
β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Setting and Study Design
2.3. Zebrafish Maintenance and Embryos Production
2.4. Treatments
2.5. Dechorionation Procedure
2.6. Sonication Procedure
2.7. Birefringence Test
2.8. DanioVision
2.9. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.10. Mitochondrial Respiratory Analysis
2.11. Statistical Analysis
3. Results
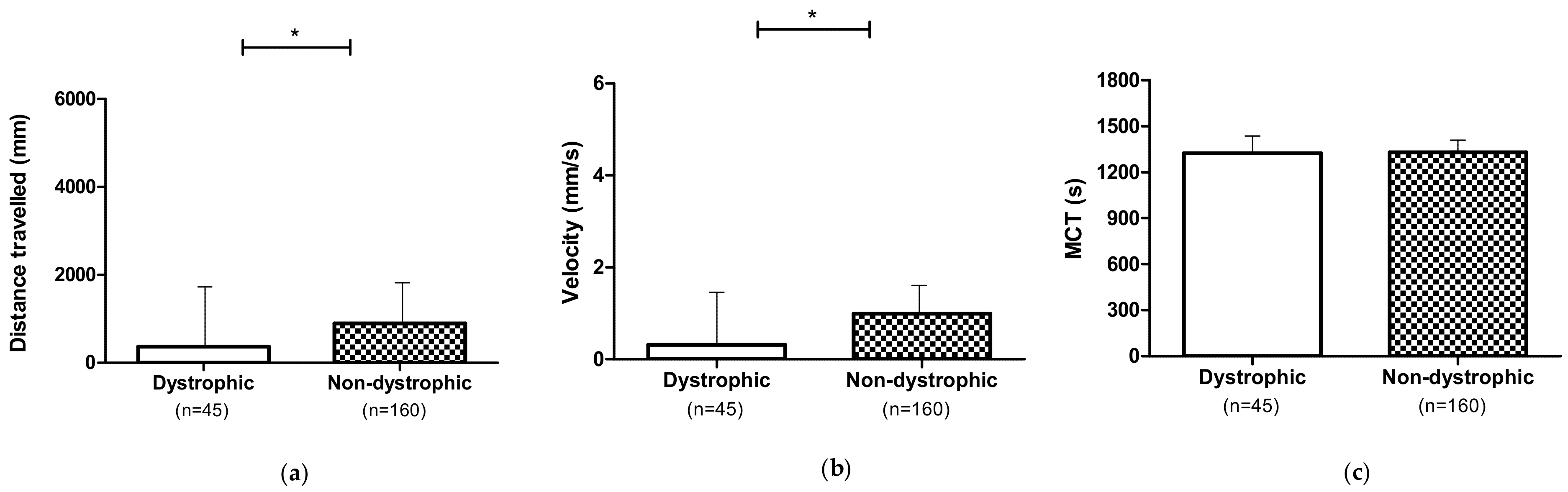
3.1. Locomotor Performances of the Dystrophic and Non-Dystrophic Phenotypes
3.2. Evaluation of the Best Method for Administration of 1,3-1,6 β-Glucans
3.2.1. Effect of Embryo Dechorionation (D)
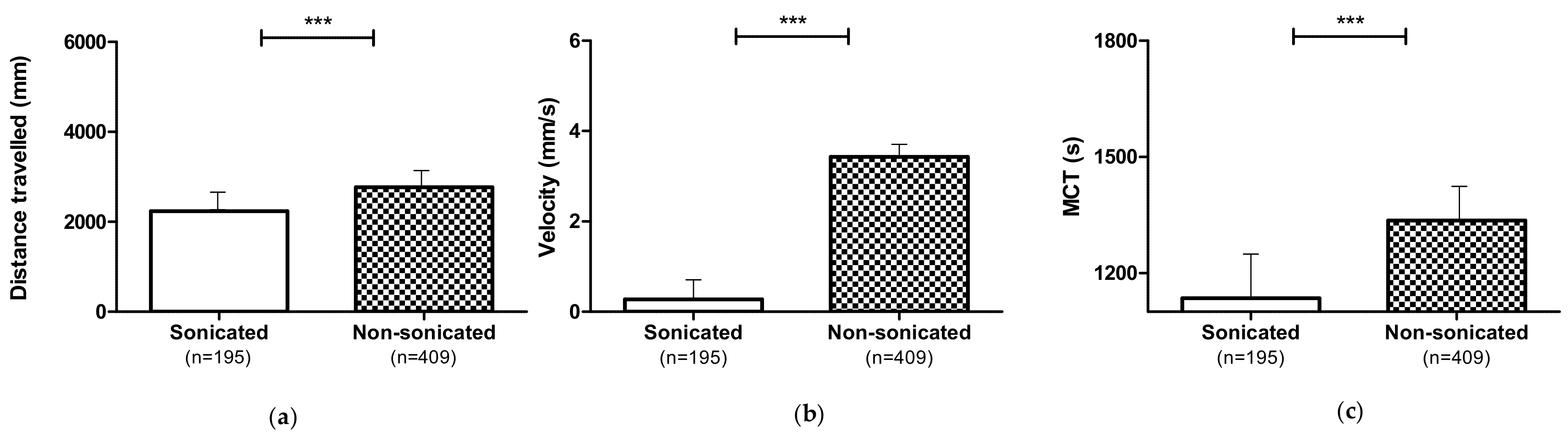
3.2.2. Effect of 1,3-1,6 β-Glucans Sonication (S)
3.2.3. Combined Effect of Embryo Dechorionation and Sonication
3.3. Effect of 1,3-1,6 β-Glucans Concentration on the Relative Incidence of Muscle Phenotypes (Dystrophic and Non-Dystrophic)
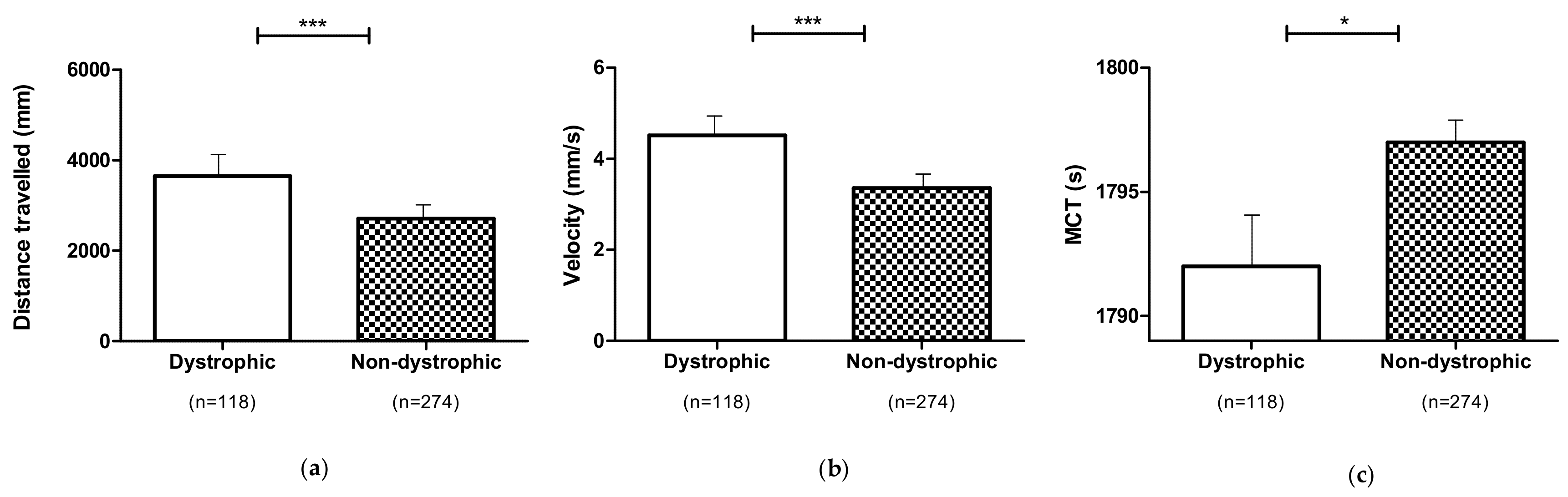
3.4. Locomotor Performances of Dystrophic and Non-Dystrophic Phenotypes (DNS and NDS Procedures)
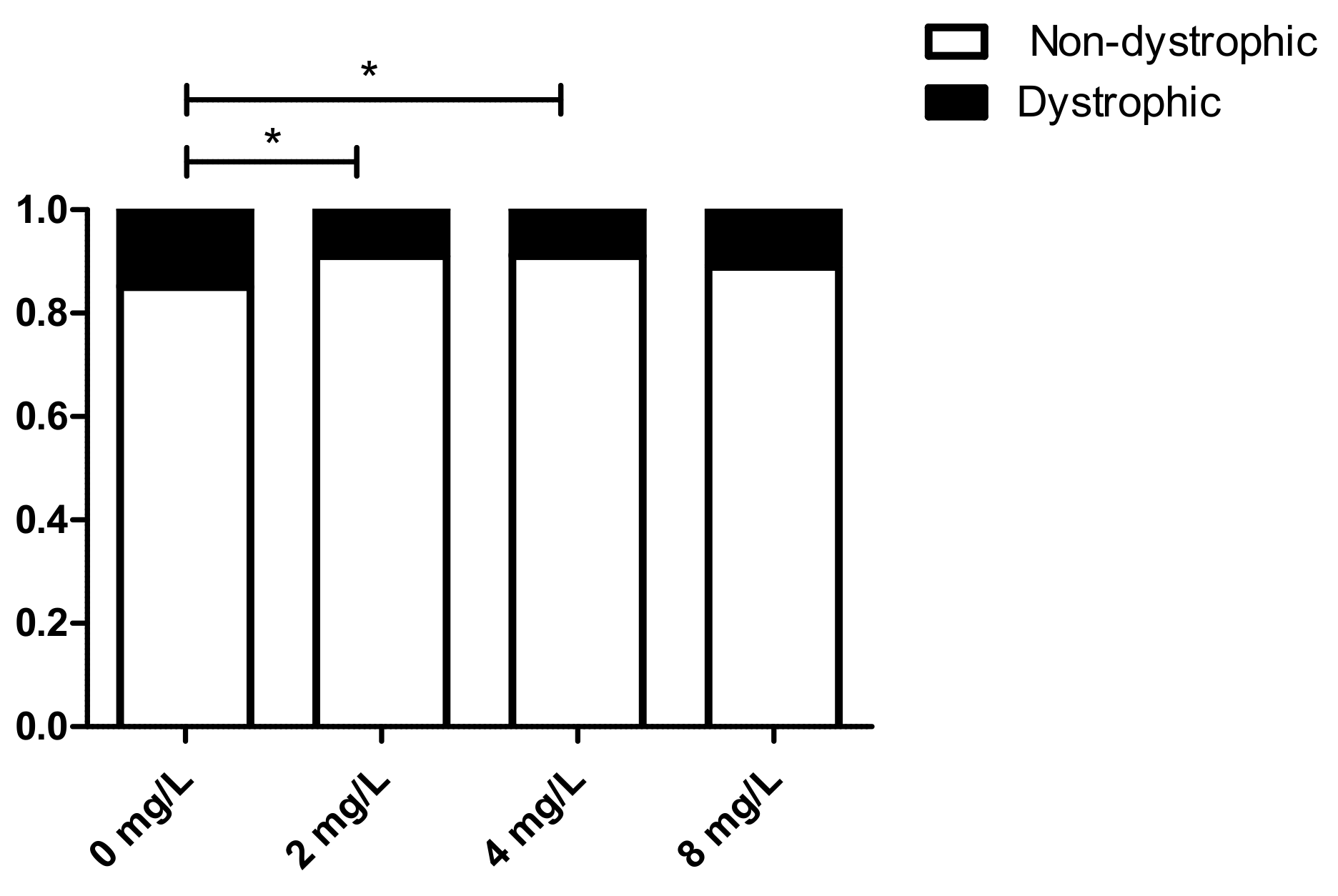
3.5. Effect of 1,3-1,6 β-Glucans Concentration on the Relative Incidence of Dystrophic and Non-Dystrophic Phenotypes (DNS and NDS Procedures)
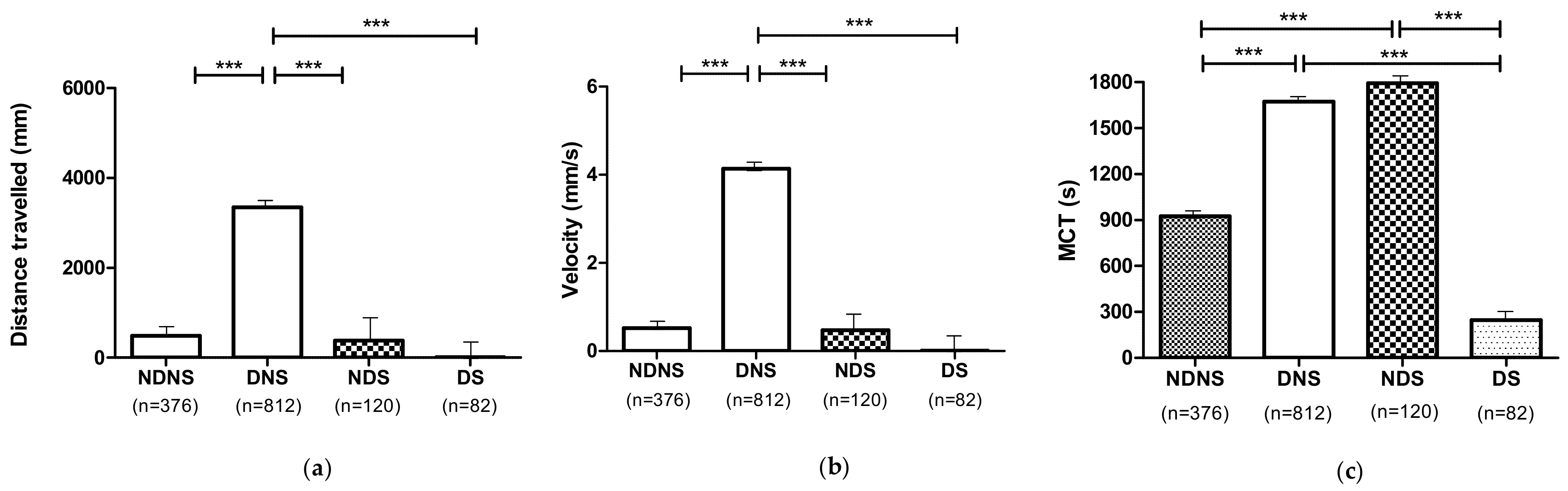
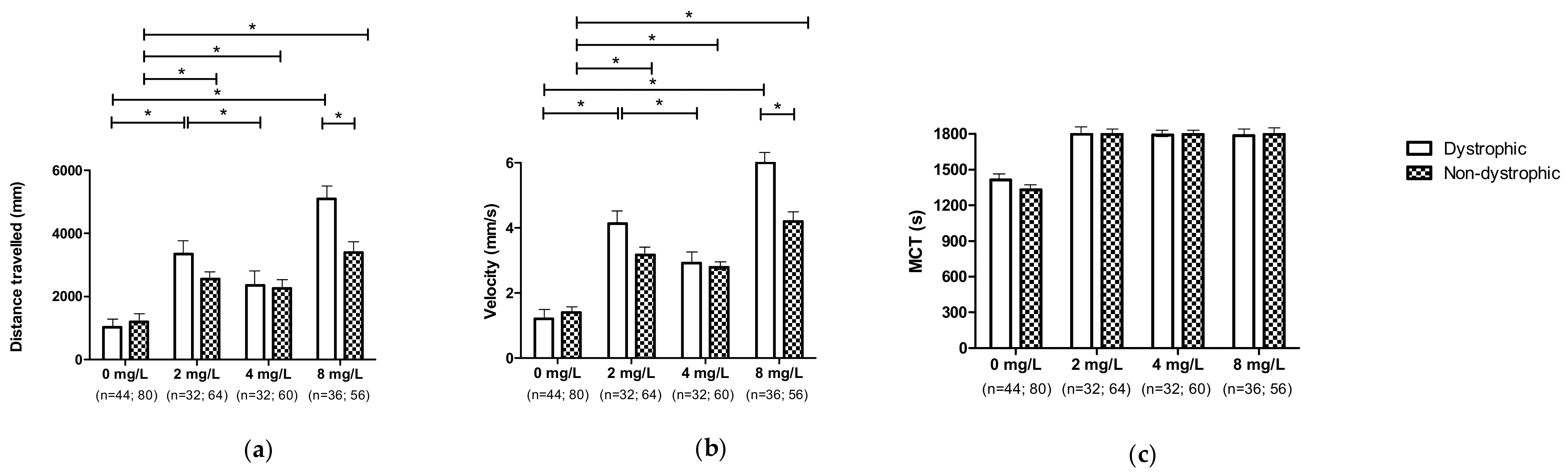
3.6. Effect of 1,3-1,6 β-Glucans Concentration on Locomotor Performances (DNS and NDS Procedures)
3.6.1. Effect of 1,3-1,6 β-Glucans Concentration on Locomotor Performances Independent of the Individual Phenotypes
3.6.2. Effect of 1,3-1,6 β-Glucans Concentration on Locomotor Performances According to the Different Phenotypes
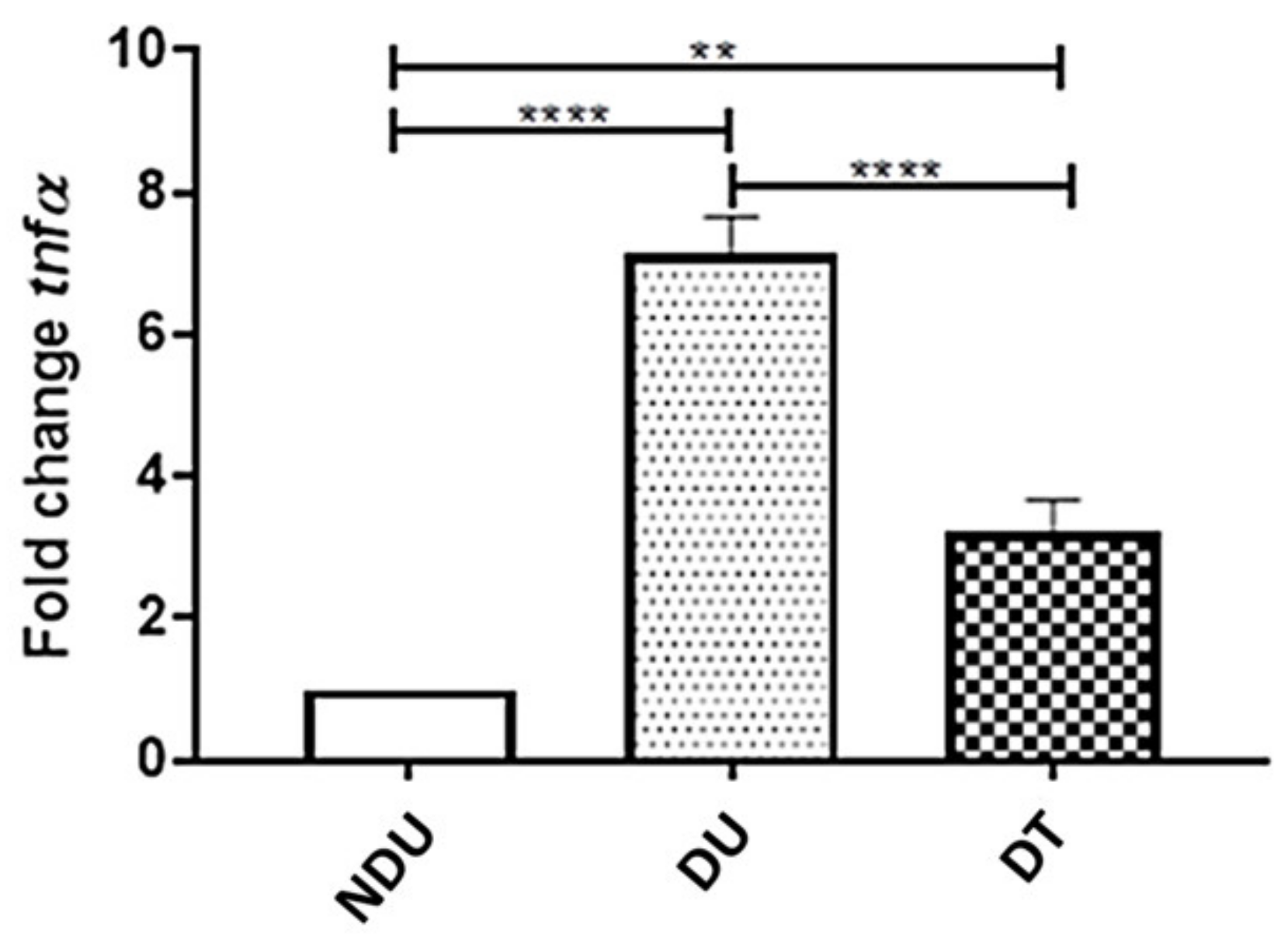
3.7. Effect of 1,3-1,6 β-Glucans on the Inflammatory Cytokine Tumor Necrosis Factor α (TNF α)
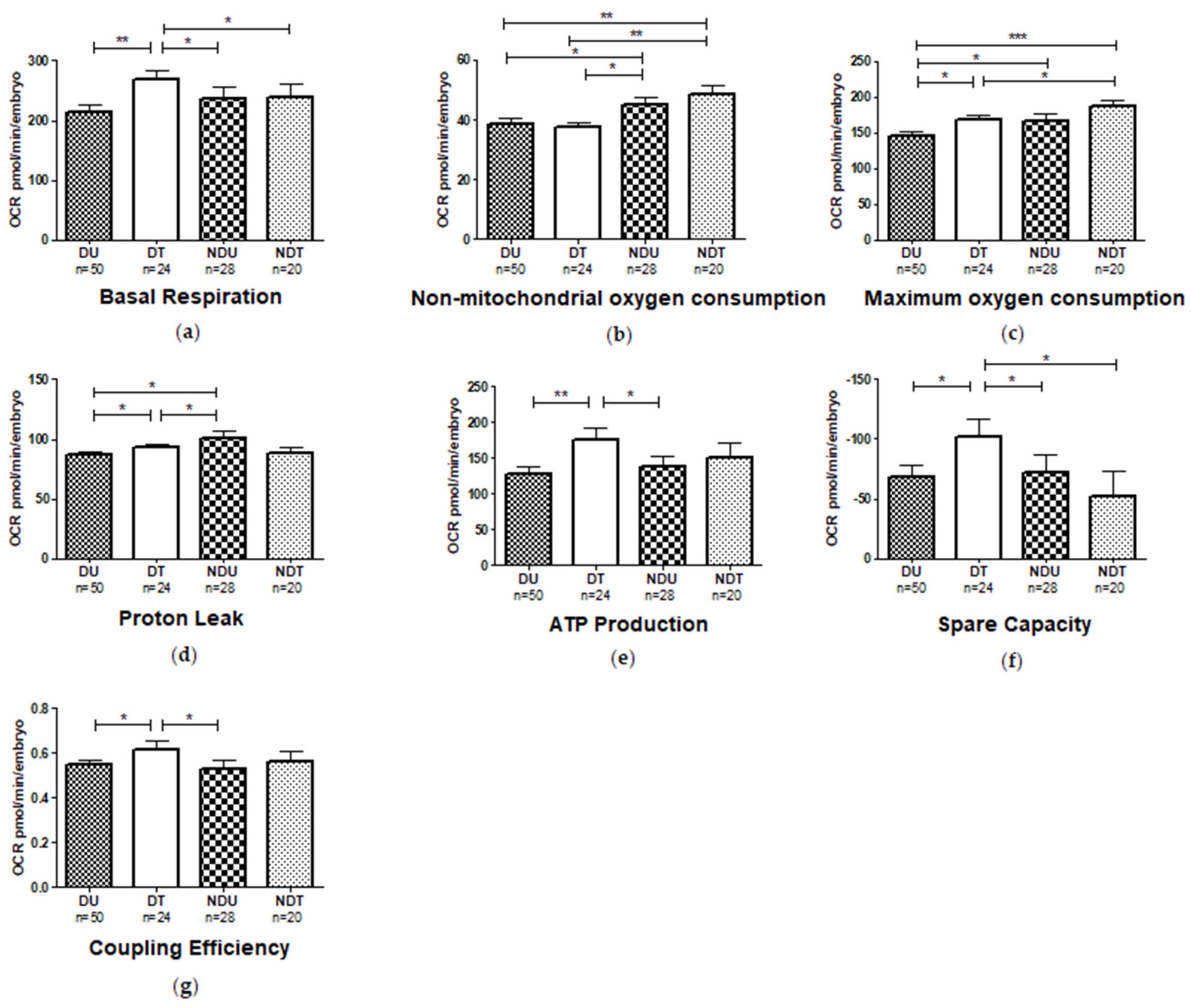
3.8. Mitochondrial Respiratory Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allsop, K.G.; Ziter, F.A. Loss of Strength and Functional Decline in Duchenne’s Dystrophy. Arch. Neurol. 1981, 38, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Dubowitz, V. Enigmatic conflict of clinical and molecular diagnosis in Duchenne/Becker muscular dystrophy. Neuromuscul. Disord. 2006, 16, 865–866. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.G.; Franzini-Armstrong, C. (Eds.) Dystrophinopathies, 2nd ed.; McGraw-Hill: New York, NY, USA, 1994; Volume 2. [Google Scholar]
