Circulating Metabolites Associated with Alcohol Intake in the European Prospective Investigation into Cancer and Nutrition Cohort

 , ,
, ,  , , , , , , , , ,
, , , , , , , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Alcohol Intake and Other Lifestyle Variables
2.3. Alcohol Intake and Other Lifestyle Variables
2.4. Statistical Analysis
2.4.1. Heterogeneity by Sex and Smoking Status
2.4.2. Sensitivity Analyses
3. Results
3.1. Participant Characteristics
3.2. Metabolome Characteristics and PC-PR2
3.3. Associations of Alcohol with Metabolites
3.4. Heterogeneity by Sex and Smoking Status
3.5. Sensitivity Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Status Report on Alcohol and Health; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Baliunas, D.O.; Taylor, B.J.; Irving, H.; Roerecke, M.; Patra, J.; Mohapatra, S.; Rehm, J. Alcohol as a risk factor for type 2 diabetes: A systematic review and meta-analysis. Diabetes Care 2009, 32, 2123–2132. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Roerecke, M. Cardiovascular effects of alcohol consumption. Trends Cardiovasc. Med. 2017, 27, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Ricci, C.; Wood, A.; Muller, D.; Gunter, M.J.; Agudo, A.; Boeing, H.; van der Schouw, Y.T.; Warnakula, S.; Saieva, C.; Spijkerman, A.; et al. Alcohol intake in relation to non-fatal and fatal coronary heart disease and stroke: EPIC-CVD case-cohort study. BMJ 2018. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Su, L.J.; Fiehn, O.; Maruvada, P.; Moore, S.C.; O’Keefe, S.J.; Wishart, D.S.; Zanetti, K.A. The use of metabolomics in population-based research. Adv. Nutr. 2014, 5, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Brennan, L.; Fiehn, O.; Hankemeier, T.; Kristal, B.S.; van Ommen, B.; Pujos-Guillot, E.; Verheij, E.; Wishart, D.; Wopereis, S. Mass-spectrometry-based metabolomics: Limitations and recommendations for future progress with particular focus on nutrition research. Metabolomics 2009, 5, 435–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liesenfeld, D.B.; Habermann, N.; Owen, R.W.; Scalbert, A.; Ulrich, C.M. Review of mass spectrometry-based metabolomics in cancer research. Cancer Epidemiol. Biomark. Prev. 2013, 22, 2182–2201. [Google Scholar] [CrossRef] [PubMed]
- Jaremek, M.; Yu, Z.; Mangino, M.; Mittelstrass, K.; Prehn, C.; Singmann, P.; Xu, T.; Dahmen, N.; Weinberger, K.M.; Suhre, K.; et al. Alcohol-induced metabolomic differences in humans. Transl. Psychiatry 2013, 3, E276. [Google Scholar] [CrossRef] [PubMed]
