria. In Geomicrobiology: Interactions between Microbes and Minerals, Reviews in Mineralogy; Banfield, J.F., Nealson, K.H., Eds.; Mineralogical Society of America: Washington, DC, USA, 1997; Volume 35, p. 161. [Google Scholar]
- Mann, S. Molecular recognition in biomineralization. Nature 1988, 332, 119–124. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Fyfe, W.S. Metal fixation by bacterial cell walls. Can. J. Earth Sci. 1985, 22, 1893–1898. [Google Scholar] [CrossRef]
- Siegel, S.M.; Galun, M.; Siegel, B.Z. Filamentous fungi as metal biosorbents: A review. Water Air Soil Pollut. 1990, 53, 335–344. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | pH | Eh (mV) | Cond. (mS/cm2) | T (°C) | K (mg/L) | Fe2+ (mg/L) | Fe3+ (mg/L) | SO42− (mg/L) | Cu (mg/L) | As (mg/L) | Co (mg/L) | Si (mg/L) | Al (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M13 | 2.7 | 456.4 | 5.5 | 17.1 | 2.0 | 228.6 | 960.3 | 1742.5 | 13.5 | 1.5 | 1.3 | 1269.9 | 299.4 |
| Mineral Phase | SI |
|---|---|
| Nontronite-Ca | 10.0 |
| Clinoptile-Ca | 7.5 |
| Quartz | 2.7 |
| Barite | 1.6 |
| CuFeO2(c) | 1.1 |
| Beidellite | 0.5 |
| Goethite | 0.16 |
| Hematite | 0.12 |
| Jarosite-K | −0.48 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oggerin, M.; Tornos, F.; Rodriguez, N.; Pascual, L.; Amils, R. Fungal Iron Biomineralization in Río Tinto. Minerals 2016, 6, 37. https://doi.org/10.3390/min6020037
Oggerin M, Tornos F, Rodriguez N, Pascual L, Amils R. Fungal Iron Biomineralization in Río Tinto. Minerals. 2016; 6(2):37. https://doi.org/10.3390/min6020037
Chicago/Turabian StyleOggerin, Monike, Fernando Tornos, Nuria Rodriguez, Laura Pascual, and Ricardo Amils. 2016. "Fungal Iron Biomineralization in Río Tinto" Minerals 6, no. 2: 37. https://doi.org/10.3390/min6020037