- De Carvalho, E.V.; Hukuda, M.E.; Escorcio, R.; Voos, M.C.; Caromano, F.A. Development and Reliability of the Functional Evaluation Scale for Duchenne Muscular Dystrophy, Gait Domain: A Pilot Study. Physiother. Res. Int. 2015, 20, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Martini, J.; Voos, M.C.; Hukuda, M.E.; de Resende, M.B.D.; Caromano, F.A. Compensatory movements during functional activities in ambulatory children with Duchenne muscular dystrophy. Arq. Neuropsiquiatr. 2014, 72, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, E.M.; Davidson, A.E.; Trickey-Glassman, A.; Backus, C.; Hong, Y.; Sakowski, S.A.; Dowling, J.J.; Feldman, E.L. Two Dynamin-2 Genes Are Required for Normal Zebrafish Development. PLoS ONE 2013, 8, E55888. [Google Scholar] [CrossRef] [Green Version]
- Gloss, D.; Moxley, R.T.; Ashwal, S.; Oskoui, M. Practice guideline update summary: Corticosteroid treatment of Duchenne muscular dystrophy. Neurology 2016, 86, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Matthews, E.; Brassington, R.; Kuntzer, T.; Jichi, F.; Manzur, A.Y. Corticosteroids for the treatment of Duchenne muscular dystrophy. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef] [Green Version]
- Babbs, A.; Chatzopoulou, M.; Edwards, B.; Squire, S.E.; Wilkinson, I.V.L.; Wynne, G.M.; Russell, A.J.; Davies, K.E. From diagnosis to therapy in Duchenne muscular dystrophy. Biochem. Soc. Trans. 2020, 48, 813–821. [Google Scholar] [CrossRef]
- Kole, R.; Krieg, A.M. Exon skipping therapy for Duchenne muscular dystrophy. Adv. Drug Deliv. Rev. 2015, 87, 104–107. [Google Scholar] [CrossRef]
- Bachrach, E.; Li, S.; Perez, A.L.; Schienda, J.; Liadaki, K.; Volinski, J.; Flint, A.; Chamberlain, J.; Kunkel, L.M. Systemic delivery of human microdystrophin to regenerating mouse dystrophic muscle by muscle progenitor cells. Proc. Natl. Acad. Sci. USA 2004, 101, 3581–3586. [Google Scholar] [CrossRef] [Green Version]
- Montanaro, F.; Liadaki, K.; Schienda, K.; Flint, A.; Gussoni, E.; Kunkel, L.M. Demystifying SP cell purification: Viability, yield, and phenotype are defined by isolation parameters. Exp. Cell Res. 2004, 298, 144–154. [Google Scholar] [CrossRef]
- Kourakis, S.; Timpani, C.A.; Campelj, D.G.; Hafner, P.; Gueven, N.; Fischer, D.; Rybalka, E. Standard of care versus new-wave corticosteroids in the treatment of Duchenne muscular dystrophy: Can we do better? Orphanet J. Rare Dis. 2021, 16, 117. [Google Scholar] [CrossRef]
