
Modulatory Properties of Food and Nutraceutical Components Targeting NLRP3 Inflammasome Activation

 , , , , , ,
, , , , , ,  , and
, and
Abstract
:1. Introduction
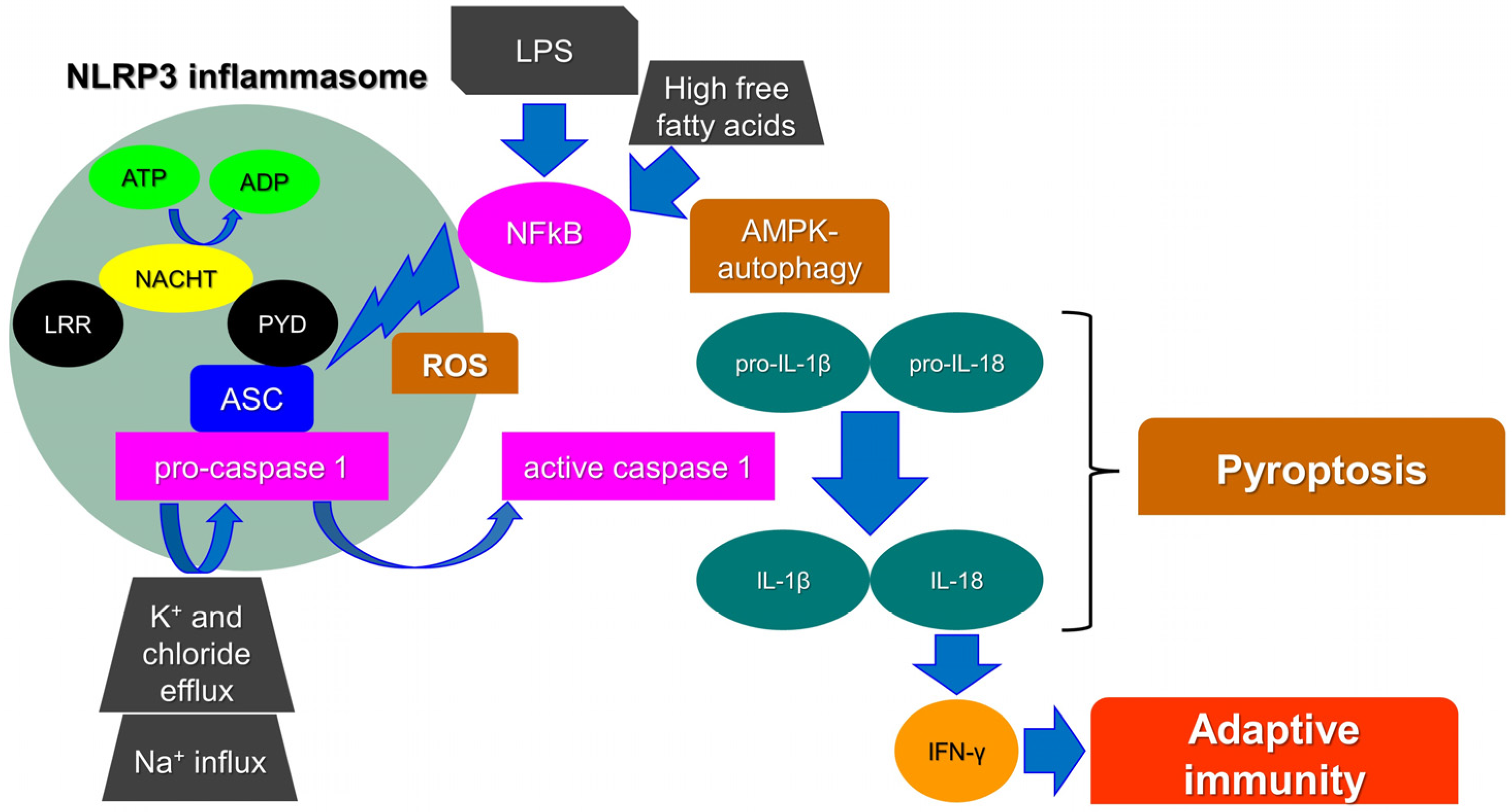
1.1. The NLRP3 Inflammasome
1.2. Food and Inflammation (Diet and Inflammation)
1.3. Role of Diet in Inflammatory Response
1.4. Fatty Acids and Inflammatory Response
- -
- Phosphorylation of insulin receptor substrate-1 (IRS-1) in serine and its degradation and elimination through the ubiquitin pathway, thus blocking insulin signal transmission;
- -
- Activation of the transcription factor NF-kB, by its dissociation from the cytoplasmic inhibitor (IkB) and transfer to the nucleus, with over-regulation of the pro-inflammatory gene expression.
