
Analysis of the In Vitro Toxicity of Nanocelluloses in Human Lung Cells as Compared to Multi-Walled Carbon Nanotubes

 , , , , , , ,
, , , , , , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Characterization of Cellulose Micro/Nanofibrils and Nanocrystals
2.2. Preparation of the Nanocelluloses and MWCNT Exposure Suspensions
2.3. Cell Culture and Exposure of Nanofibers
2.4. Cytotoxicity Assessment
2.4.1. MTT Assay
2.4.2. Clonogenic Assay
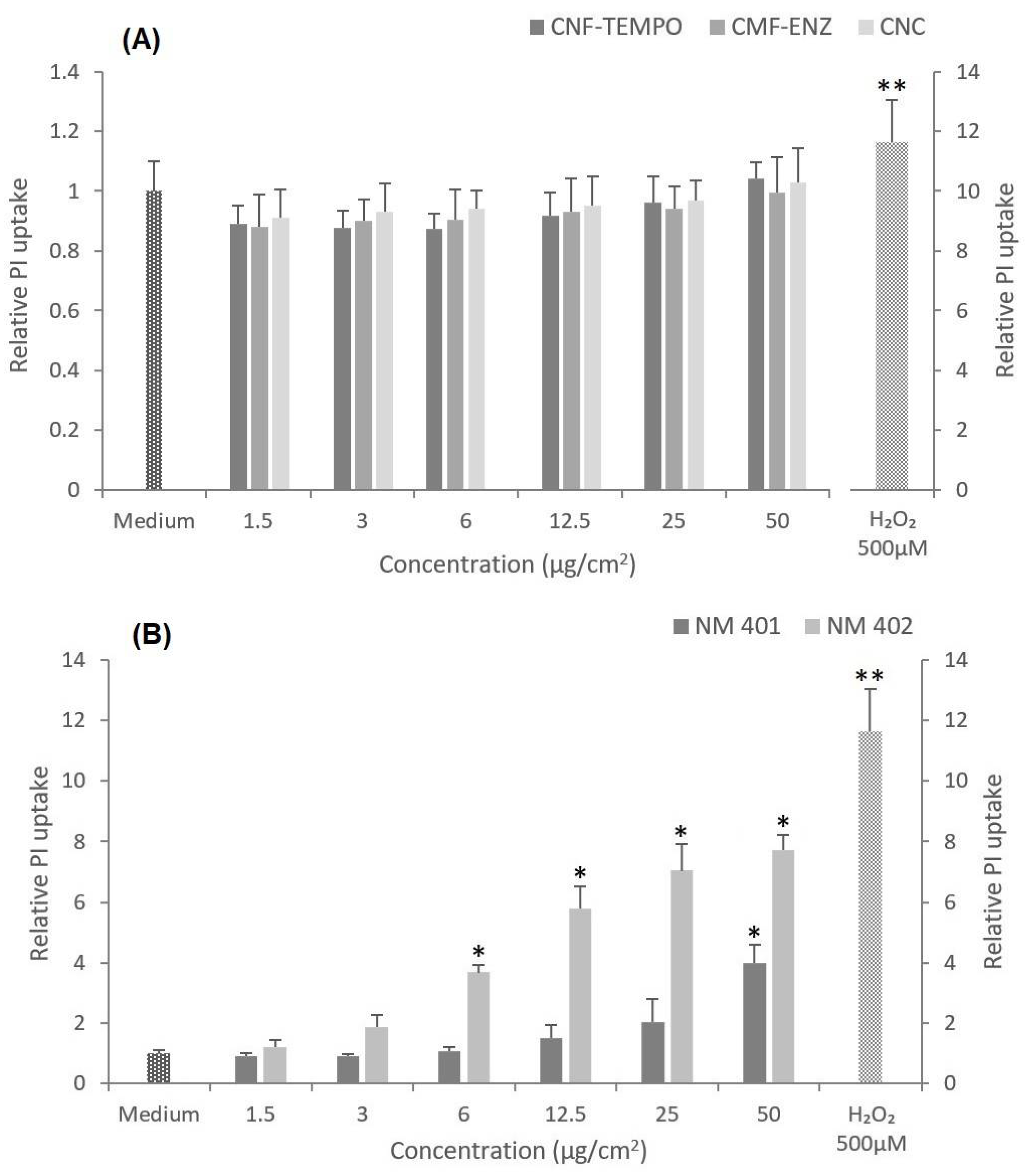
2.4.3. Propidium Iodide (PI) Membrane Integrity Assay
2.5. Reactive Oxygen Species (ROS) Production
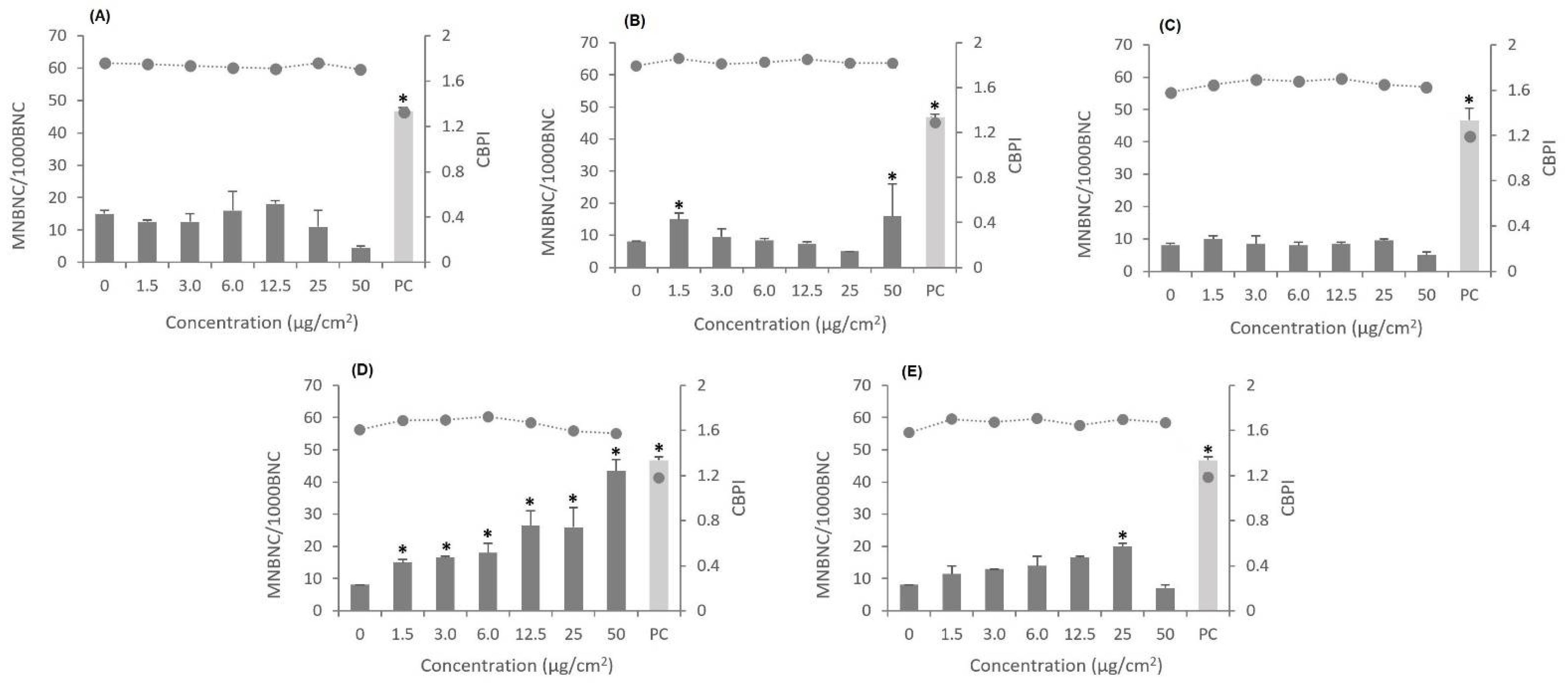
2.6. Cytokinesis-Blocked Micronucleus (CBMN) Assay
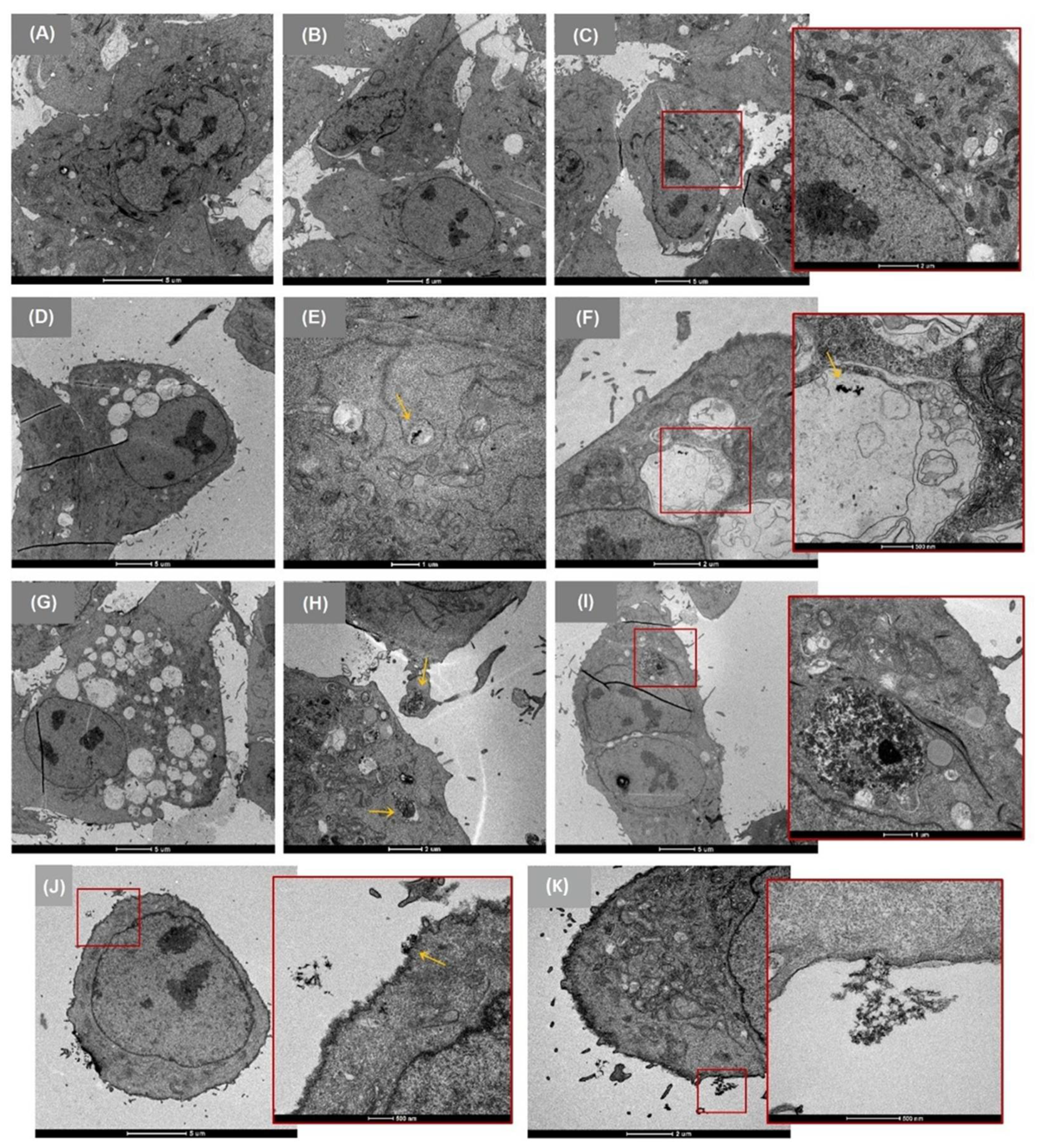
2.7. Cellular Uptake by TEM Imaging of Cells Exposed to CMNM
2.8. Statistical Analysis
3. Results
3.1. Characterization of Nanocellulose
3.2. Cellular Uptake
3.3. Cytotoxicity
3.4. Oxidative Stress
3.5. Genotoxic Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Varshney, V.K.; Naithani, S. Chemical functionalization of cellulose derived from nonconventional sources. In Cellulose Fibers: Bio-and Nano-Polymer Composites; Springer: Berlin/Heidelberg, Germany, 2011; pp. 43–60. [Google Scholar]
- Nechyporchuk, O.; Belgacem, M.N.; Bras, J. Production of cellulose nanofibrils: A review of recent advances. Ind. Crops Prod. 2016, 93, 2–25. [Google Scholar] [CrossRef]
- Kargarzadeh, H.; Mariano, M.; Gopakumar, D.; Ahmad, I.; Thomas, S.; Dufresne, A.; Huang, J.; Lin, N. Advances in cellulose nanomaterials. Cellulose 2018, 25, 2151–2189. [Google Scholar] [CrossRef]
- Ventura, C.; Pinto, F.; Lourenço, A.F.; Ferreira, P.J.T.; Louro, H.; Silva, M.J. On the toxicity of cellulose nanocrystals and nanofibrils in animal and cellular models. Cellulose 2020, 27, 5509–5544. [Google Scholar] [CrossRef]
- Lin, N.; Dufresne, A. Nanocellulose in biomedicine: Current status and future prospect. Eur. Polym. J. 2014, 59, 302–325. [Google Scholar] [CrossRef] [Green Version]
- Foster, E.J.; Moon, R.J.; Agarwal, U.P.; Bortner, M.J.; Bras, J.; Camarero-Espinosa, S.; Chan, K.J.; Clift, M.J.D.; Cranston, E.D.; Eichhorn, S.J.; et al. Current characterization methods for cellulose nanomaterials. Chem. Soc. Rev. 2018, 47, 2609–2679. [Google Scholar] [CrossRef] [Green Version]
- Shatkin, J.A.W.; Theodore, H.; Bilek, E.M.; Cowie, J. Market projections of cellulose nanomaterial-enabled products—Part 1: Applications. Tappi J. 2014, 13, 9–16. [Google Scholar] [CrossRef]
- Salas, C.; Nypelö, T.; Rodriguez-Abreu, C.; Carrillo, C.; Rojas, O.J. Nanocellulose properties and applications in colloids and interfaces. Curr. Opin. Colloid Interface Sci. 2014, 19, 383–396. [Google Scholar] [CrossRef]
- Bitounis, D.; Pyrgiotakis, G.; Bousfield, D.; Demokritou, P. Dispersion preparation, characterization, and dosimetric analysis of cellulose nano-fibrils and nano-crystals: Implications for cellular toxicological studies. NanoImpact 2019, 15, 100171. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Prabakaran, M.; Ke, M.; Gang, X.; Chung, I.M.; Um, I.C.; Gopiraman, M.; Kim, I.S. Highly dispersed nanoscale hydroxyapatite on cellulose nanofibers for bone regeneration. Mater. Lett. 2016, 168, 56–61. [Google Scholar] [CrossRef]
- Patricia, C. Polysaccharide-based films and coatings for food packaging: A review. Food Hydrocoll. 2017, 68, 136–148. [Google Scholar] [CrossRef]
- Lou, Y.R.; Kanninen, L.; Kuisma, T.; Niklander, J.; Noon, L.A.; Burks, D.; Urtti, A.; Yliperttula, M. The use of nanofibrillar cellulose hydrogel as a flexible three-dimensional model to culture human pluripotent stem cells. Stem Cells Dev. 2014, 23, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisin, E.R.; Yanamala, N.; Rodin, D.; Menas, A.; Farcas, M.; Russo, M.; Guppi, S.; Khaliullin, T.O.; Iavicoli, I.; Harper, M.; et al. Enhanced morphological transformation of human lung epithelial cells by continuous exposure to cellulose nanocrystals. Chemosphere 2020, 250, 126170. [Google Scholar] [CrossRef] [PubMed]