- Davis, J.; Samuels, E.; Mullins, L. Nutrition Considerations in Duchenne Muscular Dystrophy. Nutr. Clin. Pract. 2015, 30, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Messina, S.; Bitto, A.; Aguennouz, M.; Vita, G.L.; Polito, F.; Irrera, N.; Altavilla, D.; Marini, H.; Migliorato, A.; Squadrito, F.; et al. The soy isoflavone genistein blunts nuclear factor kappa-B, MAPKs and TNF-α activation and ameliorates muscle function and morphology in mdx mice. Neuromuscul. Disord. 2011, 21, 579–589. [Google Scholar] [CrossRef]
- De Palma, C.; Morisi, F.; Cheli, S.; Pambianco, S.; Cappello, V.; Vezzoli, M.; Rovere-Querini, P.; Moggio, M.; Ripolone, M.; Francolini, M.; et al. Autophagy as a new therapeutic target in Duchenne muscular dystrophy. Cell Death Dis. 2012, 3, e418. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.C.; Ramos, S.V.; Turnbull, P.C.; Rebalka, I.A.; Cao, A.; Monaco, C.M.F.; Varah, N.E.; Edgett, B.A.; Huber, J.S.; Tadi, P.; et al. Early myopathy in Duchenne muscular dystrophy is associated with elevated mitochondrial H2O2 emission during impaired oxidative phosphorylation. J. Cachexia Sarcopenia Muscle 2019, 10, 643–661. [Google Scholar] [CrossRef] [Green Version]
- Rybalka, E.; Timpani, C.A.; Cooke, M.B.; Williams, A.D.; Hayes, A. Defects in Mitochondrial ATP Synthesis in Dystrophin-Deficient Mdx Skeletal Muscles May Be Caused by Complex I Insufficiency. PLoS ONE 2014, 9, e115766. [Google Scholar] [CrossRef] [Green Version]
- Woodman, K.; Coles, C.; Lamandé, S.; White, J. Nutraceuticals and Their Potential to Treat Duchenne Muscular Dystrophy: Separating the Credible from the Conjecture. Nutrients 2016, 8, 713. [Google Scholar] [CrossRef]
- Sebori, R.; Kuno, A.; Hosoda, R.; Hayashi, T.; Horio, Y. Resveratrol Decreases Oxidative Stress by Restoring Mitophagy and Improves the Pathophysiology of Dystrophin-Deficient mdx Mice. Oxid. Med. Cell. Longev. 2018, 2018, 9179270. [Google Scholar] [CrossRef] [Green Version]
- Licitra, R.; Marchese, M.; Brogi, L.; Fronte, B.; Pitto, L.; Santorelli, F.M. Nutraceutical Screening in a Zebrafish Model of Muscular Dystrophy: Gingerol as a Possible Food Aid. Nutrients 2021, 13, 998. [Google Scholar] [CrossRef]
- Tzianabos, A.O. Polysaccharide Immunomodulators as Therapeutic Agents: Structural Aspects and Biologic Function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef]
- Cuervo, A.M.; Macian, F. Autophagy, nutrition and immunology. Mol. Asp. Med. 2012, 33, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish Physiol. Biochem. 2013, 39, 431–457. [Google Scholar] [CrossRef]
- Novak, M.; Vetvicka, V. β-Glucans, History, and the Present: Immunomodulatory Aspects and Mechanisms of Action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef]
