
Bioactivity and Mycochemical Profile of Extracts from Mycelial Cultures of Ganoderma spp.

 ,
,  , , ,
, , ,  ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Mycelial Cultures
2.2. Mycochemical Analyses
2.3. Antioxidant Activity
2.4. Inhibition of Cholinesterase and Amylase
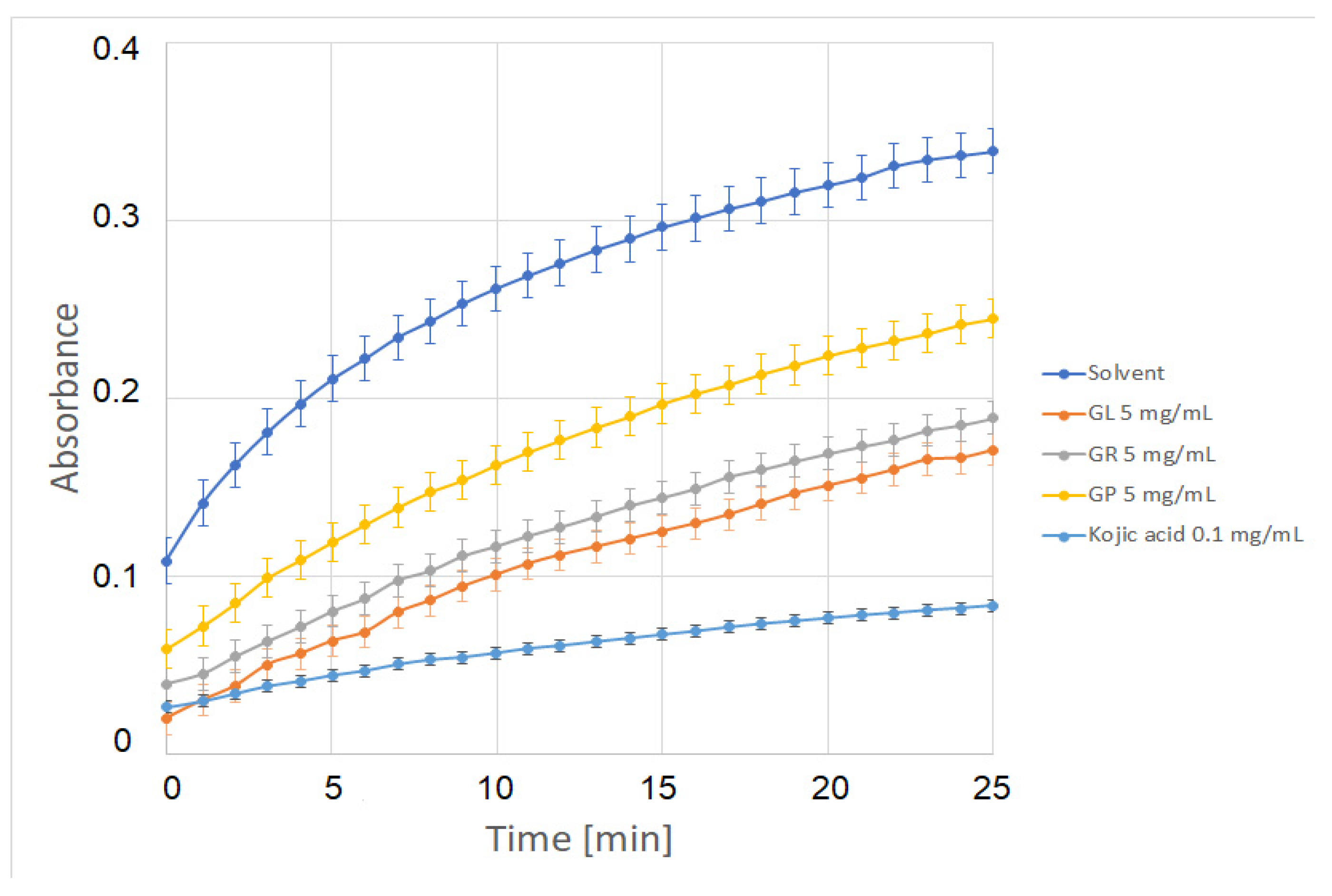
2.5. Tyrosinase Inhibition Assay and Tests in Mouse Melanoma B16F10 Cells
3. Materials and Methods
3.1. Origin of Mycelial Cultures
3.2. Initial Mycelial Cultures
3.3. Experimental Mycelial Cultures
3.4. Extraction of Biomass
3.5. Mycochemical Analysis
3.6. TPC Evaluation
3.7. TFC Evaluation
3.8. DPPH Radical Scavenging Assay
3.9. ABTS Assay
3.10. FRAP Assay
3.11. CUPRAC Assay
3.12. PHD Assay
3.13. MCA Assay
3.14. α-Amylase Inhibitory Activity
3.15. Cholinesterase Inhibitory Activity
3.16. Tyrosinase Inhibition Assay
3.17. In Vitro Viability Assessment by the MTT Assay
3.18. In Vitro Melanin Production Assay
3.19. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Karsten, P. Enumeratio Boletinarum et Polyporarum Fennicarum systemate novo dispositorum. Rev. Mycol. 1881, 3, 16–18. [Google Scholar]
- Wang, L.; Li, J.Q.; Zhang, J.; Li, Z.M.; Liu, H.G.; Wang, Y.Z. Traditional uses, chemical components and pharmacological activities of the genus Ganoderma P. Karst.: A review. RSC Adv. 2020, 10, 42084–42097. [Google Scholar] [CrossRef]
- Basnet, B.B.; Liu, L.; Bao, L.; Liu, H. Current and future perspective on antimicrobial and anti-parasitic activities of Ganoderma sp.: An update. Mycology 2017, 8, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonathan, S.G.; Awotona, F.E. Studies on antimicrobial potentials of three Ganoderma species. Afr. J. Biomed. Res. 2010, 13, 133–139. [Google Scholar]
- Xia, Q.; Zhang, H.; Sun, X.; Zhao, H.; Wu, L.; Zhu, D.; Yang, G.; Shao, Y.; Zhang, X.; Mao, X.; et al. A comprehensive review of the structure elucidation and biological activity of triterpenoids from Ganoderma spp. Molecules 2014, 19, 17478–17535. [Google Scholar] [CrossRef] [PubMed]
- Shiao, M.S. Natural products of the medicinal fungus Ganoderma lucidum: Occurrence, biological activities, and pharmacological functions. Chem. Rec. 2003, 3, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Ansor, N.M.; Abdullah, N.; Aminudin, N. Anti-angiotensin converting enzyme (ACE) proteins from mycelia of Ganoderma lucidum (Curtis) P. Karst. BMC Complement Altern. Med. 2013, 13, 1–8. [Google Scholar]
