
Isolation and Identification of Antioxidative Peptide from Goose Liver Hydrolysate to Ameliorate Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes

Abstract
:1. Introduction
2. Results and Discussion
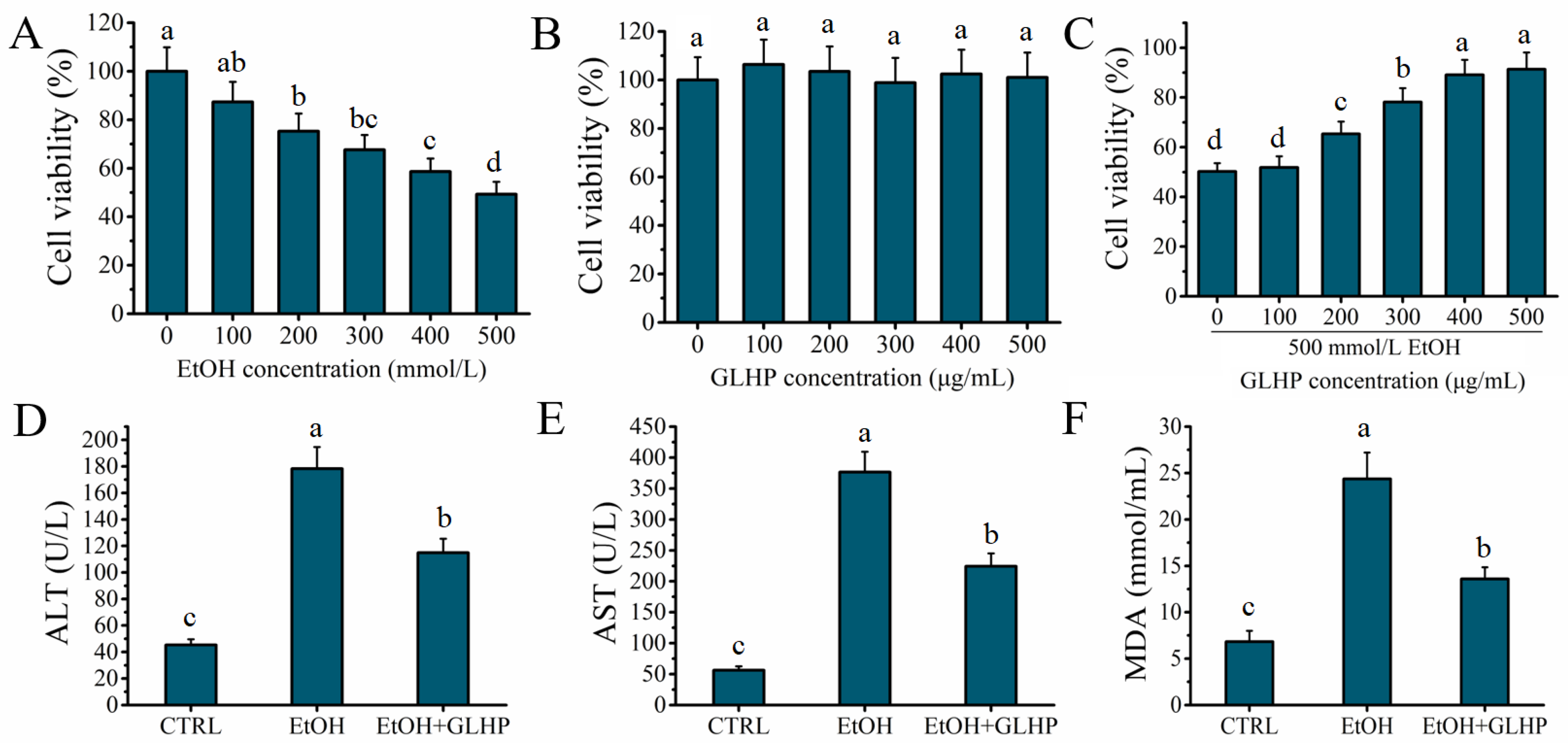
2.1. GLHP Ameliorated Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes
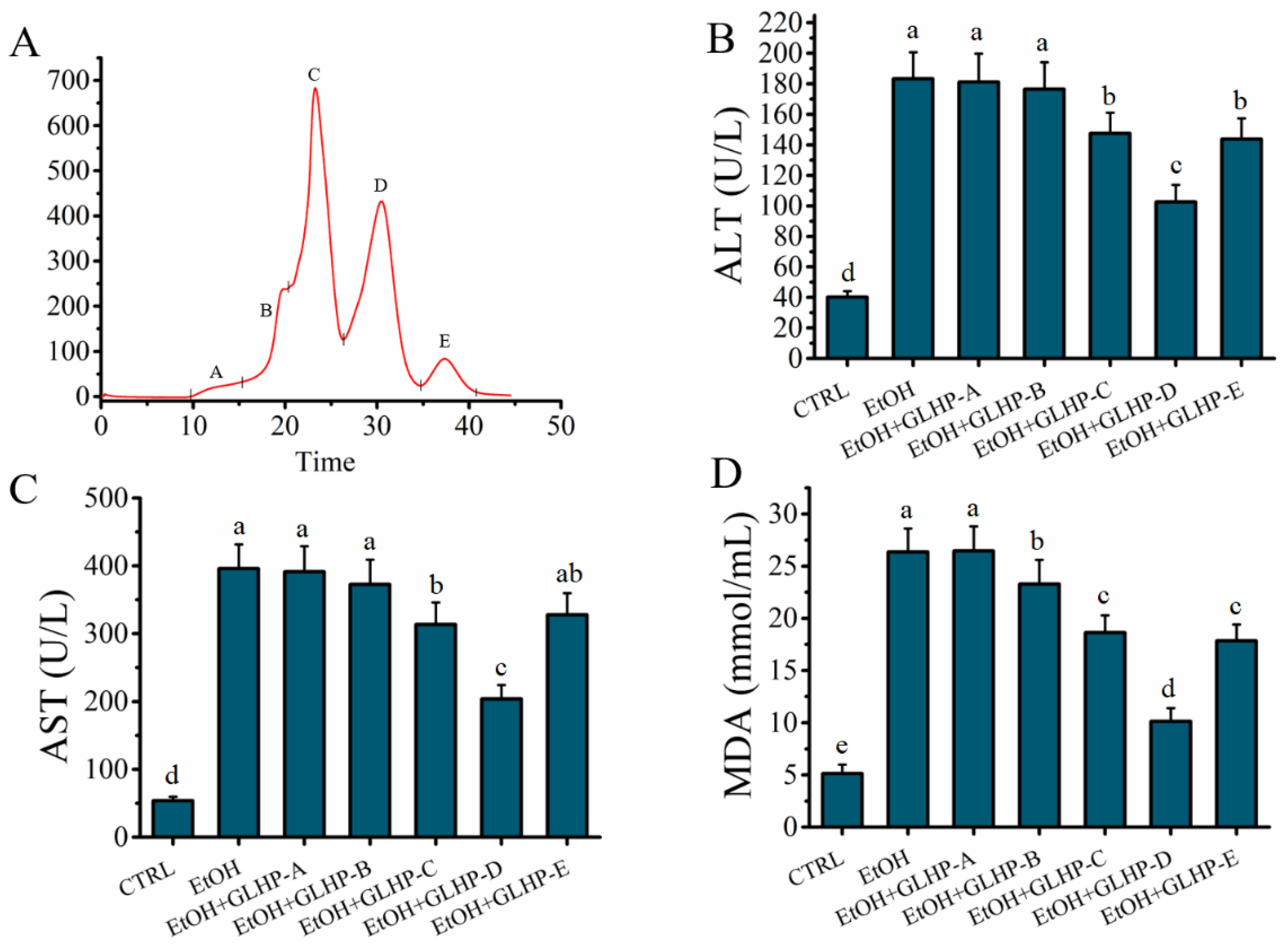
2.2. GLHP Was Separated by Size Exclusion Chromatography (SEC)
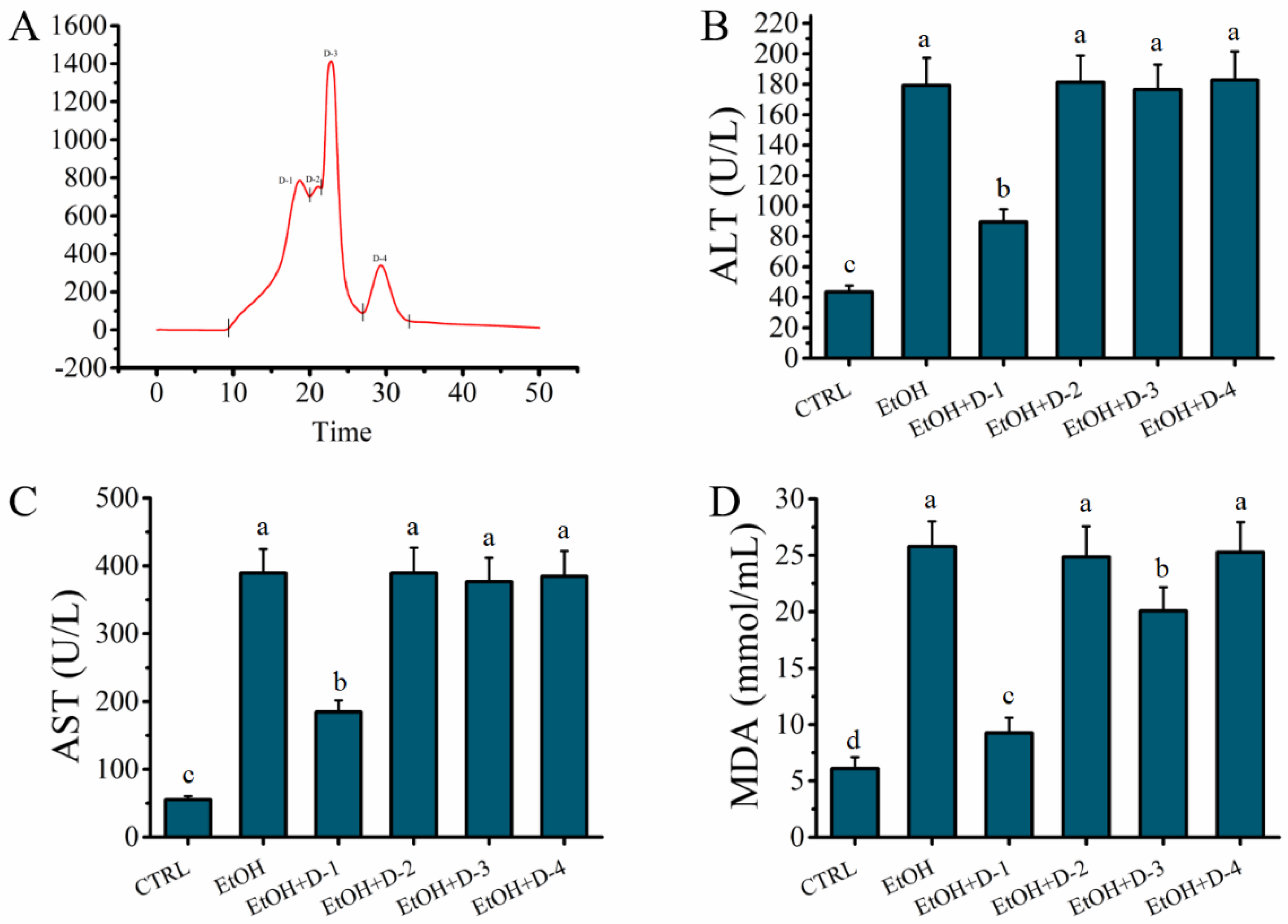
2.3. Fraction D of GLHP Was Separated by Ion Exchange Chromatography (IEC)
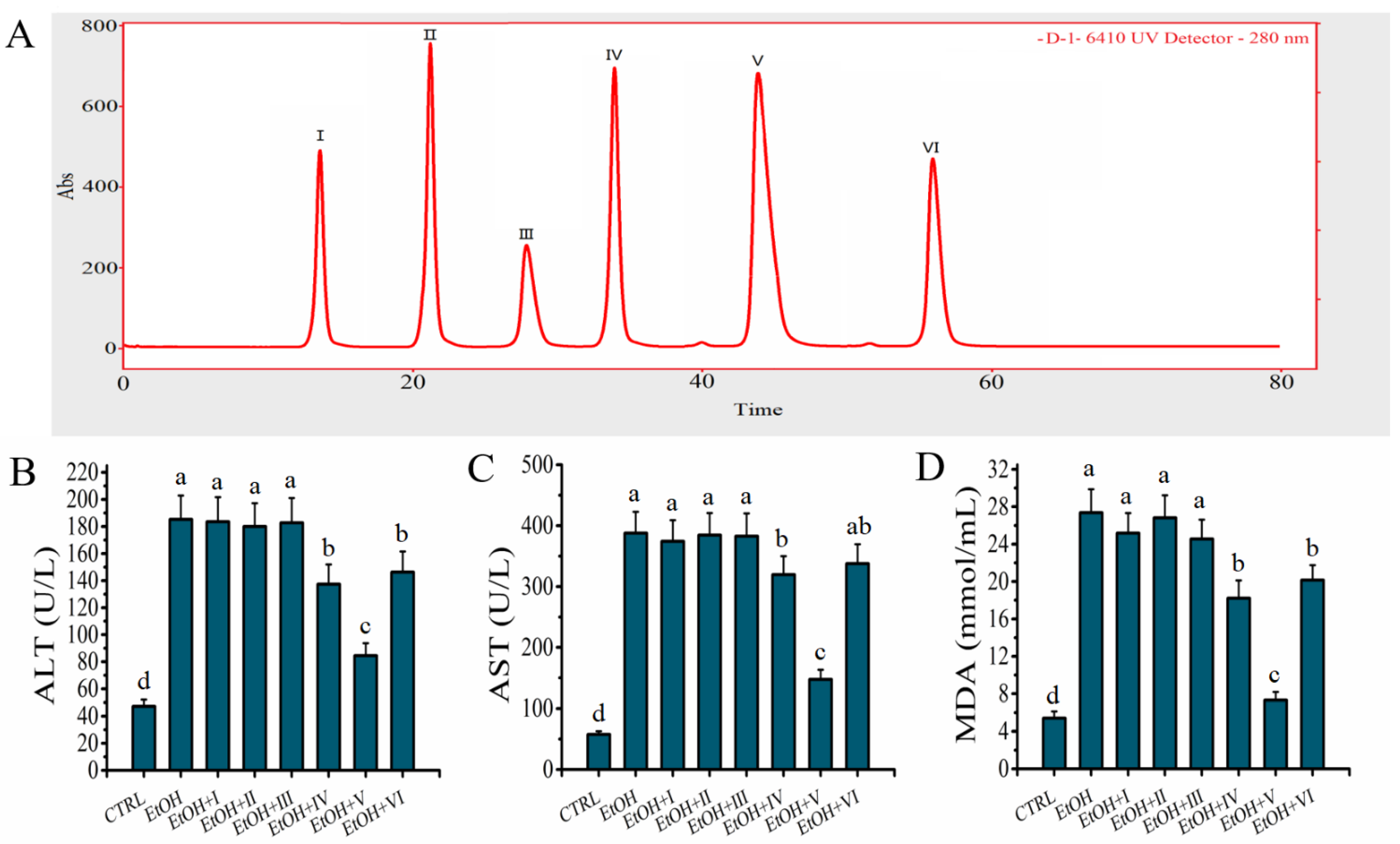
2.4. D-1 Was Separated by RP-LC
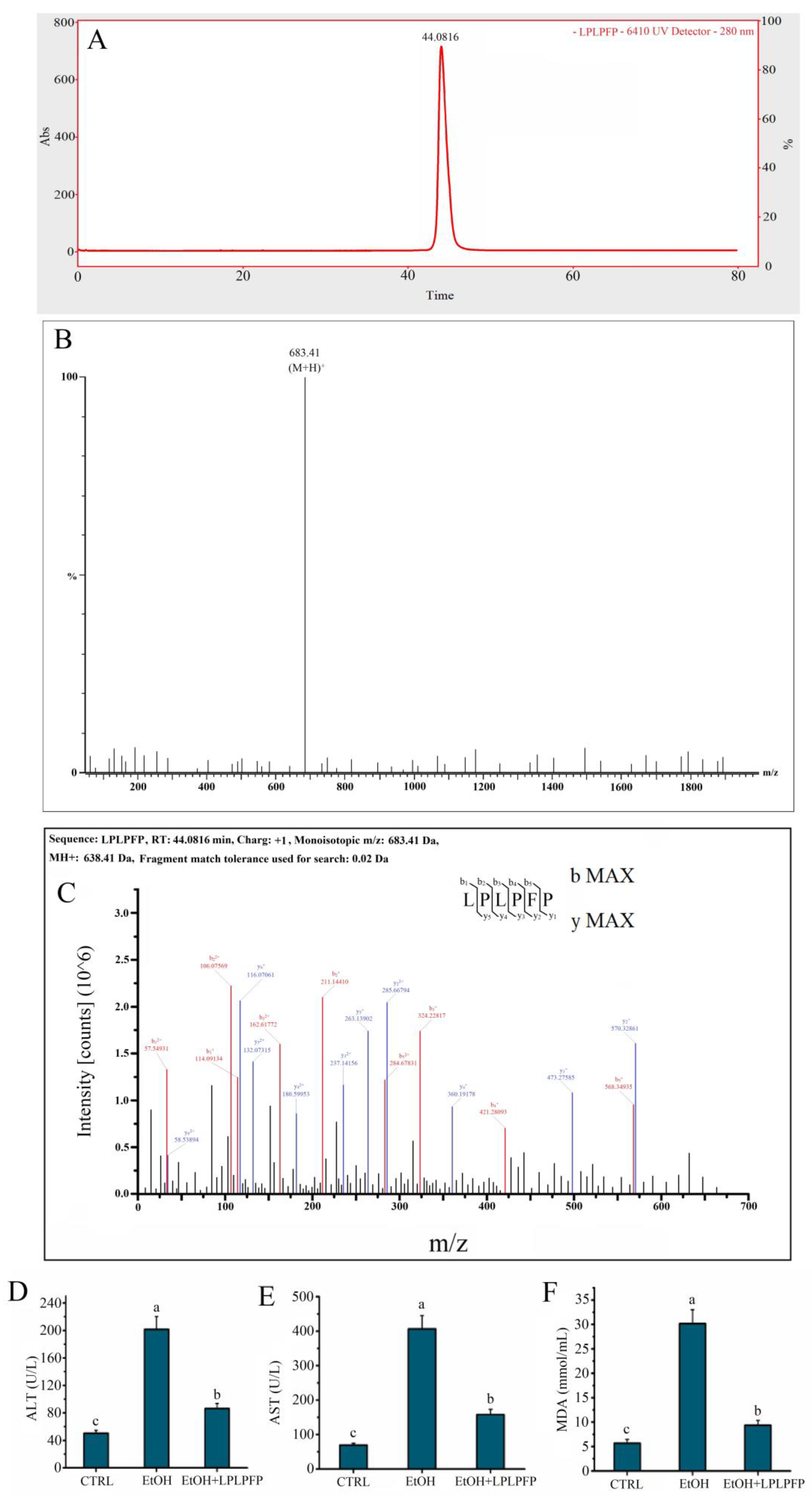
2.5. The Target Peptide Was Determined by LC-MS/MS
2.6. The Antioxidant Activity of LPLPFP
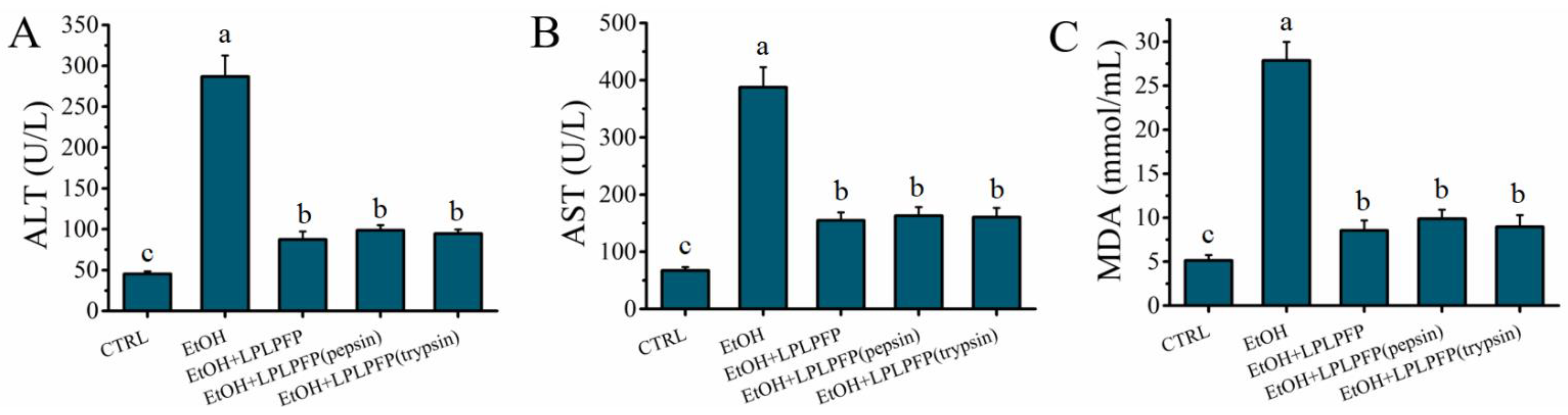
2.7. Effects of Pepsin-Trypsin Simulated GI Digestion on the Stability of LPLPFP
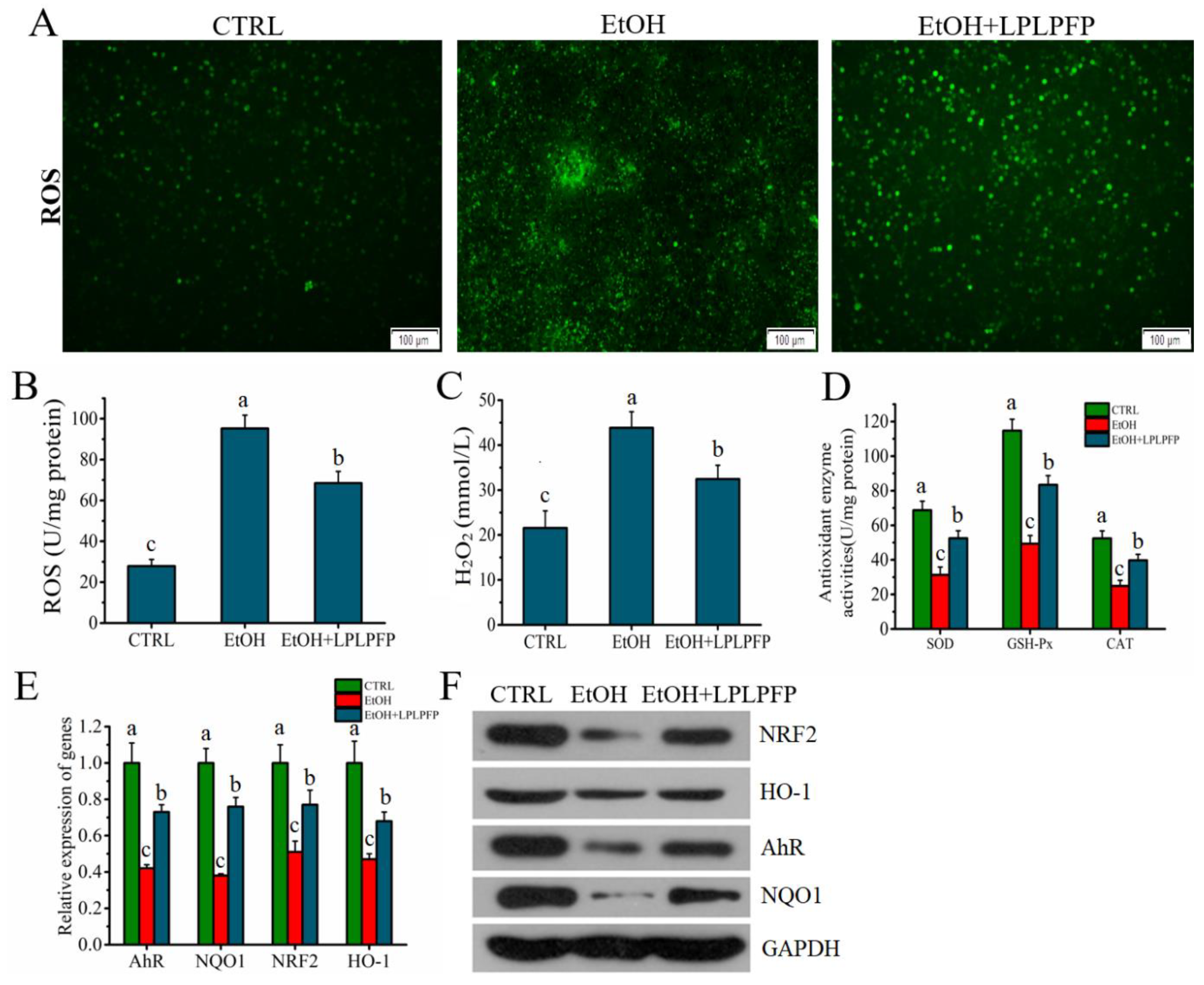
2.8. The Mechanism of LPLPFP Ameliorated Alcohol-Mediated Oxidative Stress in HHL-5 Hepatocytes
