Treadmill Exercise before and during Pregnancy Improves Bone Deficits in Pregnant Growth Restricted Rats without the Exacerbated Effects of High Fat Diet


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
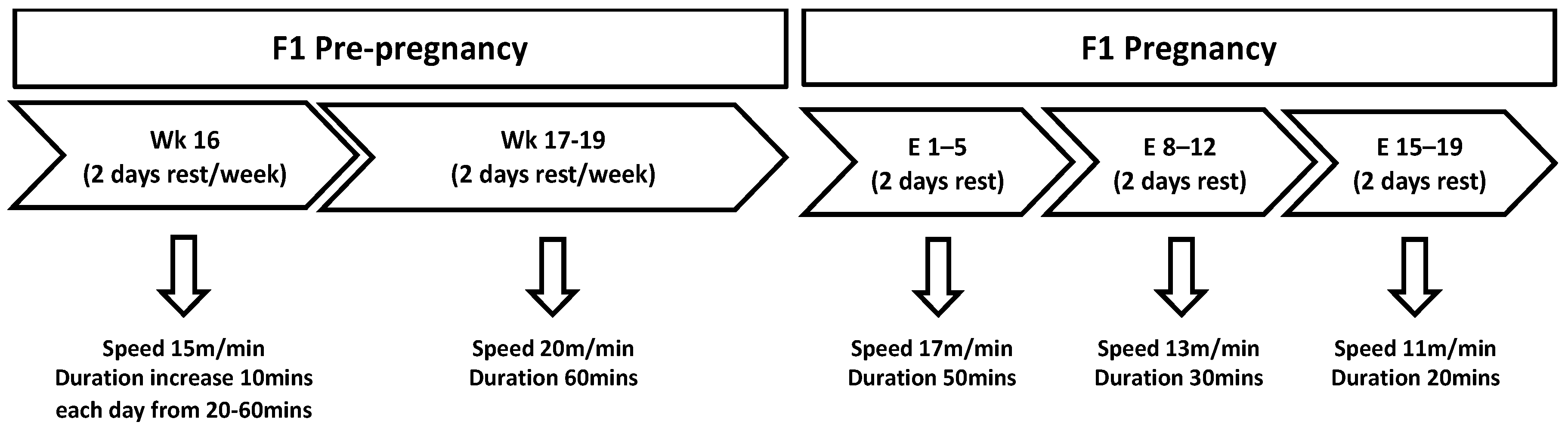
2.2. Exercise Protocol
2.3. Post-Mortem Blood and Tissue Collection
2.4. Peripheral Quantitative Computed Tomography (pQCT)
2.5. Histological Processing, Staining, and Histomorphometry
2.6. Mechanical Testing
2.7. Plasma Analysis
2.8. Statistical Analyses
3. Results
3.1. Body Weight
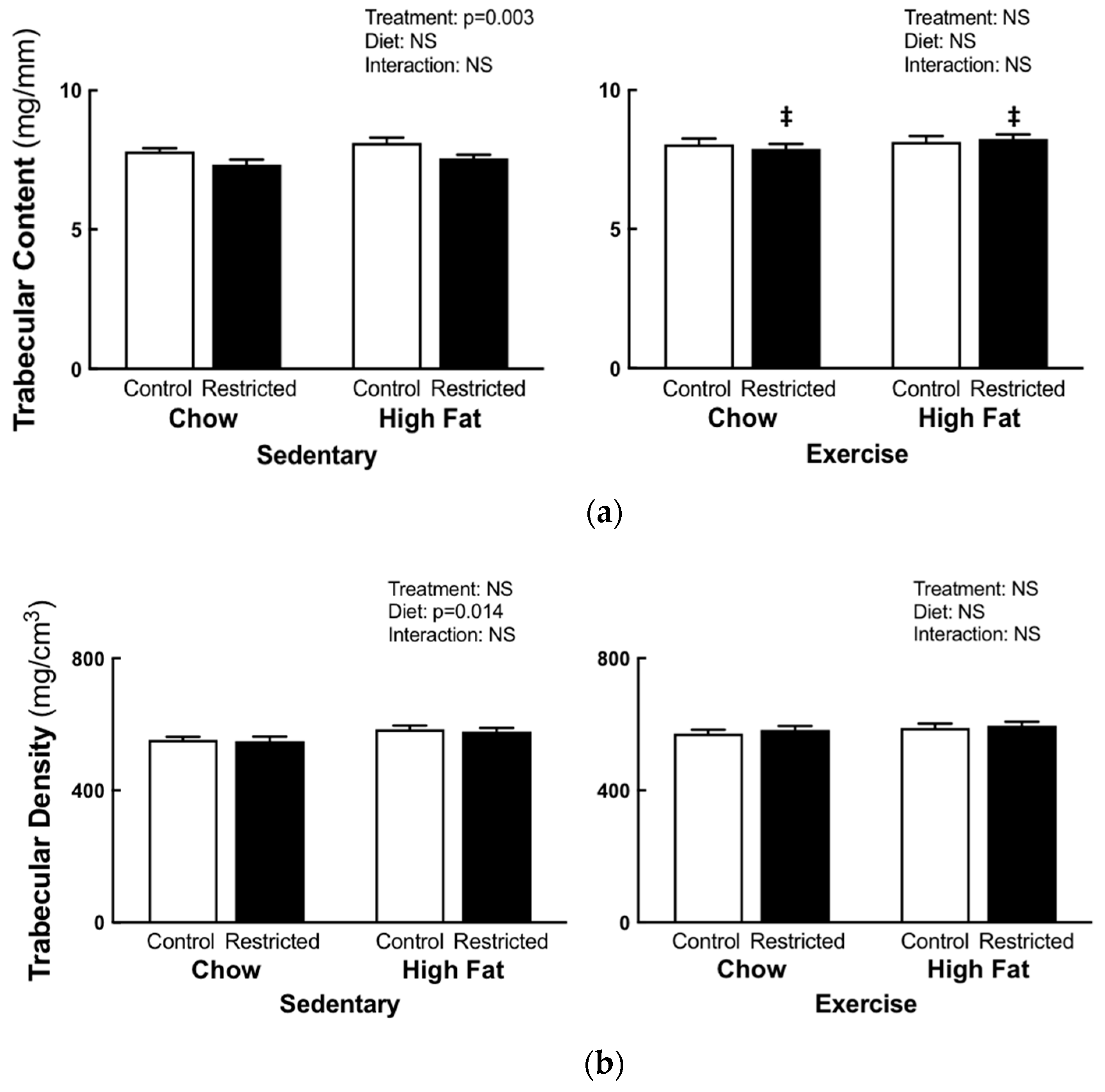
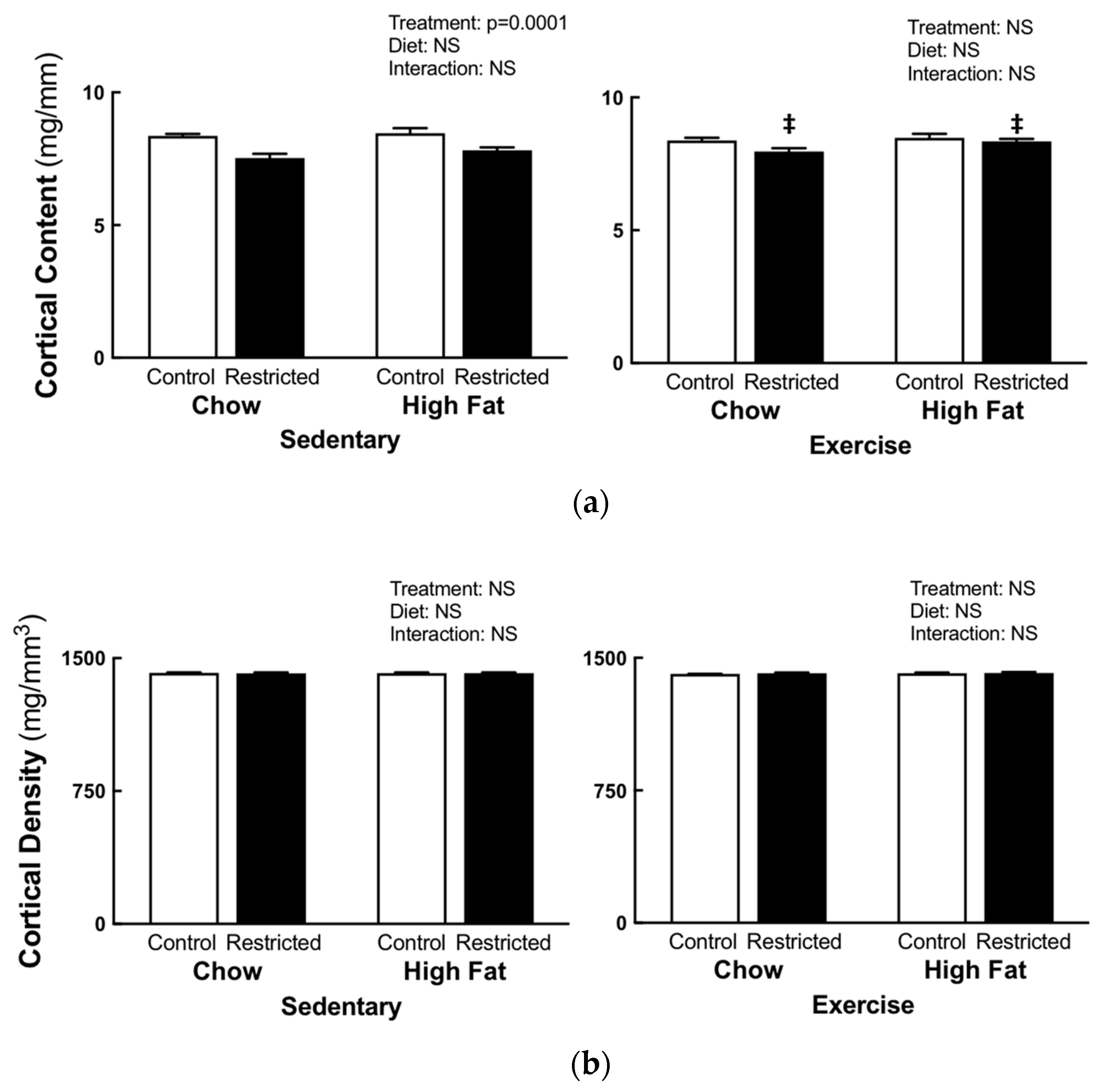
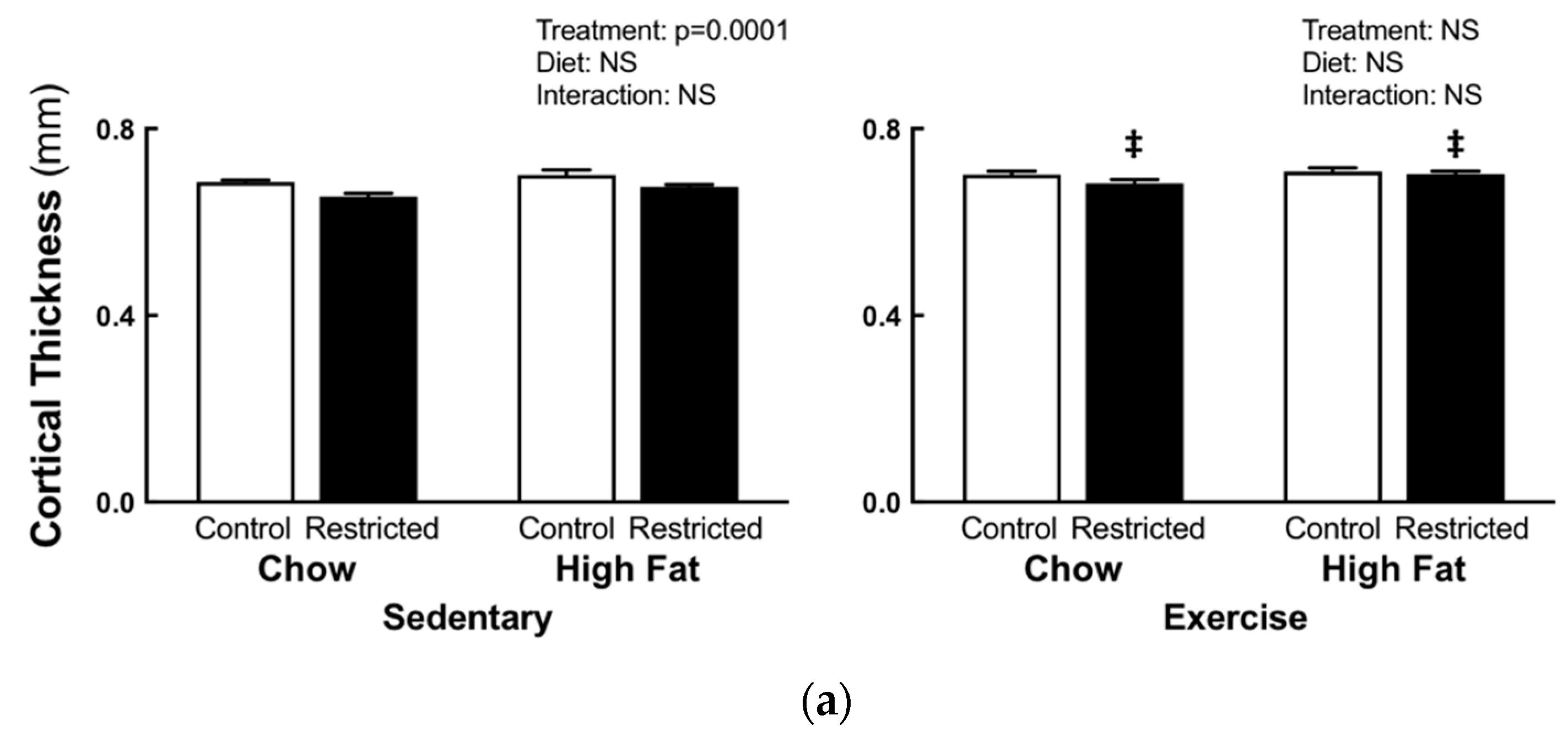
3.2. Peripheral Quantitative Computed Tomography
3.3. Histomorphometric Analysis and Mechanical Testing
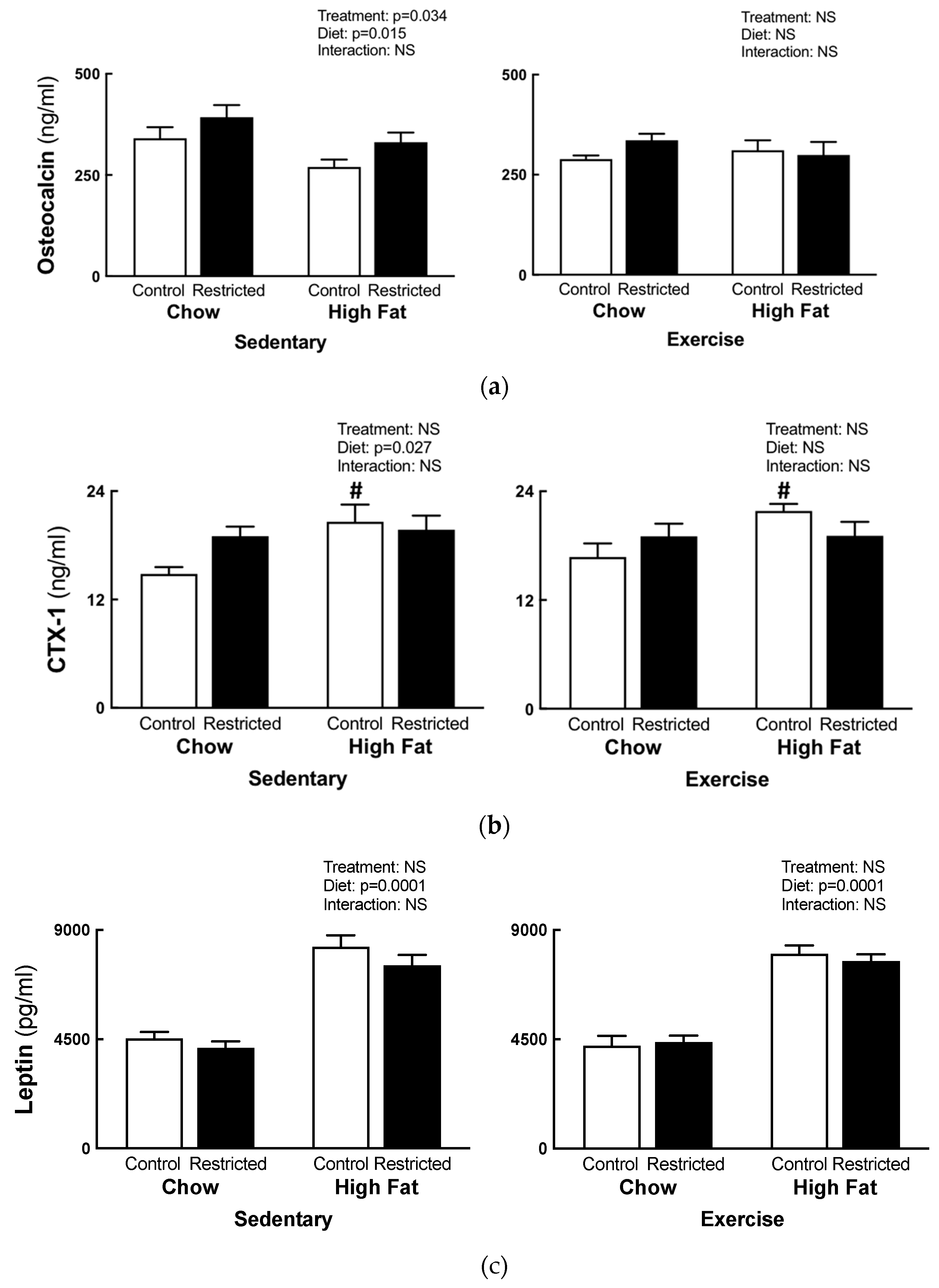
3.4. Plasma Analysis
4. Discussion
4.1. Bone Health in Pregnant Growth Restricted Females
4.2. Bone Health Following High-fat Feeding
4.3. Exercise Effects on Bone Health
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tojo, Y.; Kurabayashi, T.; Honda, A.; Yamamoto, Y.; Yahata, T.; Takakuwa, K.; Tanaka, K. Bone structural and metabolic changes at the end of pregnancy and lactation in rats. Am. J. Obstet. Gynecol. 1998, 178, 180–185. [Google Scholar] [CrossRef]
- Miller, S.C.; Bowman, B.M. Comparison of bone loss during normal lactation with estrogen deficiency osteopenia and immobilization osteopenia in the rat. Anat. Rec. 1998, 251, 265–274. [Google Scholar] [CrossRef]
- Monteiro, A.C.T.; Paes, S.T.; Dos Santos, J.A.; De Lira, K.D.S.; De Moraes, S.R.A. Effects of physical exercise during pregnancy and protein malnutrition during pregnancy and lactation on the development and growth of the offspring’s femur. J. Pediatr. (Rio J.) 2010, 86, 233–238. [Google Scholar] [CrossRef]
- Kimura, T.; Hino, K.; Kono, T.; Takano, A.; Nitta, N.; Ushio, N.; Hino, S.; Takase, R.; Kudo, M.; Daigo, Y.; et al. Maternal undernutrition during early pregnancy inhibits postnatal growth of the tibia in the female offspring of rats by alteration of chondrogenesis. Comp. Endocrinol. 2018, 260, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Romano, T.; Hryciw, D.H.; Westcott, K.T.; Wlodek, M.E. Puberty onset is delayed following uteroplacental insufficiency and occurs earlier with improved lactation and growth for pups born small. Reprod. Fertil. Dev. 2017, 29, 307. [Google Scholar] [CrossRef]
