Lactobacillus reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: The PROSIR Study

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
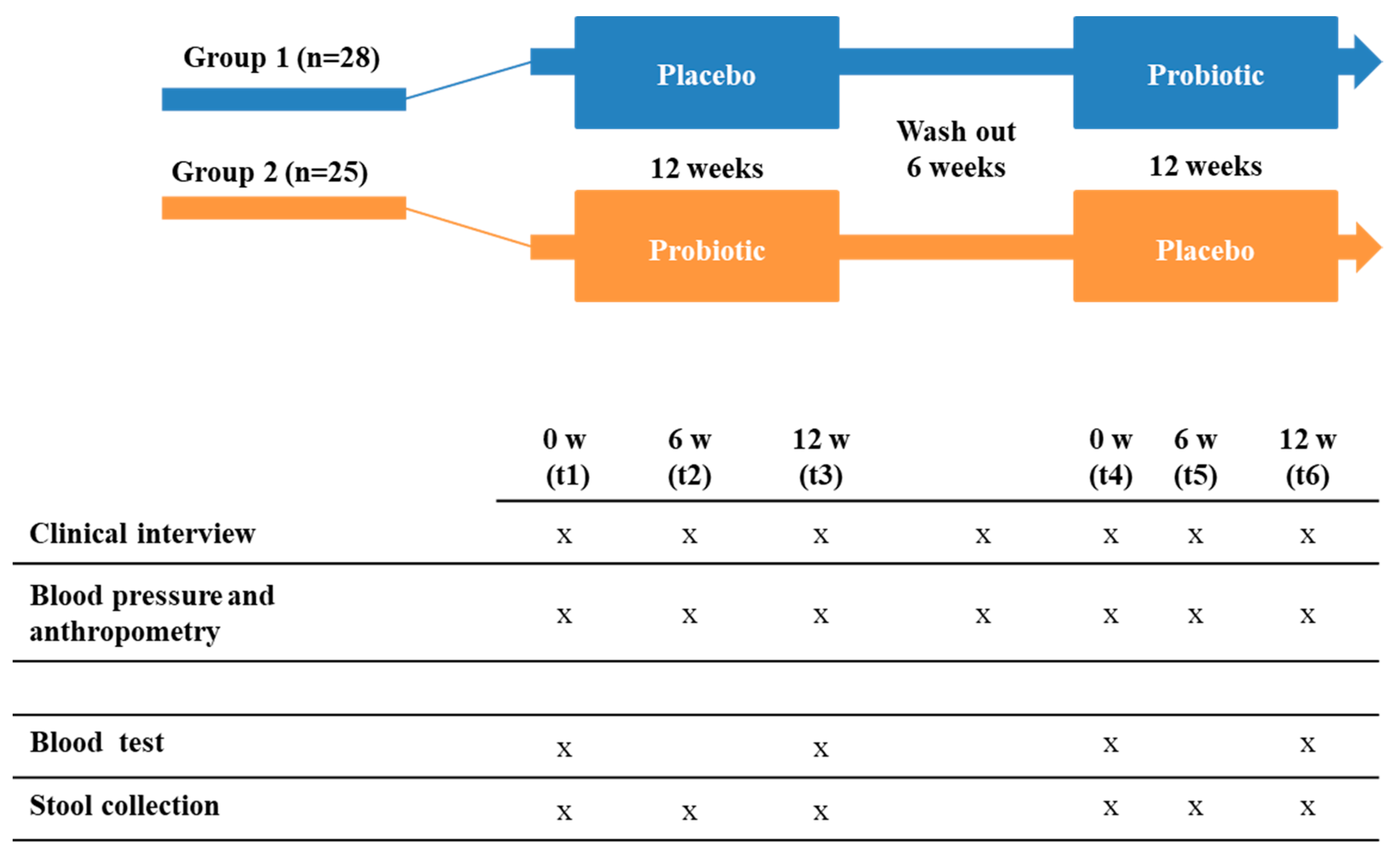
2.2. Subjects and Experimental Design
2.3. Anthropometric, Biochemical, Inflammatory, and Cardiovascular Data
2.4. Fecal Samples, DNA Extraction, and Next-Generation Sequencing
2.5. Taxonomic Analysis
2.6. Statistical Analysis
3. Results
3.1. Anthropometric, Biochemical, and Inflammatory Data
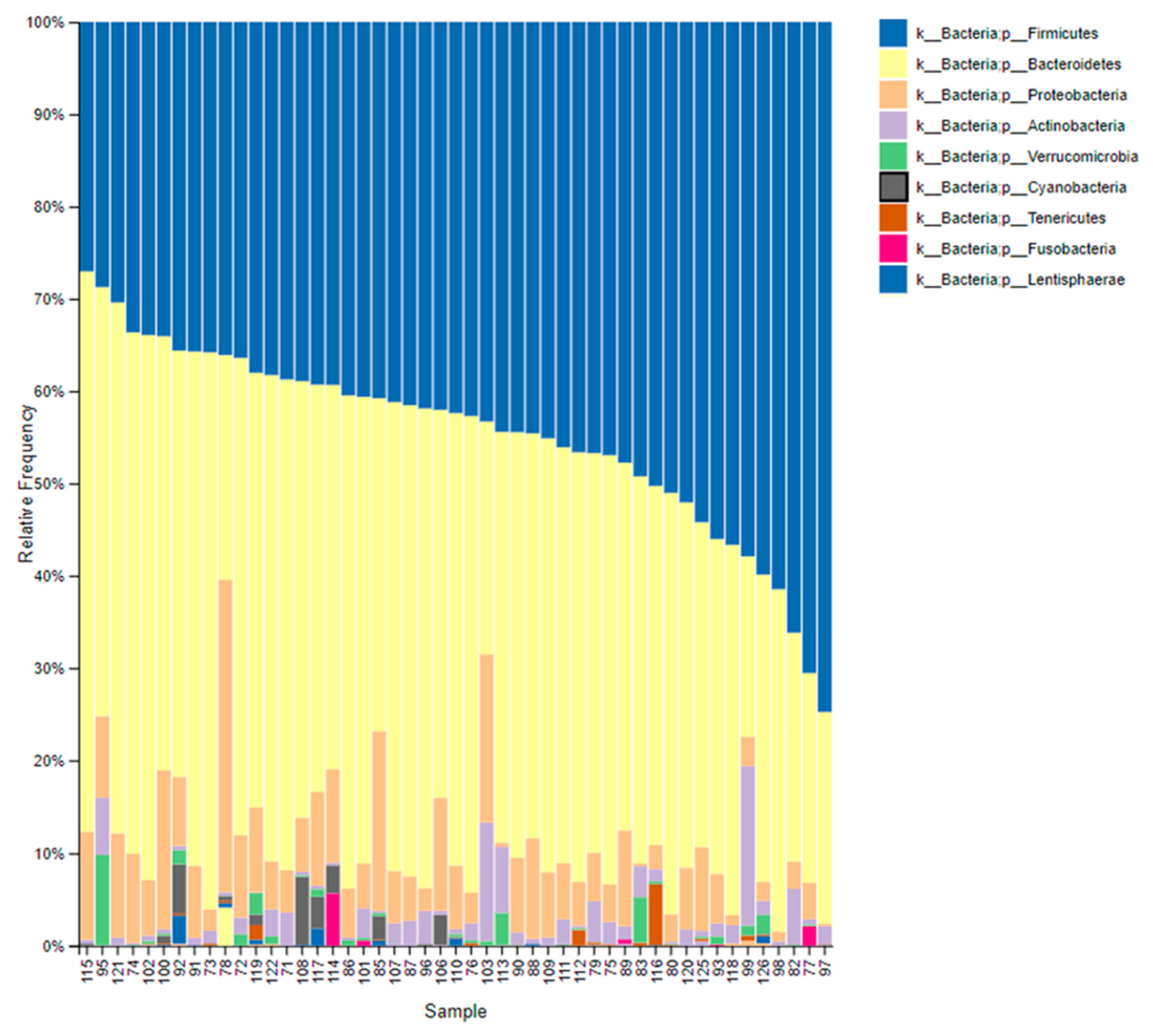
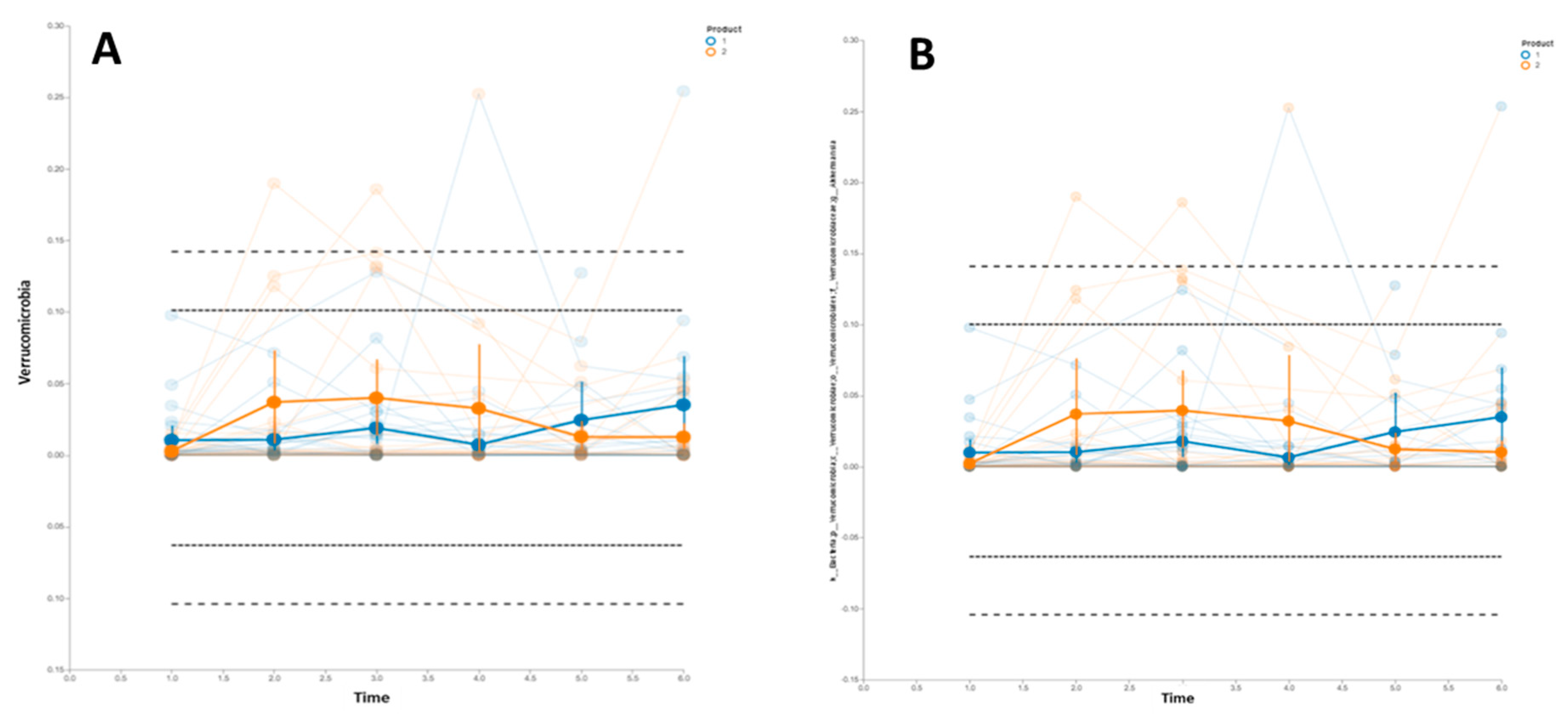
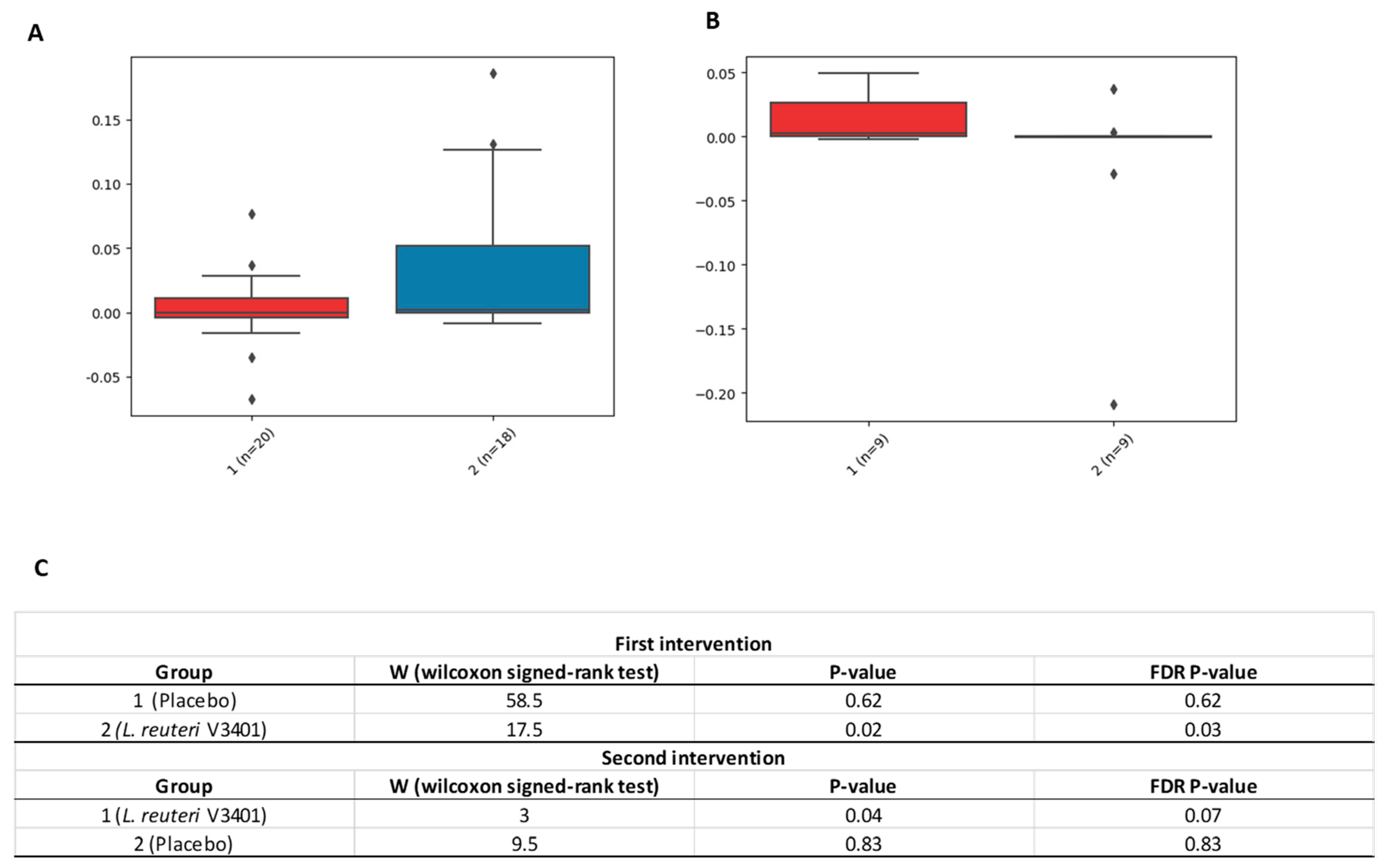
3.2. Gastrointestinal Microbiome Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. Results Probl. Cell Differ. 2017, 960, 1–17. [Google Scholar]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Zupancic, M.L.; Cantarel, B.L.; Liu, Z.; Drabek, E.F.; Ryan, K.A.; Cirimotich, S.; Jones, C.; Knight, R.; Walters, W.A.; Knights, D.; et al. Analysis of the gut microbiota in the older order Amish and its relation to the metabolic syndrome. PLoS ONE 2012, 7, e43052. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; MetaHIT Consortium; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation inhigh-fat diet induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infact. Dis. 2008, 46, S58–S61. [Google Scholar] [CrossRef]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miglioranza Scavuzzi, B.; Miglioranza, L.H.; Henrique, F.C.; Pitelli Paroschi, T.; Lozovoy, M.A.; Simao, A.N.; Dichi, I. The role of probiotics on each component of the metabolic syndrome and other