The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence


 , ,
, , {kind=link}
Abstract
:1. Introduction
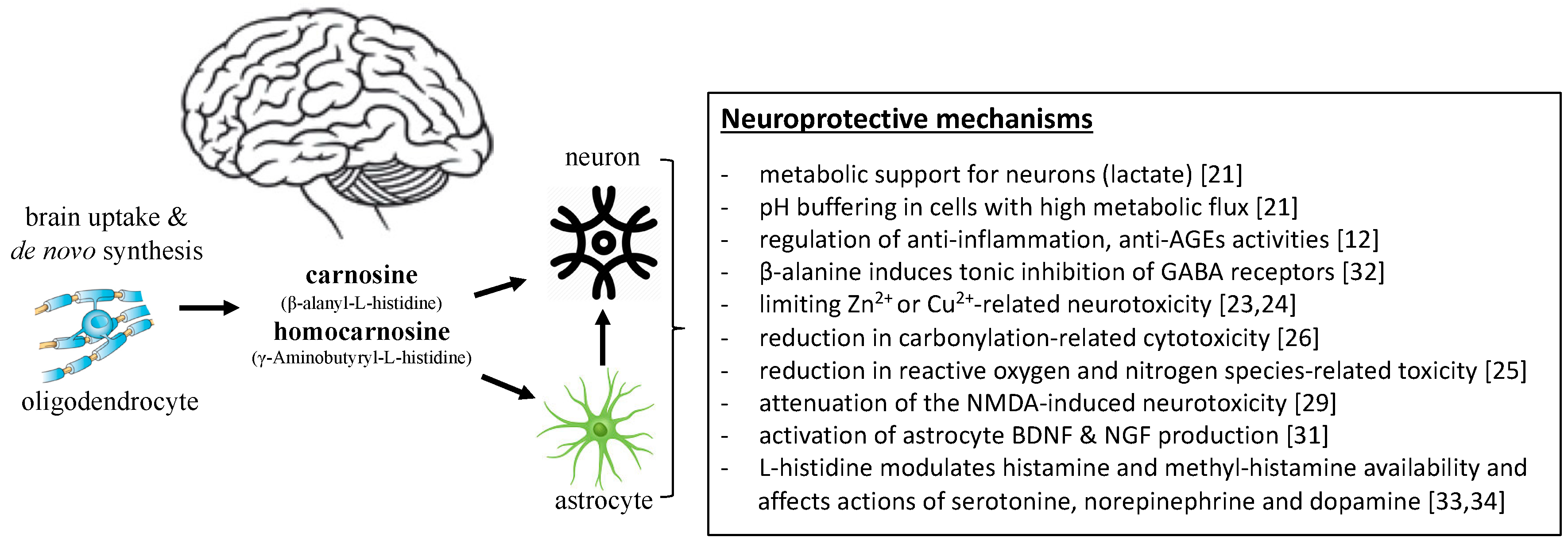
2. Proposed Mechanisms Mediating the Role of Carnosine in the Brain
3. Cell Studies of the Role of Carnosine in Brain-Related Disorders
3.1. Neurological Disorders
Brain Ischemia
3.2. Neurodegenerative Disorders
3.2.1. Alzheimer’s Disease
3.2.2. Parkinson’s Disease
3.3. Psychiatric Disorders
Major Depressive Disorder
3.4. Summary of Evidence from Cell Studies
4. Animal Studies of the Role of Carnosine in Brain-Related Disorders
4.1. Neurological and Neurodevelopmental Disorders
4.1.1. Brain Ischemia
4.1.2. Epilepsy
4.1.3. Autism Spectrum Disorder
4.1.4. Multiple Sclerosis
4.2. Neurodegenerative Disorders
4.2.1. Alzheimer’s Disease
4.2.2. Vascular Dementia
4.2.3. Parkinson’s Disease
4.3. Psychiatric Disorders
Mood Disorders
4.4. Summary of Evidence from Animal Studies
5. Human Observational Studies of the Role of Carnosine in Brain-Related Disorders
5.1. Neurological and Neurodevelopmental Disorders
5.1.1. Autism Spectrum Disorder
5.1.2. Multiple Sclerosis
5.1.3. Epilepsy and Hemorrhage
5.2. Neurodegenerative Disorders
5.2.1. Alzheimer’s Disease
5.2.2. Parkinson’s Disease
5.3. Psychiatric Disorders
5.3.1. Major Depressive Disorder
5.3.2. Schizophrenia
5.4. Summary of Evidence from Human Observational Studies
6. Human Randomized Controlled Trials of the Role of Carnosine in Brain-Related Disorders
6.1. Neurological and Neurodevelopmental Disorders
Autism Spectrum Disorder
6.2. Neurodegenerative Disorders
6.2.1. Cognitive Functions and Alzheimer’s Disease
6.2.2. Parkinson’s Disease
6.3. Psychiatric Disorders
6.3.1. Schizophrenia
6.3.2. Attention-Deficit/Hyperactivity Disorder
6.3.3. Obsessive-Compulsive Disorder
6.3.4. Behaviour
6.3.5. Fatigue
6.4. Summary of Evidence from Human Randomized Control Trials
6.5. Limitations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Noncommunicable Diseases Country Profiles 2011; World Health Organization: Geneva, Switzerland, 2011.
- Kwan, P.; Brodie, M.J. Early Identification of Refractory Epilepsy. N. Engl. J. Med. 2002. [Google Scholar] [CrossRef] [PubMed]
- Banaschewski, T.; Roessner, V.; Banaschewski, D.T.; Dittmann, R.W.; Santosh, P.J.; Rothenberger, A. Non–stimulant medications in the treatment of ADHD. Eur. Child. Adolesc. Psychiatry 2004, 13, i102–i116. [Google Scholar] [CrossRef] [PubMed]
- Simpson, H.B.; Huppert, J.D.; Petkova, E.; Foa, E.B.; Liebowitz, M.R. Response Versus Remission in Obsessive-Compulsive Disorder. J. Clin. Psychiatry 2006, 67, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, B.W.; Holland, P.; Bao, W.; Ninan, P.T.; Keller, M.B. Recovery and subsequent recurrence in patients with recurrent major depressive disorder. J. Psychiatry Res. 2012, 46, 708–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinnissen, M.; Dietrich, A.; Hoofdakker, B.J.V.D.; Hoekstra, P.J. Clinical and pharmacokinetic evaluation of risperidone for the management of autism spectrum disorder. Expert Opin. Drug Metab. Toxicol. 2014, 11, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Saltin, B. Exercise as medicine - evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25, 1–72. [Google Scholar] [CrossRef] [Green Version]
- Sale, C.; Artioli, G.G.; Gualano, B.; Saunders, B.; Hobson, R.M.; Harris, R.C. Carnosine: From exercise performance to health. Amino Acids 2013, 44, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, A.; Kurella, E.; Stvolinsky, S. Biological role of carnosine metabolism in excitable tissues: Speculations and facts (a commentary). Pathophysiology 1994, 1, 215–219. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and Pathophysiology of Carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Boldyrev, A.; Stvolinsky, S.; Fedorova, T.; Suslina, Z. Carnosine As a Natural Antioxidant and Geroprotector: From Molecular Mechanisms to Clinical Trials. Rejuvenation Res. 2010, 13, 156–158. [Google Scholar] [CrossRef]
- Berezhnoy, D.S.; Stvolinsky, S.L.; Lopachev, A.V.; Devyatov, A.A.; Lopacheva, O.M.; Kulikova, O.I.; Abaimov, D.A.; Fedorova, T.N. Carnosine as an effective neuroprotector in brain pathology and potential neuromodulator in normal conditions. Amino Acids 2019, 51, 139–150. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, R.A.; O’Kane, R.L.; Simpson, I.A.; Viña, J.R. Structure of the Blood–Brain Barrier and Its Role in the Transport of Amino Acids. J. Nutr. 2018, 136, 218S–226S. [Google Scholar] [CrossRef] [PubMed]
- De Marchis, S.; Modena, C.; Peretto, P.; Migheli, A.; Margolis, F.L.; Fasolo, A. Carnosine-related dipeptides in neurons and glia. Biochemistry 2000, 65, 824–833. [Google Scholar] [PubMed]
- Bauer, K. Carnosine and Homocarnosine, the Forgotten, Enigmatic Peptides of the Brain. Neurochem. Res. 2005, 30, 1339–1345. [Google Scholar] [CrossRef]
- De Marchis, S.; Melcangi, R.C.; Modena, C.; Cavaretta, I.; Peretto, P.; Agresti, C.; Fasolo, A. Identification of the glial cell types containing carnosine-related peptides in the rat brain. Neurosci. Lett. 1997, 237, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.-L.; Yang, L.-X.; Wu, X.-H.; Li, Q.; Ding, M.-P.; Fan, Y.-Y.; Zhang, W.-P.; Luo, J.-H.; Chen, Z. Effects of carnosine on amygdaloid-kindled seizures in Sprague–Dawley rats. Neuroscience 2005, 135, 939–947. [Google Scholar] [CrossRef]
- Kawahara, M.; Tanaka, K.-I.; Kato-Negishi, M. Zinc, Carnosine, and Neurodegenerative Diseases. Nutrients 2018, 10, 147. [Google Scholar] [CrossRef]
- Hipkiss, A.R. Possible Benefit of Dietary Carnosine towards Depressive Disorders. Aging Dis. 2015, 6, 300–303. [Google Scholar] [CrossRef]
- Caruso, G.; Caraci, F.; Jolivet, R.B. Pivotal role of carnosine in the modulation of brain cells activity: Multimodal mechanism of action and therapeutic potential in neurodegenerative disorders. Prog. Neurobiol. 2019, 175, 35–53. [Google Scholar] [CrossRef]
- Nicoletti, V.G.; Santoro, A.M.; Grasso, G.; Vagliasindi, L.I.; Giuffrida, M.L.; Cuppari, C.; Purrello, V.S.; Stella, A.M.G.; Rizzarelli, E. Carnosine interaction with nitric oxide and astroglial cell protection. J. Neurosci. 2007, 85, 2239–2245. [Google Scholar] [CrossRef] [PubMed]
- Horning, M.S.; Blakemore, L.J.; Trombley, P.Q. Endogenous mechanisms of neuroprotection: Role of zinc, copper, and carnosine. Brain Res. 2000, 852, 56–61. [Google Scholar] [CrossRef]