- Hakkarainen, T.; Koivuniemi, R.; Kosonen, M.; Escobedo-Lucea, C.; Sanz-Garcia, A.; Vuola, J.; Valtonen, J.; Tammela, P.; Mäkitie, A.; Luukko, K.; et al. Nanofibrillar cellulose wound dressing in skin graft donor site treatment. J. Control. Release Off. J. Control. Release Soc. 2016, 244, 292–301. [Google Scholar] [CrossRef] [PubMed]
- DeLoid, G.M.; Sohal, I.S.; Lorente, L.R.; Molina, R.M.; Pyrgiotakis, G.; Stevanovic, A.; Zhang, R.; McClements, D.J.; Geitner, N.K.; Bousfield, D.W.; et al. Reducing Intestinal Digestion and Absorption of Fat Using a Nature-Derived Biopolymer: Interference of Triglyceride Hydrolysis by Nanocellulose. ACS Nano 2018, 12, 6469–6479. [Google Scholar] [CrossRef]
- Ventura, C.; Lourenco, A.F.; Sousa-Uva, A.; Ferreira, P.J.T.; Silva, M.J. Evaluating the genotoxicity of cellulose nanofibrils in a co-culture of human lung epithelial cells and monocyte-derived macrophages. Toxicol. Lett. 2018, 291, 173–183. [Google Scholar] [CrossRef]
- Kangas, H.; Lahtinen, P.; Sneck, A.; Saariaho, A.-M.; Laitinen, O.; Hellén, E. Characterization of fibrillated celluloses. A short review and evaluation of characteristics with a combination of methods. Nord. Pulp Pap. Res. J. 2014, 29, 129–143. [Google Scholar] [CrossRef]
- Chinga-Carrasco, G.; Miettinen, A.; Hendriks, C.L.L.; Gamstedt, E.K.; Kataja, M. Structural characterisation of kraft pulp fibres and their nanofibrillated materials for biodegradable composite applications. Nanocompos. Polym. Anal. Methods 2011, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Abdul Khalil, H.P.S.; Bhat, A.H.; Ireana Yusra, A.F. Green composites from sustainable cellulose nanofibrils: A review. Carbohydr. Polym. 2012, 87, 963–979. [Google Scholar] [CrossRef]
- Garcia de Rodriguez, N.L.; Thielemans, W.; Dufresne, A. Sisal cellulose whiskers reinforced polyvinyl acetate nanocomposites. Cellulose 2006, 13, 261–270. [Google Scholar] [CrossRef]
- Beck-Candanedo, S.; Roman, M.; Gray, D.G. Effect of reaction conditions on the properties and behavior of wood cellulose nanocrystal suspensions. Biomacromolecules 2005, 6, 1048–1054. [Google Scholar] [CrossRef]
- Bai, W.; Holbery, J.; Li, K. A technique for production of nanocrystalline cellulose with a narrow size distribution. Cellulose 2009, 16, 455–465. [Google Scholar] [CrossRef]
- Zhu, H.; Luo, W.; Ciesielski, P.N.; Fang, Z.; Zhu, J.Y.; Henriksson, G.; Himmel, M.E.; Hu, L. Wood-Derived Materials for Green Electronics, Biological Devices, and Energy Applications. Chem. Rev. 2016, 116, 9305–9374. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, Y.; Wang, X.; Li, M.; Lei, H.; Xu, H. Cellulose nanocrystals prepared from wheat bran: Characterization and cytotoxicity assessment. Int. J. Biol. Macromol. 2019, 140, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Luzi, F.; Puglia, D.; Sarasini, F.; Tirillò, J.; Maffei, G.; Zuorro, A.; Lavecchia, R.; Kenny, J.M.; Torre, L. Valorization and extraction of cellulose nanocrystals from North African grass: Ampelodesmos mauritanicus (Diss). Carbohydr. Polym. 2019, 209, 328–337. [Google Scholar] [CrossRef]
- Tibolla, H.; Pelissari, F.M.; Martins, J.T.; Lanzoni, E.M.; Vicente, A.A.; Menegalli, F.C.; Cunha, R.L. Banana starch nanocomposite with cellulose nanofibers isolated from banana peel by enzymatic treatment: In vitro cytotoxicity assessment. Carbohydr. Polym. 2019, 207, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Clift, M.J.D.; Foster, E.J.; Vanhecke, D.; Studer, D.; Wick, P.; Gehr, P.; Rothen-Rutishauser, B.; Weder, C. Investigating the Interaction of Cellulose Nanofibers Derived from Cotton with a Sophisticated 3D Human Lung Cell Coculture. Biomacromolecules 2011, 12, 3666–3673. [Google Scholar] [CrossRef] [Green Version]