- Hirotani, M.; Ino, C.; Furuya, T.; Shiro, M. Ganoderic acids T, S and R, new triterpenoids from the cultured mycelia of Ganoderma lucidum. Chem. Pharm. Bull. 1986, 345, 2282–2285. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, P.A.; Seviour, R.J.; Schmid, F. Growth of filamentous fungi in submerged culture: Problems and possible solutions. Crit. Rev. Biotechnol. 2000, 20, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Mooralitharan, S.; Hanafiah, Z.M.; Manan, T.S.B.A.; Hasan, H.A.; Jensen, H.S.; Wan-Mohtar, W.A.A.Q.I.; Mohtar, W.H.M.W. Optimization of mycoremediation treatment for the chemical oxygen demand (COD) and ammonia nitrogen (AN) removal from domestic effluent using wild-Serbian Ganoderma lucidum (WSGL). Environ. Sci. Pollut. Res. 2021, 28, 32528–32544. [Google Scholar] [CrossRef]
- Yue, G.G.L.; Fung, K.P.; Tse, G.M.K.; Leung, P.C.; Lau, C.B.S. Comparative studies of various Ganoderma species and their different parts with regard to their antitumor and immunomodulating activities in vitro. J. Altern. Complement. Med. 2006, 12, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Sułkowska-Ziaja, K.; Szewczyk, A.; Galanty, A.; Gdula-Argasińska, J.; Muszyńska, B. Chemical composition and biological activity of extracts from fruiting bodies and mycelial cultures of Fomitopsis betulina. Mol. Biol. Rep. 2018, 45, 2535–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fijałkowska, A.; Muszyńska, B.; Sułkowska-Ziaja, K.; Kała, K.; Pawlik, A.; Stefaniuk, D.; Matuszewska, A.; Piska, K.; Pękala, E.; Kaczmarczyk, P.; et al. Medicinal potential of mycelium and fruiting bodies of an arboreal mushroom Fomitopsis officinalis in therapy of lifestyle diseases. Sci. Rep. 2020, 10, 20081. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, S.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Khadem, S.; Marles, R.J. Monocyclic phenolic acids; hydroxy– and polyhydroxybenzoic acids: Occurrence and recent bioactivity studies. Molecules 2010, 15, 7985–8005. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Jovin, E.; Malbaša, R.; Matavuly, M.; Popović, M. Medicinal and edible lignicolous fungi as natural sources of antioxidative and antibacterial agents. Phytother. Res. 2010, 24, 1473–1481. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Beara, I.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-inflammatory, antioxidant and enzyme inhibition activities in correlation with mycochemical profile of selected indigenous Ganoderma spp. from Balkan Region (Serbia). Chem. Biodivers. 2021, 18, e2000828. [Google Scholar] [CrossRef]