- C Ooi, V.E.; Liu, F. Immunomodulation and Anti-Cancer Activity of Polysaccharide-Protein Complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Fronte, B.; Kim, C.; Bagliacca, M.; Casini, L.; De Zoysa, M. 1,3-1-6β-glucans enhance tissue regeneration in zebrafish (Danio rerio): Potential advantages for aquaculture applications. Aquac. Res. 2019, 50, 3163–3170. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Sonck, E.; Stuyven, E.; Goddeeris, B.; Cox, E. The effect of β-glucans on porcine leukocytes. Vet. Immunol. Immunopathol. 2010, 135, 199–207. [Google Scholar] [CrossRef]
- Vetvicka, V. Glucan-immunostimulant, adjuvant, potential drug. World J. Clin. Oncol. 2011, 2, 115–119. [Google Scholar] [CrossRef]
- Vetvicka, V.; Thornton, B.P.; Ross, G.D. Soluble beta-glucan polysaccharide binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of iC3b-opsonized target cells. J. Clin. Investig. 1996, 98, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Barsanti, L.; Passarelli, V.; Evangelista, V.; Frassanito, A.M.; Gualtieri, P. Chemistry, physico-chemistry and applications linked to biological activities of β-glucans. Nat. Prod. Rep. 2011, 28, 457–466. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Damiano, S.; Montanino, C.; Del Buono, A.; La Rosa, G.; Guida, B.; Santillo, M. Effect of beta- and alpha-glucans on immune modulating factors expression in enterocyte-like Caco-2 and goblet-like LS 174T cells. Int. J. Biol. Macromol. 2020, 153, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G.; Wehling-Henricks, M. Evolving Therapeutic Strategies for Duchenne Muscular Dystrophy: Targeting Downstream Events. Pediatr. Res. 2004, 56, 831–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Guzmán, O.D.; Rodríguez-Cruz, M.; Escobar Cedillo, R.E. Systemic Inflammation in Duchenne Muscular Dystrophy: Association with Muscle Function and Nutritional Status. BioMed Res. Int. 2015, 2015, 891972. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Underhill, D.M. β-glucan signaling connects phagocytosis to autophagy. Glycobiology 2013, 23, 1047–1051. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, A.; Drywień, E.M.; Hamulka, J.; Sadkowski, T. Nutraceutical functions of beta-glucans in human nutrition. Rocz. Panstw. Zakładu Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- Bayrak, O.; Turgut, F.; Karatas, O.F.; Cimentepe, E.; Bayrak, R.; Catal, F.; Atis, O.; Akcay, A.; Unal, D. Oral β-Glucan Protects Kidney against Ischemia/Reperfusion Injury in Rats. Am. J. Nephrol. 2008, 28, 190–196. [Google Scholar] [CrossRef]