- Romano, T.; Wark, J.D.; Wlodek, M.E. Developmental programming of bone deficits in growth-restricted offspring. Reprod. Fertil. Dev. 2015, 27, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Romano, T.; Wark, J.D.; Wlodek, M.E. Physiological skeletal gains and losses in rat mothers during pregnancy and lactation are not observed following uteroplacental insufficiency. Reprod. Fertil. Dev. 2014, 26, 385–394. [Google Scholar] [CrossRef]
- Romano, T.; Wark, J.D.; Wlodek, M.E. Calcium supplementation does not rescue the programmed adult bone deficits associated with perinatal growth restriction. Bone 2010, 47, 1054–1063. [Google Scholar] [CrossRef]
- Romano, T.; Wark, J.D.; Owens, J.A.; Wlodek, M.E. Prenatal growth restriction and postnatal growth restriction followed by accelerated growth independently program reduced bone growth and strength. Bone 2009, 45, 132–141. [Google Scholar] [CrossRef]
- Engelbregt, M.J.; Van Weissenbruch, M.M.; Lips, P.; Van Lingen, A.; Roos, J.C.; Waal, H.A.D.-V.D. Body composition and bone measurements in intra-uterine growth retarded and early postnatally undernourished male and female rats at the age of 6 months: comparison with puberty. Bone 2004, 34, 180–186. [Google Scholar] [CrossRef]
- Robinson, J.S.; Moore, V.M.; Owens, J.A.; McMillen, I. Origins of fetal growth restriction. Eur. J. Obstet. Gynecol. Reprod. Boil. 2000, 92, 13–19. [Google Scholar] [CrossRef]
- Berghella, V. Prevention of Recurrent Fetal Growth Restriction. Obstet. Gynecol. 2007, 110, 904–912. [Google Scholar] [CrossRef]
- Martin, J.A.; Hamilton, B.E.; Sutton, P.D.; Ventura, S.J.; Menacker, F.; Kimeyer, S.; Munson, M.L. Centers for Disease Control and Prevention National Center for Health Statistics National Vital Statistics System. Births: Final data for 2005. Natl. Vital Stat. Rep. 2007, 56, 1–103. [Google Scholar]
- Gallo, L.A.; Tran, M.; Master, J.S.; Moritz, K.M.; Wlodek, M.E. Maternal adaptations and inheritance in the transgenerational programming of adult disease. Cell Tissue Res. 2012, 349, 863–880. [Google Scholar] [CrossRef]
- Oliver, H.; Jameson, K.A.; Sayer, A.A.; Cooper, C.; Dennison, E.M. Growth in early life predicts bone strength in late adulthood: The Hertfordshire Cohort Study. Bone 2007, 41, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Kensara, O.A.; Wootton, S.A.; Phillips, D.I.; Patel, M.; Jackson, A.A.; Elia, M.; Hertfordshire Study Group. Fetal programming of body composition: relation between birth weight and body composition measured with dual-energy X-ray absorptiometry and anthropometric methods in older Englishmen. Am. J. Clin. Nutr. 2005, 82, 980–987. [Google Scholar] [Green Version]