- Lacruz, M.E.; Kluttig, A.; Tiller, D.; Medenwald, D.; Giegling, I.; Rujescu, D.; Prehn, C.; Adamski, J.; Frantz, S.; Greiser, K.H.; et al. Cardiovascular Risk Factors Associated With Blood Metabolite Concentrations and Their Alterations During a 4-Year Period in a Population-Based Cohort. Circ. Cardiovasc. Genet. 2016, 9, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Pallister, T.; Jennings, A.; Mohney, R.P.; Yarand, D.; Mangino, M.; Cassidy, A.; MacGregor, A.; Spector, T.D.; Menni, C. Characterizing Blood Metabolomics Profiles Associated with Self-Reported Food Intakes in Female Twins. PLoS ONE 2016, 11, e0158568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuper, H.; Tzonou, A.; Kaklamani, E.; Hsieh, C.; Lagiou, P.; Adami, H.; Trichopoulos, D.; Stuver, S.O. Tobacco smoking, alcohol consumption and their interaction in the causation of hepatocellular carcinoma. Int. J. Cancer 2000, 85, 498–502. [Google Scholar] [CrossRef]
- Maasland, D.H.; van den Brandt, P.A.; Kremer, B.; Goldbohm, R.A.; Schouten, L.J. Alcohol consumption, cigarette smoking and the risk of subtypes of head-neck cancer: Results from the Netherlands Cohort Study. BMC Cancer 2014, 14, 187. [Google Scholar] [CrossRef] [PubMed]
- Boffetta, P.; Hashibe, M. Alcohol and cancer. Lancet Oncol. 2006, 7, 149–156. [Google Scholar] [CrossRef]
- Riboli, E.; Kaaks, R. The EPIC Project: Rationale and study design. European Prospective Investigation into Cancer and Nutrition. Int. J. Epidemiol. 1997, 26 (Suppl. 1), S61–S64. [Google Scholar] [CrossRef]
- Riboli, E.; Hunt, K.J.; Slimani, N.; Ferrari, P.; Norat, T.; Fahey, M.; Charrondiere, U.R.; Hemon, B.; Casagrande, C.; Vignat, J.; et al. European Prospective Investigation into Cancer and Nutrition (EPIC): Study populations and data collection. Public Health Nutr. 2002, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Cross, A.J.; Abubakar, M.; Jenab, M.; Aleksandrova, K.; Boutron-Ruault, M.C.; Dossus, L.; Racine, A.; Kuhn, T.; Katzke, V.A.; et al. A Nested Case-Control Study of Metabolically Defined Body Size Phenotypes and Risk of Colorectal Cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC). PLoS Med. 2016, 13, e1001988. [Google Scholar] [CrossRef] [PubMed]
- Stepien, M.; Duarte-Salles, T.; Fedirko, V.; Floegel, A.; Barupal, D.K.; Rinaldi, S.; Achaintre, D.; Assi, N.; Tjonneland, A.; Overvad, K.; et al. Alteration of amino acid and biogenic amine metabolism in hepatobiliary cancers: Findings from a prospective cohort study. Int. J. Cancer 2016, 138, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Fanidi, A.; Muller, D.C.; Bassett, J.K.; Midttun, O.; Vollset, S.E.; Travis, R.C.; Palli, D.; Mattiello, A.; Sieri, S.; et al. Circulating biomarkers of one-carbon metabolism in relation to renal cell carcinoma incidence and survival. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.A.; Fensom, G.K.; Rinaldi, S.; Scalbert, A.; Appleby, P.N.; Achaintre, D.; Gicquiau, A.; Gunter, M.J.; Ferrari, P.; Kaaks, R.; et al. Pre-diagnostic metabolite concentrations and prostate cancer risk in 1077 cases and 1077 matched controls in the European Prospective Investigation into Cancer and Nutrition. BMC Med. 2017, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, M.M.; Rehm, J.; Klipstein-Grobusch, K.; Boeing, H.; Schutze, M.; Drogan, D.; Overvad, K.; Tjonneland, A.; Halkjaer, J.; Fagherazzi, G.; et al. The association of pattern of lifetime alcohol use and cause of death in the European prospective investigation into cancer and nutrition (EPIC) study. Int. J. Epidemiol. 2013, 42, 1772–1790. [Google Scholar] [CrossRef] [PubMed]
- Haftenberger, M.; Schuit, A.J.; Tormo, M.J.; Boeing, H.; Wareham, N.; Bueno-de-Mesquita, H.B.; Kumle, M.; Hjartaker, A.; Chirlaque, M.D.; Ardanaz, E.; et al. Physical activity of subjects aged 50–64 years involved in the European Prospective Investigation into Cancer and Nutrition (EPIC). Public Health Nutr. 2002, 5, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Wareham, N.J.; Jakes, R.W.; Rennie, K.L.; Schuit, J.; Mitchell, J.; Hennings, S.; Day, N.E. Validity and repeatability of a simple index derived from the short physical activity questionnaire used in the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Public Health Nutr. 2003, 6, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haftenberger, M.; Lahmann, P.H.; Panico, S.; Gonzalez, C.A.; Seidell, J.C.; Boeing, H.; Giurdanella, M.C.; Krogh, V.; Bueno-de-Mesquita, H.B.; Peeters, P.H.; et al. Overweight, obesity and fat distribution in 50- to 64-year-old participants in the European Prospective Investigation into Cancer and Nutrition (EPIC). Public Health Nutr. 2002, 5, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Carayol, M.; Licaj, I.; Achaintre, D.; Sacerdote, C.; Vineis, P.; Key, T.J.; Onland Moret, N.C.; Scalbert, A.; Rinaldi, S.; Ferrari, P. Reliability of Serum Metabolites over a Two-Year Period: A Targeted Metabolomic Approach in Fasting and Non-Fasting Samples from EPIC. PLoS ONE 2015, 10, e0135437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siskos, A.P.; Jain, P.; Romisch-Margl, W.; Bennett, M.; Achaintre, D.; Asad, Y.; Marney, L.; Richardson, L.; Koulman, A.; Griffin, J.L.; et al. Interlaboratory Reproducibility of a Targeted Metabolomics Platform for Analysis of Human Serum and Plasma. Anal. Chem. 2017, 89, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Drogan, D.; Wang-Sattler, R.; Prehn, C.; Illig, T.; Adamski, J.; Joost, H.G.; Boeing, H.; Pischon, T. Reliability of serum metabolite concentrations over a 4-month period using a targeted metabolomic approach. PLoS ONE 2011, 6, e21103. [Google Scholar] [CrossRef] [PubMed]
- Fages, A.; Ferrari, P.; Monni, S.; Dossus, L.; Floegel, A.; Mode, N.; Johansson, M.; Travis, R.C.; Bamia, C.; Sánchez-Pérez, M.-J.; et al. Investigating sources of variability in metabolomic data in the EPIC study: The Principal Component Partial R-square (PC-PR2) method. Metabolomics 2014, 10, 1074–1083. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar]