cardiovascular risks. Expert Opin. Ther. Targets 2015, 19, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, Q.; De Haan, B.J.; Zhang, H.; Faas, M.M.; De Vos, P. Identification of TLR2/TLR6 signalling lactic acid bacteria for supporting immune regulation. Sci. Rep. 2016, 6, 34561. [Google Scholar] [CrossRef] [PubMed]
- Bernini, L.J.; Simão, A.N.; Alfieri, D.F.; Lozovoy, M.A.; Mari, N.L.; de Souza, C.H.; Dichi, I.; Costa, G.N. Beneficial effects of Bifidobacterium lactis on lipid profile and cytokines in patients with metabolic syndrome: A randomized trial. Effects of probiotics on metabolic syndrome. Nutrition 2016, 32, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Barreto, F.M.; Simão, A.N.; Morimoto, H.K.; Lozovoy, M.A.; Dichi, I.; Miglioranza, H.D.S. Beneficial effects of Lactobacillus plantarum on glycemia and homocysteine levels in postmenopausal women with metabolic syndrome. Nutrition 2014, 30, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Sañudo Otero, A.I.; Criado García, R.; Rodríguez Nogales, A.; Garach Domech, A.; Olivares Martín, M.; Gálvez Peralta, J.J.; De La Escalera Hueso, S.; Duarte Pérez, J.M.; Zarzuelo Zurita, A.; Bañuelos Hortigüela, O. Probiotic Strains Having Cholesterol Absorbing Capacity, Methods and Uses Thereof. Google Patents No. EP3031930A1, 15 June 2016. [Google Scholar]
- Tenorio-Jiménez, C.; Martínez-Ramírez, M.J.; Tercero-Lozano, M.; Arraiza-Irigoyen, C.; Del Castillo-Codes, I.; Olza, J.; Plaza-Díaz, J.; Fontana, L.; Migueles, J.H.; Olivares, M.; et al. Evaluation of the effect of Lactobacillus reuteri V3401 on biomarkers of inflammation, cardiovascular risk and liver steatosis in obese adults with metabolic syndrome: A randomized clinical trial (PROSIR). BMC Complement. Altern. Med. 2018, 18, 306. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 7, e1. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QUIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. Peer J. Prepr. 2018, 6, e27295v2. [Google Scholar]
- Arteaga, F.; Ferrer, F. Dealing with missing data in MSPC: Several methods, different interpretations, some examples. J. Chemom. 2002, 16, 408–418. [Google Scholar] [CrossRef]
- Wellek, S.; Blettner, M. On the proper use of the crossover design in clinical trials: Part 18 of a series on evaluation of scientific publications. Dtsch. Arztebl. Int. 2012, 109, 276–281. [Google Scholar] [CrossRef]
- Bibby, J.; Kent, J.; Mardia, K. Multivariate Analysis; Academic Press: London, UK, 1979. [Google Scholar]
- Barker, M.; Rayens, W. Partial least squares for discrimination. J. Chemom. 2003, 17, 166–173. [Google Scholar] [CrossRef]
- Smilde, A.K.; Jansen, J.J.; Hoefsloot, H.C.; Lamers, R.J.; Van Der Greef, J.; Timmerman, M.E. ANOVA-simultaneous component analysis (ASCA): A new tool for analyzing designed metabolomics data. Bioinformatics 2005, 21, 3043–3048. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Dillon, M.R.; Zhang, Y.; Rideout, J.R.; Bolyen, E.; Li, H.; Albert, P.S.; Caporaso, J.G. q2-longitudinal: Longitudinal and Paired-Sample Analyses of Microbiome Data. mSystems 2018, 