- Jimenez-Chillaron, J.C.; Isganaitis, E.; Charalambous, M.; Gesta, S.; Pentinat-Pelegfrin, T.; Faucette, R.R.; Otis, J.P.; Chow, A.; Diaz, R.; Ferguson-Smith, A.; et al. Intergenerational transmission of glucose intolerance and obesity by in utero undernutrition in mice. Diabetes 2009, 58, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Anevska, K.; Gallo, L.A.; Tran, M.; Jefferies, A.J.; Wark, J.D.; Wlodek, M.E.; Romano, T. Pregnant growth restricted female rats have bone gains during late gestation which contributes to second generation adolescent and adult offspring having normal bone health. Bone 2015, 74, 199–207. [Google Scholar] [CrossRef]
- Wlodek, M.E.; Westcott, K.T.; O’Dowd, R.; Serruto, A.; Wassef, L.; Moritz, K.M.; Moseley, J.M. Uteroplacental restriction in the rat impairs fetal growth in association with alterations in placental growth factors including PTHrP. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R1620–R1627. [Google Scholar] [CrossRef]
- O’Dowd, R.; Kent, J.C.; Moseley, J.M.; Wlodek, M.E. Effects of uteroplacental insufficiency and reducing litter size on maternal mammary function and postnatal offspring growth. Am. J. Physiol. Integr. Comp. Physiol. 2008, 294, R539–R548. [Google Scholar] [CrossRef]
- Cao, J.J. Effects of obesity on bone metabolism. J. Orthop. Surg. 2011, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Frost, H. Obesity, and bone strength and “mass”: A tutorial based on insights from a new paradigm. Bone 1997, 21, 211–214. [Google Scholar] [CrossRef]
- Villareal, D.T.; Apovian, C.M.; Kushner, R.F.; Klein, S. Obesity in older adults: technical review and position statement of the American Society for Nutrition and NAASO, The Obesity Society. Am. J. Clin. Nutr. 2005, 82, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Malvi, P.; Piprode, V.; Chaube, B.; Pote, S.T.; Mittal, M.; Chattopadhyay, N.; Wani, M.R.; Bhat, M.K. High fat diet promotes achievement of peak bone mass in young rats. Biochem. Biophys. Commun. 2014, 455, 133–138. [Google Scholar] [CrossRef]
- Li, W.; Xu, P.; Wang, C.; Ha, X.; Gu, Y.; Wang, Y.; Zhang, J.; Xie, J. The effects of fat-induced obesity on bone metabolism in rats. Obes. Clin. Pr. 2017, 11, 454–463. [Google Scholar] [CrossRef]
- Goulding, A.; Grant, A.M.; Williams, S.M. Bone and Body Composition of Children and Adolescents With Repeated Forearm Fractures. J. Bone 2005, 20, 2090–2096. [Google Scholar] [CrossRef]