- Şener, G.; Toklu, H.; Ercan, F.; Erkanlı, G. Protective effect of β-glucan against oxidative organ injury in a rat model of sepsis. Int. Immunopharmacol. 2005, 5, 1387–1396. [Google Scholar] [CrossRef]
- Silva, D.O.; Lobato, R.V.; Andrade, E.; Orlando, D.; Borges, B.; Zangeronimo, M.; de Sousa, R.; Pereira, L. Effects of β-Glucans Ingestion on Alveolar Bone Loss, Intestinal Morphology, Systemic Inflammatory Profile, and Pancreatic β-Cell Function in Rats with Periodontitis and Diabetes. Nutrients 2017, 9, 1016. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Mo, X.; Huang, H.; Chen, X.; Liu, H.; Peng, Z.; Chen, L.; Rong, S.; Yang, W.; Xu, S.; et al. Yeast β-glucan alleviates cognitive deficit by regulating gut microbiota and metabolites in Aβ1–42-induced AD-like mice. Int. J. Biol. Macromol. 2020, 161, 258–270. [Google Scholar] [CrossRef]
- Geller, A.; Shrestha, R.; Yan, J. Yeast-Derived β-glucan in cancer: Novel uses of a traditional therapeutic. Int. J. Mol. Sci. 2019, 20, 3618. [Google Scholar] [CrossRef] [Green Version]
- Shaki, F.; Pourahmed, J. Mitochondrial Toxicity of Depleted Uranium: Protection by Beta-Glucan. Iran. J. Pharm. Res. 2013, 12, 131–140. [Google Scholar]
- Babincová, M.; Bačová, Z.; Machová, E.; Kogan, G. Antioxidant Properties of Carboxymethyl Glucan: Comparative Analysis. J. Med. Food 2002, 5, 1084–1088. [Google Scholar] [CrossRef]
- Babincová, M.; Machová, E.; Kogan, G. Carboxymethylated Glucan Inhibits Lipid Peroxidation in Liposomes. Z. Naturforsch. C 1999, 54, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Kofuji, K.; Aoki, A.; Tsubaki, K.; Konishi, M.; Isobe, T.; Murata, Y. Antioxidant Activity of β-Glucan. ISRN Pharm. 2012, 2012, 125864. [Google Scholar] [CrossRef] [Green Version]
- Schronerová, K.; Babincová, M.; Machová, E.; Kogan, G. Carboxymethylated (1–> 3)-β-D-Glucan Protects Liposomes Against Ultraviolet Light-Induced Lipid Peroxidation. J. Med. Food 2007, 10, 189–193. [Google Scholar] [CrossRef]
- Bassett, D.I. Dystrophin is required for the formation of stable muscle attachments in the zebrafish embryo. Development 2003, 130, 5852–5860. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, G.; Karpf, J.A.; Myers, J.A.; Alexander, M.S.; Guyon, J.R.; Kunkel, L.M. Drug screening in a zebrafish model of Duchenne muscular dystrophy. Proc. Natl. Acad. Sci. USA 2011, 108, 5331–5336. [Google Scholar] [CrossRef] [Green Version]