3, e00219-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripolt, N.; Leber, B.; Blattl, D.; Eder, M.; Wonisch, W.; Scharnagl, H.; Stojakovic, T.; Obermayer-Pietsch, B.; Wascher, T.; Pieber, T.; et al. Short communication: Effect of supplementation with Lactobacillus casei Shirota on insulin sensitivity, β-cell function, and markers of endothelial function and inflammation in subjects with metabolic syndrome—A pilot study. J. Dairy Sci. 2013, 96, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, R.; Ravindran, R.; Dhanasekaran, S. Emerging Role of Adipocytokines in Type 2 Diabetes as Mediators of Insulin Resistance and Cardiovascular Disease. Can. J. Diabetes 2018, 42, 446–456.e1. [Google Scholar] [CrossRef] [PubMed]
- Weigert, C.; Hennige, A.M.; Lehmann, R.; Brodbeck, K.; Baumgartner, F.; Schaüble, M.; Häring, H.U.; Schleicher, E.D. Direct Cross-talk of Interleukin-6 and Insulin Signal Transduction via Insulin Receptor Substrate-1 in Skeletal Muscle Cells. J. Biol. Chem. 2006, 281, 7060–7067. [Google Scholar] [CrossRef] [Green Version]
- Glowinska, B.; Urban, M.; Peczynska, J.; Florys, B.; Głowińska-Olszewska, B. Soluble adhesion molecules (sICAM-1, sVCAM-1) and selectins (sE selectin, sP selectin, sL selectin) levels in children and adolescents with obesity, hypertension, and diabetes. Metabolism 2005, 54, 1020–1026. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, I.P.; Zaragoza-García, O.; Vences-Valázquez, A.; Castro-Alarcón, N.; Muñoz-Valle, J.F.; Parra-Rojas, I. Circulating levels of MCP-1, VEGF-A, sICAM-1, sVCAM-1, sE-selectin and sVE-cadherin: Relationship with components of metabolic síndrome in young