- Prieto-Alhambra, D.; Premaor, M.O.; Avilés, F.F.; Hermosilla, E.; Martinez-Laguna, D.; Carbonell-Abella, C.; Nogués, X.; Compston, J.E.; Diez-Perez, A.; Prieto-Alhambra, D.; et al. The association between fracture and obesity is site-dependent: A population-based study in postmenopausal women. J. Bone 2012, 27, 294–300. [Google Scholar] [CrossRef]
- Cohen, A.; Dempster, D.W.; Recker, R.R.; Lappe, J.M.; Zhou, H.; Zwahlen, A.; Müller, R.; Zhao, B.; Guo, X.; Lang, T.; et al. Abdominal Fat Is Associated With Lower Bone Formation and Inferior Bone Quality in Healthy Premenopausal Women: A Transiliac Bone Biopsy Study. J. Clin. Endocrinol. Metab. 2013, 98, 2562–2572. [Google Scholar] [CrossRef]
- Patsch, J.M.; Kiefer, F.W.; Varga, P.; Pail, P.; Rauner, M.; Stupphann, D.; Resch, H.; Moser, D.; Zysset, P.K.; Stulnig, T.M.; et al. Increased bone resorption and impaired bone microarchitecture in short-term and extended high-fat diet-induced obesity. Metabolism 2011, 60, 243–249. [Google Scholar] [CrossRef]
- Lu, X.M.; Zhao, H.; Wang, E.H. A high-fat diet induces obesity and impairs bone acquisition in young male mice. Mol. Med. Rep. 2013, 7, 1203–1208. [Google Scholar] [CrossRef]
- Cao, J.J.; Gregoire, B.R.; Shen, C.-L. A High-Fat Diet Decreases Bone Mass in Growing Mice with Systemic Chronic Inflammation Induced by Low-Dose, Slow-Release Lipopolysaccharide Pellets. J. Nutr. 2017, 147, 1909–1916. [Google Scholar] [CrossRef]
- Doucette, C.R.; Horowitz, M.C.; Berry, R.; MacDougald, O.A.; Koza, R.A.; Rosen, C.J.; Anunciado-Koza, R.; Anunciado-Koza, R. A high fat diet increases bone marrow adipose tissue (MAT) but does not alter trabecular or cortical bone mass in C57BL/6J mice. J. Cell. Physiol. 2015, 230, 2032–2037. [Google Scholar] [CrossRef]
- Saville, P.D.; Whyte, M.P. Muscle and bone hypertrophy. Positive effect of running exercise in the rat. Clin. Orthop. Relat. 1969, 65, 81–88. [Google Scholar]
- Woo, S.L.; Kuei, S.C.; Amiel, D.; Gomez, M.A.; Hayes, W.C.; White, F.C.; Akeson, W.H. The effect of prolonged physical training on the properties of long bone: A study of Wolff’s Law. J. Bone Jt. Surg. 1981, 63, 780–787. [Google Scholar] [CrossRef]
- Chen, M.M.; Yeh, J.K.; Aloia, J.F.; Tierney, J.M.; Sprintz, S. Effect of treadmill exercise on tibial cortical bone in aged female rats: A histomorphometry and dual energy X-ray absorptiometry study. Bone 1994, 15, 313–319. [Google Scholar] [CrossRef]
- Snyder, A.; Zierath, J.R.; Hawley, J.A.; Sleeper, M.D.; Craig, B.W. The effects of exercise mode, swimming vs. Running, upon bone growth in the rapidly growing female rat. Mech. Ageing Dev. 1992, 66, 59–69. [Google Scholar] [CrossRef]