- Lipscomb, L.; Piggott, R.W.; Emmerson, T.; Winder, S.J. Dasatinib as a treatment for Duchenne muscular dystrophy. Hum. Mol. Genet. 2016, 25, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Vieira, N.M.; Spinazzola, J.M.; Alexander, M.S.; Moreira, Y.B.; Kawahara, G.; Gibbs, D.E.; Mead, L.C.; Verjovski-Almeida, S.; Zatz, M.; Kunkel, L.M. Repression of phosphatidylinositol transfer protein α ameliorates the pathology of Duchenne muscular dystrophy. Proc. Natl. Acad. Sci. USA 2017, 114, 6081–6085. [Google Scholar] [CrossRef] [Green Version]
- Schiavone, M.; Zulian, A.; Menazza, S.; Petronilli, V.; Argenton, F.; Merlini, L.; Sabatelli, P.; Bernardi, P. Alisporivir rescues defective mitochondrial respiration in Duchenne muscular dystrophy. Pharmacol. Res. 2017, 125, 122–131. [Google Scholar] [CrossRef]
- Guyon, J.R.; Steffen, L.S.; Howell, M.H.; Pusack, T.J.; Lawrence, C.; Kunkel, L.M. Modeling human muscle disease in zebrafish. Biochim. Biophys. Acta Mol. Basis Dis. 2007, 1772, 205–215. [Google Scholar] [CrossRef]
- Henn, K.; Braunbeck, T. Dechorionation as a tool to improve the fish embryo toxicity test (FET) with the zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 153, 91–98. [Google Scholar] [CrossRef]
- Hunter, K.W.; Gault, R.A.; Berner, M.D. Preparation of microparticulate beta-glucan from Saccharomyces cerevisiae for use in immune potentiation. Lett. Appl. Microbiol. 2002, 35, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Gordon, S. Immune recognition of fungal β-glucans. Cell. Microbiol. 2005, 7, 471–479. [Google Scholar] [CrossRef]
- Li, S.; Sauer, W.C.; Huang, S.X.; Gabert, V.M. Effect of beta-glucanase supplementation to hulless barley- or wheat-soybean meal diets on the digestibilities of energy, protein, beta-glucans, and amino acids in young pigs. J. Anim. Sci. 1996, 74, 1649–1656. [Google Scholar] [CrossRef]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bond, S.T.; McEwen, K.A.; Yoganantharajah, P.; Gibert, Y. Live Metabolic Profile Analysis of Zebrafish Embryos Using a Seahorse XF 24 Extracellular Flux Analyzer. Methods Mol. Biol. 2018, 1797, 393–401. [Google Scholar] [CrossRef]
- Gibert, Y.; McGee, S.L.; Ward, A.C. Metabolic Profile Analysis of Zebrafish Embryos. J. Vis. Exp. 2013, e4300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-H.; Huang, K.-S.; Yu, C.-H.; Gong, H.-Y. Metabolic profile analysis of a single developing zebrafish embryo via monitoring of oxygen consumption rates within a microfluidic device. Biomicrofluidics 2013, 7, 64107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabre, I.; Bruneteau, M.; Ricci, P.; Micheal, G. Isolement et etude structurale de glucanes de Phytophthora parasitica. Eur. J. Biochem. 