
Non-Coding RNAs in the Crosstalk between Breast Cancer Cells and Tumor-Associated Macrophages


1
Section of Histology and Medical Embryology, Department of Anatomical, Histological, Forensic and Orthopedic Sciences, Sapienza University of Rome, 00161 Rome, Italy
2
Oncogenomic and Epigenetic Unit, IRCCS Regina Elena National Cancer Institute-IFO, 00144 Rome, Italy
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2022, 8(1), 16; https://doi.org/10.3390/ncrna8010016
Submission received: 14 December 2021
/
Revised: 19 January 2022
/
Accepted: 1 February 2022
/
Published: 6 February 2022
(This article belongs to the Collection Secreted Non-Coding RNAs as Signaling Molecules Driving Cell-to-Cell Communication in Cancer)
Abstract
:Non-coding RNAs (ncRNAs) play a pivotal role in regulating the tumor microenvironment (TME) by controlling gene expression at multiple levels. In tumors, ncRNAs can mediate the crosstalk between cancer cells and other cells in the TME, such as immune cells, stromal cells, and endothelial cells, influencing tumor development and progression. Tumor-associated macrophages (TAMs) are among the most abundant inflammatory cells infiltrating solid cancers that promote tumorigenesis, and their infiltration correlates with a poor prognosis in many tumors. Cancer cells produce different ncRNAs that orchestrate TAM recruitment and polarization toward a tumor-promoting phenotype. Tumor-reprogrammed macrophages shape the TME by promoting angiogenesis and tissue remodeling, and suppressing the anti-tumor activity of adaptive immune cells. TAMs can also produce ncRNA molecules that boost cancer cell proliferation and direct their phenotype and metabolic changes facilitating cancer progression and metastasis. This review will focus on the crosstalk between cancer cells and TAMs mediated by microRNAs and long non-coding RNAs during breast cancer (BC) initiation and progression.
1. Introduction
The communication of cancer cells with the surrounding environment is crucial for tumor development and progression. Cancer cells direct the formation of a TME that sustains their nutrient demand by spreading through the blood to other organs, and the escape from immune surveillance [1,2,3]. The TME includes cancer cells, blood vessels, immune cells, and stromal cells with the adjacent extracellular matrix (ECM) [3,4]. TAMs represent one of the primary infiltrating myeloid cells that are part of the TME, and their presence correlates with tumor aggressiveness and bad prognosis in breast cancer and other tumors [5].
TAMs partly originate from tissue-resident macrophages, but they mostly derive from circulating monocytic precursors recruited to the tumor site by chemotactic signals released by cancer cells [6]. These chemotactic signals include cytokines such as CSF-1, and chemokines such as CCL-2 and CCL-5, which recruit inflammatory monocytes and direct TAM differentiation [6,7]. Although TAMs can support or inhibit cancer progression depending on their activation state and polarization, many studies have shown that they mostly polarize toward a tumor-promoting phenotype [6,7]. Macrophages can differentiate into the pro-inflammatory M1 phenotype or the anti-inflammatory M2 phenotype [8]. Classical M1 macrophages are driven by IFNγ and bacteria during TH1 (type I helper) responses against pathogens, and produce inflammatory cytokines and tissue-damaging factors [9]. M1-like macrophages exert an anti-tumor activity during tumor onset by distinguishing and killing malignant cells. However, in established tumors, cancer cells drive macrophage shift into the alternative M2 phenotype [6,10]. M2 macrophages mediate tissue repair and remodeling after inflammatory responses in healthy tissues [11]. However, in the TME, macrophages promote cancer cell proliferation, epithelial-to-mesenchymal transformation (EMT), and stemness [12]. Moreover, TAMs favor metastasis formation by inducing angiogenesis and tissue remodeling, and contribute to the generation of an immunosuppressive environment that protects cancer cells from being killed by other immune cells [7,13].
Many studies have extensively identified macrophages as CD68-expressing cells in tumor tissues. The M2-like population is commonly distinguished for the expression of CD163, CD204, and CD206; on the contrary, M1-like macrophages express HLA-DR, iNOS, and pSTAT1, although these markers alone are not exclusive for M1 macrophages [10].
Classical M1 and alternative M2 macrophages are only two extremes of a broad spectrum of possible phenotypes. In tumors, the multitude of signals from the TME leads to TAM phenotypic plasticity and heterogeneous functions ranging between M1 and M2 phenotypes [6,14].
In recent years, many studies have shown that non-coding RNAs’ (ncRNAs) biosynthesis and processing are significantly altered in malignant cells, contributing to atypical cancer cell behavior [15,16].
Non-coding RNAs are a class of RNAs that lack a protein-coding region but can influence gene expression in healthy and pathological contexts. NcRNAs include small ncRNAs shorter than 200 nucleotides (nt) and long ncRNAs (lncRNAs) longer than 200 nt. Shorter RNAs can be further divided into small nucleolar RNAs (snoRNAs), small interfering RNAs (siRNAs), small nuclear RNAs (snRNAs), transfer RNAs (tRNAs), Piwi-interacting RNAs (piRNAs), and the most studied microRNAs (miRNAs) [17]. miRNAs can regulate post-transcriptional gene expression by repressing or degrading complementary mRNAs, and originate from a long primary miRNA (pri-miRNA) processed by the nuclear ribonuclease Drosha to a long precursor miRNA (pre-miRNA). Finally, the pre-miRNA is exported from the nucleus and cleaved by the RNase enzyme Dicer to produce the mature double-stranded miRNA [17,18]. The active single-stranded miRNA is then loaded into the RISC (RNA-induced silencing complex), where a member of the Argonaute (Ago) family of proteins allows its functional activity [19,20].
LncRNAs biosynthesis is similar to that of miRNAs for many aspects. LncRNAs can control gene expression by directly interacting with proteins, lipids, DNA, and RNA. However, because of their ability to bind to different molecules, lncRNAs functions are more complex than that of miRNAs. They can regulate many cellular processes, such as translation, splicing, and chromatin status, by interacting with RNA/DNA or influencing transcription factor stability and localization [21,22,23].
NcRNAs influence different aspects of tumor onset and development by promoting or suppressing tumorigenesis or affecting drug resistance. Moreover, the release of ncRNAs inside exosomes mediates the communication between cancer cells and other cells in the TME [24]. Researchers have identified several miRNAs in breast cancer that regulate the expression levels of ERα [25], cell survival and apoptosis [26], EMT [27], and cell stemness [28]. Like miRNAs, lncRNAs can be up-downregulated or differentially expressed in the various breast cancer subtypes and drive breast cancer progression by regulating pro-metastatic factors [29]. This review will focus on the miRNAs and the lncRNAs involved in breast cancer cell-TAM crosstalk.
2. Role of Non-Coding RNAs in TAM Recruitment and Polarization in Breast Cancer
Non-coding RNAs produced by cancer cells may regulate monocyte recruitment to the tumor site and direct their differentiation into M1 or M2 macrophages by influencing the expression of pro-or anti-inflammatory genes in macrophages [30,31].
Monocyte recruitment is guided by chemotactic signals released by cancer cells. One of the most potent factors involved in this process is CCL2, upregulated in many cancers [32,33]. Recently, Frank et al. found that miR-375 induces CCL2 expression in breast cancer cells [34]. MiR-375 is released by apoptotic BC cells and stimulates the production of CCL2 in other cancer cells by unknown mechanisms. Furthermore, miR-375 binds to the macrophage CD36 receptor and is transferred to TAMs, where it directly binds and downregulates TNS3 and PXN, two migration-inhibitory proteins, increasing the macrophage’s ability to infiltrate the tumor, resulting in cancer progression [34]. CCL2 mediates macrophage recruitment also at metastatic sites [35]. LncRNA associated with breast cancer brain metastases (lnc-BM) is significantly upregulated in brain metastasis and influences TAM recruitment by inducing the production of CCL2 via JAK2/STAT3 pathway activation in cancer cells [36]. The cytokine colony-stimulating factor-1 (CSF1) stimulates macrophage recruitment and survival, and its expression is prognostic for mortality in breast cancer [37,38]. Mir-149 directly targets and inhibits CSF1 mRNA in breast cancer cells, limiting macrophage recruitment to the primary tumor. The reduced TAM infiltration has also been associated with decreased lung metastasis formation in a mouse model with orthotopically injected MDA-MB-231 cells [39]. Once in the tumor site, monocytes rapidly differentiate and are polarized into M1 or M2-like macrophages, depending on the stimuli from the TME [6,7]. MiR-149 suppresses the expression of two M2 markers in macrophages, the mannose receptor C-type 1 (MRC1) and arginase-1 (ARG1). Although the authors did not investigate the mechanism, miR-149 possibly acts in a paracrine manner by regulating the production of soluble factors secreted by BC cells that influence macrophage polarization [39].
The miR-200 tumor suppressor family are well-known negative regulators of Epithelial-mesenchymal transition (EMT), and their expression is downregulated in many aggressive cancers, including breast cancer [40]. Williams et al. reported that the restoration of miR-200c expression in BC cells stimulates GM-CSF secretion, which induces M1-macrophage polarization as demonstrated by the upregulation of the M1 marker Nos2 and the downregulation of the M2 marker Arg1 [41]. Indeed, GM-CSF is an important regulator of the pro-inflammatory M1 phenotype [42]. On the contrary, other studies have shown that miR-200 expression is associated with poor prognosis in breast cancer patients [43,44]. Accordingly, Meng et al. reported that miR-200c upregulates PAI-2 expression in BC cells and promotes the secretion of the anti-inflammatory cytokine IL-10, which directly stimulates M2-like polarization [45].
Several cell types present in the TME, such as cancer cells, fibroblasts, and immune cells, produce a high level of IL-6 cytokine, which correlates with low survival rates in breast cancer [46]. IL-6 produced by breast cancer cells can induce M2-macrophage polarization by leading to the inhibition of miR-19a-3p expression in TAMs. Mechanistically, the downregulation of miR-19a-3p releases its inhibitory activity on the Fra1 gene, activating the downstream STAT3 pathway which drives M2-polarization [47].
Wang et al. reported that miR-100 is highly expressed in TAMs from breast cancer and in the surrounding cancer tissue. High expression of miR-100 in TAMs favors M2 TAM polarization by inhibiting the mTOR pathway and leading to the upregulation of the M2 marker CD206 [48].