- Iwamoto, J.; Shimamura, C.; Takeda, T.; Abe, H.; Ichimura, S.; Sato, Y.; Toyama, Y. Effects of treadmill exercise on bone mass, bone metabolism, and calciotropic hormones in young growing rats. J. Bone Miner. Metab. 2004, 22, 26–31. [Google Scholar] [CrossRef]
- Liu, S.Y.; Li, Z.; Xu, S.-Y.; Xu, L.; Yang, M.; Ni, G.-X. Intensity-dependent effect of treadmill running on differentiation of rat bone marrow stromal cells. Mol. Med. Rep. 2018, 17, 7746–7756. [Google Scholar] [CrossRef]
- Laker, R.C.; Wlodek, M.E.; Wadley, G.D.; Gallo, L.A.; Meikle, P.J.; McConell, G.K. Exercise early in life in rats born small does not normalize reductions in skeletal muscle pgc-1α in adulthood. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1221–E1230. [Google Scholar] [CrossRef]
- Mangwiro, Y.T.M.; Cuffe, J.S.M.; Briffa, J.F.; Mahizir, D.; Anevska, K.; Jefferies, A.J.; Hosseini, S.; Romano, T.; Moritz, K.M.; Wlodek, M.E. Maternal exercise in rats upregulates the placental insulin-like growth factor system with diet- and sex-specific responses: Minimal effects in mothers born growth restricted. J. Physiol. 2018, 596, 5947–5964. [Google Scholar] [CrossRef]
- Anevska, K.; Cheong, J.N.; Wark, J.D.; Wlodek, M.E.; Romano, T. Maternal stress does not exacerbate long-term bone deficits in female rats born growth restricted, with differential effects on offspring bone health. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R161–R170. [Google Scholar] [CrossRef]
- Brady, R.D.; Grills, B.L.; Church, J.E.; Walsh, N.C.; McDonald, A.C.; Agoston, D.V.; Sun, M.; O’Brien, T.J.; Shultz, S.R.; McDonald, S.J. Closed head experimental traumatic brain injury increases size and bone volume of callus in mice with concomitant tibial fracture. Sci. Rep. 2016, 6, 34491. [Google Scholar] [CrossRef]
- Brady, R.D.; Grills, B.L.; Romano, T.; Wark, J.D.; O’Brien, T.J.; Shultz, S.R.; McDonald, S.J. Sodium selenate treatment mitigates reduction of bone volume following traumatic brain injury in rats. J. Musculoskelet. Neuronal Interact. 2016, 16, 369–376. [Google Scholar]
- Seibel, M.J. Biochemical markers of bone turnover part 1: Biochemistry and variability. Clin. Biochem. Rev. 2005, 26, 97–122. [Google Scholar]
- Krawczewski Carhuatanta, K.A.; Demuro, G.; Tschop, M.H.; Pfluger, P.T.; Benoit, S.C.; Obici, S. Voluntary exercise improves high-fat diet-induced leptin resistance independent of adiposity. Endocrinology 2011, 152, 2655–2664. [Google Scholar] [CrossRef]
