Muscle Antioxidant Enzymes Activity and Gene Expression Are Altered by Diet-Induced Increase in Muscle Essential Fatty Acid (α-linolenic acid) Concentration in Sheep Used as a Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animal Numbers and Dietary Treatments
2.2. Animal Slaughter, Muscle Sample Collection and Fatty Acid Analysis
2.3. Determination of Antioxidant Enzyme Activity
2.4. Gene Expression Analyses
2.4.1. Sample Preparation, RNA Isolation and cDNA Synthesis
2.4.2. Real-Time Quantitative PCR
2.5. Statistical Analyses
3. Results
3.1. Influence of Diet on Muscle Fatty Acid Composition
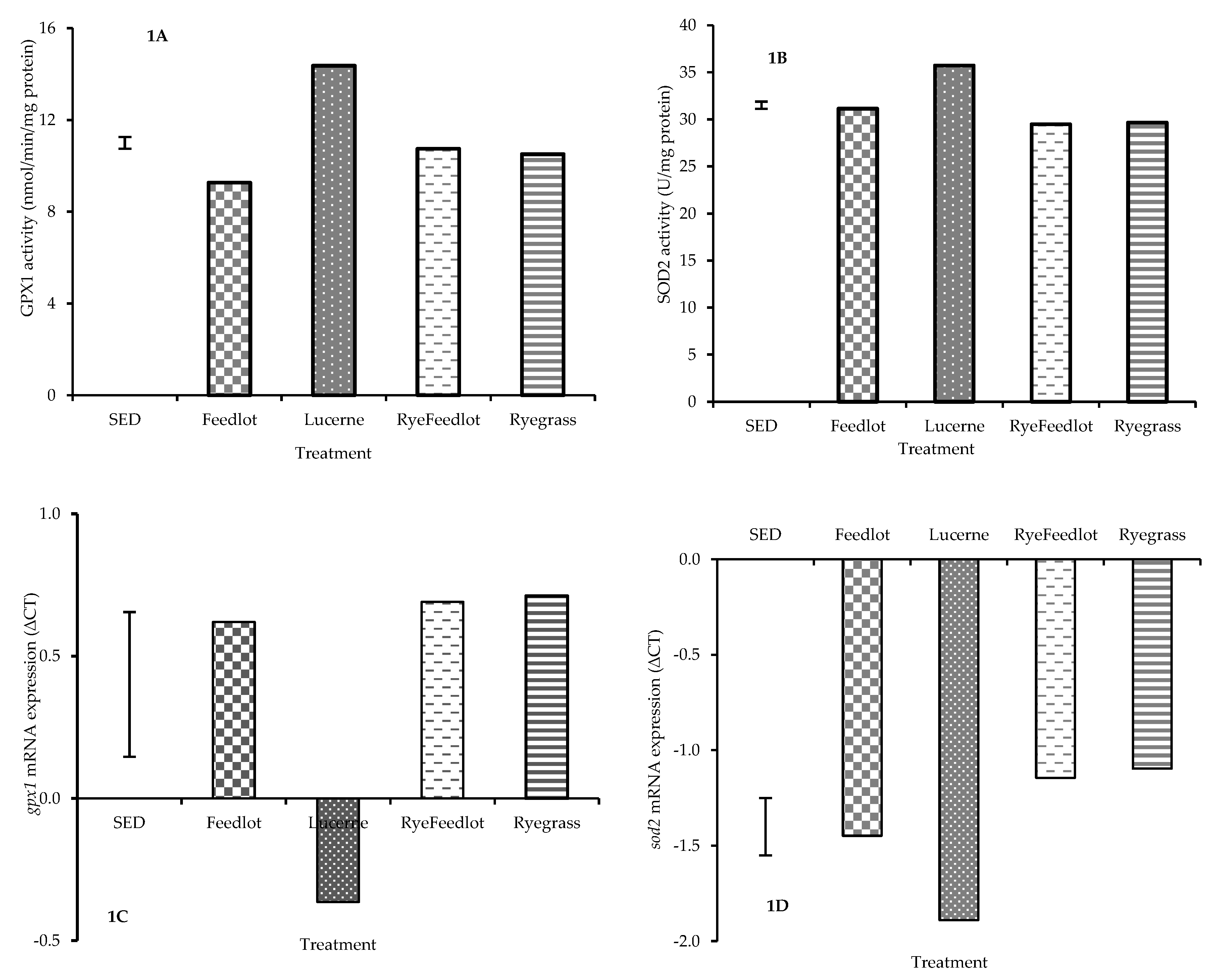
3.2. Influence of Diet on Muscle Antioxidant Enzyme Activity and Gene Expression
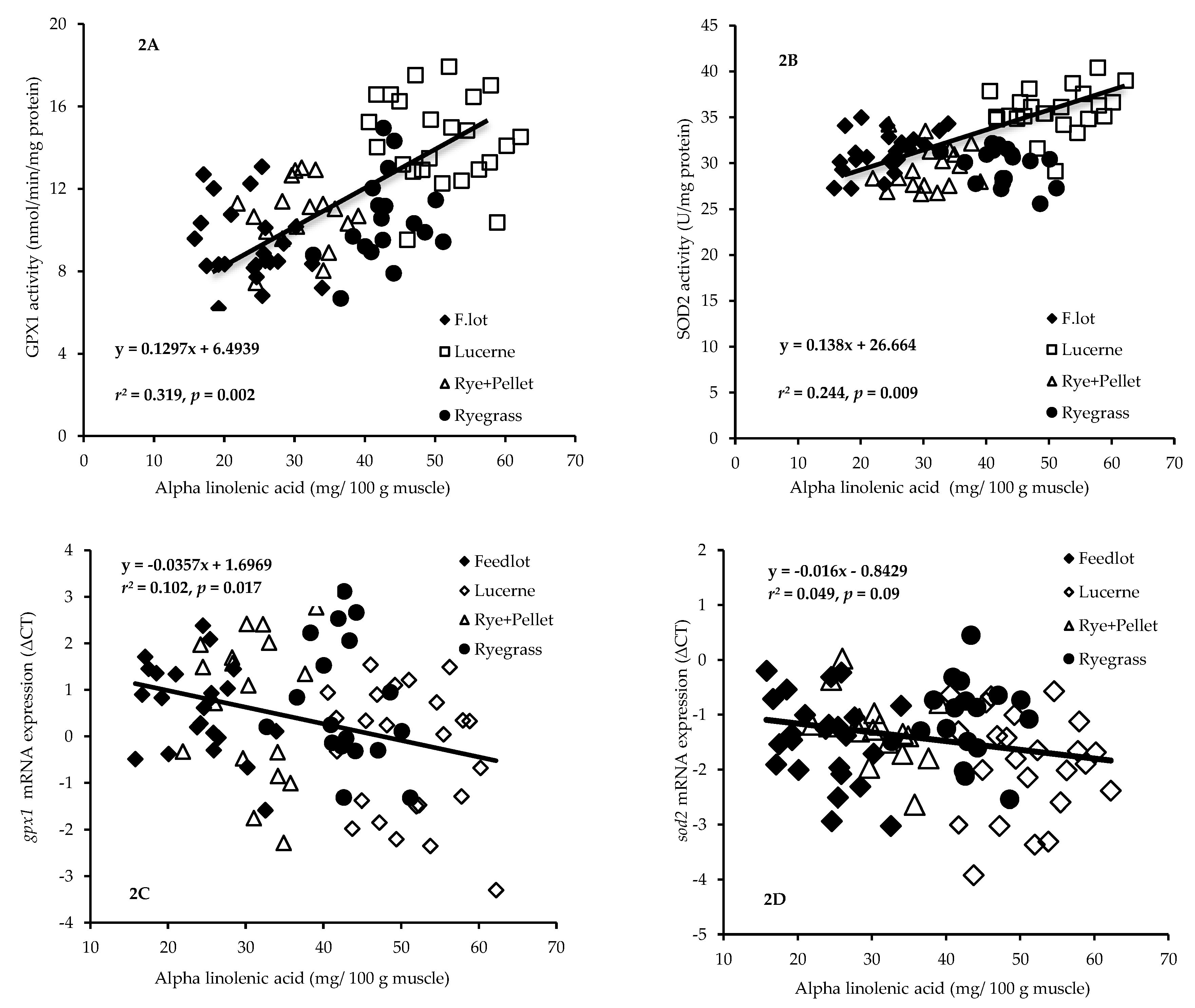
3.3. Relationship between Muscle Essential Fatty Acids (n-3 and n-6 PUFA) and Antioxidant Enzyme Activity or Gene Expression
4. Discussion
4.1. Diet and Muscle Fatty Acid Composition
4.2. Gene Expression and Activity of Enzymes Associated with Dietary Intervention
4.3. Relationships between Diet-Induced Increase in Muscle ALA and Gene Expression/Activity of Antioxidant Enzymes of GPX and SOD
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buckley, J.D.; Howe, P.C. Long-chain omega-3 polyunsaturated fatty acids may be beneficial for reducing obesity—A review. Nutrients 2010, 2, 1212–1230. [Google Scholar] [CrossRef] [PubMed]
- Daley, C.A.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutrition J. 2010, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Kopecky, J.; Rossmeisl, M.; Flachs, P.; Kuda, O.; Brauner, P.; Jilkova, Z.; Stankova, B.; Tvrzicka, E.; Bryhn, M. Symposium on ‘Frontiers in adipose tissue biology’ n-3 PUFA: Bioavailability and modulation of adipose tissue function. Proc. Nutr. Soc. 2009, 68, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N. Nutritional Modification of Muscle Long Chain Omega-3 Fatty Acids in Lamb: Effects on Growth and Composition and Quality of Meat. Ph.D. Thesis, The University of Melbourne, Victoria, Australia, 1999. [Google Scholar]