- Catalán, J.; Norppa, H. Safety Aspects of Bio-Based Nanomaterials. Bioengineering 2017, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Stefaniak, A.B.; Seehra, M.S.; Fix, N.R.; Leonard, S.S. Lung biodurability and free radical production of cellulose nanomaterials. Inhal. Toxicol. 2014, 26, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Ilves, M.; Vilske, S.; Aimonen, K.; Lindberg, H.K.; Pesonen, S.; Wedin, I.; Nuopponen, M.; Vanhala, E.; Højgaard, C.; Winther, J.R.; et al. Nanofibrillated cellulose causes acute pulmonary inflammation that subsides within a month. Nanotoxicology 2018, 12, 729–746. [Google Scholar] [CrossRef]
- Hadrup, N.; Knudsen, K.B.; Berthing, T.; Wolff, H.; Bengtson, S.; Kofoed, C.; Espersen, R.; Hojgaard, C.; Winther, J.R.; Willemoes, M.; et al. Pulmonary effects of nanofibrillated celluloses in mice suggest that carboxylation lowers the inflammatory and acute phase responses. Environ. Toxicol. Pharmacol. 2019, 66, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, H.K.; Catalán, J.; Aimonen, K.J.; Wolff, H.; Wedin, I.; Nuopponen, M.; Savolainen, K.M.; Norppa, H. Evaluation of the genotoxic potential of different types of nanofibrillated celluloses. TechConnect Briefs 2017, 1, 229–232. [Google Scholar]
- Ventura, C.; Pereira, J.F.S.; Matos, P.; Marques, B.; Jordan, P.; Sousa-Uva, A.; Silva, M.J. Cytotoxicity and genotoxicity of MWCNT-7 and crocidolite: Assessment in alveolar epithelial cells versus their coculture with monocyte-derived macrophages. Nanotoxicology 2020, 14, 479–503. [Google Scholar] [CrossRef] [PubMed]
- Lynch, I.; Weiss, C.; Valsami-Jones, E. A strategy for grouping of nanomaterials based on key physico-chemical descriptors as a basis for safer-by-design NMs. Nano Today 2014, 9, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Aimonen, K.; Suhonen, S.; Hartikainen, M.; Lopes, V.R.; Norppa, H.; Ferraz, N.; Catalán, J. Role of Surface Chemistry in the In Vitro Lung Response to Nanofibrillated Cellulose. Nanomaterials (Basel) 2021, 11, 389. [Google Scholar] [CrossRef] [PubMed]
- Stoudmann, N.; Schmutz, M.; Hirsch, C.; Nowack, B.; Som, C. Human hazard potential of nanocellulose: Quantitative insights from the literature. Nanotoxicology 2020, 14, 1241–1257. [Google Scholar] [CrossRef]
- Yanamala, N.; Kisin, E.R.; Menas, A.L.; Farcas, M.T.; Khaliullin, T.O.; Vogel, U.B.; Shurin, G.V.; Schwegler-Berry, D.; Fournier, P.M.; Star, A.; et al. In Vitro Toxicity Evaluation of Lignin-(Un)coated Cellulose Based Nanomaterials on Human A549 and THP-1 Cells. Biomacromolecules 2016, 17, 3464–3473. [Google Scholar] [CrossRef]
- Yanamala, N.; Farcas, M.T.; Hatfield, M.K.; Kisin, E.R.; Kagan, V.E.; Geraci, C.L.; Shvedova, A.A. In Vivo Evaluation of the Pulmonary Toxicity of Cellulose Nanocrystals: A Renewable and Sustainable Nanomaterial of the Future. ACS Sustain. Chem. Eng. 2014, 2, 1691–1698. [Google Scholar] [CrossRef] [Green Version]
- Samulin Erdem, J.; Alswady-Hoff, M.; Ervik, T.K.; Skare, Ø.; Ellingsen, D.G.; Zienolddiny, S. Cellulose nanocrystals modulate alveolar macrophage phenotype and phagocytic function. Biomaterials 2019, 203, 31–42. [Google Scholar] [CrossRef]
- Despres, H.W.; Sabra, A.; Anderson, P.; Hemraz, U.D.; Boluk, Y.; Sunasee, R.; Ckless, K. Mechanisms of the immune response cause by cationic and anionic surface functionalized cellulose nanocrystals using cell-based assays. Toxicol. In Vitro 2019, 55, 124–133. [Google Scholar] [CrossRef]
- Catalan, J.; Rydman, E.; Aimonen, K.; Hannukainen, K.S.; Suhonen, S.; Vanhala, E.; Moreno, C.; Meyer, V.; Perez, D.D.; Sneck, A.; et al. Genotoxic and inflammatory effects of nanofibrillated cellulose in murine lungs. Mutagenesis 2017, 32, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Catalan, J.; Ilves, M.; Jarventaus, H.; Hannukainen, K.S.; Kontturi, E.; Vanhala, E.; Alenius, H.; Savolainen, K.M.; Norppa, H. Genotoxic and immunotoxic effects of cellulose nanocrystals in vitro. Environ. Mol. Mutagen. 2015, 56, 171–182. [Google Scholar] [CrossRef]