- Trombley, P.Q.; Horning, M.S.; Blakemore, L.J. Carnosine modulates zinc and copper effects on amino acid receptors and synaptic transmission. NeuroReport 1998, 9, 3503–3507. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Martinez-Becerra, F.; Antonio, L.; Johnson, R.T.; De Campos, R.P.S.; Siegel, J.M.; Wijesinghe, M.B.; Lazzarino, G.; Lunte, S.M.; et al. Carnosine modulates nitric oxide in stimulated murine RAW 264.7 macrophages. Mol. Cell. Biochem. 2017, 431, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Carini, M.; Aldini, G.; Beretta, G.; Arlandini, E.; Facino, R.M. Acrolein-sequestering ability of endogenous dipeptides: Characterization of carnosine and homocarnosine/acrolein adducts by electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2003, 38, 996–1006. [Google Scholar] [CrossRef]
- Lopachev, A.V.; Lopacheva, O.M.; Abaimov, D.A.; Koroleva, O.V.; Vladychenskaya, E.A.; Erukhimovich, A.A.; Fedorova, T.N. Neuroprotective Effect of Carnosine on Primary Culture of Rat Cerebellar Cells under Oxidative Stress. Biochemistry 2016, 81, 511–520. [Google Scholar] [CrossRef] [PubMed]
- A Boldyrev, A.; Johnson, P.; Wei, Y.; Tan, Y.; O Carpenter, D. Carnosine and taurine protect rat cerebellar granular cells from free radical damage. Neurosci. Lett. 1999, 263, 169–172. [Google Scholar] [CrossRef]
- Shen, Y.; Hu, W.-W.; Fan, Y.-Y.; Dai, H.-B.; Fu, Q.-L.; Wei, E.-Q.; Luo, J.-H.; Chen, Z. Carnosine protects against NMDA-induced neurotoxicity in differentiated rat PC12 cells through carnosine-histidine-histamine pathway and H1/H3 receptors. Biochem. Pharmacol. 2007, 73, 709–717. [Google Scholar] [CrossRef]
- Fleisher-Berkovich, S.; Abramovitch-Dahan, C.; Ben-Shabat, S.; Apte, R.; Beit-Yannai, E. Inhibitory effect of carnosine and N-acetyl carnosine on LPS-induced microglial oxidative stress and inflammation. Peptides 2009, 30, 1306–1312. [Google Scholar] [CrossRef]
- Yamashita, S.; Sato, M.; Matsumoto, T.; Kadooka, K.; Hasegawa, T.; Fujimura, T.; Katakura, Y. Mechanisms of carnosine-induced activation of neuronal cells. Biosci. Biotechnol. Biochem. 2017, 82, 683–688. [Google Scholar] [CrossRef]
- Chesnoy-Marchais, D. Persistent GABAA/C responses to gabazine, taurine and beta-alanine in rat hypoglossal motoneurons. Neuroscience 2016, 330, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Nakamura, T.; Shibakusa, T.; Sugita, M.; Naganuma, F.; Iida, T.; Miura, Y.; Mohsen, A.; Harada, R.; Yanai, K. Insufficient Intake of L-Histidine Reduces Brain Histamine and Causes Anxiety-Like Behaviors in Male Mice. J. Nutr. 2014, 144, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Flancbaum, L.; Fitzpatrick, J.C.; Brotman, D.N.; Marcoux, A.-M.; Kasziba, E.; Fisher, H. The presence and significance of carnosine in histamine-containing tissues of several mammalian species. Agents Actions 1990, 31, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Majid, A. Neuroprotection in Stroke: Past, Present, and Future. ISRN Neurol. 2014, 2014, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou-Yang, L.; Liu, Y.; Wang, B.Y.; Cao, P.; Zhang, J.J.; Huang, Y.Y.; Lyu, J.X. Carnosine suppresses oxygen-glucose deprivation/recovery-induced proliferation and migration of reactive astrocytes of rats in vitro. Acta. Pharmacol. Sin. 2018, 39, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Bae, O.N.; Serfozo, K.; Baek, S.H.; Lee, K.Y.; Dorrance, A.; Rumbeiha, W.; Fitzgerald, S.D.; Farooq, M.U.; Naravelta, B.; Bhatt, A.; et al. Safety and efficacy evaluation of carnosine, an endogenous neuroprotective agent for ischemic stroke. Stroke 2013, 44, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Tian, Y.; Yang, J.; Shi, X.; Ouyang, L.; Gao, J.; Lu, J. Dual effects of carnosine on energy metabolism of cultured cortical astrocytes under normal and ischemic conditions. Regul. Pept. 2014, 192, 45–52. [Google Scholar] [CrossRef]
- Baek, S.-H.; Noh, A.R.; Kim, K.-A.; Akram, M.; Shin, Y.-J.; Kim, E.-S.; Yu, S.W.; Majid, A.; Bae, O.-N. Modulation of mitochondrial function and autophagy mediates carnosine neuroprotection against ischemic brain damage. Stroke 2014, 45, 2438–2443. [Google Scholar] [CrossRef]
- Shi, X.; Wang, B.; Liu, Y.; Zhang, J.; Huang, Y.; Cao, P.; Shen, Y.; Lyu, J. Carnosine modulates glutamine synthetase expression in senescent astrocytes exposed to oxygen-glucose deprivation/recovery. Brain Res. Bull. 2017, 130, 138–145. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, K.; Fan, Y.; He, P.; Wang, X.; Hu, W.; Chen, Z. Carnosine protects brain microvascular endothelial cells against rotenone-induced oxidative stress injury through histamine H1and H2receptorsin vitro. Clin. Exp. Pharmacol. Physiol. 2012, 39, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Khama-Murad, A.; Mokrushin, A.; Pavlinova, L. Neuroprotective properties of l-carnosine in the brain slices exposed to autoblood in the hemorrhagic stroke model in vitro. Regul. Pept. 2011, 167, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Bae, O.-N.; Majid, A. Role of histidine/histamine in carnosine-induced neuroprotection during ischemic brain damage. Brain Res. 2013, 1527, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Puerto, A.M.; Russo, C.; Fregni, F. Alzheimer’s Disease. Mol. Imaging Clin. Neurosci. 2018, 138, 297–338. [Google Scholar]
- Aloisi, A.; Barca, A.; Romano, A.; Guerrieri, S.; Storelli, C.; Rinaldi, R.; Verri, T. Anti-Aggregating Effect of the Naturally Occurring Dipeptide Carnosine on Aβ1-42 Fibril Formation. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Attanasio, F.; Convertino, M.; Magno, A.; Caflisch, A.; Corazza, A.; Haridas, H.; Esposito, G.; Cataldo, S.; Pignataro, B.; Milardi, D.; et al. Carnosine Inhibits Aβ42Aggregation by Perturbing the H-Bond Network in and around the Central Hydrophobic Cluster. ChemBioChem 2013, 14, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Corona, C.; Frazzini, V.; Silvestri, E.; Lattanzio, R.; La Sorda, R.; Piantelli, M.; Canzoniero, L.M.T.; Ciavardelli, D.; Rizzarelli, E.; Sensi, S.L. Effects of Dietary Supplementation of Carnosine on Mitochondrial Dysfunction, Amyloid Pathology, and Cognitive Deficits in 3xTg-AD Mice. PLoS ONE 2011, 6, e17971. [Google Scholar] [CrossRef] [PubMed]
- Preston, J.E.; Hipkiss, A.R.; Himsworth, D.T.; Romero, I.A.; Abbott, J.N. Toxic effects of b -amyloid (25–35) on immortalised rat brain endothelial cell: Protection by carnosine, homocarnosine and b -alanine. Neurosc. Lett. 1998, 242, 105–108. [Google Scholar] [CrossRef]