- Simopoulos, A.P. New products from the agri-food industry: The return of n-3 fatty acids into the food supply. Lipids 1999, 34, 297–301. [Google Scholar] [CrossRef]
- Nuernberg, K.; Dannenberger, D.; Nuernberg, G.; Ender, K.; Voigt, J.; Scollan, N.D.; Wood, J.D.; Nute, G.R.; Richardson, R.I. Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longissimus muscle in different cattle breeds. Livest. Prod. Sci. 2005, 94, 137–147. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Sinclair, A.J.; Egan, A.R.; Blakeley, S.J.; Li, D.; Leury, B.J. Effect of dietary modification of muscle long-chain n-3 fatty acid on plasma insulin and lipid metabolites, carcass traits, and fat deposition in lambs. J. Anim. Sci. 2001, 79, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Burnett, V.F.; Norng, S.; Hopkins, D.L.; Plozza, T.; Jacobs, J.L. Muscle antioxidant (vitamin E) and major fatty acid groups, lipid oxidation and retail colour of meat from lambs fed a roughage-based diet with flaxseed or algae. Meat Sci. 2016, 111, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Hopkins, D.L.; Jacobs, J.L. Increasing omega-3 levels in meat from ruminants under pasture-based systems: An invited review. Rev. Sci. Tech. Off. Int. Epiz. 2018, 37, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Omega-3 fatty acids in health and disease and in growth and development. Am. J. Clin. Nutr. 1991, 54, 438–463. [Google Scholar] [CrossRef]
- Benatti, P.; Peluso, G.; Nicolai, R.; Calvani, M. Polyunsaturated fatty acids: Biochemical, nutritional and epigenetic properties. J. Am. Coll. Nutr. 2004, 23, 281–302. [Google Scholar] [CrossRef]
- Sampath, H.; Ntambi, J.M. Polyunsaturated fatty acid regulation of genes of lipid metabolism. Annu. Rev. Nutr. 2005, 25, 317–340. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Chikwanha, O.C.; Vahmani, P.; Muchenje, V.; Dugan, M.E.R.; Mapiye, C. Nutritional enhancement of sheep meat fatty acid profile for human health and wellbeing. Food Res. Int. 2018, 104, 25–38. [Google Scholar] [CrossRef]
- Decker, E.A.; Akoh, C.C.; Wilkes, R.S. Incorporation of (n-3) fatty acids in foods: Challenges and opportunities. J. Nutr. 2012, 142, 610S–613S. [Google Scholar] [CrossRef]