- Vartiainen, J.; Pöhler, T.; Sirola, K.; Pylkkänen, L.; Alenius, H.; Hokkinen, J.; Tapper, U.; Lahtinen, P.; Kapanen, A.; Putkisto, K.; et al. Health and environmental safety aspects of friction grinding and spray drying of microfibrillated cellulose. Cellulose 2011, 18, 775–786. [Google Scholar] [CrossRef]
- Pitkänen, M.; Kangas, H.; Laitinen, O.; Sneck, A.; Lahtinen, P.; Peresin, M.S.; Niinimäki, J. Characteristics and safety of nano-sized cellulose fibrils. Cellulose 2014, 21, 3871–3886. [Google Scholar] [CrossRef]
- Nordli, H.R.; Chinga-Carrasco, G.; Rokstad, A.M.; Pukstad, B. Producing ultrapure wood cellulose nanofibrils and evaluating the cytotoxicity using human skin cells. Carbohydr. Polym. 2016, 150, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Menas, A.L.; Yanamala, N.; Farcas, M.T.; Russo, M.; Friend, S.; Fournier, P.M.; Star, A.; Iavicoli, I.; Shurin, G.V.; Vogel, U.B.; et al. Fibrillar vs crystalline nanocellulose pulmonary epithelial cell responses: Cytotoxicity or inflammation? Chemosphere 2017, 171, 671–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, K.; Carlsson, D.O.; Ålander, E.; Lindström, T.; Strømme, M.; Mihranyan, A.; Ferraz, N. Translational study between structure and biological response of nanocellulose from wood and green algae. RSC Adv. 2014, 4, 2892–2903. [Google Scholar] [CrossRef]
- Čolić, M.; Mihajlović, D.; Mathew, A.; Naseri, N.; Kokol, V. Cytocompatibility and immunomodulatory properties of wood based nanofibrillated cellulose. Cellulose 2015, 22, 763–778. [Google Scholar] [CrossRef]
- Alexandrescu, L.; Syverud, K.; Gatti, A.; Chinga-Carrasco, G. Cytotoxicity tests of cellulose nanofibril-based structures. Cellulose 2013, 20, 1765–1775. [Google Scholar] [CrossRef]
- De Lima, R.; Oliveira Feitosa, L.; Rodrigues Maruyama, C.; Abreu Barga, M.; Yamawaki, P.C.; Vieira, I.J.; Teixeira, E.M.; Correa, A.C.; Caparelli Mattoso, L.H.; Fernandes Fraceto, L. Evaluation of the genotoxicity of cellulose nanofibers. Int. J. Nanomed. 2012, 7, 3555–3565. [Google Scholar] [CrossRef] [Green Version]
- Kirsten, R.A.; Jan, M.A.; Pieter-Jan, D.T.; Eveline, V.E.; Nadia, W.A.; Frederic, V.S.; Christophe, P.J.; Ludwig, D.T.; Elke, V.D.; Astrup, J.K.; et al. Multi-Walled Carbon Nanotubes, NM-400, NM-401, NM-402, NM-403: Characterisation and Physico-Chemical Properties; European Commission: JRC Science and Policy Reports; Publications Office of the European Union; 2014; p. 118. Available online: https://op.europa.eu/en/publication-detail/-/publication/8ad34329-79d0-4b6e-9a6e-96cf2f5b4f7a (accessed on 21 March 2022).
- Louro, H.; Pinhão, M.; Santos, J.; Tavares, A.; Vital, N.; Silva, M.J. Evaluation of the cytotoxic and genotoxic effects of benchmark multi-walled carbon nanotubes in relation to their physicochemical properties. Toxicol. Lett. 2016, 262, 123–134. [Google Scholar] [CrossRef]
- Rubio, L.; El Yamani, N.; Kazimirova, A.; Dusinska, M.; Marcos, R. Multi-walled carbon nanotubes (NM401) induce ROS-mediated HPRT mutations in Chinese hamster lung fibroblasts. Environ. Res. 2016, 146, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Di Ianni, E.; Erdem, J.S.; Møller, P.; Sahlgren, N.M.; Poulsen, S.S.; Knudsen, K.B.; Zienolddiny, S.; Saber, A.T.; Wallin, H.; Vogel, U.; et al. In vitro-in vivo correlations of pulmonary inflammogenicity and genotoxicity of MWCNT. Part. Fibre Toxicol. 2021, 18, 25. [Google Scholar] [CrossRef]
- Foster, K.A.; Oster, C.G.; Mayer, M.M.; Avery, M.L.; Audus, K.L. Characterization of the A549 Cell Line as a Type II Pulmonary Epithelial Cell Model for Drug Metabolism. Exp. Cell Res. 1998, 243, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, H.; Sha, S.; Li, J.; Zhou, Z.; Cao, Y. Evaluation of in vitro toxicity of silica nanoparticles (NPs) to lung cells: Influence of cell types and pulmonary surfactant component DPPC. Ecotoxicol. Environ. Saf. 2019, 186, 109770. [Google Scholar] [CrossRef]