- Wu, J.W.; Liu, K.-N.; How, S.-C.; Chen, W.-A.; Lai, C.-M.; Liu, H.-S.; Hu, C.-J.; Wang, S.S.-S. Carnosine’s Effect on Amyloid Fibril Formation and Induced Cytotoxicity of Lysozyme. PLoS ONE 2013, 8, e81982. [Google Scholar] [CrossRef]
- Brownrigg, T.D.; Theisen, C.S.; Fibuch, E.E.; Seidler, N.W. Carnosine protects against the neurotoxic effects of a serotonin-derived melanoid. Neurochem. Res. 2011, 36, 465–475. [Google Scholar] [CrossRef]
- Martinez-Cengotitabengoa, M.; MacDowell, K.S.; Alberich, S.; Diaz, F.; Garcia-Bueno, B.; Rodriguez-Jimenez, R.; Bioque, M.; Berrocoso, E.; Parellada, M.; Lobo, A.; et al. BDNF and NGF Signalling in Early Phases of Psychosis: Relationship with Inflammation and Response to Antipsychotics after 1 Year. Schizophr. Bull. 2016, 42, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Cole, T.B.; Palmiter, R.D.; Suh, S.W.; Koh, J.-Y. Contribution by synaptic zinc to the gender-disparate plaque formation in human Swedish mutant APP transgenic mice. Proc. Natl. Acad. Sci. USA 2002, 99, 7705–7710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Dong, X.; Sun, Y. Carnosine-LVFFARK-NH2 Conjugate: A Moderate Chelator but Potent Inhibitor of Cu2 + -Mediated Amyloid # -Protein Aggregation. ACS Chem. Neurosci. 2018, 9, 2689–2700. [Google Scholar] [CrossRef] [PubMed]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neuro-Visc. Relat. 2017, 124, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shi, L.; Zhang, L. Neuroprotective effect of carnosine against salsolinol - induced Parkinson’s disease. Exp. Ther. Med. 2017, 664–670. [Google Scholar] [CrossRef]
- Gotlib, I.H.; Lemoult, J.; Colich, N.L.; Foland-Ross, L.C.; Hallmayer, J.; Joormann, J.; Lin, J.; Wolkowitz, O.M. Telomere length and cortisol reactivity in children of depressed mothers. Mol. Psychiatry 2015, 20, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.K.; Ridout, S.J.; Price, L.H.; Sen, S.; Tyrka, A.R. Depression and telomere length: A meta-analysis. J. Affect. Disord. 2016, 191, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Gheshlagh, R.G.; Parizad, N.; Sayehmiri, K. The Relationship Between Depression and Metabolic Syndrome: Systematic Review and Meta-Analysis Study. Iran. Red Crescent Med. J. 2016, 18, e26523. [Google Scholar] [CrossRef]
- Shao, L.; Li, Q.-H.; Tan, Z. l-Carnosine reduces telomere damage and shortening rate in cultured normal fibroblasts. Biochem. Biophys. Commun. 2004, 324, 931–936. [Google Scholar] [CrossRef]
- McFarland, G.; Holliday, R. Retardation of the Senescence of Cultured Human Diploid Fibroblasts by Carnosine. Exp. Cell. 1994, 212, 167–175. [Google Scholar] [CrossRef]
- Tsoi, B.; He, R.-R.; Yang, D.-H.; Li, Y.-F.; Li, X.-D.; Li, W.-X.; Abe, K.; Kurihara, H. Carnosine Ameliorates Stress-Induced Glucose Metabolism Disorder in Restrained Mice. J. Pharmacol. Sci. 2011, 117, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.K.; Laud, P.J.; Bahor, Z.; Rajanikant, G.; Majid, A. Systematic review and stratified meta-analysis of the efficacy of carnosine in animal models of ischemic stroke. Br. J. Pharmacol. 2016, 36, 1686–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrota, D.; Fedorova, T.; Stvolinsky, S.; Babusikova, E.; Likavčanová, K.; Drgova, A.; Strapkova, A.; Boldyrev, A. Carnosine Protects the Brain of Rats and Mongolian Gerbils against Ischemic Injury: After-Stroke-Effect. Neurochem. Res. 2005, 30, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Hara, K.; Maki, M.; Masuda, T.; Sanberg, C.D.; Sanberg, P.R.; Bickford, P.C.; Borlongan, C.V. Dietary Supplementation Exerts Neuroprotective Effects in Ischemic Stroke Model. Rejuvenation Res. 2008, 11, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.H.; Ma, Q.; Wang, X.; Xu, G.H. Protective effects of beef decoction rich in carnosine on cerebral ischemia injury by permanent middle cerebral artery occlusion in rats. Exp. Ther. Med. 2018, 15, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Pekçetin, Ç.; Kiray, M.; Ergur, B.U.; Tuğyan, K.; Bagriyanik, H.A.; Erbil, G.; Baykara, B.; Camsari, U.M. Carnosine attenuates oxidative stress and apoptosis in transient cerebral ischemia in rats. Acta Biol. Hung. 2009, 60, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, L.; Cheng, X.; Yang, Y.; Luan, B.; Jia, L.; Xu, F.; Zhang, Z. Carnosine pretreatment protects against hypoxia–ischemia brain damage in the neonatal rat model. Eur. J. Pharmacol. 2011, 667, 202–207. [Google Scholar] [CrossRef]
- Wang, J.-P.; Yang, Z.-T.; Liu, C.; He, Y.-H.; Zhao, S.-S. L-carnosine inhibits neuronal cell apoptosis through signal transducer and activator of transcription 3 signaling pathway after acute focal cerebral ischemia. Brain Res. 2013, 1507, 125–133. [Google Scholar] [CrossRef]
- Min, J.; Rajanikant, K.; Greenberg, E.; Bandagi, R.; Zemke, D.; Mousa, A.; Kassab, M.; Farooq, M.U.; Gupta, R.; Majid, A.; et al. Differential Neuroprotective Effects of Carnosine, Anserine, and N-Acetyl Carnosine against Permanent Focal Ischemia. J. Neurosci. 2008, 86, 2984–2991. [Google Scholar] [CrossRef]
- Adachi, N.; Liu, K.; Ninomiya, K.; Matsuoka, E.; Motoki, A.; Irisawa, Y.; Nishibori, M. Reduction of the infarct size by simultaneous administration of l-histidine and diphenhydramine in ischaemic rat brains. Resuscitation 2011, 82, 219–221. [Google Scholar] [CrossRef]
- Adachi, N.; Liu, K.; Motoki, A.; Hiraga, N.; Irisawa, Y.; Semba, K.; Arai, T. A comparison of protective effects between l-histidine and hypothermia against ischemia-induced neuronal damage in gerbil hippocampus. Eur. J. Pharmacol. 2006, 546, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Adachi, N.; Liu, K.; Arai, T. Prevention of brain infarction by postischemic administration of histidine in rats. Brain Res. 2005, 1039, 220–223. [Google Scholar] [CrossRef]
- Kaur, I.; Jaggi, A.S.; Singh, N.; Kumar, A. Evidence for the role of histaminergic pathways in neuroprotective mechanism of ischemic postconditioning in mice. Fundam. Clin. Pharmacol. 2017, 31, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; He, P.; Fan, Y.; Zhang, J.X.; Yan, H.J.; Hu, W.W.; Ohtsu, H.; Chen, Z. Free Radical Biology & Medicine Carnosine protects against permanent cerebral ischemia in histidine decarboxylase knockout mice by reducing glutamate excitotoxicity. Free Radic. Biol. Med. 2010, 48, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Dichter, M.A. Emerging Insights Into Mechanisms of Epilepsy: Implications for New Antiepileptic Drug Development. Epilepsia 1994, 35, S51–S57. [Google Scholar] [CrossRef]