population. Med. Clin. (Bar) 2016, 147, 427–434. [Google Scholar] [CrossRef]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- de Vos, W.M. Microbe profile: Akkermansia muciniphila: A conserved intestinal symbiont that acts as the gatekeeper of our mucosa. Microbiology 2017, 163, 646–648. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 27, e71108. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, V.; Leber, B.; Lemesch, S.; Trajanoski, S.; Bashir, M.; Horvath, A.; Tawdrous, M.; Stojakovic, T.; Fauler, G.; Fickert, P.; et al. Lactobacillus casei Shirota supplementation does not restore gut microbiota composition and gut barrier in metabolic syndrome: A randomized pilot study. PLoS ONE 2015, 10, e0141399. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Placebo | Probiotic | Probiotic | Placebo | |||||
| t1 | t3 | t4 | t6 | t1 | t3 | t4 | t6 | |
| Weight (kg) | 109.02 ± 26.7 | 105.70 ± 26.2 | 101.50 ± 24.5 | 101.08 ± 24.2 | 103.49 ± 15.2 | 96.56 ± 16.2 | 93.91 ± 16.9 | 92.0 ± 17.3 |
| BMI (kg/m²) | 38.76 ± 7.2 | 37.57 ± 7.1 | 36.77 ± 6.8 | 36.56 ± 6.6 | 38.30 ± 7.3 | 35.69 ± 7.1 | 34.57 ± 6.9 | 33.80 ± 6.6 |
| SBP (mm Hg) | 137.68 ± 16.9 | 133.28 ± 15.4 | 133.11 ± 20.4 | 132.21 ± 14.6 | 139 ± 22.6 | 129.95 ± 16.0 | 131.30 ± 20.0 | 131.40 ± 18.6 |
| DBP (mm Hg) | 84.28 ± 9.6 | 81.96 ± 7.7 | 81.68 ± 11.0 | 82.11 ± 10.5 | 87.95 ± 14.3 | 78.18 ±10.4 | 78.85 ± 12.4 | 81.60 ± 11.2 |
| Glucose (mg/dL) | 103.29 ± 11.0 | 108.08 ± 11.5 | 106.74 ± 8.9 | 105.53 ± 10.5 | 101.0 ± 13.9 | 103.68 ± 13.4 | 101.22 ± 11.8 | 103.78 ± 16.5 |