- Verhaeghe, J.; Van Herck, E.; Van Bree, R.; Van Assche, F.A.; Bouillon, R. Osteocalcin during the reproductive cycle in normal and diabetic rats. J. Endocrinol. 1989, 120, 143–151. [Google Scholar] [CrossRef]
- Oury, F.; Khrimian, L.; Denny, C.A.; Gardin, A.; Chamouni, A.; Goeden, N.; Huang, Y.Y.; Lee, H.; Srinivas, P.; Gao, X.B.; et al. Maternal and offspring pools of osteocalcin influence brain development and functions. Cell 2013, 155, 228–241. [Google Scholar] [CrossRef]
- Ducy, P. The role of osteocalcin in the endocrine cross-talk between bone remodelling and energy metabolism. Diabetologia 2011, 54, 1291–1297. [Google Scholar] [CrossRef] [Green Version]
- Firth, E.C.; Gamble, G.D.; Cornish, J.; Vickers, M.H. Neonatal leptin treatment reverses the bone-suppressive effects of maternal undernutrition in adult rat offspring. Sci. Rep. 2017, 7, 7686. [Google Scholar] [CrossRef]
- Chen, J.R.; Lazarenko, O.P.; Wu, X.; Tong, Y.; Blackburn, M.L.; Shankar, K.; Badger, T.M.; Ronis, M.J.J. Obesity reduces bone density associated with activation of ppargamma and suppression of wnt/beta-catenin in rapidly growing male rats. PLoS ONE 2010, 5, e13704. [Google Scholar] [CrossRef]
- Lau, B.Y.; Fajardo, V.A.; McMeekin, L.; Sacco, S.M.; Roy, B.D.; Peters, S.J.; Ward, W.E.; Leblanc, P.J. Influence of high-fat diet from differential dietary sources on bone mineral density, bone strength, and bone fatty acid composition in rats. Appl. Physiol. Nutr. Metab. 2010, 35, 598–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathey, J.; Horcajada-Molteni, M.-N.; Chanteranne, B.; Picherit, C.; Puel, C.; Lebecque, P.; Cubizoles, C.; Davicco, M.-J.; Coxam, V.; Barlet, J.-P. Bone Mass in Obese Diabetic Zucker Rats: Influence of Treadmill Running. Calcif. Tissue Int. 2002, 70, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, J.; Yeh, J.; Aloia, J. Differential effect of treadmill exercise on three cancellous bone sites in the young growing rat. Bone 1999, 24, 163–169. [Google Scholar] [CrossRef]
- Yeh, J.K.; Aloia, J.F.; Chen, M.; Ling, N.; Koo, H.C.; Millard, W.J. Effect of growth hormone administration and treadmill exercise on serum and skeletal igf-i in rats. Am. J. Physiol. Endocrinol. Metab. 1994, 266, E129–135. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Ferron, M. The contribution of bone to whole-organism physiology. Nature 2012, 481, 314. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okimoto, N.; Okazaki, Y.; Tanaka, Y.; Nakamura, T.; Suzuki, M. Effects of tower climbing exercise on bone mass, strength, and turnover in growing rats. J. Bone Miner. Res. 2001, 16, 166–174. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Postnatal Weight (g) | Control | Restricted |