- Schwartz, J.C.; Arrang, J.M.; Garbarg, M.; Pollard, H.; Ruat, M. Histaminergic transmission in the mammalian brain. Physiol. Rev. 1991, 71, 1–51. [Google Scholar] [CrossRef]
- Gietzen, D.W.; Lindström, S.H.; Sharp, J.W.; The, P.S.; Donovan, M.J. Indispensable amino acid-deficient diets induce seizures in ketogenic diet-fed rodents, demonstrating a role for amino acid balance in dietary treatments for epilepsy. J. Nutr. 2018, 148, 480–489. [Google Scholar] [CrossRef]
- Qi, Z.; Yu, X.; Xu, P.; Hao, Y.; Pan, X.; Zhang, C. l-Homocarnosine, l-carnosine, and anserine attenuate brain oxidative damage in a pentylenetetrazole-induced epilepsy model of ovariectomized rats. 3 Biotech. 2018, 8, 363. [Google Scholar] [CrossRef]
- Wu, X.-H.; Ding, M.-P.; Zhu-Ge, Z.-B.; Zhu, Y.-Y.; Jin, C.-L.; Chen, Z. Carnosine, a precursor of histidine, ameliorates pentylenetetrazole-induced kindled seizures in rat. Neurosci. Lett. 2006, 400, 146–149. [Google Scholar] [CrossRef]
- Zhu, Y.-Y.; Zhu-Ge, Z.-B.; Wu, D.-C.; Wang, S.; Liu, L.-Y.; Ohtsu, H.; Chen, Z. Carnosine inhibits pentylenetetrazol-induced seizures by histaminergic mechanisms in histidine decarboxylase knock-out mice. Neurosci. Lett. 2007, 416, 211–216. [Google Scholar] [CrossRef]
- Kozan, R.; Sefil, F.; Bağırıcı, F.; Bağirici, F. Anticonvulsant effect of carnosine on penicillin-induced epileptiform activity in rats. Brain Res. 2008, 1239, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.-B.; Li, B.; Gu, J.; Cheng, J.-S.; Yang, R. Electro-acupuncture improves epileptic seizures induced by kainic acid in taurine-depletion rats. Acupunct. Electro-Ther. Res. 2005, 30, 207–217. [Google Scholar] [CrossRef]
- Kamiński, R.M.; Zółkowska, D.; Kozicka, M.; Kleinrok, Z.; Czuczwar, S.J. L-Histidine is a beneficial adjuvant for antiepileptic drugs against maximal electroshock-induced seizures in mice. Amino Acids 2004, 26, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.-Y.; Zheng, X.-J.; Wang, J. Effects of carnosine on the evoked potentials in hippocampal CA1 region. J. Zhejiang Univ. B 2009, 10, 505–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiotto, A.; Calderan, A.; Ruzza, P.; Borin, G. Carnosine and Carnosine-Related Antioxidants: A Review. Curr. Med. Chem. 2005, 12, 2293–2315. [Google Scholar] [CrossRef] [PubMed]
- Berney, T.P. Autism – an evolving concept. Br. J. Psychiatry 2000, 176, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; De Theije, C.G.; Da Silva, S.L.; Van Der Horst, H.; Reinders, M.T.; Broersen, L.M.; Willemsen, L.E.; Kas, M.J.; Garssen, J.; Kraneveld, A.D.; et al. mTOR plays an important role in cow’s milk allergy-associated behavioral and immunological deficits. Neuropharmacology 2015, 97, 220–232. [Google Scholar] [CrossRef]
- Wu, J.; De Theije, C.G.; Da Silva, S.L.; Abbring, S.; Van Der Horst, H.; Broersen, L.M.; Willemsen, L.; Kas, M.; Garssen, J.; Kraneveld, A.D. Dietary interventions that reduce mTOR activity rescue autistic-like behavioral deficits in mice. Brain, Behav. Immun. 2017, 59, 273–287. [Google Scholar] [CrossRef]
- Machado-vieira, R.; Zanetti, M.V.; Teixeira, A.L.; Uno, M.; Valiengo, L.L.; Soeiro-de-Souza, M.G.; Oba-Shinjo, S.M.; de Sousa, R.T.; Zarate, C.A., Jr.; Gattaz, W.F. Decreased Akt 1/mTOR pathway mRNA expression in short-term bipolar disorder. Eur. Neuropsychopharmacol. 2018, 25, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Price, J.B.; Bronars, C.; Erhardt, S.; Cullen, K.R.; Schwieler, L.; Berk, M.; Walder, K.; McGee, S.L.; Frye, M.A.; Tye, S.J. Bioenergetics and synaptic plasticity as potential targets for individualizing treatment for depression. Neurosci. Biobehav. Rev. 2018, 90, 212–220. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, L.; Wu, X.; Liu, G.; Peng, Y.; Xin, X.; Jiao, B.; Kong, X. Carnosine Inhibits the Proliferation of Human Gastric Carcinoma Cells by Retarding Akt/mTOR/p70S6K Signaling. J. Cancer 2014, 5, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, R.; Hartung, H.-P.; Toyka, K.V. Animal models for autoimmune demyelinating disorders of the nervous system. Mol. Med. Today 2000, 2, 88–91. [Google Scholar] [CrossRef]
- Keytsman, C.; Blancquaert, L.; Wens, I.; Missine, M.; Van Noten, P.; Vandenabeele, F.; Derave, W.; Eijnde, B.O. Muscle carnosine in experimental autoimmune encephalomyelitis L:"and multiple sclerosis. Mult. Scler. Relat. Disord. 2018, 21, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Le Page, C.; Ferry, A.; Rieu, M. Effect of muscular exercise on chronic relapsing experimental autoimmune encephalomyelitis. J. Appl. Physiol. 1994, 77, 2341–2347. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Green, B.D. Temporal effects of neuron-specific beta-secretase 1 (BACE1) knock-in on the mouse brain metabolome: Implications for Alzheimer’s disease. Neuroscience 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- Sase, A.; Dahanayaka, S.; Höger, H.; Wu, G.; Lubec, G. Changes of hippocampal beta-alanine and citrulline levels are paralleling early and late phase of retrieval in the Morris Water Maze. Behav. Brain 2013, 249, 104–108. [Google Scholar] [CrossRef]
- Attanasio, F.; Cataldo, S.; Fisichella, S.; Nicoletti, S.; Nicoletti, V.G.; Pignataro, B.; Savarino, A.; Rizzarelli, E. Protective Effects ofl- andd-Carnosine on α-Crystallin Amyloid Fibril Formation: Implications for Cataract Disease. Biochemistry 2009, 48, 6522–6531. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Enya, A.; Enomoto, K.; Ding, Q.; Hisatsune, T. Anserine (beta-alanyl-3-methyl-L-histidine) improves neurovascular-unit dysfunction and spatial memory in aged AβPPswe/PSEN1dE9 Alzheimer’s-model mice. Sci. Rep. 2017, 7, 12571. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, N.; Bhatia, P.; Kumar, A.; Jaggi, A.S.; Singh, N. Potential of carnosine, a histamine precursor in rat model of bilateral common carotid artery occlusion-induced vascular dementia. Fundam. Clin. Pharmacol. 2018, 32, 516–531. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.A.; Chu, Y.; Grider, A.; Coffield, J.A. Supplementation with L -Histidine during Dietary Zinc Repletion Improves Short-Term Memory in Zinc-Restricted Young Adult Male Rats. J. Nutr. 2000, 130, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, T.K.; Pence, B.D.; Ossyra, J.M.; Gibbons, T.E.; Pérez, S.; McCusker, R.H.; Kelley, K.W.; Johnson, R.W.; Woods, J.A.; Rhodes, J.S. Exercise but not (-)-Epigallocatechin-3-gallate or β-Alanine enhances physical fitness, brain plasticity, and behavioral performance in mice. Physiol. Behav. 2015, 145, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Fernàndez-Busquets, X.; Ponce, J.; Bravo, R.; Arimon, M.; Martiáñez, T.; Gella, A.; Cladera, J.; Durany, N. Modulation of amyloid beta peptide(1-42) cytotoxicity and aggregation in vitro by glucose and chondroitin sulfate. Curr. Res. 2010, 7. [Google Scholar]