| Insulin (mU/mL) | 17.50 ± 10.6 | 16.18 ± 11.3 | 22.44 ± 10.3 | 21.74 ± 11.7 | 14.24 ± 8.5 | 12.42 ± 10.9 | 14.04 ± 6.1 | 17.47 ± 7.8 |
| HOMA index | 4.48 ± 2.8 | 4.41 ± 3.3 | 5.91 ± 2.8 | 5.66 ± 3.5 | 3.71 ± 2.7 | 3.52 ± 3.6 | 3.64 ± 1.7 | 4.46 ± 2.2 |
| Glycated Hemoglobin (%) | 5.59 ± 0.4 | 6.04 ± 2.2 | 5.44 ± 0.3 | 5.49 ± 0.3 | 5.68 ± 0.4 | 5.90 ± 1.9 | 5.46 ± 0.3 | 5.44 ± 0.3 |
| Total cholesterol (mg/dL) | 232.42 ± 43.0 | 207.08 ± 36.0 | 202.79 ± 45.8 | 224.16 ± 45.5 | 233.41 ± 46.5 | 203.64 ± 37.9 | 209.56 ± 58.0 | 220.89 ± 53.8 |
| Triacylglycerols (mg/dL) | 119.25 ± 47.6 | 122.46 ± 59.9 | 109.00 ± 47.3 | 118.89 ± 52.2 | 130.55 ± 47.5 | 128.23 ± 57.6 | 112.89 ± 42.7 | 100.56 ± 62.6 |
| LDL (mg/dL) | 156.79 ± 35.7 | 131.71 ± 30.0 | 128.11 ± 32.5 | 144.42 ± 39.0 | 161.0 ± 41.6 | 132.91 ± 32.1 | 136.22 ± 47.7 | 145.72 ± 44.5 |
| HDL (mg/dL) | 50.54 ± 14.6 | 50.46 ± 12.2 | 52.47 ± 13.3 | 54.11 ± 10.2 | 45.95 ± 9.5 | 44.68 ± 7.9 | 50.44 ± 10.3 | 54.61 ± 11.5 |
| GOT (U/L) | 25.75 ± 7.7 | 22.83 ± 6.3 | 22.63 ± 6.8 | 22.32 ± 5.7 | 23.55 ± 12.4 | 25.50 ± 14.9 | 20.56 ± 6.1 | 21.33 ± 6.2 |
| GPT (U/L) | 34.79 ± 17.4 | 28.17 ± 14.3 | 22.42 ± 11.9 | 27.89 ± 12.7 | 24.38 ± 8.9 | 22.24 ± 8.4 | 18.00 ± 7.5 | 22.50 ± 11.0 |
| γ-GT (U/L) | 36.29 ± 13.6 | 37.08 ± 16.7 | 38.42 ± 18.9 | 36.74 ± 21.2 | 26.05 ± 12.7 | 24.91 ± 14.2 | 24.89 ± 14.3 | 24.00 ± 15.4 |
| Group 1 | Group 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Placebo | Probiotic | Probiotic | Placebo | |||||
| t1 | t2 | t3 | t5 | t1 | t2 | t3 | t5 | |
| CRP (mg/dL) | 5.13 ± 3.7 | 5.78 ± 4.8 | 6.97 ± 7.4 | 5.52 ± 4.4 | 5.88 ± 4.4 | 6.12 ± 6.3 | 3.66 ± 2.7 | 4.24 ± 3.8 |