Tao et al. showed that the expression of the lncRNA linc00514 in breast cancer tissues correlates with tumor aggressiveness and controls TAM polarization via Notch signaling activation [49]. In BC cells, linc00514 induces a STAT3-mediated increase in Jagged1, promoting the secretion of the anti-inflammatory cytokines IL-4 and IL-6, which facilitate M2 polarization by increasing the expression of the M2 markers CD206 and CD163 in TAMs [49].
Xist is a tumor-suppressor and poorly expresses lncRNA in BC cells [50]. Recently, Zhao et al. showed that Xist is also expressed in macrophages, and it participates in M1-phenotype maintenance. Xist functions as a sponge for miR-101, which inhibits the expression of C/EBPα and KLF6, two regulators of M1 macrophage polarization. The knockdown of Xist in M1 macrophages promotes the switch to the M2 phenotype, as demonstrated by the upregulation of the M2 marker CD206, confirming its tumor-suppressor role not only in BC cells but also in TAMs [51]. Several studies have reported the oncogenic function of miR-181 in breast cancer [52]. MiR-181 can be released into BC cell-derived exosomes, and promotes the generation of an immunosuppressive environment by enhancing the infiltration of myeloid-derived suppressor cells, which play a crucial role in cancer immune escape [53]. In TAMs, miR-181a suppresses the expression of C/EBPα and KLF6, promoting M2-polarization and increasing TAM pro-tumoral behavior [54].
The lncRNA GNAS-AS1 is upregulated in ER+ breast cancer tissues and TAMs from BC tissues. The overexpression of GNAS-AS1 facilitates M2 macrophage polarization in vitro by directly sponging miR-433-3p, a negative regulator of GATA3, which induces M2-macrophage polarization and promotes anti-inflammatory activity [55]. GATA3 is also targeted and inhibited by miR-720, an anti-oncogenic miRNA downregulated in TAMs from metastatic breast cancer patients. The restoration of miR-720 expression in M2-macrophages drastically reduces the production of anti-inflammatory cytokines and the ability of macrophages to stimulate BC cell migration [56].
NcRNAs have also been studied as drugs to re-educate TAMs in vivo [57]. Huang et al. constructed a nanoparticle loaded with miRNA mimic let-7b, which can bind to the TLR7 on TAM membrane. In vitro, let-7b acts as an agonist of TLR7 and inhibits the secretion of IL-10, a potent immunosuppressive cytokine that induces M2-like polarization. Once delivered in vivo in a mouse model of breast cancer, the let-7b-complex is released in the tumor acidic microenvironment and limits tumor growth by reversing M2-like TAM phenotypes. Moreover, let-7b can also activate the TAM-induced Th1 response in the treated mice, restoring the anti-tumor immune response [58]. Figure 1 and Table 1 and Table 2 recapitulate the role of miRNAs and lncRNAs in TAM recruitment and polarization in breast cancer.
3. Non-Coding RNAs in the Regulation of BC Cells and TAM Metabolic Reprogramming
Cancer cells profoundly alter their metabolism to cope with the high biosynthetic demand of proliferating cells and adopt aerobic glycolysis, also known as the Warburg effect, to enhance glucose uptake and utilization [81]. In aerobic glycolysis, glucose is processed into lactate even in the presence of oxygen, providing ATP and building blocks very quickly to support tumor cell survival and proliferation [81,82]. In this context, TAM metabolism is also reprogrammed to sustain the newly developed tumor microenvironment [83,84]. Despite TAM phenotype changes over time during tumor progression, M1-like macrophages generally display a highly glycolytic metabolism associated with lactate secretion and ROS production. Instead, M2-like macrophages typically rely on oxidative phosphorylation and fatty acid oxidation [83,84]. Cancer cells and TAMs establish a mutual metabolic communication, with cancer cells promoting the M2-like phenotype through the secretion of lactate, and TAMs increasing glucose availability for tumor cells [84]. Non-coding RNAs are crucial mediators of this interplay [85,86].
Cancer cells increase HIF-1α protein levels in response to hypoxia to counteract cellular stress. HIF-1α is well known for its ability to modify cancer cell metabolism by inducing the overexpression of different glycolytic enzymes [87]. In healthy cells, PHD2 rapidly hydroxylates and degrades HIF-1α through the ubiquitin/proteasome pathway [88]. However, in cancer cells, HIF-1α degradation is prevented, leading to cancer cell metabolic switch and tumor progression [89]. Chen et al. demonstrated that TAMs isolated from breast cancer tissue release extracellular vesicles (EVs) containing the lncRNA HISLA, which enters tumor cells and stabilizes HIF-1α leading to aerobic glycolysis upregulation. Mechanistically, HISLA functions by directly interacting with PHD2 and inhibiting its hydroxylase activity on HIF-1α, leading to its stabilization [77].
Lactate dehydrogenase B (LDHB) is a glycolysis-related enzyme that catalyzes the interconversion of pyruvate and lactate, whose function in macrophages has been poorly investigated [59]. Frank et al. demonstrated that miR-375 derived from breast cancer cells is captured by TAMs, where it reduces LDHB expression levels, likely by directly targeting its transcript. The decrease in LDHB leads to the upregulation of aerobic glycolysis and lactate production in TAMs. The downregulation of LDHB also results in the activation of SREBP2, an enzyme that regulates cholesterol synthesis. The increased production of lactate and sterol by TAMs in the TME boosts tumor progression by stimulating cancer cell proliferation and survival. Therefore, miR-375 can be a promising target to selectively interfere with TAM tumor-promoting functions by reverting their metabolic status [59].
MiR-503-3p is an miRNA upregulated in breast cancer patients with a poor prognosis [90]. Huang et al. reported that exosomal miR-503-3p secreted by TAMs directly targets DACT2 transcript, a tumor suppressor gene acting as an antagonist of β-catenin in BC cells. The inhibition of miR-503-3p upregulates DACT2 and inhibits Wnt/ β-catenin pathway in BC cells, resulting in glycolysis suppression and mitochondrial oxidative phosphorylation increase [60].
The inhibition of miR-33 regulates macrophage metabolism and M2-polarization by reducing glycolysis and promoting Fatty Acid Oxidation (FAO) in atherosclerosis [91]. Moradi-Chaleshtori et al. employed the delivery of 4T1 BC cell-derived exosomes containing miR-33 to macrophages to regulate their polarization. MiR-33 delivery resulted in macrophage polarization switch from M2 to M1 phenotype, and the upregulation of the M1 markers Nos2, Irf5, CD86, TNF-α, and IL-1β. The delivery of miR-33 induces M1 macrophage polarization also by regulating TAM metabolism. These M1-reprogrammed macrophages acquired the ability to inhibit 4T1 cell migration and invasion, identifying miR-33 delivery as a potential drug treatment for breast cancer [61].
Adipocyte/macrophage fatty acid-binding proteins (FABPs) are lipid chaperons that integrate inflammatory and metabolic responses in macrophages [92]. The upregulation of FABPs increases the pro-tumoral activity of a subtype of TAMs (CD11b+F4/80+Ly6C-MHCII-CD11c-) by upregulating the IL-6/STAT3 pathway via the modulation of NFkB/miR-29b. Mechanistically, FABPs downregulate miR-29b in TAMs via an increased NFkB activity, releasing the inhibitory effect of miR-29b on the pro-tumoral factor STAT3 and inducing macrophage M2 polarization. Therefore, the authors proposed FABPs as new prognostic markers and potential therapeutic targets in breast cancers [62].
Breast cancer cells secrete miR-122, which suppresses glucose metabolism by reducing the glycolytic enzyme pyruvate kinase expression in the premetastatic niche. In this manner, miR-122 prepares the environment for the tumor cell arrival by increasing glucose availability and thus favoring cancer cell proliferation and dissemination. Indeed, the authors showed that miR-122 secreted from the primary tumor could affect glucose metabolism in the lungs and brain before metastases onset [63]. In this context, miR-122 may also influence macrophage metabolism and affect their pro-tumoral phenotype in the premetastatic niche. Figure 2, Table 1, and Table 2 summarize the role of miRNAs and lncRNAs in TAM and BC cell metabolic reprogramming during cancer progression.
4. Non-Coding RNAs in the Regulation of TAM-Induced Angiogenesis
The formation of new blood vessels is indispensable for the successful growth of tumor cells, which require a considerable supply of nutrients, oxygen, and other factors to sustain a high growth rate [93,94]. The increased blood vessel density in the tumor environment allows cancer cells to intravasate and enter circulation, the primary event for metastasis formation [93,94]. The direct correlation between TAM presence and a pro-angiogenic environment has been extensively documented in breast cancer [95,96]. In this context, cancer cells reprogram TAMs to a pro-angiogenic phenotype to support all the stages for blood vessel formation. Indeed, TAMs promote ECM remodeling through metalloproteinase secretion and stimulate endothelial cell proliferation and migration by secreting growth factors such as EGF, VEGF, and pro-angiogenic cytokines such as TNFα, CCL8, and IL-8 [95,97]. NcRNAs control TAM and BC cell angiogenic properties by regulating the expression of growth factors and the release of cytokines in the TME [97,98].
Our group recently demonstrated that ID4 expression in breast cancer cells drives TAM reprogramming by increasing the expression of two pro-angiogenic factors, granulin (GRN) and HIF. We demonstrated that ID4 expression in BC cells mediates the release of VEGF, which acts in a paracrine manner and promotes the downregulation of the anti-angiogenic miR-107, miR-15b, and miR-195 in macrophages. The downregulation of these miRNAs leads to GRN and HIF-1 expression in macrophages, promoting their pro-angiogenic abilities and breast cancer progression [64,99,100].
M2 macrophages also secrete factors that regulate BC cell pro-angiogenic activity. Dong et al. demonstrated that VEGF secreted by M2 macrophages induces the upregulation of the lncRNA PCAT6 in human BC cells. PCAT6 induces the expression of VEGFR2, leading to the activation of the Akt/mTOR pathway. Mechanistically, PCAT6 recruits the ubiquitin protease USP14 on VEGFR2 inhibiting its proteasome-mediated degradation. The activation of the Akt/mTOR pathway in BC cells triggers a pro-angiogenic response, both in vivo and in vitro, facilitating their tumorigenic behavior [78].
The administration of ncRNAs has emerged as a potential therapeutic strategy to stop tumor progression through angiogenesis suppression [101]. MiR-29b is a tumor suppressor miRNA in many cancers, where it regulates cell proliferation, apoptosis, and differentiation, and its expression also decreases in breast cancer tissue compared with normal mammary tissue [65]. Li et al. showed that the systemic administration of miR-29b in a mouse model of breast cancer significantly suppresses tumor growth by reducing angiogenesis. In BC cells, miR-29b downregulates Akt3 protein expression, which is known to induce VEGF and to promote angiogenesis. At the same time, the authors observed that the TAM infiltration was significantly reduced after miR-29b administration, indicating that miR-29b directly or indirectly acts on macrophage recruitment. The reduced number of TAMs further mitigates the secretion of pro-angiogenic factors in the TME [65].