- Loske, C.; Gerdemann, A.; Schepl, W.; Wycislo, M.; Schinzel, R.; Palm, D.; Riederer, P.; Münch, G. Transition metal-mediated glycoxidation accelerates cross-linking of β-amyloid peptide. JBIC J. Biol. Inorg. Chem. 2000, 267, 4171–4178. [Google Scholar] [CrossRef]
- Colín-Barenque, L.; Bizarro-Nevares, P.; Villalva, A.G.; Pedraza-Chaverri, J.; Medina-Campos, O.N.; Jimenez-Martínez, R.; Rodríguez-Rangel, D.S.; Reséndiz, S.; Fortoul, T.I. Neuroprotective effect of carnosine in the olfactory bulb after vanadium inhalation in a mouse model. Int. J. Exp. Pathol. 2018, 99, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C. Mechanistic Insights Into Diabetes Mellitus and Oxidative Stress. Med. Chem. 2007, 14, 1729–1738. [Google Scholar]
- Ma, J.; Chen, J.; Bo, S.; Lu, X.; Zhang, J. Protective effect of carnosine after chronic cerebral hypoperfusion possibly through suppressing astrocyte activation. Am. J. Transl. 2015, 7, 2706–2715. [Google Scholar]
- Fišar, Z.; Jirák, R.; Zvěřová, M.; Setnička, V.; Habartová, L.; Hroudová, J.; Vaníčková, Z.; Raboch, J. Plasma amyloid beta levels and platelet mitochondrial respiration in patients with Alzheimer’s disease. Clin. Biochem. 2019. [Google Scholar] [CrossRef]
- Oliveira, W.H.; Nunes, A.K.; França, M.E.; Santos, L.A.; Lós, D.B.; Rocha, S.W.; Barbosa, K.P.; Rodrigues, G.B.; Peixoto, C.A. Effects of metformin on inflammation and short-term memory in streptozotocin-induced diabetic mice. Brain Res. 2016, 1644, 149–160. [Google Scholar] [CrossRef]
- Nagayach, A.; Patro, N.; Patro, I. Astrocytic and microglial response in experimentally induced diabetic rat brain. Metab. Brain 2014, 29, 747–761. [Google Scholar] [CrossRef]
- Ahshin-Majd, S.; Zamani, S.; Kiamari, T.; Kiasalari, Z.; Baluchnejadmojarad, T.; Roghani, M. Carnosine ameliorates cognitive deficits in streptozotocin-induced diabetic rats: Possible involved mechanisms. Peptides 2016, 86, 102–111. [Google Scholar] [CrossRef]
- Herculano, B.; Tamura, M.; Ohba, A.; Shimatani, M.; Kutsuna, N.; Hisatsune, T. β-alanyl-L-histidine rescues cognitive deficits caused by feeding a high fat diet in a transgenic mouse model of Alzheimer’s disease. J. Alzheimer’s 2013, 33, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, P.B.; Counts, S.E.; Nyenhuis, D. Vascular cognitive impairment and dementia. Biochim. Biophys. Acta Mol. Basis Dis. 2016. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.-Y.; Hou, W.-W.; Yan, H.-J.; Huang, S.-W.; Hu, W.-W.; Ma, J.; Xiong, J.; Hou, W.; Yan, H.; Sun, Y.; et al. Protective Effect of Carnosine on Subcortical Ischemic Vascular Dementia in Mice. CNS Neurosci. Ther. 2012, 18, 745–753. [Google Scholar]
- Afshin-Majd, S.; Khalili, M.; Roghani, M.; Mehranmehr, N.; Baluchnejadmojarad, T. Carnosine Exerts Neuroprotective Effect Against 6-Hydroxydopamine Toxicity in Hemiparkinsonian Rat. Mol. Neurobiol. 2014, 51, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-J.; Kuo, W.-W.; Liu, W.-H.; Yin, M.-C. Antioxidative and Anti-Inflammatory Protection from Carnosine in the Striatum of MPTP-Treated Mice. J. Agric. Food Chem. 2010, 58, 11510–11516. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, O.; Berezhnoy, D.; Stvolinsky, S.; Lopachev, A.; Orlova, V.; Fedorova, T. Neuroprotective effect of the carnosine – α-lipoic acid nanomicellar complex in a model of early-stage Parkinson’s disease. Regul. Toxicol. Pharmacol. 2018, 95, 254–259. [Google Scholar] [CrossRef]
- Bermúdez, M.-L.; Skelton, M.R.; Genter, M.B. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the Thy1-aSyn mouse model of Parkinson’s disease. Mol. Genet. Metab. 2018, 125, 305–313. [Google Scholar] [CrossRef]
- Gasser, T.; Hardy, J.; Mizuno, Y. Milestones in PD genetics. Mov. Disord. 2011, 26, 1042–1048. [Google Scholar] [CrossRef]
- Ishiwari, K.; Mingote, S.; Correa, M.; Trevitt, J.T.; Carlson, B.B.; Salamone, J.D. The GABA uptake inhibitor β-alanine reduces pilocarpine-induced tremor and increases extracellular GABA in substantia nigra pars reticulata as measured by microdialysis. J. Neurosci. Methods 2004, 140, 39–46. [Google Scholar] [CrossRef]
- Ericson, M. Clarke ÆRBC. b -alanine elevates dopamine levels in the rat nucleus accumbens: Antagonism by strychnine. Amino Acids 2010, 1051–1055. [Google Scholar] [CrossRef]
- Hipkiss, A.R. On the Relationship between Energy Metabolism, Proteostasis, Aging and Parkinson’s Disease: Possible Causative Role of Methylglyoxal and Alleviative Potential of Carnosine. Aging Dis. 2017, 8, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Chondrogianni, N.; Petropoulos, I.; Grimm, S.; Georgila, K.; Catalgol, B.; Friguet, B.; Grune, T.; Gonos, E.S. Protein damage, repair and proteolysis. Mol. Asp. Med. 2014, 35, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Li, Y.; Xiao, S.; Song, D.; Qing, H.; Li, Q.; Rajput, A.H. Occurrence and distribution of salsolinol-like compound, 1-acetyl-6,7-dihydroxy-1,2,3,4-tetrahydroisoquinoline (ADTIQ) in Parkinsonian brains. J. Neural. Transm. 2012, 119, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Uchiki, T.; Weikel, K.A.; Jiao, W.; Shang, F.; Caceres, A.; Pawlak, D.; Handa, J.T.; Brownlee, M.; Nagaraj, R.; Taylor, A. Glycation-altered proteolysis as a pathobiologic mechanism that links dietary glycemic index, aging, and age-related disease (in nondiabetics). Aging Cell. 2012, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.; Cuny, J.; Nawroth, G.; Djuric, Z.; Humpert, P.M.; Zeier, M.; Bierhaus, A.; Nawroth, P.P. Is diabetes an acquired disorder of reactive glucose metabolites and their intermediates? Diabetologia 2012, 55, 1151–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelenberg, A.J.; Hopkins, H.S. Assessing and Treating Depression in Primary Care Medicine. Am. J. Med. 2007, 120, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Bansal, Y.; Kuhad, A. Mitochondrial Dysfunction in Depression. Curr. Neuropharmacol. 2016, 14, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.; Walder, K.; McGee, S.L.; Dean, O.M.; Tye, S.J.; Maes, M.; Berk, M. A model of the mitochondrial basis of bipolar disorder. Neurosci. Biobehav. Rev. 2017, 74, 1–20. [Google Scholar] [CrossRef]