- Newson, R.B. Frequentist q-values for multiple-test procedures. Stata J. 2010, 10, 568–584. [Google Scholar]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef]
- Ruppert, D.; Wand, M.P.; Carroll, R.J. Semiparametric Regression, Cambridge Series in Statistical and Probabilistic Mathematics; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Jann, B.; Gutierrez, R. pspline: Stata Module Providing a Penalized Spline Scatterplot Smoother Based on Linear Mixed Model Technology. Available online: http://ideas.repec.org/c/boc/bocode/s456972.html (accessed on 19 February 2018).
- StataCorp. Stata Statistical Software: Release 12, version 12.1; StataCorp LP: College Station, TX, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Version 3.3.3. 2017. Available online: https://www.R-project.org/ (accessed on 19 February 2018).
- Jenkins, R.W.; Canals, D.; Hannun, Y.A. Roles and regulation of secretory and lysosomal acid sphingomyelinase. Cell Signal. 2009, 21, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.; Honig, S.; Liebisch, G.; Luth, A.; Kleuser, B.; Gulbins, E.; Schmitz, G.; Kornhuber, J. Alterations of plasma glycerophospholipid and sphingolipid species in male alcohol-dependent patients. Biochim. Biophys. Acta 2015, 1851, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wang, J.Y.; Hertervig, E.; Cheng, Y.; Nilsson, A.; Duan, R.D. Activation of neutral sphingomyelinase participates in ethanol-induced apoptosis in Hep G2 cells. Alcohol Alcohol. 2000, 35, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Deaciuc, I.V.; Nikolova-Karakashian, M.; Fortunato, F.; Lee, E.Y.; Hill, D.B.; McClain, C.J. Apoptosis and dysregulated ceramide metabolism in a murine model of alcohol-enhanced lipopolysaccharide hepatotoxicity. Alcohol. Clin. Exp. Res. 2000, 24, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Pietilainen, K.H.; Rog, T.; Seppanen-Laakso, T.; Virtue, S.; Gopalacharyulu, P.; Tang, J.; Rodriguez-Cuenca, S.; Maciejewski, A.; Naukkarinen, J.; Ruskeepaa, A.L.; et al. Association of lipidome remodeling in the adipocyte membrane with acquired obesity in humans. PLoS Biol. 2011, 9, e1000623. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Reimers, E.; Santolaria-Fernandez, F.; Martin-Gonzalez, M.C.; Fernandez-Rodriguez, C.M.; Quintero-Platt, G. Alcoholism: A systemic proinflammatory condition. World J. Gastroenterol. 2014, 20, 14660–14671. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Suzuki, H.; Sakaguchi, S. Common pathogenic mechanism in development progression of liver injury caused by non-alcoholic or alcoholic steatohepatitis. J. Toxicol. Sci. 2007, 32, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Wolk, A.; Vessby, B.; Ljung, H.; Barrefors, P. Evaluation of a biological marker of dairy fat intake. Am. J. Clin. Nutr. 1998, 68, 291–295. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.R.; Li, F.; Sharpe, M.R.; Williams, C.D.; Curry, S.C.; Ma, X.; Jaeschke, H. Circulating Acylcarnitines as Biomarkers of Mitochondrial Dysfunction after Acetaminophen Overdose in Mice and Humans. Arch. Toxicol. 2014, 88, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Krahenbuhl, S.; Reichen, J. Carnitine metabolism in patients with chronic liver disease. Hepatology 1997, 25, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R.K.; Hoppel, C.L. Elevated plasma carnitine in hepatic cirrhosis. Hepatology 1983, 3, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Shambaugh, G.E., 3rd. Urea biosynthesis I. The urea cycle and relationships to the citric acid cycle. Am. J. Clin. Nutr. 1977, 30, 2083–2087. [Google Scholar] [CrossRef] [PubMed]
- De Koning, T.J.; Klomp, L.W. Serine-deficiency syndromes. Curr. Opin. Neurol. 2004, 17, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.P. Why most discovered true associations are inflated. Epidemiology 2008, 19, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yu, B.; Alexander, D.; Steffen, L.M.; Nettleton, J.A.; Boerwinkle, E. Metabolomic patterns and alcohol consumption in African Americans in the Atherosclerosis Risk in Communities Study. Am. J. Clin. Nutr. 2014, 99, 1470–1478. [Google Scholar] [CrossRef] [PubMed]
- Guertin, K.A.; Moore, S.C.; Sampson, J.N.; Huang, W.Y.; Xiao, Q.; Stolzenberg-Solomon, R.Z.; Sinha, R.; Cross, A.J. Metabolomics in nutritional epidemiology: Identifying metabolites associated with diet and quantifying their potential to uncover diet-disease relations in populations. Am. J. Clin. Nutr. 2014, 100, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, P.; Cook, S.; Wang, Q.; Tiainen, M.; Tynkkynen, T.; Kangas, A.J.; Soininen, P.; Laitinen, J.; Viikari, J.; Kahonen, M.; et al. Metabolic profiling of alcohol consumption in 9778 young adults. Int. J. Epidemiol. 2016, 45, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Del Boca, F.K.; Darkes, J. The validity of self-reports of alcohol consumption: State of the science and challenges for research. Addiction 2003, 98, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kaaks, R.; Slimani, N.; Riboli, E. Pilot phase studies on the accuracy of dietary intake measurements in the EPIC project: Overall evaluation of results. European Prospective Investigation into Cancer and Nutrition. Int. J. Epidemiol. 1997, 26 (Suppl. 1), S26–S36. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Kastenmuller, G.; He, Y.; Belcredi, P.; Moller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; Ceglarek, U.; et al. Differences between human plasma and serum metabolite profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef] [PubMed]