3. Materials and Methods
3.1. Preparation of GLHP
3.2. Isolation and Identification of Bioactive Peptides from GLHP
3.3. Measurement of Antioxidant Activity
3.4. Cell Culture and Treatments
3.5. Simulated Gastrointestinal (GI) Digestion
3.6. Determination of Cell Viability
3.7. Determination of Oxidative Stress Indicators
3.8. RT-PCR Analysis
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, N.; Collaborators, G.A. Alcohol use and burden for 195 countries and territories, 1990–2016: A systematic analysis for the global burden of disease study 2016 GDB 2016 alcohol collaborators. Lancet 2018, 392, 1015–1035. [Google Scholar] [CrossRef] [Green Version]
- Nie, W.; Zhou, K.; Wang, Y.; Wang, Z.M.; Xie, Y.; Zhou, H.; Xu, B.C. Isolation and identification of bioactive peptides from Xuanwei ham that rescue oxidative stress damage induced by alcohol in HHL-5 hepatocytes. Food Funct. 2020, 11, 9710–9720. [Google Scholar] [CrossRef]
- Balasubramaniyan, V.; Sailaja, J.K.; Nalini, N. Role of leptin on alcohol-induced oxidative stress in swiss mice. Pharmacol. Res. 2003, 47, 211–216. [Google Scholar] [CrossRef]
- Cederbaum, A.I. Cytochrome P450 2E1- dependent oxidant stress and upregulation of antioxidant defense in liver cells. J. Gastroenterol. Hepatol. 2006, 21 (Suppl. S3), S22–S25. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B. Teaching the basics of redox biology to medical and graduate students: Oxidants, antioxidants and disease mechanisms. Redox Biol. 2013, 1, 244–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, A.; Kaplowitz, N.; Tsukamoto, H.; Kamimura, S.; Fernandez-Checa, J.C. Hepatic mitochondrial glutathione depletion and progression of experimental alcoholic liver disease in rats. Hepatology 1992, 16, 1423–1427. [Google Scholar] [CrossRef]
- Gong, P.; Cederbaum, A.I.; Nieto, N. Heme oxygenase-1 protects HepG2 cells against cytochrome P450 2E1-dependent toxicity. Free Radic. Biol. Med. 2004, 36, 307–318. [Google Scholar] [CrossRef]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef]