- Nieto, G.; Ros, G. Modification of fatty acid composition in meat through diet: Effect on lipid peroxidation and relationship to nutritional quality—A review. Lipid Peroxidation 2012, 12, 239–258. [Google Scholar] [CrossRef]
- Scollan, N.D.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Norng, S.; Burnett, V.F.; Dunshea, F.R.; Jacobs, J.L.; Hopkins, D.L. The synergism of biochemical components controlling lipid oxidation in lamb muscle. Lipids 2014, 49, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Vossen, E.; Ntawubizi, M.; Raes, K.; Smet, K.; Huyghebaert, G.; Arnouts, S.; De Smet, S. Effect of dietary antioxidant supplementation on the oxidative status of plasma in broilers. J. Anim. Physiol. Anim. Nutr. 2011, 95, 198–205. [Google Scholar] [CrossRef]
- Bekhit, A.E.A.; Hopkins, D.L.; Fahri, F.T.; Ponnampalam, E.N. Oxidative processes in muscle systems and fresh meat: Sources, markers, and remedies. Comp. Rev. Food Sci. Food Saf. 2013, 12, 565–597. [Google Scholar] [CrossRef]
- Halliwell, B. Review: Lipid peroxidation, antioxidants and cardiovascular disease: How should we move forward? Cardiovas. Res. 2000, 47, 410–418. [Google Scholar] [CrossRef]
- Young, I.S. Oxidative stress and vascular disease: Insights from isoprostane measurement. Clin. Chem. 2005, 51, 14–15. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Xindan Li, X.; Kong, M.; Li, L.; Zhang, G.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Ponnampalam, E.N.; Leury, B.J.; Liu, F.; Dunshea, F.R. Antioxidant dynamics in the live animal and implications for ruminant health and product (meat/milk) quality. Anim. Prod. Sci. 2014, 54, 1425–1536. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Review: Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Burnett, V.F.; Norng, S.; Warner, R.D.; Jacobs, J.L. Vitamin E and fatty acid content of lamb meat from perennial pasture or annual pasture systems with supplements. Anim. Prod. Sci. 2012, 52, 255–262. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes, 8th ed.; Australian Government: Canberra, Australia, 2013.