- Breier, M.; Wahl, S.; Prehn, C.; Fugmann, M.; Ferrari, U.; Weise, M.; Banning, F.; Seissler, J.; Grallert, H.; Adamski, J.; et al. Targeted metabolomics identifies reliable and stable metabolites in human serum and plasma samples. PLoS ONE 2014, 9, e89728. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
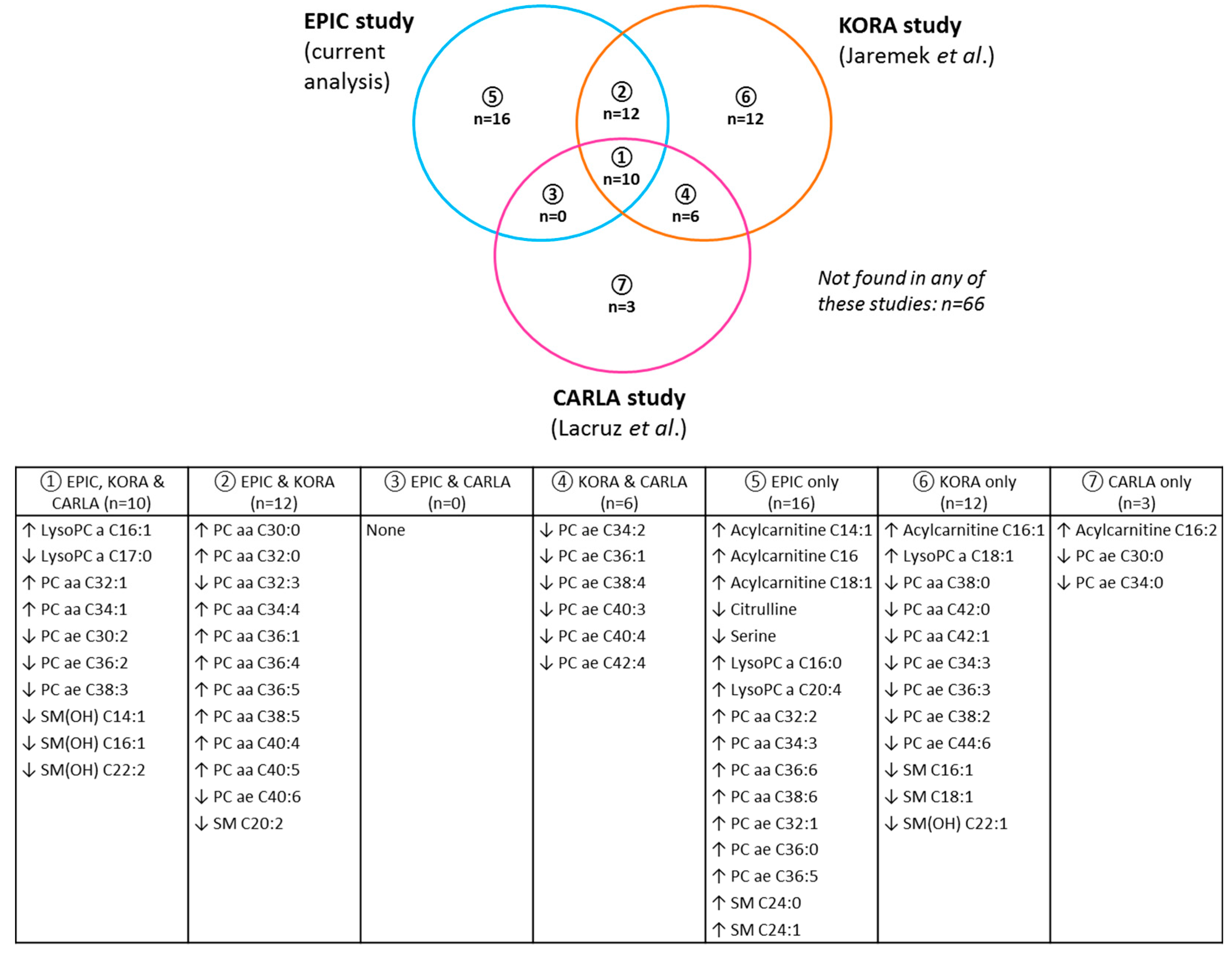{kind=link}
| Discovery Set (n = 1983, 66.7%) | Replication Set (n = 991, 33.3%) | Total Dataset (n = 2974, 100%) | ||||
|---|---|---|---|---|---|---|
| Age, mean (SD) | 58.5 | (7.6) | 58.0 | (7.9) | 58.3 | (7.7) |
| Sex, n (%) | ||||||
| Men | 1497 | (75.5) | 734 | (74.1) | 2231 | (75.0) |
| Women | 486 | (24.5) | 257 | (25.9) | 743 | (25.0) |
| Education level, n (%) b | ||||||
| None/primary | 891 | (46.8) | 408 | (43.1) | 1299 | (45.5) |
| Secondary | 236 | (12.4) | 117 | (12.4) | 353 | (12.4) |
| Technical/professional | 410 | (21.5) | 239 | (25.2) | 649 | (22.8) |
| University or higher | 369 | (19.4) | 183 | (19.3) | 552 | (19.4) |
| Alcohol intake in men (g/day), median (5th, 95th perc) | 14.0 | (0.0, 67.9) | 14.0 | (0.0, 67.9) | 13.7 | (0.0, 64.0) |
| Alcohol intake in women (g/day), median (5th, 95th perc) | 3.2 | (0.0, 26.4) | 2.2 | (0.0, 25.7) | 2.8 | (0.0, 25.7) |
| Categories of alcohol intake, n (%) | ||||||
| Non-drinkers (<0.1 g/day) | 227 | (11.5) | 118 | (11.9) | 345 | (11.6) |
| Light drinkers (0.1–4.9 g/day) | 485 | (24.5) | 241 | (24.3) | 726 | (24.4) |
| Moderate drinkers (5–39.9 g/day) | 1026 | (51.7) | 495 | (50.0) | 1521 | (51.1) |
| Heavy drinkers (≥40 g/day) | 245 | (12.4) | 137 | (13.8) | 382 | (12.8) |
| Body mass index (kg/m2), mean (SD) | 26.8 | (3.8) | 26.9 | (3.6) | 26.8 | (3.7) |
| Physical activity, n (%) c | ||||||
| Inactive | 542 | (27.7) | 281 | (28.7) | 823 | (28.1) |
| Moderately inactive | 652 | (33.4) | 328 | (33.5) | 980 | (33.4) |
| Moderately active | 406 | (20.8) | 214 | (21.9) | 620 | (21.1) |
| Active | 354 | (18.1) | 156 | (15.9) | 510 | (17.4) |
| Smoking status, n (%) d | ||||||
| Current smoker | 445 | (22.7) | 219 | (22.6) | 664 | (22.7) |
| Former smoker | 764 | (38.9) | 365 | (37.7) | 1129 | (38.5) |
| Never smoker | 754 | (38.4) | 385 | (39.7) | 1139 | (38.9) |
| Meat intake (g/day), median (5th, 95th perc) | 106.1 | (34.1, 220.2) | 108.4 | (31.2, 233.0) | 106.6 | (32.9, 222.9) |
| Fish intake (g/day), median (5th, 95th perc) | 28.8 | (2.5, 113.3) | 28.8 | (3.0, 113.0) | 28.8 | (2.5, 113.2) |
| Energy intake (kcal/day), median (5th, 9th perc) | 2187.4 | (1330.0, 3480.4) | 2190.5 | (1304.9, 3451.3) | 2188.3 | (1327.7, 3480.4) |
| Fasting status, n (%) e | ||||||
| ≥6 h | 775 | (40.2) | 391 | (40.4) | 1166 | (40.2) |