- Xiao, C.; Zhou, F.; Zheng, L.; Cai, Y.; Zhao, M. Chicken breast-derived alcohol dehydrogenase-activating peptides in response to physicochemical changes and digestion simulation: The vital role of hydrophobicity. Food Res. Int. 2020, 136, 109592. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Du, Y.Y.; Xu, F.R.; Zhou, K.; Wang, Z.M.; Al-Dalali, S.; Wang, Y.; Li, X.M.; Ma, Y.H.; Xie, Y.; et al. Oligopeptides from Jinhua ham prevent alcohol-induced liver damage by regulating intestinal homeostasis and oxidative stress in mice. Food Funct. 2021, 12, 10053–10070. [Google Scholar] [CrossRef]
- Xing, L.; Li, G.; Toldrá, F.; Zhang, W. The physiological activity of bioactive peptides obtained from meat and meat by-products. Adv. Food Nutr. Res. 2021, 97, 97,147–185. [Google Scholar] [CrossRef]
- Wang, X.; Wang, D.Y.; Xu, W.M.; Zou, Y.; Fang, W.M.; Tian, H.W.; Zhou, F.Y. Research on characteristics and flavor of three kinds of poultry blood tofu gel. Sci. Technol. Food Ind. 2021, 42, 76–82. [Google Scholar] [CrossRef]
- Xing, L.; Liu, R.; Gao, X.; Zheng, J.; Wang, C.; Zhou, G. The proteomics homology of antioxidant peptides extracted from dry-cured xuanwei and jinhua ham. Food Chem. 2018, 266, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Cacciuttolo, M.A.; Trinh, L.; Lumpkin, J.A.; Rao, G. Hyperoxia induces DNA damage in mammalian cells. Free Radical Biol. Med. 1993, 14, 267–276. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Zhang, W.G.; Zhou, G.H.; Xu, X.L.; Kang, Z.L.; Yin, Y. Isolation and Identification of Antioxidant Peptides from Jinhua Ham. J. Agric. Food Chem. 2013, 61, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.C. Purification of antioxidative peptides prepared from enzymatic hydrolysates of tuna dark muscle by-product. Food Chem. 2010, 122, 42–48. [Google Scholar] [CrossRef]