MiR-155 is a well-known oncomiRNA induced by hypoxia and VEGF, which can promote TAM recruitment in breast cancer [102]. Kong et al. reported that the overexpression of miR-155 induces angiogenesis in a mouse model of breast cancer by directly targeting VHL mRNA. VHL targets the members of HIFα family; thus, its downregulation results in HIF1α and HIF2α expression and the induction of angiogenesis. In addition, miR-155 overexpression results in a massive TAM infiltration, although the mechanisms regulating this process have not been clarified yet [66]. TAMs can, in turn, produce miR-155 and miR-155-stimulating factors, such as VEGF, causing a positive feedback loop that boosts angiogenesis [103,104].
MiR-21 and miR-29a are two potent pro-angiogenic factors involved in TAM reprogramming. Mathsyaraja et al. showed that these factors are upregulated in a CSF1-ETS2-dependent pathway in infiltrating myeloid cells from a metastatic model of murine breast cancer. MiR-21 and miR-29a mediate the pro-angiogenic switch in TAMs by downregulating the expression of the anti-angiogenic genes Pdcd4, Spry1, Timp3 Col4a2, and Sparc. The expression of miR-21 and miR-29a also correlates with the upregulation of M2 markers in macrophages and with an increased TAM pro-tumoral potential [67]. Table 1 and Table 2 outline the roles of miRNAs and lncRNAs in TAM and BC cell pro-angiogenic activity during breast cancer progression.
5. Non-Coding RNAs in the Crosstalk between TAMs and BC Cells during Metastasis Formation
The metastatic spreading of cancer cells requires the cooperation between cancer cells and the TME [1]. Before metastasis onset, cancer cells acquire a motile mesenchymal phenotype by undergoing EMT, entering blood vessels, and finally extravasating and invading other tissues where they need to find a favorable niche to grow [105]. TAMs foster metastasis formation by promoting cancer cell EMT, angiogenesis and ECM remodeling, increasing the chances for cancer cells to extravasate and enter the circulation. Furthermore, TAMs provide factors to limit the immune response in the pre-metastatic niche to prepare the microenvironment for the arrival of metastatic cancer cells. [13,106]. Sanchez-Gonzalez et al. showed that miR-149 downregulation in BC cells promotes the secretion of CSF1, a cytokine that stimulates the secretion of EGFR ligands by TAMs. EGFR ligands then bind to the receptor expressed by cancer cells and activate the EGFR pathway, increasing BC cells’ malignant behavior and resulting in lung metastasis formation in an SCID mouse orthotopically injected with human BC cells. MiR-149 downregulation also favors tumor progression by increasing TAM recruitment to the tumor site [39]. MiR-100 expression in TAMs increases IL-1ra secretion via the modulation of the mTOR-Stat5a pathway. The authors showed that IL-1ra increases cancer cell invasive ability and stemness by inducing the activation of the Hedgehog signal pathway. In vivo experiments finally showed that miR-100 inhibition significantly reduced lung metastasis formation in a 4T1 BC mouse model [48]. Long noncoding RNA associated with breast cancer brain metastasis (lnc-BM) is upregulated in metastatic cells where it induces the expression of CCL2 via the activation of the JAK2/STAT3 pathway. The secretion of CCL2 recruits TAMs, which secrete oncostatin M (OSM) and IL-6, inducing an additional activation of the JAK2/STAT3 pathway, activating a positive feedback loop. The activation of the JAK/STAT pathway in cancer cells facilitates cell adhesion to capillaries and extravasation to the brain through the upregulation of the endothelial adhesive molecule ICAM. The treatment of a mouse xenograft model of BC with nanoparticles carrying lnc-BM siRNA inhibited brain metastasis formation [36].
Tumors are often characterized by a persistent and non-resolutive inflammation that correlates with a poor prognosis [107]. Guo et al. demonstrated that the delivery of miR-183-5p from 4T1 BC cells to macrophages mediated by exosomes increases the secretion of pro-inflammatory cytokines IL-1b, IL-6, and TNF-a from macrophages. Mechanistically, miR-183-5p targets PPP2CA in TAMs, leading to the upregulation of the NF-kB pathway, which is responsible for the activation of the pro-inflammatory pathway. The release of pro-inflammatory cytokines in the TME is detrimental and results in tumor progression and lung metastasis. The authors finally showed that the injection of BC cells with downregulated miR-183-5p in a mouse model results in a significant reduction in lung metastasis [68]. Another study showed that exosomal miR-183-5p transmission from BC cells to TAMs targets and inhibits the H3K27 histone demethylase KDM6B, inhibiting the expression of M1-related pro-inflammatory genes. The co-administration of BC cells and TAMs pre-treated with exosomes from cells overexpressing miR-183-5p promoted lung metastasis in vivo, highlighting the ability of this miRNA to induce TAM-mediated metastasis formation [69].
Progranulin (PGRN) is a secreted glycoprotein upregulated in many human cancers [108]. Recently, it has been shown that TAMs isolated from PGRN-/- mice harboring breast cancer release exosomes that can affect BC cell behavior by reducing EMT and invasive ability. These TAM-released exosomes contained high levels of miR-5100, which could be taken up by BC cells targeting CXCL12, a chemokine known to promote BC cell invasiveness [70]. In an in vivo model of PY8119 breast cancer, the absence of PGRN resulted in a reduced grade of TAM infiltration and reduced lung metastasis formation [109].
TAMs are a primary source of CCL18, a chemokine associated with breast cancer progression and poor prognosis [110]. Lin et al. demonstrated that CCL18 decreases miR-98 expression in BC cells via Lin28b/ NF-κB activation, promoting the activation of the EMT pathway. To investigate the role of CCL18 on metastasis formation, the authors injected CCL18 in a mouse model harboring a breast tumor derived from MDA-MB-231 cells and observed an induction of lung and liver metastasis formation, together with the downregulation of miR-98 [71]. Another study showed that the ectopic overexpression of miR-181b in TAMs could counteract the pro-tumoral effects of TAM-released CCL18 by inactivating the NF-κB pathway in BC cells in vitro [72]. Let-7a is a downregulated miRNA in breast cancer that can inhibit the effects of CCL18 in BC cells by downregulating Lin28 expression. Therefore, let-7a delivery in BC tissue could be a potential strategy to limit the detrimental effects of TAMs [111]. Figure 3, Table 1, and Table 2 summarize the role of miRNAs and lncRNAs in TAM-induced breast cancer metastasis.
6. The Role of miRNAs and lncRNAs in TAM-Induced Chemoresistance in Breast Cancer
Drug resistance is the leading cause of relapse and death in cancer [112]. TAMs may play a significant role in response to chemotherapy by accumulating in the tumor tissue and mediating the pro-regenerative and anti-apoptotic responses that confer resistance to therapies to cancer cells [113]. In a recent study, Liu et al. reported that IRENA is the most upregulated lncRNA in TAMs after neoadjuvant chemotherapy in breast cancer patients. The authors demonstrated that chemotherapy activates the inflammatory response and upregulates the Jak1–STAT1-induced IRENA lncRNA. IRENA then induces the pro-inflammatory NF-κB pathway in TAMs by promoting the dimerization and phosphorylation of PKR, a protein that activates several inflammatory-related pathways [114]. Although inflammatory M1 macrophages are often associated with antitumor activity, the activation of the NF-κB pathway in the context of a chemotherapeutic treatment promoted TAM-mediated cancer cell chemoresistance. The authors showed that the IRENA knockdown in the PyMT murine model of breast cancer improved the cancer response to chemotherapy treatment [79].
MiR-770 is selectively overexpressed in chemo-sensitive breast cancer tissues and mediates doxorubicin sensitivity by regulating gene expression in BC cells and interfering with the TME. First, miR-770 promotes BC cells apoptosis after treatment with doxorubicin and reduces metastasis formation by directly targeting STMN1mRNA and inhibiting the EMT pathway. Secondly, BC cells can transfer miR-770 to TAMs, where it upregulates the M1 markers MCP-1, iNOS and CD80 while decreasing the expression of M2-macrophage markers. The M1-reprogrammed macrophages helped to reverse the BC cell’s resistance to doxorubicin and sensitize them to cell death, underlining the importance of the TME in the chemoresistance process [73].
The expression of the oncomiRNA miR-21 has been associated with trastuzumab and chemotherapy resistance in HER2-positive breast cancers [74]. MiR-21 mediates chemoresistance by acting in BC cells and TAM recruitment. In BC cells, miR-21 promotes survival by targeting PTEN and PDCD. In the TME, PTEN downregulation increased the secretion of IL-6 from BC cells, which correlated with higher TAM infiltration that contributed to the acquisition of chemoresistance in BC cells [74].
Tan et al. uncovered the role of miR-708 in chemo-resistant breast cancer stem cells. MiR-708 is downregulated in chemo-resistant stem cells, where it inhibits self-renewal by targeting the CD47. The overexpression of miR-708 in BC cells with increased chemosensitivity to docetaxel and increased macrophage phagocytic activity, restoring the immune-mediated elimination of chemo-resistant cells [75].
MiR-100 expression in TAMs increases IL-1ra secretion via the mTOR-Stat5a pathway. The release of IL-1ra in the TME facilitates cancer stem cell development via the activation of the Hedgehog pathway and promotes resistance to cisplatin treatment. The inhibition of miR-100 in a 4T1 breast cancer model significantly improved the cisplatin response, resulting in reduced metastasis formation [48].
Breast cancer cells resistant to adriamycin secrete exosomes containing miR-222, which are captured by macrophages, promoting M2 polarization by targeting PTEN and inducing the Akt pathway. By inducing M2 macrophages, chemo-resistant cells modify the TME to sustain tumor growth and metastasis formation [76].
The lncRNA LINC00337 is upregulated in breast cancer cells and promotes paclitaxel resistance by affecting the TAM phenotype. LINC00337 promotes CCL2, IL-13, and M-CSF secretion inducing macrophage recruitment and M2 polarization, as demonstrated by the upregulation of the M2 markers CD163 and ARGM2 macrophages, which then increase the malignant behavior of chemo-resistant cancer cells by promoting their survival and migratory abilities [80]. Table 1 and Table 2 recapitulate the role of ncRNAs in TAM-induced breast cancer chemoresistance.