| 3–5.9 h | 371 | (19.2) | 193 | (19.9) | 564 | (19.5) |
| <3 h | 784 | (40.6) | 385 | (39.7) | 1169 | (40.3) |
| Sub-study, n (%) | ||||||
| Colorectal cancer controls | 334 | (16.8) | 157 | (15.8) | 491 | (16.5) |
| Kidney cancer controls | 401 | (20.2) | 234 | (23.6) | 635 | (21.4) |
| Hepatobiliary cancer controls | 209 | (10.5) | 118 | (11.9) | 327 | (11.0) |
| Prostate cancer controls | 1039 | (52.4) | 482 | (48.6) | 1521 | (51.1) |
| Discovery Analysis (n = 1983) b | Replication Analysis (n = 991) b | |||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | β | (SE) | p-Value | (FDR q-Value) c | β | (SE) | p-Value | (Bonf.-adj. p-Value) d |
| Acylcarnitine C14:1 | 0.05 | (0.02) | 2.2 × 10−3 2.2 × 10−3 | (4.7 × 10−3) | 0.08 | (0.02) | 6.5 × 10−4 | (4.7 × 10−2) |
| Acylcarnitine C16 | 0.11 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.13 | (0.02) | 4.4 × 10−9 | (3.2 × 10−7) |
| Acylcarnitine C18:1 | 0.06 | (0.02) | 2.4 × 10−4 | (6.2 × 10−4) | 0.08 | (0.02) | 6.7 × 10−4 | (4.8 × 10−2) |
| Citrulline | −0.10 | (0.02) | 8.2 × 10−10 | (5.9 × 10−9) | −0.10 | (0.02) | 1.2 × 10−5 | (8.9 × 10−4) |
| LysoPC a C16:0 | 0.09 | (0.01) | 1.8 × 10−10 | (1.5 × 10−9) | 0.08 | (0.02) | 6.4 × 10−5 | (4.6 × 10−3) |
| LysoPC a C16:1 | 0.11 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.12 | (0.02) | 2.5 × 10−8 | (1.8 × 10−6) |
| LysoPC a C17:0 | −0.09 | (0.01) | 1.0 × 10−10 | (9.5 × 10−10) | −0.11 | (0.02) | 2.0 × 10−7 | (1.4 × 10−5) |
| LysoPC a C20:4 | 0.06 | (0.02) | 3.5 × 10−5 | (1.2 × 10−4) | 0.08 | (0.02) | 1.2 × 10−4 | (8.5 × 10−3) |
| PC aa C30:0 | 0.10 | (0.01) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.12 | (0.02) | 4.3 × 10−8 | (3.1 × 10−6) |
| PC aa C32:0 | 0.12 | (0.01) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.13 | (0.02) | 5.3 × 10−10 | (3.8 × 10−8) |
| PC aa C32:1 | 0.20 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.22 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) |
| PC aa C32:2 | 0.07 | (0.02) | 7.3 × 10−6 | (3.0 × 10−5) | 0.10 | (0.02) | 1.6 × 10−5 | (1.2 × 10−3) |
| PC aa C32:3 | −0.04 | (0.01) | 2.7 × 10−3 | (5.8 × 10−3) | −0.08 | (0.02) | 4.8 × 10−5 | (3.5 × 10−3) |
| PC aa C34:1 | 0.17 | (0.01) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.14 | (0.02) | 2.0 × 10−11 | (1.4 × 10−9) |
| PC aa C34:3 | 0.07 | (0.01) | 4.0 × 10−7 | (2.1 × 10−6) | 0.08 | (0.02) | 4.7 × 10−5 | (3.3 × 10−3) |
| PC aa C34:4 | 0.11 | (0.02) | 1.5 × 10−10 | (1.3 × 10−9) | 0.13 | (0.02) | 1.9 × 10−8 | (1.3 × 10−6) |
| PC aa C36:4 | 0.14 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.15 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) |
| PC aa C36:5 | 0.16 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.17 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) |