- Wang, S.Y.; Shi, X.C.; Laborda, P. Indole-based melatonin analogues: Synthetic approaches and biological activity. Review. Eur. J. Med. Chem. 2020, 185, 111847. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Sułkowska-Ziaja, K. Impact of food processing on non-hallucinogenic indole derivatives in edible mushrooms. In Processing and Impact on Active Components in Food; Elsevier/ Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2015. [Google Scholar]
- Sułkowska-Ziaja, K.; Maślanka, A.; Szewczyk, A.; Muszyńska, B. Physiologically active compounds in four species of Phellinus. Nat. Prod. Commun. 2017, 12, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Barreira, J.C.M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Development of a Novel Methodology for the Analysis of Ergosterol in Mushrooms. Food Anal. Methods 2014, 7, 217–223. [Google Scholar] [CrossRef]
- Rosecke, J.; Konig, W.A. Constituents of various wood-rotting basidiomycetes. Phytochemistry 2000, 54, 603–610. [Google Scholar] [CrossRef]
- Lv, G.P.; Zhao, J.; Duan, J.A.; Tang, Y.P.; Li, S.P. Comparison of sterols and fatty acids in two species of Ganoderma. Chem. Cent. J. 2012, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.; Grünvald, P.; Ladányi, M.; Papp, V.; Papp, I.; Némedi, E.; Mirmazloum, I. Heat treatment of Reishi medicinal mushroom (Ganoderma lingzhi) basidiocarp enhanced its β-glucan solubility, antioxidant capacity and lactogenic properties. Foods 2021, 10, 2015. [Google Scholar] [CrossRef]
- Tan, E.S.S.; Leo, T.K.; Tan, C.K. Effect of tiger milk mushroom (Lignosus rhinocerus) supplementation on respiratory health, immunity and antioxidant status: An open-label prospective study. Sci. Rep. 2021, 11, 1–10. [Google Scholar]
- Zengin, G.; Sarikurkcu, C.; Gunes, E.; Uysal, A.; Ceylan, R.; Uysal, S.; Gungor, H.; Aktumsek, A. Two Ganoderma species: Profiling of phenolic compounds by HPLC–DAD, antioxidant, antimicrobial and inhibitory activities on key enzymes linked to diabetes mellitus, Alzheimer’s disease and skin disorders. Food Funct. 2015, 6, 2794–2802. [Google Scholar] [CrossRef] [PubMed]
- Saltarelli, R.; Palma, F.; Gioacchini, A.M.; Calcabrini, C.; Mancini, U.; De Bellis, R.; Stocchi, V.; Potenza, L. Phytochemical composition, antioxidant and antiproliferative activities and effects on nuclear DNA of ethanolic extract from an Italian mycelial isolate of Ganoderma lucidum. J. Ethnopharmacol. 2019, 231, 464–473. [Google Scholar] [CrossRef]
- Sharpe, E.; Farragher-Gnadt, A.P.; Igbanugo, M.; Huber, T.; Michelotti, J.C.; Milenkowic, A.; Ludlam, S.; Walker, M.; Hanes, D.; Bradley, R.; et al. Comparison of antioxidant activity and extraction techniques for commercially and laboratory prepared extracts from six mushroom species. J. Sci. Food Agric. 2021, 4, 100130. [Google Scholar] [CrossRef]