| IL-6 (pg/mL) | 2.91 ± 1.8 | 3.33 ± 2.4 | 3.12 ± 1.9 | 2.62 ± 2.0 | 2.07 ± 1.2 | 1.79 ± 1.2 | 1.37 ± 0.8 | 1.72 ± 1.0 |
| IL-8 (pg/mL) | 2.86 ± 1.7 | 2.80 ± 1.2 | 4.11 ± 7.1 | 4.23 ± 9.4 | 2.66 ± 1.1 | 2.73 ± 1.2 | 2.27 ± 1.0 | 2.28 ± 1.1 |
| TNF-α (pg/mL) | 4.70 ± 2.5 | 4.91 ± 2.6 | 4.59 ± 2.1 | 3.51 ± 1.7 | 4.05 ± 1.9 | 4.15 ± 2.1 | 3.05 ± 1.2 | 3.28 ± 2.0 |
| Adiponectin (mg/L) | 6.55 ± 5.1 | 5.95 ± 4.7 | 5.82 ± 3.8 | 6.56 ± 3.4 | 5.69 ± 3.7 | 6.91 ± 6.5 | 7.20 ± 4.4 | 8.26 ± 6.1 |
| tPAI1 (µg/L) | 9.24 ± 4.9 | 9.55 ± 4.3 | 10.55 ± 4.8 | 11.56 ± 7.1 | 9.31 ± 3.2 | 8.36 ± 2.6 | 9.08 ± 3.2 | 9.51 ± 3.6 |
| P-selectin (ng/mL) | 46.78 ± 19.7 | 46.70 ± 21.2 | 49.26 ± 22.8 | 63.47 ± 38.3 | 48.06 ± 16.7 | 40.51 ± 11.4 | 60.73 ± 35.4 | 58.09 ± 21.6 |
| Resistin (µg/L) | 17.71 ± 8.1 | 17.70 ± 13.7 | 16.99 ± 5.2 | 17.45 ± 10.4 | 15.33 ± 7.2 | 11.89 ± 5.2 | 11.08 ± 4.0 | 12.60 ± 5.2 |
| HGF (pg/mL) | 161.12 ± 97.9 | 155.73 ± 93.6 | 131.45 ± 65.9 | 162.45 ± 88.3 | 175.06 ± 75.1 | 170.17 ± 72.7 | 160.47 ± 68.8 | 157.95 ± 57.5 |
| Leptin (µg/L) | 28.56 ± 14.8 | 24.07 ± 12.3 | 23.71 ± 13.8 | 18.42 ± 10.5 | 21.97 ± 11.8 | 17.68 ± 10.9 | 13.67 ± 9.3 | 14.24 ± 10.4 |
| MCP-1 (pg/mL) | 107.53 ± 39.3 | 106.57 ± 31.7 | 120.07 ± 60.1 | 118.20 ± 46.8 | 108.86 ± 39.3 | 114.61 ± 51.5 | 116.31 ± 43.6 | 112.51 ± 41.2 |
| sICAM (ng/mL) | 73.65 ± 37.2 | 73.71 ± 41.1 | 67.86 ± 35.2 | 65.47 ± 32.2 | 74.80 ± 26.9 | 71.50 ± 33.1 | 75.17 ± 40.2 | 73.0 ± 41.2 |
| MPO (ng/mL) | 17.69 ± 5.9 | 19.96 ± 10.7 | 20.70 ± 13.2 | 30.53 ± 21.2 | 15.56 ± 7.8 | 18.14 ± 13.0 | 17.31 ± 12.4 | 19.46 ± 10.2 |