|---|---|---|
| PN1 | 4.22 ± 0.04 | 3.49 ± 0.02 * |
| PN7 | 9.88 ± 0.16 | 7.99 ± 0.17 * |
| PN14 | 22.23 ± 0.26 | 18.83 ± 0.38 * |
| PN35 | 76.36 ± 0.62 | 68.36 ± 0.80 * |
| Two-Way ANOVA | ||||||
|---|---|---|---|---|---|---|
| Chow | HFD | Treatment | Diet | Interaction | ||
| 15 week Body Weight (g) | ||||||
| Sedentary | Control | 232 ± 4.0 | 239 ± 7.0 | p = 0.0001 | NS | NS |
| Restricted | 213 ± 2.0 | 217 ± 7.8 | ||||
| Exercise | Control | 225 ± 4.1 | 242 ± 2.3 | p = 0.020 | p = 0.0001 | NS |
| Restricted | 217 ± 2.6 | 234 ± 2.5 | ||||
| 19 week Body Weight (g) | ||||||
| Sedentary | Control | 248 ± 4.5 | 266 ± 7.0 | p = 0.001 | p = 0.026 | NS |
| Restricted | 230 ± 2.4 | 240 ± 8.3 | ||||
| Exercise | Control | 258 ± 3.1 | 275 ± 5.3 | p = 0.022 | p = 0.0001 | NS |
| Restricted | 247 ± 3.3 | 269 ± 2.1 | ||||
| Mating Body Weight (g) | ||||||
| Sedentary | Control | 258 ± 4.7 | 270 ± 8.0 | p = 0.002 | NS | NS |
| Restricted | 233 ± 6.0 | 249 ± 7.5 | ||||
| Exercise | Control | 264 ± 3.7 | 280 ± 6.8 | p = 0.042 | p = 0.001 | NS |
| Restricted | 251 ± 4.3 | 272 ± 4.5 | ||||
| E20 Body Weight (g) | ||||||
| Sedentary | Control | 347 ± 5.3 | 360 ± 10.0 | p = 0.0001 | p = 0.030 | NS |
| Restricted | 314 ± 4.1 | 335 ± 8.6 | ||||
| Exercise | Control | 346 ± 3.8 | 377 ± 7.7 | p = 0.011 | p = 0.0001 | NS |
| Restricted | 332 ± 5.7 | 359 ± 5.9 | ||||
| Pregnancy Weight Gain (g) | ||||||
| Sedentary | Control | 89 ± 3.5 | 90 ± 4.1 | p = 0.027 | NS | NS |
| Restricted | 76 ± 2.1 | 87 ± 2.3 | ||||
| Exercise | Control | 82 ± 1.8 | 97 ± 3.1 | NS | p = 0.006 | NS |
| Restricted | 81 ± 2.9 | 87 ± 5.4 | ||||
| Two-Way ANOVA | ||||||
|---|---|---|---|---|---|---|
| Chow | HFD | Treatment | Diet | Interaction | ||
| R. Femur Length (mm) | ||||||
| Sedentary | Control | 34.3 ± 0.2 | 34.2 ± 0.2 | NS | NS | NS |
| Restricted | 33.5 ± 0.2 | 33.4 ± 0.2 | ||||
| Exercise | Control | 34.9 ± 0.1 | 34.6 ± 0.3 | NS | NS | NS |
| Restricted | 34.1 ± 0.1 | 34.6 ± 0.2 | ||||
| Bone Area (%) | ||||||
| Sedentary | Control | 16 ± 3.24 | 22.7 ± 2.2 | NS | NS | NS |
| Restricted | 18.6 ± 2.4 | 22 ± 2.9 | ||||
| Exercise | Control | 19.5 ± 1.6 | 21.7 ± 2.7 | NS | NS | NS |
| Restricted | 19 ± 1.3 | 20.5 ± 3.7 | ||||
| Calcified Cartilage (%) | ||||||