7. Conclusions
Breast cancer is the most common tumor in women worldwide and a very heterogeneous disease [115]. Although the prognosis of non-metastatic breast cancer is generally favorable, the most aggressive forms remain incurable, and, in the case of chemo-resistant breast cancers, the treatments may alleviate the symptoms rather than eradicate the disease [115]. Therefore, new therapeutic approaches considering the molecular heterogeneities of the different breast cancer subtypes are needed to improve patients’ life expectancies [116]. The immunosuppressive and tumor-promoting tumor microenvironment allows cancer cells to escape the immune-mediated killing mechanisms and provides pro-survival signals, laying the foundations for metastasis formation and chemoresistance [1,117]. TAMs represent one of the primary cell populations in the TME that contribute to each stage of cancer progression [7,118]. Deregulated ncRNAs in BC cells promote TAM recruitment and polarization to the M2 phenotype and educate TAMs to promote cancer progression. At the same time, ncRNA alteration in TAMs affects BC cell behavior [98]. This crosstalk can be mediated by ncRNAs secreted in the TME and collected by nearby cells or by factors released in the TME that control ncRNA expression in other cells. Different studies showed that targeting TAMs is beneficial for breast cancer. Therapies under investigation involve blocking TAM recruitment, eliminating TAMs already present in the TME, or interfering with their polarization status [119].
TAM elimination is challenging, as the drugs used for this purpose may not be selective for macrophages. The manipulation of ncRNA expression could be a potential strategy to interfere with TAM recruitment or to reprogram M2-polarized macrophages [119]. NcRNAs are often released inside exosomes, so one possibility is the use of antibodies to target exosomes from cancer cells that can be recognized for the expression of particular proteins [120]. Another possibility involves the use of small molecules such as Antisense Oligonucleotides (ASOs) or small interfering RNA (siRNA) that inhibit or degrade ncRNA molecules [121]. On the contrary, the delivery of anti-oncogenic ncRNAs through exosomes or nanoparticles might restore altered molecular pathways in cancer [120,122]. Today, the use of RNA-based therapies for cancer treatment is still premature, due to the difficulties in drug delivery and the risk of off-target effects. However, in the future, the manipulation of ncRNA expression, alone or in combination with other treatments, could be a promising therapeutic approach to interfere with TAM pro-tumoral functions in breast cancer and other cancers.
Author Contributions
Conceptualization and writing—original draft preparation, A.B., C.T., G.F., F.F.; writing—review and editing, A.B., C.T., G.F., F.F.; visualization, A.B., C.T.; supervision, G.F., F.F.; project administration, G.F., F.F.; funding acquisition, G.F., F.F. All authors have read and agreed to the published version of the manuscript.
Funding
Please add: This research was funded by: AIRC IG 2018—ID. 21406 project, ‘Progetti Ateneo’ Sapienza University of Rome and PRIN 2017-Prot. 2017TATYMP_003 to F.F.; AIRC IG 2018 —ID. 21434 project to G.F.
Acknowledgments
We thank Martyna Sniegocka for the critical reading of the manuscript and for fruitful discussions. We apologize for not directly citing many crucial references; these references can, however, be found in the cited previous reviews.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhao, J.; Zhang, L.; Wei, F.; Lian, Y.; Wu, Y.; Gong, Z.; Zhang, S.; Zhou, J.; Cao, K.; et al. Role of tumor microenvironment in tumorigenesis. J. Cancer 2017, 8, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Brassart-Pasco, S.; Brézillon, S.; Brassart, B.; Ramont, L.; Oudart, J.-B.; Monboisse, J.C. Tumor Microenvironment: Extracellular Matrix Alterations Influence Tumor Progression. Front. Oncol. 2020, 10, 397. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, D.; Cang, H.; Guo, B. Crosstalk between cancer and immune cells: Role of tumor-associated macrophages in the tumor microenvironment. Cancer Med. 2019, 8, 4709–4721. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef]
- Jayasingam, S.D.; Citartan, M.; Thang, T.H.; Mat Zin, A.A.; Ang, K.C.; Ch’ng, E.S. Evaluating the Polarization of Tumor-Associated Macrophages Into M1 and M2 Phenotypes in Human Cancer Tissue: Technicalities and Challenges in Routine Clinical Practice. Front. Oncol. 2020, 9, 1512. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tan, W.; Wang, C. Tumor-associated macrophage-derived cytokines enhance cancer stem-like characteristics through epithelial–mesenchymal transition. OncoTargets Ther. 2018, 11, 3817–3826. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 1–16. [Google Scholar] [CrossRef]
- Oshi, M.; Tokumaru, Y.; Asaoka, M.; Yan, L.; Satyananda, V.; Matsuyama, R.; Matsuhashi, N.; Futamura, M.; Ishikawa, T.; Yoshida, K.; et al. M1 Macrophage and M1/M2 ratio defined by transcriptomic signatures resemble only part of their conventional clinical characteristics in breast cancer. Sci. Rep. 2020, 10, 16554. [Google Scholar] [CrossRef]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: Platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 1–19. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Müller, M.; Fazi, F.; Ciaudo, C. Argonaute Proteins: From Structure to Function in Development and Pathological Cell Fate Determination. Front. Cell Dev. Biol. 2020, 7, 360. [Google Scholar] [CrossRef] [Green Version]
- Bellissimo, T.; Tito, C.; Ganci, F.; Sacconi, A.; Masciarelli, S.; Di Martino, G.; Porta, N.; Cirenza, M.; Sorci, M.; De Angelis, L.; et al. Argonaute 2 drives miR-145-5p-dependent gene expression program in breast cancer cells. Cell Death Dis. 2019, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Iaiza, A.; Tito, C.; Ianniello, Z.; Ganci, F.; Laquintana, V.; Gallo, E.; Sacconi, A.; Masciarelli, S.; De Angelis, L.; Aversa, S.; et al. METTL3-dependent MALAT1 delocalization drives c-Myc induction in thymic epithelial tumors. Clin. Epigenetics 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tito, C.; Ganci, F.; Sacconi, A.; Masciarelli, S.; Fontemaggi, G.; Pulito, C.; Gallo, E.; Laquintana, V.; Iaiza, A.; De Angelis, L.; et al. LINC00174 is a novel prognostic factor in thymic epithelial tumors involved in cell migration and lipid metabolism. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication. Non-Coding RNA 2018, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, E.W.; Yang, X. microRNA Regulation in Estrogen Receptor-Positive Breast Cancer and Endocrine Therapy. Biol. Proced. Online 2018, 20, 17. [Google Scholar] [CrossRef]
- Sharma, S.; Patnaik, P.K.; Aronov, S.; Kulshreshtha, R. ApoptomiRs of Breast Cancer: Basics to Clinics. Front. Genet. 2016, 7, 175. [Google Scholar] [CrossRef] [Green Version]
- Zare, M.; Bastami, M.; Solali, S.; Alivand, M.R. Aberrant miRNA promoter methylation and EMT-involving miRNAs in breast cancer metastasis: Diagnosis and therapeutic implications. J. Cell. Physiol. 2017, 233, 3729–3744. [Google Scholar] [CrossRef]
- Shimono, Y.; Mukohyama, J.; Nakamura, S.-I.; Minami, H. MicroRNA Regulation of Human Breast Cancer Stem Cells. J. Clin. Med. 2015, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Amelio, I.; Bernassola, F.; Candi, E. Emerging roles of long non-coding RNAs in breast cancer biology and management. Semin. Cancer Biol. 2020, 72, 36–45. [Google Scholar] [CrossRef]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef]