- Banerjee, S.; Ghosh, T.K.; Poddar, M.K. Carnosine reverses the aging-induced down regulation of brain regional serotonergic system. Mech. Ageing Dev. 2015, 152, 5–14. [Google Scholar] [CrossRef]
- Dolu, N.; Acer, H.; Kara, A.Y. Investigation of Dose-Related Effects of Carnosine on Anxiety with Sympathetic Skin Response and T-maze. Acta Med. 2014, 57, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Tomonaga, S.; Yamane, H.; Onitsuka, E.; Yamada, S.; Sato, M.; Takahata, Y.; Morimatsu, F.; Furuse, M. Pharmacology, Biochemistry and Behavior Carnosine-induced antidepressant-like activity in rats. Pharmacol. Biochem. Behav. 2008, 89, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.V.; Krishna, D.R.; Palit, G. Histaminergic H1 receptors mediate L-histidine-induced anxiety in elevated plus-maze test in mice. Behav. Pharmacol. 2007, 18, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Canto-De-Souza, L.; Garção, D.; Romaguera, F.; Mattioli, R. Dorsal hippocampal microinjection of chlorpheniramine reverses the anxiolytic-like effects of l-histidine and impairs emotional memory in mice. Neurosci. Lett. 2015, 587, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, S.F.; Ohtsu, H.; Benice, T.S.; Rizk-Jackson, A.; Raber, J. Age-dependent measures of anxiety and cognition in male histidine decarboxylase knockout (Hdc−/−) mice. Brain Res. 2006, 1071, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Tomonaga, S.; Toyoda, A. Effects of Diet Quality and Psychosocial Stress on the Metabolic Profiles of Mice. J. Proteome 2017, 16, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Tomonaga, S.; Tachibana, T.; Takagi, T.; Saito, E.-S.; Zhang, R.; Denbow, D.; Furuse, M. Effect of central administration of carnosine and its constituents on behaviors in chicks. Brain Res. Bull. 2004, 63, 75–82. [Google Scholar] [CrossRef]
- Murakami, T.; Furuse, M. The impact of taurine- and beta-alanine-supplemented diets on behavioral and neurochemical parameters in mice: Antidepressant versus anxiolytic-like effects. Amino Acids 2010, 39, 427–434. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Gepner, Y.; Cohen, H. β-Alanine supplementation reduces anxiety and increases neurotrophin expression in both young and older rats. Nutr. Res. 2019, 62, 51–63. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Ostfeld, I.; Stout, J.R.; Harris, R.C.; Kaplan, Z.; Cohen, H. β - Alanine supplemented diets enhance behavioral resilience to stress exposure in an animal model of PTSD. Amino Acids 2015, 47, 1247–1257. [Google Scholar] [CrossRef]
- Stein, T.P.; Barnes, V.; Rhodes, N.; Guo, L.; Ming, X. Metabolic Perturbance in Autism Spectrum Disorders: A Metabolomics Study. J. Proteome 2012, 11, 5856–5862. [Google Scholar]
- Aldred, S.; Moore, K.M.; Fitzgerald, M.; Waring, R.H. Plasma Amino Acid Levels in Children with Autism and Their Families. J. Autism Dev. Disord. 2003, 33, 93–97. [Google Scholar] [CrossRef]
- Bala, K.A.; Doğan, M.; Mutluer, T.; Kaba, S.; Aslan, O.; Balahoroğlu, R.; Çokluk, E.; Üstyol, L.; Kocaman, S. Plasma amino acid profile in autism spectrum disorder (ASD). Eur. Rev. Med. Pharmacol. Sci. 2016, 20. [Google Scholar]
- Burrus, C.J. A biochemical rationale for the interaction between gastrointestinal yeast and autism. Med. Hypotheses 2012, 79, 784–785. [Google Scholar] [CrossRef] [PubMed]
- Delaye, J.B.; Patin, F.; Lagrue, E.; Le Tilly, O.; Bruno, C.; Vuillaume, M.L.; Raynaud, M.; Benz-De Bretagne, I.; Laumonnier, F.; Vourc’h, P.; et al. Post-hoc analysis of plasma amino acid profiles: Towards a specific pattern in autism spectrum disorder and intellectual disability. Ann. Clin. Biochem. 2018, 55, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Audhya, T.; McDonough-Means, S.; A Rubin, R.; Quig, D.; Geis, E.; Gehn, E.; Loresto, M.; Mitchell, J.; Atwood, S.; et al. Nutritional and metabolic status of children with autism vs. neurotypical children, and the association with autism severity. Nutr. Metab. 2011, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Jurynczyk, M.; Probert, F.; Yeo, T.; Tackley, G.; Claridge, T.D.W.; Cavey, A.; Woodhall, M.R.; Arora, S.; Winkler, T.; Schiffer, E.; et al. Metabolomics reveals distinct, antibody-independent, molecular signatures of MS, AQP4-antibody and MOG-antibody disease. Acta Neuropathol. Commun. 2017, 5, 95. [Google Scholar] [CrossRef] [PubMed]
- Wassif, W.; Sherwood, R.; Amir, A.; Idowu, B.; Summers, B.; Leigh, N.; Peters, T. Serum carnosinase activities in central nervous system disorders. Clin. Chim. Acta 1994, 225, 57–64. [Google Scholar] [CrossRef]
- Rao, M.L.; Stefan, H.; Scheid, C.; Kuttler, A.D.S.; Fröscher, W. Serum Amino Acids, Liver Status, and Antiepileptic Drug Therapy in Epilepsy. Epilepsia 1993, 34, 347–354. [Google Scholar] [CrossRef]
- Jung, C.S.; Lange, B.; Zimmermann, M.; Seifert, V. CSF and Serum Biomarkers Focusing on Cerebral Vasospasm and Ischemia after Subarachnoid Hemorrhage. Res. Treat. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Petroff, O.A.C.; Mattson, R.H.; Behar, K.L.; Hyder, F.; Rothman, D.L.; Hyder, D.S.F. Vigabatrin increases human brain homocarnosine and improves seizure control. Ann. Neurol. 1998, 44, 948–952. [Google Scholar] [CrossRef]
- Kori, M.; Ayidin, B.; Unal, S.; Arga, K.Y.; Kazan, D. Metabolic Biomarkers and Neurodegeneration: A Pathway Enrichment Analysis of Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. OMICS 2016, 20. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, A.N.; Harrington, R.J.; Tsai, A.; Liao, P.; Harrington, M.G. Free amino acid and dipeptide changes in the body fluids from Alzheimer’s disease subjects. Amino Acids 2007, 32, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Balion, C.M.; Benson, C.; Raina, P.S.; Papaioannou, A.; Patterson, C.; Ismaila, A.S. Brain type carnosinase in dementia: A pilot study. BMC Neurol. 2007, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Perrin, R.J.; Craig-Schapiro, R.; Malone, J.P.; Shah, A.R.; Gilmore, P.; Davis, A.E.; Roe, C.M.; Peskind, E.R.; Li, G.; Galasko, D.R.; et al. Identification and Validation of Novel Cerebrospinal Fluid Biomarkers for Staging Early Alzheimer’s Disease. PLoS ONE 2011, 6, e16032. [Google Scholar] [CrossRef] [PubMed]