- Saito, T.; Isogai, A. Wet Strength Improvement of TEMPO-Oxidized Cellulose Sheets Prepared with Cationic Polymers. Ind. Eng. Chem. Res. 2007, 46, 773–780. [Google Scholar] [CrossRef]
- Lourenço, A.F.; Gamelas, J.A.F.; Nunes, T.; Amaral, J.; Mutjé, P.; Ferreira, P.J. Influence of TEMPO-oxidised cellulose nanofibrils on the properties of filler-containing papers. Cellulose 2017, 24, 349–362. [Google Scholar] [CrossRef]
- Lourenço, A.F.; Gamelas, J.A.; Sarmento, P.; Ferreira, P.J. Enzymatic nanocellulose in papermaking–The key role as filler flocculant and strengthening agent. Carbohydr. Polym. 2019, 224, 115200. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Q.; Hirth, K.; Baez, C.; Agarwal, U.P.; Zhu, J.Y. Tailoring the yield and characteristics of wood cellulose nanocrystals (CNC) using concentrated acid hydrolysis. Cellulose 2015, 22, 1753–1762. [Google Scholar] [CrossRef]
- Rosa, R.R.; Fernandes Susete, N.; Saraiva, D.V.; Silva, P.E.S.; Godinho Maria, H.; Pereira, S.R.S.; Sousa, A.P.M. Cellulose Nanocrystals (CNCs) Aqueous Colloidal Suspensions: Evidence of a Density Inversion at the Isotropic-Liquid Crystal Phase Transition; Provisional Application for Patent at 21-04-2022; Raiz-Instituto de Investigação da Floresta e Papel Universidade Nova de Lisboa: Eixo, Portugal, 2021. [Google Scholar]
- Gamelas, J.A.F.; Pedrosa, J.; Lourenço, A.F.; Mutjé, P.; González, I.; Chinga-Carrasco, G.; Singh, G.; Ferreira, P.J.T. On the morphology of cellulose nanofibrils obtained by TEMPO-mediated oxidation and mechanical treatment. Micron 2015, 72, 28–33. [Google Scholar] [CrossRef]
- Henriksson, M.; Berglund, L.A.; Isaksson, P.; Lindström, T.; Nishino, T. Cellulose Nanopaper Structures of High Toughness. Biomacromolecules 2008, 9, 1579–1585. [Google Scholar] [CrossRef]
- Jensen, K.A.; Kembouche, Y.; Christiansen, E.; Jacobsen, N.R.; Wallin, H.; Guiot, C.E.; Spalla, O.; Witschger, O. Final Protocol for Producing Suitable Manufactured Nanomaterial Exposure Media—Report—The Generic NANOGENOTOX Dispersion Protocol—Standard Operation Procedure (SOP) and Background Documentation; The National Research Centre for the Working Environment (NRCWE): Copenhagen, Denmark, 2011; Available online: https://www.anses.fr (accessed on 11 March 2022).
- Tavares, A.M.; Louro, H.; Antunes, S.; Quarre, S.; Simar, S.; De Temmerman, P.J.; Verleysen, E.; Mast, J.; Jensen, K.A.; Norppa, H.; et al. Genotoxicity evaluation of nanosized titanium dioxide, synthetic amorphous silica and multi-walled carbon nanotubes in human lymphocytes. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2014, 28, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.M.; Calado, R.; Marto, J.; Bettencourt, A.; Almeida, A.J.; Gonçalves, L.M.D. Chitosan Nanoparticles as a Mucoadhesive Drug Delivery System for Ocular Administration. Mar Drugs 2017, 15, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test; OECD: Paris, France, 2016. [Google Scholar]
- OECD. OECD Guideline for the Testing of Chemicals 487-In Vitro Mammalian Cell Micronucleus Test; OECD: Paris, France, 2010. [Google Scholar]
- Mendieta, I.; Rodríguez-Nieto, M.; Nuñez-Anita, R.E.; Menchaca-Arredondo, J.L.; García-Alcocer, G.; Berumen, L.C. Ultrastructural changes associated to the neuroendocrine transdifferentiation of the lung adenocarcinoma cell line A549. Acta Histochem. 2021, 123, 151797. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.S.A.; Pohlmann, B.C.; Calado, V.; Bojorge, N.; Pereira, N., Jr. Production of nanocellulose by enzymatic hydrolysis: Trends and challenges. Eng. Life Sci. 2019, 19, 279–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, A.F.; Godinho, D.; Gamelas, J.A.F.; Sarmento, P.; Ferreira, P.J.T. Carboxymethylated cellulose nanofibrils in papermaking: Influence on filler retention and paper properties. Cellulose 2019, 26, 3489–3502. [Google Scholar] [CrossRef]