| PC aa C36:6 | 0.11 | (0.02) | 2.1 × 10−10 | (1.6 × 10−9) | 0.10 | (0.02) | 6.2 × 10−6 | (4.4 × 10−4) |
| PC aa C38:5 | 0.09 | (0.02) | 1.9 × 10−7 | (1.1 × 10−6) | 0.11 | (0.02) | 6.3 × 10−6 | (4.6 × 10−4) |
| PC aa C38:6 | 0.12 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.09 | (0.02) | 7.8 × 10−5 | (5.6 × 10−3) |
| PC ae C30:2 | −0.05 | (0.01) | 5.0 × 10−5 | (1.7 × 10−4) | −0.07 | (0.02) | 4.4 × 10−5 | (3.1 × 10−3) |
| PC ae C32:1 | 0.06 | (0.02) | 1.8 × 10−5 | (6.8 × 10−5) | 0.08 | (0.02) | 2.1 × 10−4 | (1.5 × 10−2) |
| PC ae C36:0 | 0.16 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | 0.15 | (0.02) | 9.0 × 10−11 | (6.5 × 10−9) |
| PC ae C36:2 | −0.11 | (0.02) | <1.0 × 10−12 | (<1.0 × 10−12) | −0.14 | (0.02) | 1.3 × 10−9 | (9.5 × 10−8) |
| PC ae C36:5 | 0.10 | (0.02) | 1.0 × 10−11 | (1.0 × 10−10) | 0.13 | (0.02) | 5.3 × 10−9 | (3.8 × 10−7) |
| PC ae C38:3 | −0.09 | (0.02) | 5.9 × 10−9 | (3.8 × 10−8) | −0.09 | (0.02) | 8.2 × 10−5 | (5.9 × 10−3) |
| PC ae C40:6 | −0.06 | (0.02) | 2.8 × 10−4 | (7.3 × 10−4) | −0.08 | (0.02) | 5.4 × 10−4 | (3.9 × 10−2) |
| SM C20:2 | −0.06 | (0.01) | 1.4 × 10−6 | (6.8 × 10−6) | −0.08 | (0.02) | 4.8 × 10−7 | (3.5 × 10−5) |
| SM C24:0 | 0.04 | (0.01) | 1.3 × 10−6 | (6.3 × 10−6) | 0.05 | (0.01) | 2.7 × 10−5 | (1.9 × 10−3) |
| SM C24:1 | 0.03 | (0.01) | 2.8 × 10−5 | (1.0 × 10−4) | 0.04 | (0.01) | 5.2 × 10−4 | (3.8 × 10−3) |
| SM(OH) C14:1 | −0.06 | (0.01) | 3.9 × 10−7 | (2.1 × 10−6) | −0.08 | (0.02) | 2.4 × 10−6 | (1.7 × 10−4) |
| SM(OH) C16:1 | −0.05 | (0.01) | 2.6 × 10−6 | (1.1 × 10−5) | −0.08 | (0.02) | 2.1 × 10−7 | (1.5 × 10−5) |
| SM(OH) C22:2 | −0.05 | (0.01) | 1.3 × 10−7 | (8.3 × 10−7) | −0.05 | (0.01) | 2.3 × 10−5 | (1.6 × 10−3) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Roekel, E.H.; Trijsburg, L.; Assi, N.; Carayol, M.; Achaintre, D.; Murphy, N.; Rinaldi, S.; Schmidt, J.A.; Stepien, M.; Kaaks, R.; et al. Circulating Metabolites Associated with Alcohol Intake in the European Prospective Investigation into Cancer and Nutrition Cohort. Nutrients 2018, 10, 654. https://doi.org/10.3390/nu10050654
Van Roekel EH, Trijsburg L, Assi N, Carayol M, Achaintre D, Murphy N, Rinaldi S, Schmidt JA, Stepien M, Kaaks R, et al. Circulating Metabolites Associated with Alcohol Intake in the European Prospective Investigation into Cancer and Nutrition Cohort. Nutrients. 2018; 10(5):654. https://doi.org/10.3390/nu10050654
Chicago/Turabian StyleVan Roekel, Eline H., Laura Trijsburg, Nada Assi, Marion Carayol, David Achaintre, Neil Murphy, Sabina Rinaldi, Julie A. Schmidt, Magdalena Stepien, Rudolf Kaaks, and et al. 2018. "Circulating Metabolites Associated with Alcohol Intake in the European Prospective Investigation into Cancer and Nutrition Cohort" Nutrients 10, no. 5: 654. https://doi.org/10.3390/nu10050654