| Sedentary | Control | 9.3 ± 2.2 | 9.5 ± 1.1 | NS | NS | NS |
| Restricted | 6.2 ± 2.3 | 8.4 ± 2.3 | ||||
| Exercise | Control | 6.8 ± 1.8 | 9.5 ± 0.04 | NS | NS | NS |
| Restricted | 6.6 ± 1.6 | 10.3 ± 1.5 | ||||
| Growth Plate (μm) | ||||||
| Sedentary | Control | 121 ± 5.8 | 123 ± 6.4 | NS | NS | NS |
| Restricted | 138 ± 14.6 | 129.6 ± 6.2 | ||||
| Exercise | Control | 126 ± 8.1 | 128.8 ± 2.9 | NS | NS | NS |
| Restricted | 116 ± 6.5 | 123 ± 11.5 | ||||
| Two-Way ANOVA | ||||||
|---|---|---|---|---|---|---|
| Chow | HFD | Treatment | Diet | Interaction | ||
| L. Femur Length (mm) | ||||||
| Sedentary | Control | 34.4 ± 0.2 | 34.0 ± 0.3 | NS | NS | NS |
| Restricted | 33.5 ± 0.2 | 33.5 ± 0.2 | ||||
| Exercise | Control | 34.5 ± 0.2 | 34.7 ± 0.2 | NS | NS | NS |
| Restricted | 34.0 ± 0.2 | 34.3 ± 0.2 | ||||
| Peak Force (N) | ||||||
| Sedentary | Control | 263.5 ± 16.0 | 293.9 ± 10.9 | NS | p = 0.003 | NS |
| Restricted | 247.2 ± 19.5 | 287.8 ± 12.9 | ||||
| Exercise | Control | 313.2 ± 2.8 | 286.1 ± 18.1 | NS | NS | NS |
| Restricted | 270.9 ± 12.3 | 299.0 ± 22.5 | ||||
| Bending Stress (× 106 m2) | ||||||
| Sedentary | Control | 107 ± 6.6 | 117 ± 4.4 | NS | p = 0.003 | NS |
| Restricted | 100 ± 5.7 | 125 ± 4.6 | ||||
| Exercise | Control | 115.5 ± 8.5 | 115.1 ± 6.9 | NS | NS | NS |
| Restricted | 109.6 ± 4.5 | 122.3 ± 9.1 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anevska, K.; Mahizir, D.; Briffa, J.F.; Jefferies, A.J.; Wark, J.D.; Grills, B.L.; Brady, R.D.; McDonald, S.J.; Wlodek, M.E.; Romano, T. Treadmill Exercise before and during Pregnancy Improves Bone Deficits in Pregnant Growth Restricted Rats without the Exacerbated Effects of High Fat Diet. Nutrients 2019, 11, 1236. https://doi.org/10.3390/nu11061236
Anevska K, Mahizir D, Briffa JF, Jefferies AJ, Wark JD, Grills BL, Brady RD, McDonald SJ, Wlodek ME, Romano T. Treadmill Exercise before and during Pregnancy Improves Bone Deficits in Pregnant Growth Restricted Rats without the Exacerbated Effects of High Fat Diet. Nutrients. 2019; 11(6):1236. https://doi.org/10.3390/nu11061236
Chicago/Turabian StyleAnevska, Kristina, Dayana Mahizir, Jessica F. Briffa, Andrew J. Jefferies, John D. Wark, Brian L. Grills, Rhys D. Brady, Stuart J. McDonald, Mary E. Wlodek, and Tania Romano. 2019. "Treadmill Exercise before and during Pregnancy Improves Bone Deficits in Pregnant Growth Restricted Rats without the Exacerbated Effects of High Fat Diet" Nutrients 11, no. 6: 1236. https://doi.org/10.3390/nu11061236