- Saraiva, D.V.; Chagas, R.; Abreu, B.M.d.; Gouveia, C.N.; Silva, P.E.S.; Godinho, M.H.; Fernandes, S.N. Flexible and Structural Coloured Composite Films from Cellulose Nanocrystals/Hydroxypropyl Cellulose Lyotropic Suspensions. Crystals 2020, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Lopes, V.R.; Sanchez-Martinez, C.; Strømme, M.; Ferraz, N. In vitro biological responses to nanofibrillated cellulose by human dermal, lung and immune cells: Surface chemistry aspect. Part. Fibre Toxicol. 2017, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fubini, B.; Ghiazza, M.; Fenoglio, I. Physico-chemical features of engineered nanoparticles relevant to their toxicity. Nanotoxicology 2010, 4, 347–363. [Google Scholar] [CrossRef]
- Reifarth, M.; Hoeppener, S.; Schubert, U.S. Uptake and Intracellular Fate of Engineered Nanoparticles in Mammalian Cells: Capabilities and Limitations of Transmission Electron Microscopy—Polymer-Based Nanoparticles. Adv. Mater. 2018, 30, 1703704. [Google Scholar] [CrossRef]
- Sousa de Almeida, M.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem. Soc. Rev. 2021, 50, 5397–5434. [Google Scholar] [CrossRef]
- Mahmoud, K.A.; Mena, J.A.; Male, K.B.; Hrapovic, S.; Kamen, A.; Luong, J.H.T. Effect of Surface Charge on the Cellular Uptake and Cytotoxicity of Fluorescent Labeled Cellulose Nanocrystals. ACS Appl. Mater. Interfaces 2010, 2, 2924–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shazali, N.A.H.; Zaidi, N.E.; Ariffin, H.; Abdullah, L.C.; Ghaemi, F.; Abdullah, J.M.; Takashima, I.; Nik Abd Rahman, N.M.A. Characterization and Cellular Internalization of Spherical Cellulose Nanocrystals (CNC) into Normal and Cancerous Fibroblasts. Materials (Basel) 2019, 12, 3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Hirani, A.A.; Colacino, K.R.; Lee, Y.W.; Roman, M. Cytotoxicity and cellular uptake of cellulose nanocrystals. Nano Life 2012, 2, 1241006. [Google Scholar] [CrossRef]
- Zhao, F.; Zhao, Y.; Liu, Y.; Chang, X.; Chen, C.; Zhao, Y. Cellular uptake, intracellular trafficking, and cytotoxicity of nanomaterials. Small 2011, 7, 1322–1337. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, A.; Cañete, M.; Roca, A.G.; Calero, M.; Veintemillas-Verdaguer, S.; Serna, C.J.; Morales Mdel, P.; Miranda, R. The influence of surface functionalization on the enhanced internalization of magnetic nanoparticles in cancer cells. Nanotechnology 2009, 20, 115103. [Google Scholar] [CrossRef]
- Mittag, A.; Hoera, C.; Kämpfe, A.; Westermann, M.; Kuckelkorn, J.; Schneider, T.; Glei, M. Cellular Uptake and Toxicological Effects of Differently Sized Zinc Oxide Nanoparticles in Intestinal Cells. Toxics 2021, 9, 96. [Google Scholar] [CrossRef]
- Cho, E.C.; Xie, J.; Wurm, P.A.; Xia, Y. Understanding the role of surface charges in cellular adsorption versus internalization by selectively removing gold nanoparticles on the cell surface with a I2/KI etchant. Nano Lett. 2009, 9, 1080–1084. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, M.; Okazaki, T.; Yudasaka, M. Quantification of Carbon Nanotubes Taken up by Macrophage Cells Using Optical Absorption Method. E-J. Surf. Sci. Nanotechnol. 2018, 16, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wang, L.; Wang, Z.; Cuschieri, A. Different cellular response mechanisms contribute to the length-dependent cytotoxicity of multi-walled carbon nanotubes. Nanoscale Res. Lett. 2012, 7, 361. [Google Scholar] [CrossRef] [Green Version]
- Ünlu, A.; Dinç, B.; Bektaş, M. Characterization of short-length multi-walled carbon nanotubes and cytotoxicity on MDA-MB-231 and HUVEC cell lines. Carbon Lett. 2019, 30, 143–153. [Google Scholar] [CrossRef]