- Ponnampalam, E.N.; Linden, N.P.; Mitchell, M.L.; Hopkins, D.L.; Jacobs, J.L. Production systems to deliver premium grade lambs to the growing international and Australian markets. Small. Rumin. Res. 2017, 157, 32–39. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Warner, R.D.; Kitessa, S.; McDonagh, M.B.; Pethick, D.W.; Allen, D.; Hopkins, D.L. Influence of finishing systems and sampling site on fatty acid composition and retail shelf-life of lamb. Anim. Prod. Sci. 2010, 50, 775–781. [Google Scholar] [CrossRef]
- Côrtes, C.; Palin, M.F.; Gagnon, N.; Benchaar, C.; Lacasse, P.; Petit, H.V. Mammary gene expression and activity of antioxidant enzymes and concentration of the mammalian lignan enterolactone in milk and plasma of dairy cows fed flax lignans and infused with flax oil in the abomasum. Br. J. Nutr. 2012, 108, 1390–1398. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, L.K.A.T. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar]
- Payne, R.W. The guide to GenStat, Release 15, Part 2: Statistics, 2012; VSN International: Hertfordshire, UK, 2012. [Google Scholar]
- Ponnampalam, E.N.; Lewandowski, P.; Fahri, F.T.; Burnett, V.F.; Dunshea, F.R.; Plozza, T.E.; Jacobs, J.L. Forms of n-3 (ALA, C18:3n-3 or DHA, C22:6n-3) fatty acids affect carcass yield, blood lipids, muscle n-3 fatty acids and liver gene expression in lambs. Lipids 2015, 50, 1133–1143. [Google Scholar] [CrossRef]
- Cherfaoui, M.; Durand, D.; Bonnet, M.; Bernard, L.; Bauchart, D.; Ortigues-Marty, I.; Gruffat, D. A grass-based diet favours muscle n-3 long-chain PUFA deposition without modifying gene expression of proteins involved in their synthesis or uptake in Charolais steers. Animal 2013, 7, 1833–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, S. Polyunsaturated fatty acid regulation of gene transcription: A mechanism to improve energy balance and insulin resistance. Br. J. Nutr. 2000, 83, S59–S66. [Google Scholar] [CrossRef] [PubMed]
- Haug, A.; Nyquist, N.F.; Thomassen, M.; Høstmark, A.T.; Østbye, T.K. N-3 fatty acid intake altered fat content and fatty acid distribution in chicken breast muscle but did not influence mRNA expression of lipid-related enzymes. Lipids Health Dis. 2014, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Herdmann, A.; Nuernberg, K.; Martin, J.; Nuernberg, G.; Doran, O. Effect of dietary fatty acids on expression of lipogenic enzymes and fatty acid profile in tissues of bulls. Animal 2010, 4, 755–762. [Google Scholar] [CrossRef]
- Jena, B.P.; Panda, N.; Patra, R.C.; Mishra, P.K.; Behura, N.C.; Panigrahi, B. Supplementation of vitamin E and C reduces oxidative stress in broiler breeder hens during summer. Food Nutr. Sci. 2013, 4, 33–37. [Google Scholar] [CrossRef]