- Cho, J.Y.; Sadiq, N.B.; Kim, J.C.; Lee, B.; Hamayun, M.; Lee, T.S.; Kim, H.Y. Optimization of antioxidant, anti-diabetic, and anti-inflammatory activities and ganoderic acid content of differentially dried Ganoderma lucidum using response surface methodology. Food Chem. 2021, 335, 127645. [Google Scholar]
- Heleno, S.A.; Barros, L.; Martins, A.; Queiroz, M.J.R.; Santos-Buelga, C.; Ferreira, I.C.F.R. Fruiting body, spores and in vitro produced mycelium of Ganoderma lucidum from Northeast Portugal: A comparative study of the antioxidant potential of phenolic and polysaccharidic extracts. Food Res. Int. 2012, 46, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Tel-Çayan, G.; Öztürk, M.; Duru, M.E.; Rehman, M.U.; Adhikari, A.; Türkoğlu, A.; Choudhary, M.I. Phytochemical investigation, antioxidant and anticholinesterase activities of Ganoderma adspersum. Ind Crops Prod. 2015, 76, 749–754. [Google Scholar] [CrossRef]
- Taofiq, O.; Heleno, S.A.; Calhelha, R.C.; Alves, M.J.; Barros, L.; González-Paramás, A.M.; Barreiro, M.F.; Ferreira, I.C.F.R. The potential of Ganoderma lucidum extracts as bioactive ingredients in topical formulations, beyond its nutritional benefits. Food Chem. Toxicol. 2017, 108, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Saltarelli, R.; Ceccaroli, P.; Buffalini, M.; Vallorani, L.; Casadei, L.; Zambonelli, A.; Stocchi, V.; Iotti, M.; Badalyan, S.; Stocchi, V. Biochemical characterization and antioxidant and antiproliferative activities of different Ganoderma collections. J. Mol. Microbiol. 2015, 25, 16–25. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.; Sharma, A.; Tuli, H.S.; Batra, P.; Beniwal, V.; Gupta, G.K.; Sharma, A.K. Bioactive metabolites of Ganoderma lucidum: Factors, mechanism and broad spectrum therapeutic potential. J. Herb. Med. 2019, 17, 100268. [Google Scholar] [CrossRef]
- Wei, J.C.; Wang, A.H.; Wei, Y.L.; Huo, X.K.; Tian, X.G.; Feng, L.; Ma, X.C.; Wang, C.; Huang, S.S.; Jia, J.M. Chemical characteristics of the fungus Ganoderma lucidum and their inhibitory effects on acetylcholinesterase. J. Asian Nat. Prod. Res. 2018, 20, 992–1001. [Google Scholar] [CrossRef]
- Kaur, A.; Randhawa, K.; Singh, V.; Shri, R. Bioactivity-guided isolation of acetylcholinesterase inhibitor from Ganoderma mediosinense (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 755–763. [Google Scholar] [CrossRef]