| sVCAM (ng/mL) | 494.22 ± 125.1 | 511.04 ± 154.8 | 516.47 ± 149.1 | 507.61 ± 138.7 | 489.68 ± 80.9 | 472.72 ± 71.4 | 491.62 ± 99.6 | 527.43 ± 74.6 |
| LPS (ng/mL) | 285.81 ± 107.2 | 277.90 ± 116.3 | 312.22 ± 126.0 | 326.19 ± 166.0 | 321.82 ± 105.4 | 316.83 ± 124.0 | 309.91 ± 136.7 | 308.70 ± 131.4 |
| LBP (ng/mL) | 731.26 ± 512.2 | 782.55 ± 323.0 | 635.96 ± 294.2 | 747.39 ± 272.6 | 837.47 ± 423.5 | 742.81 ± 349.2 | 833.63 ± 560.5 | 855.78 ± 663.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenorio-Jiménez, C.; Martínez-Ramírez, M.J.; Del Castillo-Codes, I.; Arraiza-Irigoyen, C.; Tercero-Lozano, M.; Camacho, J.; Chueca, N.; García, F.; Olza, J.; Plaza-Díaz, J.; et al. Lactobacillus reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: The PROSIR Study. Nutrients 2019, 11, 1761. https://doi.org/10.3390/nu11081761
Tenorio-Jiménez C, Martínez-Ramírez MJ, Del Castillo-Codes I, Arraiza-Irigoyen C, Tercero-Lozano M, Camacho J, Chueca N, García F, Olza J, Plaza-Díaz J, et al. Lactobacillus reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: The PROSIR Study. Nutrients. 2019; 11(8):1761. https://doi.org/10.3390/nu11081761
Chicago/Turabian StyleTenorio-Jiménez, Carmen, María José Martínez-Ramírez, Isabel Del Castillo-Codes, Carmen Arraiza-Irigoyen, Mercedes Tercero-Lozano, José Camacho, Natalia Chueca, Federico García, Josune Olza, Julio Plaza-Díaz, and et al. 2019. "Lactobacillus reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: The PROSIR Study" Nutrients 11, no. 8: 1761. https://doi.org/10.3390/nu11081761