- Schogor, A.L.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Lacasse, P.; Petit, H.V. Mammary gene expression and activity of antioxidant enzymes and oxidative indicators in the blood, milk, mammary tissue and ruminal fluid of dairy cows fed flax meal. Br. J. Nutr. 2013, 110, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Reddy Avula, C.P.; Fernandes, G. Modulation of antioxidant enzymes and apoptosis in mice by dietary lipids and treadmill exercise. J. Clin. Immunol. 1999, 19, 35–44. [Google Scholar] [CrossRef]
- Ruiz-Gutiérrez, V.; Pérez-Espinosa, A.; Vázquez, C.M.; Santa-María, C. Effects of dietary fats (fish, olive and high-oleic-acid sunflower oils) on lipid composition and antioxidant enzymes in rat liver. Br. J. Nutr. 1999, 82, 233–241. [Google Scholar]
- Burnett, V.F.; Seymour, G.R.; Norng, S.; Jacobs, J.L.; Ponnampalam, E.N. Lamb growth performance and carcass weight from rotationally grazed perennial pasture systems compared with annual pasture systems with supplements. Anim. Prod. Sci. 2012, 52, 248–254. [Google Scholar] [CrossRef]
- Burnett, V.F.; Jacobs, J.L.; Norng, S.; Ponnampalam, E.N. Feed intake, liveweight gain and carcass traits of lambs offered pelleted annual pasture hay supplemented with flaxseed (Linum usitatissimum) flakes or algae (Schizochytrium sp.). Anim. Prod. Sci. 2017, 57, 877–883. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Hopkins, D.L.; Khageswor, G.; Jacobs, J.L.; Plozza, T.E.; Lewandowski, P.; Bekhit, A. The use of oxidative stress biomarkers in live animals (in vivo) to predict meat quality deterioration post mortem (in vitro) caused by changes in muscle biochemical components. J. Anim. Sci. 2017, 95, 3012–3024. [Google Scholar] [PubMed]
- Karami, M.; Ponnampalam, E.N.; Hopkins, D.L. The effect of palm oil or canola oil on feedlot performance, plasma and tissue fatty acid profile and meat quality in goats. Meat Sci. 2013, 94, 165–169. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Items | Feedlot | Lucerne | Ryegrass |
|---|---|---|---|
| Dry matter, % as feed | 91.1 | 32.3 | 32.4 |
| Crude protein, % of DM | 16.1 | 18.6 | 5.2 |
| Metabolisable energy, MJ/kg DM | 12.9 | 8.6 | 10.4 |
| Acid detergent fibre, % of DM | 9.0 | 33.5 | 26.3 |
| Neutral detergent fibre, % of DM | 19.6 | 44.2 | 45.5 |
| Organic matter, % of DM | 94.7 | 91.4 | 92.8 |
| Dry matter digestibility, % of DM | 82.7 | 59.6 | 69.6 |
| Gene 1 | Accession Number (Ovine) | Primer Sequence (5′ to 3′) |
|---|---|---|
| β-actin | NM_001009784.1 | Forward: GTCCGTGACATCAAGGAGAAG Reverse: AGGAAGGAAGGCTGGAAGAG |
| cpt1β | NM_001009259.1 | Forward: CCTGGAAGAAACGCCTGATT Reverse: CAGGGTTTGGCGAAAGAAGA |
| gpx1 | XM_004018462 | Forward: AGTTTGGGCATCAGGAAAAC Reverse: CCGAAGGAAGGCGAAGAG |
| gpx4 | XM_004023249.1 | Forward: GAGTTCGCTGCTGGCTATAA Reverse: CGGCAGATCCTTCTCTATCA |
| nfκB | EF_121765.1 | Forward: ATTCAGCCCTTTGCCCATCT Reverse: ATGGGATGTCAGTGGCGTTA |
| sod1 | NM_001145185.1 | Forward: GGTTCCACGTCCATCAGTTT Reverse: CAATGGCAACACCATTTTTG |
| sod2 | NM_001280703 | Forward: TCACAGCATCTTCTGGACAA Reverse: TGCTCCTTATTGAAGCCAAG |
| upc2 | NM_001280682.1 | Forward: AAGGCCCACCTAATGACAGA Reverse: CCCAGGGCAGAGTTCATGT |