- González-domínguez, R.; García-barrera, T.; Gómez-ariza, J.L. Journal of Pharmaceutical and Biomedical Analysis Metabolite profiling for the identification of altered metabolic pathways in Alzheimer’s disease. J. Pharm. Biomed. Anal. 2015, 107, 75–81. [Google Scholar] [CrossRef]
- Ibanez, C.; Simó, C.; Martin-Álvarez, P.J.; Kivipelto, M.; Winblad, B.; Cedazo-Minguez, A.; Cifuentes, A. Toward a Predictive Model of Alzheimer’s Disease Progression Using Capillary Electrophoresis–Mass Spectrometry Metabolomics. Anal. Chem. 2012, 84, 8532–8540. [Google Scholar] [CrossRef]
- Molina, J.A.; Jiménez-Jiménez, F.J.; Gómez, P.; Vargas, C.; Navarro, J.A.; Ortí-Pareja, M.; Gasalla, T.; Benito-León, J.; Bermejo, F.; Arenas, J. Decreased cerebrospinal fluid levels of neutral and basic amino acids in patients with Parkinson’s disease. J. Neurol. Sci. 1997, 150, 123–127. [Google Scholar] [CrossRef]
- Licker, V.; Côté, M.; Lobrinus, J.A.; Rodrigo, N.; Kövari, E.; Hochstrasser, D.F.; Turck, N.; Sanchez, J.-C.; Burkhard, P.R. Proteomic profiling of the substantia nigra demonstrates CNDP2 overexpression in Parkinson’s disease. J. Proteome 2012, 75, 4656–4667. [Google Scholar] [CrossRef]
- Luan, H.; Liu, L.; Meng, N.; Tang, Z.; Chua, K.K.; Chen, L.L.; Song, J.X.; Mok, V.C.; Xie, L.X.; Li, M.; et al. LC − MS-Based Urinary Metabolite Signatures in Idiopathic Parkinson’s Disease. J. Proteome Res. 2015, 14, 467–478. [Google Scholar] [CrossRef]
- Woo, H.-I.; Chun, M.-R.; Yang, J.-S.; Lim, S.-W.; Kim, M.-J.; Kim, S.-W.; Kim, D.-K.; Lee, S.-Y.; Myung, W.-J. Plasma Amino Acid Profiling in Major Depressive Disorder Treated With Selective Serotonin Reuptake Inhibitors. CNS Neurosci. Ther. 2015, 21, 417–424. [Google Scholar] [CrossRef]
- Altamura, C.; Maes, M.; Dai, J.; Meltzer, H. Plasma concentrations of excitatory amino acids, serine, glycine, taurine and histidine in major depression. Eur. Neuropsychopharmacol. 1995, 5, 71–75. [Google Scholar] [CrossRef]
- Chen, J.-J.; Zhou, C.-J.; Liu, Z.; Fu, Y.-Y.; Zheng, P.; Yang, D.-Y.; Li, Q.; Mu, J.; Wei, Y.-D.; Zhou, J.-J.; et al. Divergent Urinary Metabolic Phenotypes between Major Depressive Disorder and Bipolar Disorder Identified by a Combined GC–MS and NMR Spectroscopic Metabonomic Approach. J. Proteome 2015, 14, 3382–3389. [Google Scholar] [CrossRef] [PubMed]
- A Lewis, D.; A Lieberman, J. Catching up on schizophrenia: Natural history and neurobiology. Neuron 2000, 28. [Google Scholar]
- Saha, S.; Chant, D.; Welham, J.; McGrath, J. A Systematic Review of the Prevalence of Schizophrenia. PLoS Med. 2005, 2, e141. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yu, Z.; Giegling, I.; Xie, L.; Hartmann, A.M.; Prehn, C.; Adamski, J.; Kahn, R.; Li, Y.; Illig, T.; et al. Schizophrenia shows a unique metabolomics signature in plasma. Transl. Psychiatry 2012, 2, e149. [Google Scholar] [CrossRef]
- Leppik, L.; Kriisa, K.; Koido, K.; Koch, K.; Kajalaid, K.; Haring, L.; Vasar, E.; Zilmer, M. Profiling of Amino Acids and Their Derivatives Biogenic Amines Before and After Antipsychotic Treatment in First-Episode Psychosis. Front. Psychol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Hatano, T.; Ohnuma, T.; Sakai, Y.; Shibata, N.; Maeshima, H.; Hanzawa, R.; Suzuki, T.; Arai, H. Plasma alanine levels increase in patients with schizophrenia as their clinical symptoms improve—Results from the Juntendo University Schizophrenia Projects (JUSP). Psychiatry Res. 2010, 177, 27–31. [Google Scholar] [CrossRef]
- Chez, M.G.; Buchanan, C.P.; Aimonovitch, M.C.; Becker, M.; Schaefer, K.; Black, C.; Komen, J. Double-Blind, Placebo-Controlled Study of L-Carnosine Supplementation in Children With Autistic Spectrum Disorders. J. Child. Neurol. 2002, 17, 833–837. [Google Scholar] [CrossRef]
- Mehrazad-Saber, Z.; Kheirouri, S.; Noorazar, S.G. Effects of l-Carnosine Supplementation on Sleep Disorders and Disease Severity in Autistic Children: A Randomized, Controlled Clinical Trial. Basic Clin. Pharmacol. Toxicol. 2018, 123, 72–77. [Google Scholar] [CrossRef]
- Hajizadeh-Zaker, R.; Ghajar, A.; Mesgarpour, B.; Afarideh, M.; Mohammadi, M.R.; Akhondzadeh, S. l-Carnosine As an Adjunctive Therapy to Risperidone in Children with Autistic Disorder: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Child. Adolesc. Psychopharmacol. 2018, 28, 74–81. [Google Scholar] [CrossRef]
- Baraniuk, J.N.; El-Amin, S.; Corey, R.; Rayhan, R.; Timbol, C. Carnosine treatment for gulf war illness: A randomized controlled trial. J. Heal. Sci. 2013, 5, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Sasahara, I.; Fujimura, N.; Nozawa, Y.; Furuhata, Y.; Sato, H. The effect of histidine on mental fatigue and cognitive performance in subjects with high fatigue and sleep disruption scores. Physiol. Behav. 2015, 147, 238–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, J.R.; Landau, G.; Stout, J.R.; Hoffman, M.W.; Shavit, N.; Rosen, P.; Moran, D.S.; Fukuda, D.H.; Shelef, I.; Carmom, E.; et al. β - Alanine ingestion increases muscle carnosine content and combat specific performance in soldiers. Amino Acids 2015, 47, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Varanoske, A.N.; Wells, A.J.; Kozlowski, G.J.; Gepner, Y.; Frosti, C.L.; Boffey, D.; Coker, N.A.; Harat, I.; Hoffman, J.R. Effects of β-alanine supplementation on physical performance, cognition, endocrine function, and inflammation during a 24 h simulated military operation. Physiol. Rep. 2018, 6, e13938. [Google Scholar] [CrossRef] [PubMed]
- Furst, T.; Massaro, A.; Miller, C.; Williams, B.T.; Lamacchia, Z.M.; Horvath, P.J. β-Alanine supplementation increased physical performance and improved executive function following endurance exercise in middle aged individuals. J. Int. Soc. Sports Nutr. 2018, 15, 32. [Google Scholar] [CrossRef] [PubMed]
- Solis, M.Y.; Cooper, S.; Hobson, R.M.; Artioli, G.G.; Otaduy, M.C.; Roschel, H.; Robertson, J.; Martin, D.; Painelli, S.V.; Harris, R.C.; et al. Effects of Beta-Alanine Supplementation on Brain Homocarnosine/Carnosine Signal and Cognitive Function: An Exploratory Study. PLoS ONE 2015, 10, e0123857. [Google Scholar] [CrossRef]
- Szcześniak, D.; Budzeń, S.; Kopeć, W.; Rymaszewska, J.; Szczȩśniak, D. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Arch. Gerontol. Geriatr. 2014, 59, 485–490. [Google Scholar] [CrossRef]