- Deveci, E.; Çayan, F.; Tel-Çayan, G.; Duru, M.E. Inhibitory activities of medicinal mushrooms on α-amylase and α-glucosidase-enzymes related to type 2 diabetes. S. Afr. J. Bot. 2021, 137, 19–23. [Google Scholar] [CrossRef]
- Yalcin, O.U.; Sarikurkcu, C.; Cengiz, M.; Gungor, H.; Zeljković, S.C. Ganoderma carnosum and Ganoderma pfeifferi: Metal concentration, phenolic content, and biological activity. Mycologia 2020, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.D.; Yong, T.Q.; Zhang, Y.F.; Hu, H.P.; Xie, Y.Z. Inhibitory Effect of Five Ganoderma Species (Agaricomycetes) against Key Digestive Enzymes Related to Type 2 Diabetes Mellitus. Int. J. Med. Mushrooms 2019, 21, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Raman, J.; Sabaratnam, V.; Heng, C.K.; Kuppusamy, U.R. Functional properties of partially characterized polysaccharide from the medicinal mushroom Ganoderma neo-japonicum (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Long, Z.; Xue, Y.; Ning, Z.; Sun, J.; Li, J.; Su, Z.; Liu, Q.; Xu, C.; Yan, J.K. Production, characterization, and bioactivities of exopolysaccharides from the submerged culture of Ganoderma cantharelloideum MH Liu. 3 Biotech 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Joo, O.S.; Hwang, C.E.; Hong, S.Y.; Sin, E.C.; Nam, S.H.; Cho, K.M. Antioxidative and digestion enzyme inhibitory activity of Ganoderma lucidum depends on the extraction solvent. Korean J. Food Preserv. 2018, 25, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Migas, P.; Borys, I. BMD–TLC—A useful separation technique for quantitative analysis of arbutin and hydroquinone in herbal drugs. Acta Chromatogr. 2021, 24, 291–300. [Google Scholar] [CrossRef]
- Noh, J.M.; Kwak, S.Y.; Seo, H.S.; Seo, J.H.; Kim, B.G.; Lee, Y.S. Kojic acid–amino acid conjugates as tyrosinase inhibitors. Bioorganic Med. Chem. Lett. 2009, 19, 5586–5589. [Google Scholar] [CrossRef] [PubMed]
- Oddoux, L. Recherches sur les Myceliums Secondaires des Homobasidie’s en Culture; Morphologie, Cytologie, Exigences Alimentaires; Patissier: Lyon, France, 1957. [Google Scholar]
- Włodarczyk, A.; Krakowska, A.; Sułkowska-Ziaja, K.; Suchanek, M.; Zięba, P.; Opoka, W.; Muszyńska, B. Pleurotus spp. Mycelia enriched in magnesium and zinc Salts as a potential functional food. Molecules 2021, 26, 162. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Nithiyanantham, S.; Locatelli, M.; Ceylan, R.; Uysal, S.; Aktumsek, A.; Selvi, P.K.; Maskovic, P. Screening of in vitro antioxidant and enzyme inhibitory activities of different extracts from two uninvestigated wild plants: Centranthus longiflorus subsp. longiflorus and Cerinthe minor subsp. auriculata. Eur. J. Integr. Med. 2016, 8, 28–292. [Google Scholar]