| Feedlot | Lucerne | Ryegrass | RyeFeedlot | SED | p-Value | |
|---|---|---|---|---|---|---|
| C10:0 | 4.8 | 5.8 | 5.0 | 4.2 | 0.68 | 0.13 |
| C12:0 | 5.9 | 7.9 | 7.0 | 7.0 | 0.65 | 0.04 |
| C14:0 | 86 | 102 | 93 | 90 | 9.1 | 0.27 |
| C14:1 | 3.5 | 3.9 | 3.3 | 3.7 | 0.39 | 0.43 |
| C15:0 | 8.8 | 11.3 | 10.6 | 9.7 | 0.64 | 0.01 |
| C15:1 | 0.25 | 0.26 | 0.26 | 0.24 | 0.02 | 0.24 |
| C16:0 | 566 | 607 | 577 | 499 | 42.5 | 0.09 |
| C16:1 | 42.4 | 41.1 | 38.1 | 35.8 | 3.1 | 0.16 |
| C17:0 | 27.7 | 29.7 | 29.4 | 25.3 | 2.1 | 0.07 |
| C18:0 | 317 | 376 | 375 | 320 | 27.4 | 0.06 |
| C18:1n-9cis | 860 | 880 | 845 | 748 | 60.1 | 0.15 |
| C18:2n-6 (LA) | 139 b | 99 a | 93 a | 100 a | 5.4 | 0.001 |
| Cis-9, trans-11 CLA 1 | 1.10 | 1.44 | 1.16 | 1.21 | 0.29 | 0.57 |
| C18:3n-3 (ALA) | 23.7 a | 50.7 d | 42.8 c | 30.8 b | 1.8 | 0.001 |
| C20:4n-6 (AA) | 34.6 b | 27.3 a | 28.0 a | 33.1 b | 1.3 | 0.001 |
| C20:5n-3 (EPA) | 12.8 a | 18.2 b | 17.3 b | 17.6 b | 0.74 | 0.001 |
| C22:5n-3 (DPA) | 12.9 a | 16.6 b | 15.9 b | 16.1 b | 0.58 | 0.001 |
| C22:6n-3 (DHA) | 3.7 a | 5.0 b | 4.8 b | 4.9 b | 0.32 | 0.001 |
| Total n-3 fatty acids | 53.4 a | 90.8 d | 81.0 c | 69.7 b | 2.43 | 0.001 |
| Total n-6 fatty acids | 180 c | 132 ab | 126 a | 139 b | 5.97 | 0.001 |
| Ratio of n-6: n-3 | 3.5 c | 1.5 a | 1.6 a | 2.0 b | 0.15 | 0.001 |
| Total fat (%) | 2.2 | 2.3 | 2.2 | 2.0 | 0.14 | 0.13 |
| Response Variables | Terms | |||||
|---|---|---|---|---|---|---|
| C18:3n-3 (ALA) | C20:5n-3 (EPA) | |||||
| F-Statistic | DF | p-Value | F-Statistic | DF | p-Value | |
| GPX1 activity | 12.01 | 123.5 | 0.002 | 5.47 | 181.1 | 0.022 |
| SOD2 activity | 7.35 | 151.8 | 0.009 | 0.01 | 180.6 | 0.931 |
| gpx1 gene expression | 6.75 | 121.8 | 0.017 | 0.11 | 164.4 | 0.744 |
| sod2 gene expression | 3.05 | 123.2 | 0.094 | 0.20 | 163.4 | 0.653 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponnampalam, E.N.; Vahedi, V.; Giri, K.; Lewandowski, P.; Jacobs, J.L.; Dunshea, F.R. Muscle Antioxidant Enzymes Activity and Gene Expression Are Altered by Diet-Induced Increase in Muscle Essential Fatty Acid (α-linolenic acid) Concentration in Sheep Used as a Model. Nutrients 2019, 11, 723. https://doi.org/10.3390/nu11040723
Ponnampalam EN, Vahedi V, Giri K, Lewandowski P, Jacobs JL, Dunshea FR. Muscle Antioxidant Enzymes Activity and Gene Expression Are Altered by Diet-Induced Increase in Muscle Essential Fatty Acid (α-linolenic acid) Concentration in Sheep Used as a Model. Nutrients. 2019; 11(4):723. https://doi.org/10.3390/nu11040723
Chicago/Turabian StylePonnampalam, Eric N., Vahid Vahedi, Khageswor Giri, Paul Lewandowski, Joe L. Jacobs, and Frank R. Dunshea. 2019. "Muscle Antioxidant Enzymes Activity and Gene Expression Are Altered by Diet-Induced Increase in Muscle Essential Fatty Acid (α-linolenic acid) Concentration in Sheep Used as a Model" Nutrients 11, no. 4: 723. https://doi.org/10.3390/nu11040723