- Ding, Q.; Tanigawa, K.; Kaneko, J.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/Carnosine Supplementation Preserves Blood Flow in the Prefrontal Brain of Elderly People Carrying APOE e4. Aging Dis. 2018, 9, 334–345. [Google Scholar] [CrossRef] [Green Version]
- Rokicki, J.; Li, L.; Imabayashi, E.; Kaneko, J.; Hisatsune, T.; Matsuda, H. Daily Carnosine and Anserine Supplementation Alters Verbal Episodic Memory and Resting State Network Connectivity in Healthy Elderly Adults. Front. Aging Neurosci. 2015, 7, 91. [Google Scholar] [CrossRef]
- Hisatsune, T.; Kaneko, J.; Kurashige, H.; Cao, Y.; Satsu, H.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H. Effect of Anserine/Carnosine Supplementation on Verbal Episodic Memory in Elderly People. J. Alzheimer’s Dis. 2016, 50, 149–159. [Google Scholar] [CrossRef]
- Small, B.J.; Rawson, K.S.; Martin, C.; Eisel, S.L.; Sanberg, C.D.; McEvoy, C.L.; Sanberg, P.R.; Shytle, R.D.; Tan, J.; Bickford, P.C. Nutraceutical Intervention Improves Older Adults’ Cognitive Functioning. Rejuvenation Res. 2014, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Cornelli, U. Treatment of Alzheimer’s Disease with a Cholinesterase Inhibitor Combined with Antioxidants. Neurodegener. Dis. 2010, 7, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Yamano, E.; Tanaka, M.; Ishii, A.; Tsuruoka, N.; Abe, K.; Watanabe, Y. Effects of chicken essence on recovery from mental fatigue in healthy males. Med. Sci. 2013, 19, 540–547. [Google Scholar] [Green Version]
- Koozehchian, M.S.; Earnest, C.P.; Jung, Y.P.; Collins, P.B.; O’Connor, A.; Dalton, R.; Shin, S.Y.; Sowinski, R.; Rasmussen, C.; Murano, P.S.; et al. Dose Response to One Week of Supplementation of a Multi-Ingredient Preworkout Supplement Containing Caffeine Before Exercise. J. Caffeine 2017, 7, 81–94. [Google Scholar] [CrossRef]
- Young, H.; Benton, D.; Carter, N. The Effect of Chicken Extract on Mood, Cognition and Heart Rate Variability. Nutrients 2015, 7, 887–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldyrev, A.; Fedorova, T.; Stepanova, M.; Dobrotvorskaya, I.; Kozlova, E.; Boldanova, N.; Bagyeva, G.; Ivanova-Smolenskaya, I.; Illarioshkin, S. Carnisone increases efficiency of DOPA therapy of Parkinson’s disease: A pilot study. Rejuvenation Res. 2008, 11, 821–827. [Google Scholar] [CrossRef]
- Ghajar, A.; Khoaie-Ardakani, M.-R.; Shahmoradi, Z.; Alavi, A.-R.; Afarideh, M.; Shalbafan, M.-R.; Ghazizadeh-Hashemi, M.; Akhondzadeh, S. L-carnosine as an add-on to risperidone for treatment of negative symptoms in patients with stable schizophrenia: A double-blind, randomized placebo-controlled trial. Psychiatry Res. 2018, 262, 94–101. [Google Scholar] [CrossRef]
- Chengappa, K.R.; Turkin, S.R.; DeSanti, S.; Bowie, C.R.; Brar, J.S.; Schlicht, P.J.; Murphy, S.L.; Hetrick, M.L.; Bilder, R.; Fleet, D. A preliminary, randomized, double-blind, placebo-controlled trial of l-carnosine to improve cognition in schizophrenia. Schizophr. Res. 2012, 142, 145–152. [Google Scholar] [CrossRef]
- Taylor, E.; Sergeant, J.; Asherson, P.; Banaschewski, T.; Buitelaar, J.; Coghill, D.; Danckaerts, M.; Rothenberger, A.; Sonuga-Barke, E.; cxvhausen, H.-C.; et al. European clinical guidelines for hyperkinetic disorder ? first upgrade. Eur. Child. Adolesc. Psychiatry 2004, 13, i7–i30. [Google Scholar] [CrossRef]
- Ghajar, A.; Aghajan-Nashtaei, F.; Afarideh, M.; Mohammadi, M.R.; Akhondzadeh, S. L-carnosine as Adjunctive Therapy in Children and Adolescents with Attention-Deficit/Hyperactivity Disorder. J. Clin. Child. Adoles. Psychol. 2018, 28, 331–338. [Google Scholar] [CrossRef]
- Rajendram, R.; Kronenberg, S.; Burton, C.L.; Arnold, P.D. Glutamate Genetics in Obsessive-Compulsive Disorder: A Review. J. Can. Acad. Child. Adolesc. Psychiatry 2017, 26, 205–213. [Google Scholar]
- Arabzadeh, S.; Shahhossenie, M.; Mesgarpour, B.; Rezaei, F.; Shalbafan, M.R.; Ghiasi, Z.; Akhondzadeh, S. L-carnosine as an adjuvant to fluvoxamine in treatment of obsessive compulsive disorder: A randomized double-blind study. Hum. Psychopharmacol. Clin. Exp. 2017, 32, e2584. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Tian, Y.; Bao, Y.; Xu, H.; Cheng, J.; Wang, B.; Shen, Y.; Chen, Z.; Lyu, J. Carnosine decreased neuronal cell death through targeting glutamate system and astrocyte mitochondrial bioenergetics in cultured neuron/astrocyte exposed to OGD/recovery. Brain Res. Bull. 2016, 124, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Shirotsuki, K.; Nonaka, Y.; Abe, K.; Adachi, S.I.; Adachi, S.; Kuboki, T.; Nakao, M. The effect for Japanese workers of a self- help computerized cognitive behaviour therapy program with a supplement soft drink. BioPsychoSoc. Med. 2017, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Kairaluoma, L.; Närhi, V.; Ahonen, T.; Westerholm, J.; Aro, M. Do fatty acids help in overcoming reading difficulties? A double-blind, placebo-controlled study of the effects of eicosapentaenoic acid and carnosine supplementation on children with dyslexia. Child. Care Health Dev. 2009, 35, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.B.; Wessely, S.; Kuh, D.; Hotopf, M. The relationship between fatigue and psychiatric disorders: Evidence for the concept of neurasthenia. J. Psychosom. 2009, 66, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.; Maes, M.; Berk, M.; Puri, B.K. Myalgic encephalomyelitis or chronic fatigue syndrome: How could the illness develop? Metab. Brain 2019, 34, 385–415. [Google Scholar] [CrossRef] [PubMed]
- Dannaway, J.; New, C.C.; New, C.H.; Maher, C.G. Exercise therapy is a beneficial intervention for chronic fatigue syndrome (PEDro synthesis). Br. J. Sports Med. 2018, 52, 542–543. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schön, M.; Mousa, A.; Berk, M.; Chia, W.L.; Ukropec, J.; Majid, A.; Ukropcová, B.; de Courten, B. The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence. Nutrients 2019, 11, 1196. https://doi.org/10.3390/nu11061196
Schön M, Mousa A, Berk M, Chia WL, Ukropec J, Majid A, Ukropcová B, de Courten B. The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence. Nutrients. 2019; 11(6):1196. https://doi.org/10.3390/nu11061196
Chicago/Turabian StyleSchön, Martin, Aya Mousa, Michael Berk, Wern L. Chia, Jozef Ukropec, Arshad Majid, Barbara Ukropcová, and Barbora de Courten. 2019. "The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence" Nutrients 11, no. 6: 1196. https://doi.org/10.3390/nu11061196