- Curtale, G.; Rubino, M.; Locati, M. MicroRNAs as Molecular Switches in Macrophage Activation. Front. Immunol. 2019, 10, 799. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Vadgama, J.V.; Wang, P. CCL2/CCR2 signaling in cancer pathogenesis. Cell Commun. Signal. 2020, 18, 82. [Google Scholar] [CrossRef]
- Lim, S.Y.; Yuzhalin, A.; Gordon-Weeks, A.N.; Muschel, R.J. Targeting the CCL2-CCR2 signaling axis in cancer metastasis. Oncotarget 2016, 7, 28697–28710. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.-C.; Ebersberger, S.; Fink, A.F.; Lampe, S.; Weigert, A.; Schmid, T.; Ebersberger, I.; Syed, S.N.; Brüne, B. Apoptotic tumor cell-derived microRNA-375 uses CD36 to alter the tumor-associated macrophage phenotype. Nat. Commun. 2019, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, T.; Qian, B.-Z.; Soong, D.; Cassetta, L.; Noy, R.; Sugano, G.; Kato, Y.; Li, J.; Pollard, J.W. CCL2-induced chemokine cascade promotes breast cancer metastasis by enhancing retention of metastasis-associated macrophages. J. Exp. Med. 2015, 212, 1043–1059. [Google Scholar] [CrossRef]
- Wang, S.; Liang, K.; Hu, Q.; Li, P.; Qingsong, H.; Yang, Y.; Yao, J.; Mangala, L.S.; Li, C.; Yang, W.; et al. JAK2-binding long noncoding RNA promotes breast cancer brain metastasis. J. Clin. Investig. 2017, 127, 4498–4515. [Google Scholar] [CrossRef] [Green Version]
- Cannarile, M.A.; Weisser, M.; Jacob, W.; Jegg, A.-M.; Ries, C.H.; Rüttinger, D. Colony-stimulating factor 1 receptor (CSF1R) inhibitors in cancer therapy. J. Immunother. Cancer 2017, 5, 53. [Google Scholar] [CrossRef]
- Elin, R.; Rebecca Dale, U.; Stein Harald, J.; Lill-Tove, B. Macrophage-colony stimulating factor (CSF1) predicts breast cancer progression and mortality. Anticancer Res. 2015, 35, 865. [Google Scholar]
- Sánchez-González, I.; Bobien, A.; Molnar, C.; Schmid, S.; Strotbek, M.; Boerries, M.; Busch, H.; Olayioye, M.A. miR-149 Suppresses Breast Cancer Metastasis by Blocking Paracrine Interactions with Macrophages. Cancer Res. 2020, 80, 1330–1341. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, L. Members of the microRNA-200 family are promising therapeutic targets in cancer. Exp. Ther. Med. 2017, 14, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.M.; Christenson, J.L.; O’Neill, K.I.; Hafeez, S.A.; Ihle, C.L.; Spoelstra, N.S.; Slansky, J.E.; Richer, J.K. MicroRNA-200c restoration reveals a cytokine profile to enhance M1 macrophage polarization in breast cancer. NPJ Breast Cancer 2021, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. GM-CSF-Dependent Inflammatory Pathways. Front. Immunol. 2019, 10, 2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, T.N.M.; Hamar, P.; Guo, C.; Basar, E.; Perdigão-Henriques, R.; Balaj, L.; Lieberman, J. miR-200–containing extracellular vesicles promote breast cancer cell metastasis. J. Clin. Investig. 2014, 124, 5109–5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuomarila, M.; Luostari, K.; Soini, Y.; Kataja, V.; Kosma, V.-M.; Mannermaa, A. Overexpression of MicroRNA-200c Predicts Poor Outcome in Patients with PR-Negative Breast Cancer. PLoS ONE 2014, 9, e109508. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Zhang, R.; Wang, Y.; Zhu, G.; Jin, T.; Li, C.; Zhang, S. miR-200c/PAI-2 promotes the progression of triple negative breast cancer via M1/M2 polarization induction of macrophage. Int. Immunopharmacol. 2019, 81, 106028. [Google Scholar] [CrossRef]
- Masjedi, A.; Hashemi, V.; Hojjat-Farsangi, M.; Ghalamfarsa, G.; Azizi, G.; Yousefi, M.; Jadidi-Niaragh, F. The significant role of interleukin-6 and its signaling pathway in the immunopathogenesis and treatment of breast cancer. Biomed. Pharmacother. 2018, 108, 1415–1424. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Z.; Chen, C.; Liu, Y.; Si, Q.; Chuang, T.-H.; Li, N.; Gomezcabrero, A.; Reisfeld, R.A.; Xiang, R.; et al. MicroRNA-19a-3p inhibits breast cancer progression and metastasis by inducing macrophage polarization through downregulated expression of Fra-1 proto-oncogene. Oncogene 2013, 33, 3014–3023. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, Y.; Guo, J.; He, H.; Mi, X.; Chen, C.; Xie, J.; Wang, S.; Wu, P.; Cao, F.; et al. miR-100 maintains phenotype of tumor-associated macrophages by targeting mTOR to promote tumor metastasis via Stat5a/IL-1ra pathway in mouse breast cancer. Oncogenesis 2018, 7, 1–17. [Google Scholar] [CrossRef]
- Tao, S.; Chen, Q.; Lin, C.; Dong, H. Linc00514 promotes breast cancer metastasis and M2 polarization of tumor-associated macrophages via Jagged1-mediated notch signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 1–17. [Google Scholar] [CrossRef]
- Zheng, R.; Lin, S.; Guan, L.; Yuan, H.; Liu, K.; Liu, C.; Ye, W.; Liao, Y.; Jia, J.; Zhang, R. Long non-coding RNA XIST inhibited breast cancer cell growth, migration, and invasion via miR-155/CDX1 axis. Biochem. Biophys. Res. Commun. 2018, 498, 1002–1008. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, Z.; Ma, R.; Zhang, Y.; Zhao, L.; Yan, Y.; Lv, X.; Zhang, L.; Su, P.; Bi, J.; et al. lncRNA-Xist/miR-101-3p/KLF6/C/EBPα axis promotes TAM polarization to regulate cancer cell proliferation and migration. Mol. Ther.-Nucleic Acids 2021, 23, 536–551. [Google Scholar] [CrossRef]
- Yang, C.; Tabatabaei, S.N.; Ruan, X.; Hardy, P. The Dual Regulatory Role of MiR-181a in Breast Cancer. Cell. Physiol. Biochem. 2017, 44, 843–856. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, W.; Zhang, R.; Liu, P.; Ye, Y.; Yu, W.; Guo, X.; Yu, J. Cancer exosome-derived miR-9 and miR-181a promote the development of early-stage MDSCs via interfering with SOCS3 and PIAS3 respectively in breast cancer. Oncogene 2020, 39, 4681–4694. [Google Scholar] [CrossRef]
- Bi, J.; Zeng, X.; Zhao, L.; Wei, Q.; Yu, L.; Wang, X.; Yu, Z.; Cao, Y.; Shan, F.; Wei, M. miR-181a Induces Macrophage Polarized to M2 Phenotype and Promotes M2 Macrophage-mediated Tumor Cell Metastasis by Targeting KLF6 and C/EBPα. Mol. Ther.-Nucleic Acids 2016, 5, e368. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-Q.; Zhou, Z.-Y.; Dong, X.; Guo, L.; Zhang, K.-J. LncRNA GNAS-AS1 facilitates ER+ breast cancer cells progression by promoting M2 macrophage polarization via regulating miR-433-3p/GATA3 axis. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Zhong, Y.; Yi, C. MicroRNA-720 suppresses M2 macrophage polarization by targeting GATA. Biosci. Rep. 2016, 36. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Pioppini, C.; Ozpolat, B.; Calin, G.A. Non-coding RNAs regulation of macrophage polarization in cancer. Mol. Cancer 2021, 20, 1–15. [Google Scholar] [CrossRef]
- Huang, Z.; Gan, J.; Long, Z.; Guo, G.; Shi, X.; Wang, C.; Zang, Y.; Ding, Z.; Chen, J.; Zhang, J.; et al. Targeted delivery of let-7b to reprogramme tumor-associated macrophages and tumor infiltrating dendritic cells for tumor rejection. Biomaterials 2016, 90, 72–84. [Google Scholar] [CrossRef]
- Frank, A.-C.; Raue, R.; Fuhrmann, D.C.; Sirait-Fischer, E.; Reuse, C.; Weigert, A.; Lütjohann, D.; Hiller, K.; Syed, S.N.; Brüne, B. Lactate dehydrogenase B regulates macrophage metabolism in the tumor microenvironment. Theranostics 2021, 11, 7570–7588. [Google Scholar] [CrossRef]
- Huang, S.; Fan, P.; Zhang, C.; Xie, J.; Gu, X.; Lei, S.; Chen, Z.; Huang, Z. Exosomal microRNA-503-3p derived from macrophages represses glycolysis and promotes mitochondrial oxidative phosphorylation in breast cancer cells by elevating DACT. Cell Death Discov. 2021, 7, 1–13. [Google Scholar] [CrossRef]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Heidari, N.; Mohammadi-Yeganeh, S.; Hashemi, S.M. Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int. Immunopharmacol. 2020, 90, 107198. [Google Scholar] [CrossRef]