1984, 142, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Tabata, K.; Itoh, W.; Yanaki, T. Molecular Weight Dependence of the Antitumor Activity of Schizophyllan. Agric. Biol. Chem. 1986, 50, 231–232. [Google Scholar] [CrossRef]
- Blaschek, W.; Käsbauer, J.; Kraus, J.; Franz, G. Pythium aphanidermatum: Culture, cell-wall composition, and isolation and structure of antitumour storage and solubilised cell-wall (1 → 3), (1 → 6)-β-d-glucans. Carbohydr. Res. 1992, 231, 293–307. [Google Scholar] [CrossRef]
- Creton, R. The calcium pump of the endoplasmic reticulum plays a role in midline signaling during early zebrafish development. Dev. Brain Res. 2004, 151, 33–41. [Google Scholar] [CrossRef]
- Lillicrap, A. The Use of Zebrafish Embryos as an Alternative Approach for Ecotoxicityn Testing. Master’s Thesis, University of Exeter, Exeter, UK, 2010. [Google Scholar]
- Paatero, I.; Casals, E.; Niemi, R.; Özliseli, E.; Rosenholm, J.M.; Sahlgren, C. Analyses in zebrafish embryos reveal that nanotoxicity profiles are dependent on surface-functionalization controlled penetrance of biological membranes. Sci. Rep. 2017, 7, 8423. [Google Scholar] [CrossRef] [Green Version]
- Cantó-Santos, J.; Grau-Junyent, J.M.; Garrabou, G. The Impact of Mitochondrial Deficiencies in Neuromuscular Diseases. Antioxidants 2020, 9, 964. [Google Scholar] [CrossRef]
- Kelly-Worden, M.; Thomas, E. Mitochondrial Dysfunction in Duchenne Muscular Dystrophy. Open J. Endocr. Metab. Dis. 2014, 4, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, J.E.; Pollard, A.K.; Lesanpezeshki, L.; Deane, C.S.; Gaffney, C.J.; Etheridge, T.; Szewczyk, N.J.; Vanapalli, S.A. Muscle strength deficiency and mitochondrial dysfunction in a muscular dystrophy model of Caenorhabditis elegans and its functional response to drugs. Dis. Model. Mech. 2018, 11, dmm.036137. [Google Scholar] [CrossRef] [Green Version]
- Heydemann, A. Skeletal Muscle Metabolism in Duchenne and Becker Muscular Dystrophy—Implications for Therapies. Nutrients 2018, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.M.; Lin, A.J.; Strumwasser, A.R.; Cory, K.; Whitney, K.; Ho, T.; Ho, T.; Lee, J.L.; Rucker, D.H.; Nguyen, C.Q.; et al. Mitochondrial Dysfunction Is an Early Consequence of Partial or Complete Dystrophin Loss in mdx Mice. Front. Physiol. 2020, 11, 690. [Google Scholar] [CrossRef]