- Pantarotto, D.; Briand, J.P.; Prato, M.; Bianco, A. Translocation of bioactive peptides across cell membranes by carbon nanotubes. Chem. Commun. (Camb.) 2004, 1, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, S.A.d.A. Biological effects of cellulose nanofibrils in a lung epithelial cell line. In FCT: Departamento de Ciências da Vida; Faculdade de Ciências e Tecnologia (FCT)—Universidade Nova de Lisboa (NOVA): RUN—Repositório Universidade Nova; 2019; Available online: https://run.unl.pt/ (accessed on 11 March 2022).
- Ni, H.; Zeng, S.; Wu, J.; Cheng, X.; Luo, T.; Wang, W.; Zeng, W.; Chen, Y. Cellulose nanowhiskers: Preparation, characterization and cytotoxicity evaluation. Bio-Med. Mater. Eng. 2012, 22, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Meschini, S.; Pellegrini, E.; Maestri, C.A.; Condello, M.; Bettotti, P.; Condello, G.; Scarpa, M. In vitro toxicity assessment of hydrogel patches obtained by cation-induced cross-linking of rod-like cellulose nanocrystals. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Male, K.B.; Leung, A.C.; Montes, J.; Kamen, A.; Luong, J.H. Probing inhibitory effects of nanocrystalline cellulose: Inhibition versus surface charge. Nanoscale 2012, 4, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Hua, K.; Ålander, E.; Lindström, T.; Mihranyan, A.; Strømme, M.; Ferraz, N. Surface Chemistry of Nanocellulose Fibers Directs Monocyte/Macrophage Response. Biomacromolecules 2015, 16, 2787–2795. [Google Scholar] [CrossRef] [PubMed]
- Hosseinidoust, Z.; Alam, M.N.; Sim, G.; Tufenkji, N.; van de Ven, T.G. Cellulose nanocrystals with tunable surface charge for nanomedicine. Nanoscale 2015, 7, 16647–16657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanif, Z.; Ahmed, F.R.; Shin, S.W.; Kim, Y.K.; Um, S.H. Size- and dose-dependent toxicity of cellulose nanocrystals (CNC) on human fibroblasts and colon adenocarcinoma. Colloids Surf. B Biointerfaces 2014, 119, 162–165. [Google Scholar] [CrossRef]
- Jalili, P.; Gueniche, N.; Lanceleur, R.; Burel, A.; Lavault, M.-T.; Sieg, H.; Böhmert, L.; Meyer, T.; Krause, B.-C.; Lampen, A.; et al. Investigation of the in vitro genotoxicity of two rutile TiO2 nanomaterials in human intestinal and hepatic cells and evaluation of their interference with toxicity assays. NanoImpact 2018, 11, 69–81. [Google Scholar] [CrossRef]
- Allegri, M.; Perivoliotis, D.K.; Bianchi, M.G.; Chiu, M.; Pagliaro, A.; Koklioti, M.A.; Trompeta, A.-F.A.; Bergamaschi, E.; Bussolati, O.; Charitidis, C.A. Toxicity determinants of multi-walled carbon nanotubes: The relationship between functionalization and agglomeration. Toxicol. Rep. 2016, 3, 230–243. [Google Scholar] [CrossRef] [Green Version]
- Di Cristo, L.; Bianchi, M.G.; Chiu, M.; Taurino, G.; Donato, F.; Garzaro, G.; Bussolati, O.; Bergamaschi, E. Comparative in Vitro Cytotoxicity of Realistic Doses of Benchmark Multi-Walled Carbon Nanotubes towards Macrophages and Airway Epithelial Cells. Nanomaterials (Basel) 2019, 9, 982. [Google Scholar] [CrossRef] [Green Version]
- García-Rodríguez, A.; Rubio, L.; Vila, L.; Xamena, N.; Velázquez, A.; Marcos, R.; Hernández, A. The Comet Assay as a Tool to Detect the Genotoxic Potential of Nanomaterials. Nanomaterials 2019, 9, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongkam, W.; Gerloff, K.; van Berlo, D.; Albrecht, C.; Schins, R.P.F. Oxidant generation, DNA damage and cytotoxicity by a panel of engineered nanomaterials in three different human epithelial cell lines. Mutagenesis 2016, 32, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.; Kling, K.; Jensen, K.A.; Clausen, P.A.; Madsen, A.M.; Wallin, H.; Vogel, U. Characterization of genotoxic response to 15 multiwalled carbon nanotubes with variable physicochemical properties including surface functionalizations in the FE1-Muta(TM) mouse lung epithelial cell line. Environ. Mol. Mutagen. 2015, 56, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gurel, V.; Morris, D.; Murray, D.W.; Zhitkovich, A.; Kane, A.B.; Hurt, R.H. Bioavailability of Nickel in Single-Wall Carbon Nanotubes. Adv. Mater. 2007, 19, 2790–2796. [Google Scholar] [CrossRef]