- Hao, J.; Yan, F.; Zhang, Y.; Triplett, A.; Zhang, Y.; Schultz, D.A.; Sun, Y.; Zeng, J.; Silverstein, K.A.; Zheng, Q.; et al. Expression of Adipocyte/Macrophage Fatty Acid–Binding Protein in Tumor-Associated Macrophages Promotes Breast Cancer Progression. Cancer Res. 2018, 78, 2343–2355. [Google Scholar] [CrossRef] [Green Version]
- Fong, M.Y.; Zhou, W.; Liu, L.; Alontaga, A.Y.; Chandra, M.; Ashby, J.; Chow, A.; O’Connor, S.T.F.; Li, S.; Chin, A.R.; et al. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat. Cell Biol. 2015, 17, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Donzelli, S.; Milano, E.; Pruszko, M.; Sacconi, A.; Masciarelli, S.; Iosue, I.; Melucci, E.; Gallo, E.; Terrenato, I.; Mottolese, M.; et al. Expression of ID4 protein in breast cancer cells induces reprogramming of tumour-associated macrophages. Breast Cancer Res. 2018, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.-X.; Permuthwey, J.; Lancaster, J.M.; Coppola, D.M.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 promotes tumour angiogenesis by targeting VHL and is associated with poor prognosis and triple-negative breast cancer. Oncogene 2013, 33, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Mathsyaraja, H.; Thies, K.; Taffany, D.A.; Deighan, C.; Liu, T.; Yu, L.; Fernandez, S.A.; Shapiro, C.L.; Otero, J.P.; Timmers, C.; et al. CSF1-ETS2-induced microRNA in myeloid cells promote metastatic tumor growth. Oncogene 2014, 34, 3651–3661. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Duan, Z.; Zhang, C.; Wang, W.; He, H.; Liu, Y.; Wu, P.; Wang, S.; Song, M.; Chen, H.; et al. Mouse 4T1 Breast Cancer Cell–Derived Exosomes Induce Proinflammatory Cytokine Production in Macrophages via miR-183. J. Immunol. 2020, 205, 2916–2925. [Google Scholar] [CrossRef]
- Xun, J.; Du, L.; Gao, R.; Shen, L.; Wang, D.; Kang, L.; Chen, C.; Zhang, Z.; Zhang, Y.; Yue, S.; et al. Cancer-derived exosomal miR-138-5p modulates polarization of tumor-associated macrophages through inhibition of KDM6B. Theranostics 2021, 11, 6847–6859. [Google Scholar] [CrossRef]
- Yue, S.; Ye, X.; Zhou, T.; Gan, D.; Qian, H.; Fang, W.; Yao, M.; Zhang, D.; Shi, H.; Chen, T. PGRN−/− TAMs-derived exosomes inhibit breast cancer cell invasion and migration and its mechanism exploration. Life Sci. 2020, 264, 118687. [Google Scholar] [CrossRef]
- Lin, X.; Chen, L.; Yao, Y.; Zhao, R.; Cui, X.; Chen, J.; Hou, K.; Zhang, M.; Su, F.; Chen, J.; et al. CCL18-mediated down-regulation of miR98 and miR27b promotes breast cancer metastasis. Oncotarget 2015, 6, 20485–20499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, Y.-X.; Chen, L.-P.; Ji, M.-L. Upregulation of microRNA-181b inhibits CCL18-induced breast cancer cell metastasis and invasion via the NF-κB signaling pathway. Oncol. Lett. 2016, 12, 4411–4418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Liang, Y.; Sang, Y.; Song, X.; Zhang, H.; Liu, Y.; Jiang, L.; Yang, Q. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN. Cell Death Dis. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Bottai, G.; Nuciforo, P.; Di Tommaso, L.; Giovannetti, E.; Peg, V.; Losurdo, A.; Pérez-Garcia, J.; Masci, G.; Corsi, F.; et al. MicroRNA-21 links epithelial-to-mesenchymal transition and inflammatory signals to confer resistance to neoadjuvant trastuzumab and chemotherapy in HER2-positive breast cancer patients. Oncotarget 2015, 6, 37269–37280. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.; Tang, H.; Jiang, X.; Ye, F.; Huang, L.; Shi, D.; Li, L.; Huang, X.; Li, L.; Xie, X.; et al. Metformin mediates induction of miR-708 to inhibit self-renewal and chemoresistance of breast cancer stem cells through targeting CD47. J. Cell. Mol. Med. 2019, 23, 5994–6004. [Google Scholar] [CrossRef]
- Chen, W.-X.; Wang, D.-D.; Zhu, B.; Zhu, Y.-Z.; Zheng, L.; Feng, Z.-Q.; Qin, X.-H. Exosomal miR-222 from adriamycin-resistant MCF-7 breast cancer cells promote macrophages M2 polarization via PTEN/Akt to induce tumor progression. Aging 2021, 13, 10415–10430. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Yang, L.; Liu, J.; Zhang, X.; Zhang, Y.; Tu, Q.; Yin, D.; Lin, D.; Wong, P.P.; et al. Extracellular vesicle-packaged HIF-1α-stabilizing lncRNA from tumour-associated macrophages regulates aerobic glycolysis of breast cancer cells. Nat. Cell Biol. 2019, 21, 498–510. [Google Scholar] [CrossRef]
- Dong, F.; Ruan, S.; Wang, J.; Xia, Y.; Le, K.; Xiao, X.; Hu, T.; Wang, Q. M2 macrophage-induced lncRNA PCAT6 facilitates tumorigenesis and angiogenesis of triple-negative breast cancer through modulation of VEGFR. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Liu, J.; Lao, L.; Chen, J.; Li, J.; Zeng, W.; Zhu, X.; Li, J.; Chen, X.; Yang, L.; Xing, Y.; et al. The IRENA lncRNA converts chemotherapy-polarized tumor-suppressing macrophages to tumor-promoting phenotypes in breast cancer. Nat. Cance 2021, 2, 457–473. [Google Scholar] [CrossRef]
- Xing, Z.; Zhang, M.; Liu, J.; Liu, G.; Feng, K.; Wang, X. LINC00337 induces tumor development and chemoresistance to paclitaxel of breast cancer by recruiting M2 tumor-associated macrophages. Mol. Immunol. 2021, 138, 1–9. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.P.; Sabatini, D.M. Cancer Cell Metabolism: Warburg and Beyond. Cell 2008, 134, 703–707. [Google Scholar] [CrossRef] [Green Version]
- Mehla, K.; Singh, P.K. Metabolic Regulation of Macrophage Polarization in Cancer. Trends Cancer 2019, 5, 822–834. [Google Scholar] [CrossRef]
- Vitale, I.; Manic, G.; Coussens, L.M.; Kroemer, G.; Galluzzi, L. Macrophages and Metabolism in the Tumor Microenvironment. Cell Metab. 2019, 30, 36–50. [Google Scholar] [CrossRef]
- Li, J.; Lu, Z.; Zhang, Y.; Xia, L.; Su, Z. Emerging roles of non-coding RNAs in the metabolic reprogramming of tumor-associated macrophages. Immunol. Lett. 2021, 232, 27–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, Q.; Xia, Q.; Cheng, J.; Huang, Z.; Li, Y.; Chen, P.; Yang, J.; Fan, X.; Liang, Y.; et al. Noncoding RNAs link metabolic reprogramming to immune microenvironment in cancers. J. Hematol. Oncol. 2021, 14, 1–26. [Google Scholar] [CrossRef]
- Jun, J.C.; Rathore, A.; Younas, H.; Gilkes, D.; Polotsky, V.Y. Hypoxia-Inducible Factors and Cancer. Curr. Sleep Med. Rep. 2017, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wielockx, B.; Meneses, A.M. PHD2: From hypoxia regulation to disease progression. Hypoxia 2016, 4, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Soni, S.; Padwad, Y.S. HIF-1 in cancer therapy: Two decade long story of a transcription factor. Acta Oncol. 2017, 56, 503–515. [Google Scholar] [CrossRef]
- Zhao, Z.; Fan, X.; Jiang, L.; Xu, Z.; Xue, L.; Zhan, Q.; Song, Y. miR-503-3p promotes epithelial–mesenchymal transition in breast cancer by directly targeting SMAD2 and E-cadherin. J. Genet. Genom. 2017, 44, 75–84. [Google Scholar] [CrossRef]
- Ouimet, M.; Ediriweera, H.N.; Gundra, U.M.; Sheedy, F.; Ramkhelawon, B.; Hutchison, S.B.; Rinehold, K.; Van Solingen, C.; Fullerton, M.D.; Cecchini, K.; et al. MicroRNA-33–dependent regulation of macrophage metabolism directs immune cell polarization in atherosclerosis. J. Clin. Investig. 2015, 125, 4334–4348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Cao, H.; Kono, K.; Gorgun, C.Z.; Furuhashi, M.; Uysal, K.T.; Cao, Q.; Atsumi, G.; Malone, H.; Krishnan, B.; et al. Adipocyte/macrophage fatty acid binding proteins control integrated metabolic responses in obesity and diabetes. Cell Metab. 2005, 1, 107–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [Green Version]
- Zuazo-Gaztelu, I.; Casanovas, O. Unraveling the Role of Angiogenesis in Cancer Ecosystems. Front. Oncol. 2018, 8, 248. [Google Scholar] [CrossRef]