- Zengin, G. A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crops. Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Saghaie, L.; Pourfarzam, M.; Fassihi, A.; Sartippour, B. Synthesis and tyrosinase inhibitory properties of some novel derivatives of kojic acid. Res. Pharm. Sci. 2013, 8, 233–242. [Google Scholar]
- Bae, S.J.; Ha, Y.M.; Park, Y.J.; Song, Y.M.; Ha, T.K.; Chun, P.; Moon, H.R.; Chung, H.Y. Design, synthesis, and evaluation of (E)-N-substituted benzylidene-aniline derivatives as tyrosinase inhibitors. Eur. J. Med. Chem. 2012, 57, 383–390. [Google Scholar] [CrossRef]
- Popiół, J.; Gunia-Krzyżak, A.; Słoczyńska, K.; Koczurkiewicz, P.; Piska, K.; Wójcik-Pszczoła, K.; Żelaszczyk, D.; Krupa, A.; Żmudzki, P.; Marona, H.; et al. The involvement of xanthone and (E)-cinnamoyl chromophores for the design and synthesis of novel sunscreening agents. Int. J. Mol. 2021, 22, 34. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Chemical Compounds | Ganoderma adspersum | Ganoderma applanatum | Ganoderma carnosum | Ganoderma lucidum | Ganoderma pfeifferi | Ganoderma resinaceum |
|---|---|---|---|---|---|---|
| Phenolic acids | ||||||
| Gallic acid | 17.62 ± 0.70 | 10.55 ± 1.44 | nd | 17.90 ± 2.57 | 34.31 ± 2.04 | nd |
| Protocatechuic acid | 3.07 ± 0.23 | 0.57 ± 0.39 | 3.95 ± 1.18 | 1.57 ± 0.07 | 1.86 ± 0.06 | 1.29 ± 0.99 |
| 3.4-Dihydroxyphenylacetic acid | 38.95 ± 3.17 | 56.60 ± 2.73 | nd | 31.00 ± 1.50 | 77.37 ± 4.04 | 42.25 ± 11.52 |
| p-Hydroxybenzoic acid | 0.62 ± 0.15 | 1.83 ± 0.03 | 0.25 ± 0.08 | 0.09 ± 0.01 | nd | 2.17 ± 0.03 |
| Caffeic acid | nd | 0.60 ± 0.10 | 1.60 ± 0.07 | nd | 0.53 ± 0.06 | nd |
| Non-hallucinogenic indoles | ||||||
| L-tryptophan | 6.02 ± 1.00 | 7.31 ± 1.02 | 3.03 ± 0.2 | 10.58 ± 2.07 | 7.26 ± 1.05 | 7.12 ± 1.06 |
| Melatonin | nd | 0.02 ± 0.004 | nd | 0.98 ± 0.02 | 0.012 ± 005 | nd |
| Sterols | ||||||
| Ergosterol | 74.78 ± 1.27 | 28.15 ± 0.48 | 16.64 ± 0.69 | 28.72 ± 1.35 | 42.68 ± 2.30 | 44.26 ± 4.56 |
| Ergosterol peroxide | * | * | * | * | * | * |
| Tyrosinase inhibitors | ||||||
| Kojic acid | nd | 0.14 ± 0.09 | nd | 0.39 ± 0.52 | nd | nd |
| Assay | Ganoderma adspersum | Ganoderma applanatum | Ganoderma carnosum | Ganoderma lucidum | Ganoderma pfeifferi | Ganoderma resinaceum |
|---|---|---|---|---|---|---|
| TPC (mg GAE/g) | 10.57 ± 0.09 d | 13.99 ± 0.08 b | 16.56 ± 0.37 a | 13.54 ± 0.37 b | 12.53 ± 0.11 c | 16.36 ± 0.38 a |