- Zhang, W.G.; Xiao, S.; Himali, S.; Lee, E.J.; Ahn, D.U. Improving functional value of meat products. Meat Sci. 2010, 86, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Mendis, E.; Jung, W.K.; Je, J.Y.; Kim, S.K. Purification of a radical scavenging peptide from fermented mussel sauce and its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Kim, S.K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin Rana catesbeiana. Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef]
- Ko, S.C.; Kim, D.; Jeon, Y.J. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef]
- Ambade, A.; Mandrekar, P. Oxidative stress and inflammation: Essential partners in alcoholic liver disease. Int. J. Hepatol. 2012, 853175. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Hao, L.; Zhang, W.; Zhong, W.; Guo, W.; Yue, R.; Sun, X.; Zhou, Z. Activation of AhR-NQO1 signaling pathway protects against alcohol-induced liver injury by improving redox balance. Cell Mol. Gastroenter Hepatol. 2021, 12, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Lee, B.; Park, K.E.; Utsuki, T.; Shin, T.; Oh, C.W.; Kim, H.R. Dieckol enhances the expression of antioxidant and detoxifying enzymes by the activation of Nrf2−MAPK signalling pathway in HepG2 cells. Food Chem. 2015, 174, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Wang, S.Y.; Lin, Y.T.; Chen, Y.C. Antioxidant activities of chicken liver hydrolysates by pepsin treatment. Int. J. Food Sci. Tech. 2014, 49, 1654–1662. [Google Scholar] [CrossRef]
- Xie, N.N.; Liu, S.S.; Wang, C.; Li, B. Stability of casein antioxidant peptide fractions during in vitro digestion/Caco-2 cell model: Characteristics of the resistant peptides. Eur. Food Res. Technol. 2014, 239, 577–586. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content % | Concentration (mg/mL) | DPPH Radical Scavenging Activity (%) | ∙OH Radical Scavenging Activity (%) | Fe2+ Chelating Ability (%) | |
|---|---|---|---|---|---|
| LPLPFP | 3.83% | 1 | 95.74 ± 2.86 a | 88.45 ± 1.52 a | 35.32 ± 4.01 a |
| BHT | / | 1 | 90.56 ± 1.73 b | 79.38 ± 2.03 b | 14.39 ± 2.46 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Chen, Z.; Wei, H.; Zhu, S.; Cai, K. Isolation and Identification of Antioxidative Peptide from Goose Liver Hydrolysate to Ameliorate Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes. Molecules 2022, 27, 7151. https://doi.org/10.3390/molecules27217151
Du Y, Chen Z, Wei H, Zhu S, Cai K. Isolation and Identification of Antioxidative Peptide from Goose Liver Hydrolysate to Ameliorate Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes. Molecules. 2022; 27(21):7151. https://doi.org/10.3390/molecules27217151
Chicago/Turabian StyleDu, Yeye, Zhihong Chen, Haiyang Wei, Shuangjie Zhu, and Kezhou Cai. 2022. "Isolation and Identification of Antioxidative Peptide from Goose Liver Hydrolysate to Ameliorate Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes" Molecules 27, no. 21: 7151. https://doi.org/10.3390/molecules27217151