- Riabov, V.; Gudima, A.; Wang, N.; Mickley, A.; Orekhov, A.; Kzhyshkowska, J. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front. Physiol. 2014, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.Y.; Pollard, J.W. Tumor-Associated Macrophages Press the Angiogenic Switch in Breast Cancer: Figure 1. Cancer Res. 2007, 67, 5064–5066. [Google Scholar] [CrossRef] [Green Version]
- Munir, M.; Kay, M.; Kang, M.; Rahman, M.; Al-Harrasi, A.; Choudhury, M.; Moustaid-Moussa, N.; Hussain, F.; Rahman, S. Tumor-Associated Macrophages as Multifaceted Regulators of Breast Tumor Growth. Int. J. Mol. Sci. 2021, 22, 6526. [Google Scholar] [CrossRef]
- Han, D.; Fang, Y.; Guo, Y.; Hong, W.; Tu, J.; Wei, W. The emerging role of long non-coding RNAs in tumor-associated macrophages. J. Cancer 2019, 10, 6738–6746. [Google Scholar] [CrossRef]
- Donzelli, S.; Sacconi, A.; Turco, C.; Gallo, E.; Milano, E.; Iosue, I.; Blandino, G.; Fazi, F.; Fontemaggi, G. Paracrine Signaling from Breast Cancer Cells Causes Activation of ID4 Expression in Tumor-Associated Macrophages. Cells 2020, 9, 418. [Google Scholar] [CrossRef] [Green Version]
- Turco, C.; Donzelli, S.; Fontemaggi, G. miR-15/107 microRNA Gene Group: Characteristics and Functional Implications in Cancer. Front. Cell Dev. Biol. 2020, 8, 427. [Google Scholar] [CrossRef]
- Song, X.; Guo, Y.; Song, P.; Duan, D.; Guo, W. Non-coding RNAs in Regulating Tumor Angiogenesis. Front. Cell Dev. Biol. 2021, 9, 751578. [Google Scholar] [CrossRef] [PubMed]
- Mattiske, S.; Suetani, R.J.; Neilsen, P.; Callen, D. The Oncogenic Role of miR-155 in Breast Cancer. Cancer Epidemiology Biomarkers Prev. 2012, 21, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Chen, Z.; Ni, Y.; Bian, C.; Huang, J.; Chen, L.; Xie, X.; Wang, J. Tumor-associated macrophages secret exosomal miR-155 and miR-196a-5p to promote metastasis of non-small-cell lung cancer. Transl. Lung Cancer Res. 2021, 10, 1338–1354. [Google Scholar] [CrossRef] [PubMed]
- Bruning, U.; Cerone, L.; Neufeld, Z.; Fitzpatrick, S.F.; Cheong, A.; Scholz, C.C.; Simpson, D.A.; Leonard, M.O.; Tambuwala, M.M.; Cummins, E.P.; et al. MicroRNA-155 Promotes Resolution of Hypoxia-Inducible Factor 1 Activity during Prolonged Hypoxia. Mol. Cell. Biol. 2011, 31, 4087–4096. [Google Scholar] [CrossRef] [Green Version]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate Partners for Tumor Cell Migration, Invasion, and Metastasis. Cell 2006, 124, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 1–46. [Google Scholar] [CrossRef]
- Arechavaleta-Velasco, F.; Perez-Juarez, C.E.; Gerton, G.L.; Diaz-Cueto, L. Progranulin and its biological effects in cancer. Med. Oncol. 2017, 34, 1–11. [Google Scholar] [CrossRef]
- Ahirwar, D.K.; Nasser, M.W.; Ouseph, M.M.; Elbaz, M.; Cuitiño, M.C.; Kladney, R.D.; Varikuti, S.; Kaul, K.; Satoskar, A.R.; Ramaswamy, B.; et al. Fibroblast-derived CXCL12 promotes breast cancer metastasis by facilitating tumor cell intravasation. Oncogene 2018, 37, 4428–4442. [Google Scholar] [CrossRef]
- Chen, J.; Yao, Y.; Gong, C.; Yu, F.; Su, S.; Chen, J.; Liu, B.; Deng, H.; Wang, F.; Lin, L.; et al. CCL18 from Tumor-Associated Macrophages Promotes Breast Cancer Metastasis via PITPNM. Cancer Cell 2011, 19, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, Y.-X.; Zhang, D.-Z.; Fang, X.-J.; Sun, P.-S.; Xue, H.-C. Let-7a mimic attenuates CCL18 induced breast cancer cell metastasis through Lin 28 pathway. Biomed. Pharmacother. 2016, 78, 301–307. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Larionova, I.; Cherdyntseva, N.; Liu, T.; Patysheva, M.; Rakina, M.; Kzhyshkowska, J. Interaction of tumor-associated macrophages and cancer chemotherapy. OncoImmunology 2019, 8, e1596004. [Google Scholar] [CrossRef] [Green Version]
- Kang, R.; Tang, D. PKR-Dependent Inflammatory Signals. Sci. Signal. 2012, 5, pe47. [Google Scholar] [CrossRef] [Green Version]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef]
- Padh, H. Novel therapeutics. Curr. Sci. 2004, 86, 254–256. [Google Scholar] [CrossRef]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 2020, 18, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Cassetta, L.; Fragkogianni, S.; Sims, A.H.; Swierczak, A.; Forrester, L.M.; Zhang, H.; Soong, D.Y.H.; Cotechini, T.; Anur, P.; Lin, E.Y.; et al. Human Tumor-Associated Macrophage and Monocyte Transcriptional Landscapes Reveal Cancer-Specific Reprogramming, Biomarkers, and Therapeutic Targets. Cancer Cell 2019, 35, 588–602.e10. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Wang, J.; Lu, D.; Xu, X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Signal Transduct. Target. Ther. 2021, 6, 1–12. [Google Scholar] [CrossRef]
- Li, C.; Ni, Y.-Q.; Xu, H.; Xiang, Q.-Y.; Zhao, Y.; Zhan, J.-K.; He, J.-Y.; Li, S.; Liu, Y.-S. Roles and mechanisms of exosomal non-coding RNAs in human health and diseases. Signal Transduct. Target. Ther. 2021, 6, 1–31. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics — challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Ingenito, F.; Roscigno, G.; Affinito, A.; Nuzzo, S.; Scognamiglio, I.; Quintavalle, C.; Condorelli, G. The Role of Exo-miRNAs in Cancer: A Focus on Therapeutic and Diagnostic Applications. Int. J. Mol. Sci. 2019, 20, 4687. [Google Scholar] [CrossRef] [Green Version]
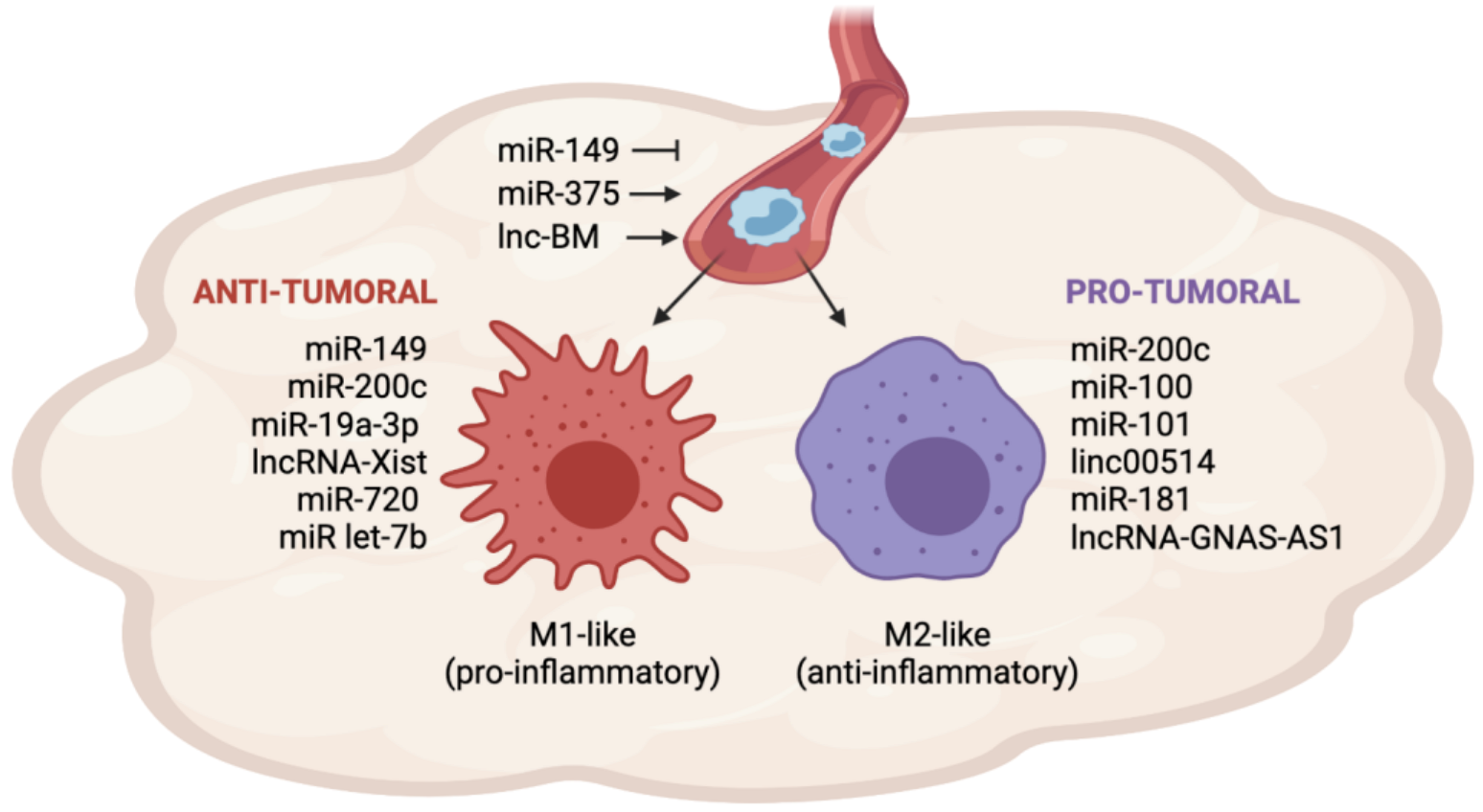
Figure 1.
miRNAs and lncRNAs involved in TAM recruitment and polarization in breast cancer. The expression of miRNAs and lncRNAs in BC cells influences TAM recruitment by regulating the secretion of cytokines and chemokines in the TME. MiR-149 inhibits monocyte recruitment, whereas miR-375 and lnc-BM promote monocyte recruitment. Once in the tumor site, monocytes can differentiate into anti-tumoral (M1-like) or pro-tumoral (M2-like) macrophages. The expression of miR-200c, miR-100, miR-101, linc00514, miR-181 and lncRNA GNAS-AS1 in BC cells or TAMs promotes the differentiation into the M2-like phenotype. Conversely, the expression of miR-149, miR-200c, miR-19a-3p, lncRNA-Xist, miR-720, and miR-let7b in BC cells or TAMs promotes the differentiation into the M1-like phenotype.
Figure 1.
miRNAs and lncRNAs involved in TAM recruitment and polarization in breast cancer. The expression of miRNAs and lncRNAs in BC cells influences TAM recruitment by regulating the secretion of cytokines and chemokines in the TME. MiR-149 inhibits monocyte recruitment, whereas miR-375 and lnc-BM promote monocyte recruitment. Once in the tumor site, monocytes can differentiate into anti-tumoral (M1-like) or pro-tumoral (M2-like) macrophages. The expression of miR-200c, miR-100, miR-101, linc00514, miR-181 and lncRNA GNAS-AS1 in BC cells or TAMs promotes the differentiation into the M2-like phenotype. Conversely, the expression of miR-149, miR-200c, miR-19a-3p, lncRNA-Xist, miR-720, and miR-let7b in BC cells or TAMs promotes the differentiation into the M1-like phenotype.

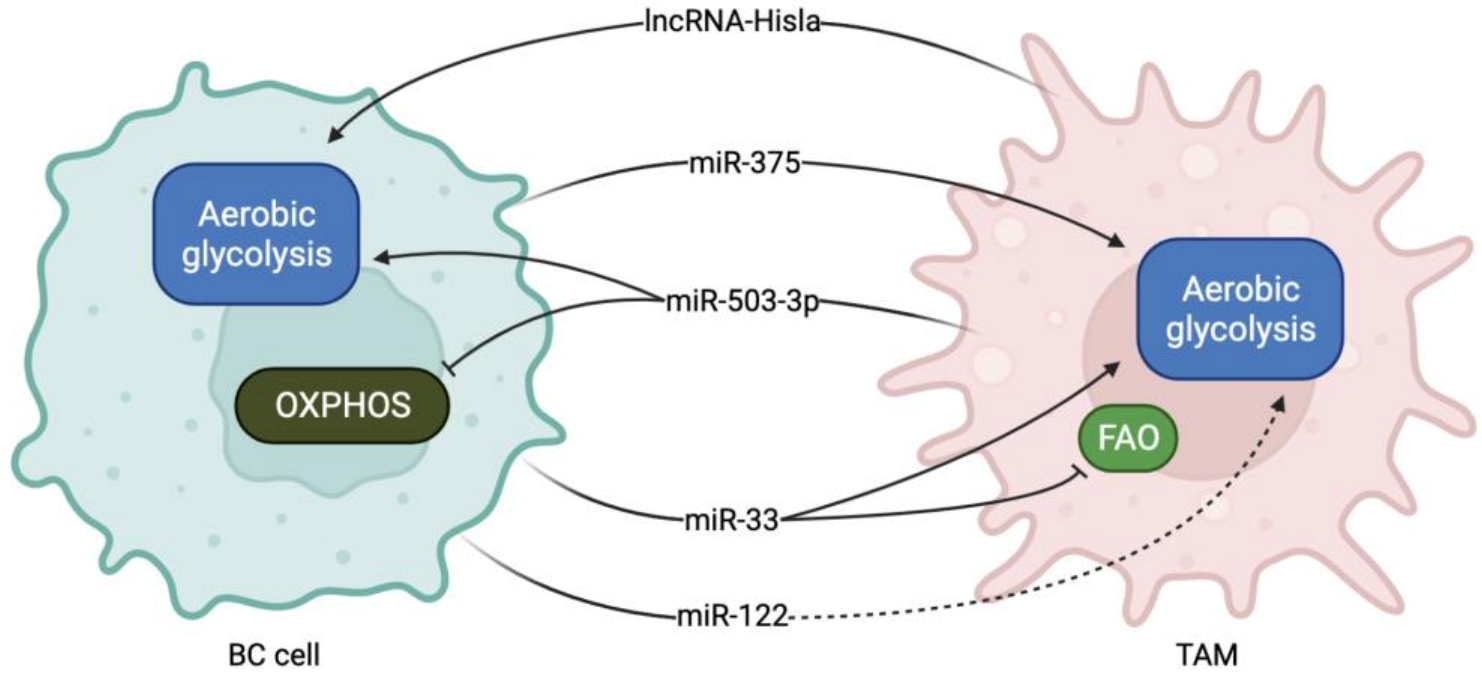
Figure 2.
miRNAs and lncRNAs involved in TAM-BC crosstalk during metabolism reprogramming. LncRNA Hisla and miR-503-3p are secreted from TAMs and collected by BC cells, where they promote aerobic glycolysis and suppress OXPHOS. Instead, miR-375, miR-33 and miR-122 are secreted by BC cells and promote glycolysis upregulation and FAO inhibition in TAMs.
Figure 2.
miRNAs and lncRNAs involved in TAM-BC crosstalk during metabolism reprogramming. LncRNA Hisla and miR-503-3p are secreted from TAMs and collected by BC cells, where they promote aerobic glycolysis and suppress OXPHOS. Instead, miR-375, miR-33 and miR-122 are secreted by BC cells and promote glycolysis upregulation and FAO inhibition in TAMs.