| TFC (mg RE/g) | 0.21 ± 0.02 c | 0.36 ± 0.03 b | 0.22 ± 0.02 c | 0.22 ± 0.01 c | 0.21 ± 0.04 c | 0.57 ± 0.02 a |
| DPPH (mg TE/g) | 5.61 ± 0.10 c | 5.83 ± 0.18 b | 5.01 ± 0.08 e | 5.27 ± 0.04 d | 5.28 ± 0.04 d | 6.74 ± 0.01 a |
| ABTS (mg TE/g) | 10.16 ± 0.02 b | 11.29 ± 0.28 a | 9.74 ± 0.03 bc | 9.61 ± 0.35 c | 9.77 ± 0.13 bc | 11.60 ± 0.36 a |
| FRAP (mg TE/g) | 7.27 ± 0.07 c | 8.10 ± 0.05 b | 7.40 ± 0.22 c | 6.91 ± 0.05 d | 8.10 ± 0.13 b | 10.16 ± 0.08 a |
| CUPRAC (mg TE/g) | 17.50 ± 0.45 e | 23.55 ± 0.24 c | 22.94 ± 0.27 c | 20.35 ± 0.84 d | 25.28 ± 0.24 b | 31.78 ± 0.33 a |
| PHD (mmol TE/g) | 0.45 ± 0.02 e | 0.52 ± 0.02 cd | 0.53 ± 0.01 bc | 0.57 ± 0.03 ab | 0.48 ± 0.01 de | 0.61 ± 0.04 a |
| MCA (mg EDTAE/g) | 8.51 ± 0.09 a | 8.98 ± 0.10 a | 8.49 ± 0.78 a | 7.74 ± 0.96 ab | 5.20 ± 0.52 c | 6.40 ± 0.79 bc |
| Assay | Ganoderma adspersum | Ganoderma applanatum | Ganoderma carnosum | Ganoderma lucidum | Ganoderma pfeifferi | Ganoderma resinaceum |
|---|---|---|---|---|---|---|
| AChE (mg GALAE/g) | 1.22 ± 0.01 a | na | 1.19 ± 0.01 c | na | 1.20 ± 0.01 b | na |
| BChE (mg GALAE/g) | 1.09 ± 0.08 a | 1.13 ± 0.13 a | 0.70 ± 0.02 c | 0.90 ± 0.03 b | 0.60 ± 0.03 c | 0.86 ± 0.08 b |
| Amylase (mmol ACAE/g) | 0.15 ± 0.01 a | 0.14 ± 0.01 bc | 0.14 ± 0.01 c | 0.14 ± 0.01 b | 0.14 ± 0.01 c | 0.13 ± 0.01 d |
| Tested Extract or Reference | Ganoderma adspersum | Ganoderma applanatum | Ganoderma carnosum | Ganoderma lucidum | Ganoderma pfeifferi | Ganoderma resinaceum | Kojic Acid |
|---|---|---|---|---|---|---|---|
| % of Inhibition ± SD | 0 | 0 | 0 | 50.53 ± 3.23 | 29.18 ± 0.44 | 45.31 ± 0.87 | 81.59 ± 0.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sułkowska-Ziaja, K.; Zengin, G.; Gunia-Krzyżak, A.; Popiół, J.; Szewczyk, A.; Jaszek, M.; Rogalski, J.; Muszyńska, B. Bioactivity and Mycochemical Profile of Extracts from Mycelial Cultures of Ganoderma spp. Molecules 2022, 27, 275. https://doi.org/10.3390/molecules27010275
Sułkowska-Ziaja K, Zengin G, Gunia-Krzyżak A, Popiół J, Szewczyk A, Jaszek M, Rogalski J, Muszyńska B. Bioactivity and Mycochemical Profile of Extracts from Mycelial Cultures of Ganoderma spp. Molecules. 2022; 27(1):275. https://doi.org/10.3390/molecules27010275
Chicago/Turabian StyleSułkowska-Ziaja, Katarzyna, Gokhan Zengin, Agnieszka Gunia-Krzyżak, Justyna Popiół, Agnieszka Szewczyk, Magdalena Jaszek, Jerzy Rogalski, and Bożena Muszyńska. 2022. "Bioactivity and Mycochemical Profile of Extracts from Mycelial Cultures of Ganoderma spp." Molecules 27, no. 1: 275. https://doi.org/10.3390/molecules27010275