- Zulian, A.; Schiavone, M.; Giorgio, V.; Bernardi, P. Forty years later: Mitochondria as therapeutic targets in muscle diseases. Pharmacol. Res. 2016, 113, 563–573. [Google Scholar] [CrossRef]
- Srivastava, N.K.; Yadav, R.; Mukherjee, S.; Pal, L.; Sinha, N. Abnormal lipid metabolism in skeletal muscle tissue of patients with muscular dystrophy: In vitro, high-resolution NMR spectroscopy-based observation in early phase of the disease. Magn. Reson. Imaging 2017, 38, 163–173. [Google Scholar] [CrossRef]
- Solsona-Vilarrasa, E.; Fucho, R.; Torres, S.; Nuñez, S.; Nuño-Lámbarri, N.; Enrich, C.; García-Ruiz, C.; Fernández-Checa, J.C. Cholesterol enrichment in liver mitochondria impairs oxidative phosphorylation and disrupts the assembly of respiratory supercomplexes. Redox Biol. 2019, 24, 101214. [Google Scholar] [CrossRef]
- Baggetto, L.G.; Clottes, E.; Vial, C. Low mitochondrial proton leak due to high membrane cholesterol content and cytosolic creatine kinase as two features of the deviant bioenergetics of Ehrlich and AS30-D tumor cells. Cancer Res. 1992, 52, 4935–4941. [Google Scholar]
- Kusmiati; Dhewantara, F.X.R. Cholesterol-Lowering Effect of Beta Glucan Extracted from Saccharomyces cerevisiae in Rats. Sci. Pharm. 2016, 84, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Timpani, C.A.; Mamchaoui, K.; Butler-Browne, G.; Rybalka, E. Nitric Oxide (NO) and Duchenne Muscular Dystrophy: NO Way to Go? Antioxidants 2020, 9, 1268. [Google Scholar] [CrossRef]
- Timpani, C.A.; Hayes, A.; Rybalka, E. Revisiting the dystrophin-ATP connection: How half a century of research still implicates mitochondrial dysfunction in Duchenne Muscular Dystrophy aeriology. Med. Hypotheses 2015, 85, 1021–1033. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brogi, L.; Marchese, M.; Cellerino, A.; Licitra, R.; Naef, V.; Mero, S.; Bibbiani, C.; Fronte, B. β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy. Nutrients 2021, 13, 1619. https://doi.org/10.3390/nu13051619
Brogi L, Marchese M, Cellerino A, Licitra R, Naef V, Mero S, Bibbiani C, Fronte B. β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy. Nutrients. 2021; 13(5):1619. https://doi.org/10.3390/nu13051619
Chicago/Turabian StyleBrogi, Letizia, Maria Marchese, Alessandro Cellerino, Rosario Licitra, Valentina Naef, Serena Mero, Carlo Bibbiani, and Baldassare Fronte. 2021. "β-Glucans as Dietary Supplement to Improve Locomotion and Mitochondrial Respiration in a Model of Duchenne Muscular Dystrophy" Nutrients 13, no. 5: 1619. https://doi.org/10.3390/nu13051619