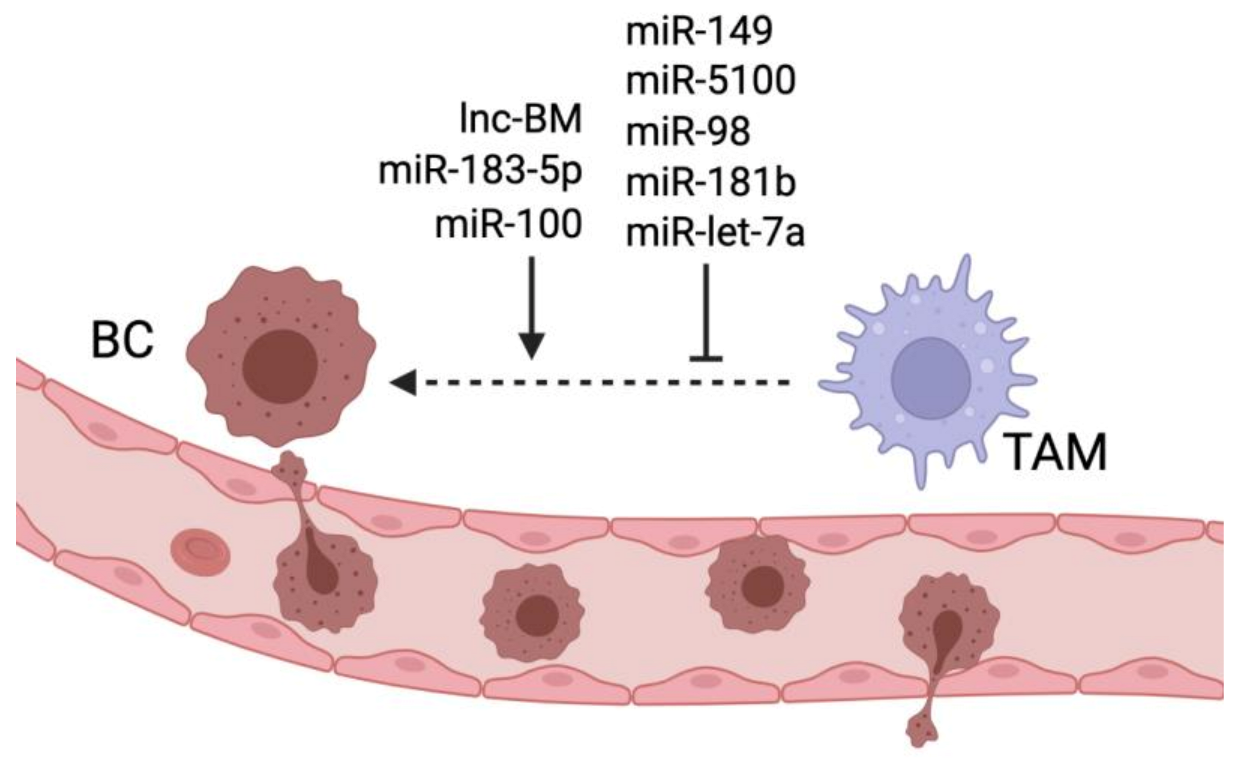
Figure 3.
miRNAs and lncRNAs regulating TAM-induced breast cancer metastasis. Lnc-BM, miR-183-5p, and miR-100 promote TAM ability to stimulate breast cancer metastasis, while miR-149, miR-5100, miR-98, miR-181b and let-7a inhibit TAM- mediated metastasis formation.
Figure 3.
miRNAs and lncRNAs regulating TAM-induced breast cancer metastasis. Lnc-BM, miR-183-5p, and miR-100 promote TAM ability to stimulate breast cancer metastasis, while miR-149, miR-5100, miR-98, miR-181b and let-7a inhibit TAM- mediated metastasis formation.


{kind=link}
{kind=link}
{kind=link}
Table 1.
miRNAs involved in BC cells-TAM crosstalk during BC progression.
| microRNA | Expression | Regulated Pathways | Biological Function | Reference |
|---|---|---|---|---|
| miR-375 | Transferred from apoptotic BC cells to TAMs | Induces CCL2 secretion in BC cells and TNS3 and PXN downregulation in TAMs | Promotes TAM migration and recruitment | [34] |
| miR-149 | Brain metastatic BC cells | Directly targets and inhibits CSF1 | Limits TAM recruitment and lung metastasis; promotes M1 polarization | [39] |
| miR-200c | BC cells | Induces GM-CSF secretion | Induces M1 polarization | [41] |
| miR-200c | BC cells | Induces nPAI-2-mediated IL-10 secretion | Induces M2 polarization | [45] |
| miR-19a-3p | TAMs | Inhibits Fra1/ STAT3 pathway | Inhibits M2 polarization | [47] |
| miR-100 | TAMs | Inhibits mTOR and induces STAT5-mediated IL1-ra secretion | Induces M2 polarization, lung metastasis and chemoresistance | [48] |
| miR-181a | TAMs | Inhibits C/EBPa and KLF6 expression | Induces M2 polarization | [54] |
| miR-720 | TAMs | Directly targets and inhibits GATA3 | Inhibits M2 polarization | [56] |
| Let-7b | Ectopically administered | Acts as an agonist of TLR7 in TAMs and inhibits IL-10 secretion | Inhibits M2 polarization | [58] |
| miR-375 | Transferred from BC cells to TAMs | Inhibits LDHB expression Induces SREBP2 activation | Induces aerobic glycolysis and lactate production Induces cholesterol synthesis | [59] |
| miR-503-3p | Transferred from TAMs cells to BC cells | Activates Wnt/β-cathenin pathway by directly targeting DACT2 | Induces glycolysis and inhibits OXPHOS | [60] |
| miR-33 | Ectopically administered | Stimulates M1-related cytokine production | Induces M1 polarization. induces glycolysis and inhibits FAO | [61] |
| miR-29b | TAMs | Inhibits STAT3 pathway | Inhibits M2 polarization | [62] |
| miR-122 | BC cells | PK downregulation | Increases glucose availability in the pre-metastatic niche for cancer cells | [63] |
| miR-107 miR-15b | TAMs | Inhibits HIF1 and GRN expression | Inhibits TAM pro-angiogenic functions | [64] |
| miR-29b | BC cells | Inhibits VEGF secretion by downregulating Akt3 | Inhibits angiogenesis | [65] |
| miR-155 | BC cells/ TAMs | Directly targets and inhibits VHL | Induces angiogenesis | [66] |
| miR-21 miR-29a | TAMs | Inhibit Pdcd4, Spry1, Timp3, Col4a2, and Sparc expression | Induces TAM pro-angiogenic reprogamming | [67] |
| miR-183-5p | Transferred from BC cells to TAMs | Directly targets and inhibits PPP2CA inducing NFkB activation | Promotes the formation of a pro-inflammatory TME and induces lung metastasis | [68] |
| miR-183-5p | Transferred from BC cells to TAMs | Inhibits KDM6B histone demethylase | Induces M2 polarization and promotes lung metastasis | [69] |
| miR-5100 | Transferred from TAMs to BC cells in PGRN-/- mice | Inhibits CXCL12 production | Reduces TAM infiltration; reduces EMT and lung metastasis | [70] |
| miR-98 | BC cells | Inhibits EMT | Inhibits lung and liver metastasis | [71] |
| miR-181b | Ectopically administred | Downregulates NFkB pathway | Inhibits lung metastasis | [72] |
| miR-770 | BC cells
Transferred from BC cells to TAMs | Inhibits STMN1 expression Induces M1 genes | Inhibits EMT and promotes cell death Promotes M1 polarization | [73] |
| miR-21 | BC cells | Inhibits PTEN and PDCD4 expression | Induces chemoresistance and TAM infiltration | [74] |
| miR-708 | Chemo-resistant BC stem cells | Directly targets and inhibits CD47 | Inhibits cancer stem cell self-renewal and increases M1 phagocitic activity reducing chemoresistance | [75] |
| miR-222 | Transferred from BC cells to TAMs | Induces Akt pathway via PTEN downregulation | Induces M2 polarization and cancer chemoresistance | [76] |
Table 2.
lncRNAs involved in BC cells-TAM crosstalk during BC progression.
| lncRNA | Expression | Regulated Pathways | Biological Function | References |
|---|---|---|---|---|
| lnc-BM | Brain metastatic BC cells | Upregulates JAK2/STAT3 pathway | Promotes TAM recruitment and brain metastasis formation | [36] |
| linc-00514 | BC cells | Induces STAT3-mediated Jagge1 upregulation | Promotes M2 polarization | [49] |
| lnc-Xist | TAMs | Sponges miR-101 and induces C/EBPα and KLF6 expression | Promotes M1 polarization | [51] |
| lnc-GNAS-AS1 | BC cells/ TAMs | Sponges miR-433-3p and induces GATA3 expression | Promotes M2 polarization | [55] |
| lnc-Hisla | Transferred from TAMs to BC cells | Upregulates HIF1α expression by inhibiting PHD2 | Induces aerobic glycolysis | [77] |
| lnc-PCAT6 | BC cells | Inhibits VEGFR2 degradation | Induces angiogenesis | [78] |
| lnc-IRENA | TAMs | Induces PKR-mediated NFkB pathway activation | Induces BC chemoresistance | [79] |
| linc-00337 | BC cells | Promotes CCL12, IL-13, M-CSF secretion | Induces chemoresistance, and M2 polarization | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Benedetti, A.; Turco, C.; Fontemaggi, G.; Fazi, F. Non-Coding RNAs in the Crosstalk between Breast Cancer Cells and Tumor-Associated Macrophages. Non-Coding RNA 2022, 8, 16. https://doi.org/10.3390/ncrna8010016
AMA Style
Benedetti A, Turco C, Fontemaggi G, Fazi F. Non-Coding RNAs in the Crosstalk between Breast Cancer Cells and Tumor-Associated Macrophages. Non-Coding RNA. 2022; 8(1):16. https://doi.org/10.3390/ncrna8010016
Chicago/Turabian StyleBenedetti, Anna, Chiara Turco, Giulia Fontemaggi, and Francesco Fazi. 2022. "Non-Coding RNAs in the Crosstalk between Breast Cancer Cells and Tumor-Associated Macrophages" Non-Coding RNA 8, no. 1: 16. https://doi.org/10.3390/ncrna8010016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.