- Norppa, H.; Falck, G.C. What do human micronuclei contain? Mutagenesis 2003, 18, 221–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M. Cytokinesis-block micronucleus assay evolves into a “cytome” assay of chromosomal instability, mitotic dysfunction and cell death. Mutat. Res. 2006, 600, 58–66. [Google Scholar] [CrossRef]
- Araldi, R.P.; de Melo, T.C.; Mendes, T.B.; de Sá Júnior, P.L.; Nozima, B.H.; Ito, E.T.; de Carvalho, R.F.; de Souza, E.B.; de Cassia Stocco, R. Using the comet and micronucleus assays for genotoxicity studies: A review. Biomed. Pharmacother. 2015, 72, 74–82. [Google Scholar] [CrossRef]
- Valentin-Severin, I.; Le Hegarat, L.; Lhuguenot, J.-C.; Le Bon, A.-M.; Chagnon, M.-C. Use of HepG2 cell line for direct or indirect mutagens screening: Comparative investigation between comet and micronucleus assays. Mutat. Res./Genet. Toxicol. Environ. Mutagenesis 2003, 536, 79–90. [Google Scholar] [CrossRef]
- Gordobil, O.; Oberemko, A.; Saulis, G.; Baublys, V.; Labidi, J. In vitro cytotoxicity studies of industrial Eucalyptus kraft lignins on mouse hepatoma, melanoma and Chinese hamster ovary cells. Int. J. Biol. Macromol. 2019, 135, 353–361. [Google Scholar] [CrossRef]
- Kumar Singh, A.; Yadav, P.; Bharagava, R.; Saratale, G.; Raj, A. Biotransformation and Cytotoxicity Evaluation of Kraft Lignin Degraded by Ligninolytic Serratia liquefaciens. Front. Microbiol. 2019, 10, 2364. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Multi-Walled Carbon Nanotubes | Thickness ± SD (nm) | Geodesic Length ± SD (nm) | Aspect Ratio ± SD | Specific Surface Area (m2/g) |
|---|---|---|---|---|
| NM-401 | 67 ± 24 a | 4048 ± 2371 a | - | 140.46 a |
| 62.8 ± 1.4 b | 3366.4 ± 1.9 b | 53.6 ± 2.0 b | - | |
| NM-402 | 11 ± 3 a | 1372 ± 836 a | 226.4 a | |
| 10.7 ± 1.3 b | 1141.3 ± 2.0 b | 107.1 ± 1.9 b | - |
| Micro/Nanocellulose Sample | Fibrillation Yield (%) | CCOOH (μmol/g) | DP | [η] (mL/g) | Fibril Diameter 1 (nm) | Zeta-Potential (mV) | ||
|---|---|---|---|---|---|---|---|---|
| PBS | CM | PBS | CM | |||||
| CNF-TEMPO | 100 | 1332 | 309 | 130 | 10.7 ± 1.9 | - | −24.6 ± 1.0 | −19.7 ± 1.5 |
| CMF-ENZ | 4.9 | 143 | 1591 | 618 | 29.7 ± 7.3 | 85.2 ± 41.2 | −11.6 ± 1.0 | −9.4 ± 0.6 |
| CNC | 44 | - | - | - | 19.7 ± 6.1 | 36.0 ± 9.0 | −17.3 ± 0.8 | −13.9 ± 0.3 |
| Assay/Endpoint | CNF-TEMPO | CMF-ENZ | CNC | NM-401 | NM-402 |
|---|---|---|---|---|---|
| MTT | - | - | - | + | + |
| PI | - | - | - | ++ | ++ |
| Clonogenic | - | - | - | ++ | + |
| ROS, 1 h | - | - | - | - | ++ |
| ROS, 24 h | - | - | - | - | ++ |
| MN | - | + | - | ++ | (+) |
| CBPI | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, F.; Lourenço, A.F.; Pedrosa, J.F.S.; Gonçalves, L.; Ventura, C.; Vital, N.; Bettencourt, A.; Fernandes, S.N.; da Rosa, R.R.; Godinho, M.H.; et al. Analysis of the In Vitro Toxicity of Nanocelluloses in Human Lung Cells as Compared to Multi-Walled Carbon Nanotubes. Nanomaterials 2022, 12, 1432. https://doi.org/10.3390/nano12091432
Pinto F, Lourenço AF, Pedrosa JFS, Gonçalves L, Ventura C, Vital N, Bettencourt A, Fernandes SN, da Rosa RR, Godinho MH, et al. Analysis of the In Vitro Toxicity of Nanocelluloses in Human Lung Cells as Compared to Multi-Walled Carbon Nanotubes. Nanomaterials. 2022; 12(9):1432. https://doi.org/10.3390/nano12091432
Chicago/Turabian StylePinto, Fátima, Ana Filipa Lourenço, Jorge F. S. Pedrosa, Lídia Gonçalves, Célia Ventura, Nádia Vital, Ana Bettencourt, Susete N. Fernandes, Rafaela R. da Rosa, Maria Helena Godinho, and et al. 2022. "Analysis of the In Vitro Toxicity of Nanocelluloses in Human Lung Cells as Compared to Multi-Walled Carbon Nanotubes" Nanomaterials 12, no. 9: 1432. https://doi.org/10.3390/nano12091432