1.5. Carbohydrates and Inflammatory Response
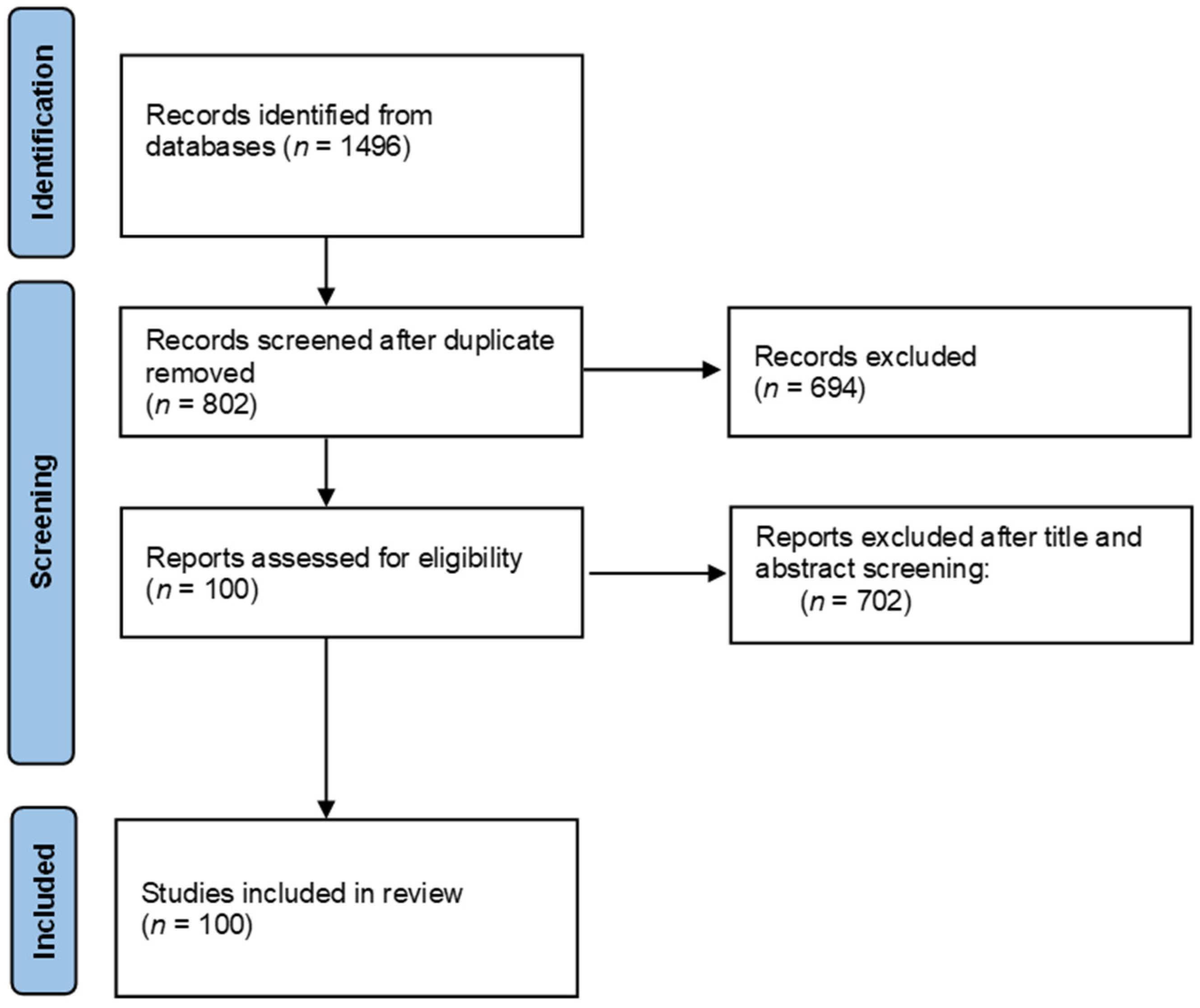
2. Materials and Methods
3. Food Components as NLPR3 Modulators
3.1. Polyphenols
3.1.1. Phenolic Acids
3.1.2. Flavones and Flavanones
3.1.3. Flavonols
3.1.4. Other Phenolics
3.1.5. Polyphenols Mixtures
3.2. Organosulfur Compounds
3.3. Terpenes and Terpenoids
3.4. Fatty Acids
3.4.1. Saturated Fatty Acids
3.4.2. Unsaturated Fatty Acids
3.5. Carotenoids
3.6. Proteins and Amino Acid Derivatives
3.7. Saponins and Sterols
3.8. Polysaccharides
3.9. Vitamins and Derivatives
3.10. Probiotics, Symbiotics, and Their Main Components
4. Food and Nutraceuticals Components Bioavailability and Toxicity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castejón-Vega, B.; Giampieri, F.; Alvarez-Suarez, J.M. Nutraceutical compounds targeting inflammasomes in human diseases. Int. J. Mol. Sci. 2020, 21, 4829. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 inflammasome: An overview of mechanisms of activation and regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fusco, R.; Siracusa, R.; Genovese, T.; Cuzzocrea, S.; Paola, R. Di Focus on the role of NLRP3 inflammasome in diseases. Int. J. Mol. Sci. 2020, 21, 4223. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.A.; Bergstralh, D.T.; Wang, Y.; Willingham, S.B.; Ye, Z.; Zimmermann, A.G.; Ting, J.P.Y. Cryopyrin/NALP3 binds ATP/dATP, is an ATPase, and requires ATP binding to mediate inflammatory signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 8041–8046. [Google Scholar] [CrossRef] [Green Version]
- Hafner-Bratkovič, I.; Sušjan, P.; Lainšček, D.; Tapia-Abellán, A.; Cerović, K.; Kadunc, L.; Angosto-Bazarra, D.; Pelegrίn, P.; Jerala, R. NLRP3 lacking the leucine-rich repeat domain can be fully activated via the canonical inflammasome pathway. Nat. Commun. 2018, 9, 5182. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, T.; Yang, Y.; Jin, T.; Jiang, W.; Zhou, R. Orchestration of NLRP3 Inflammasome Activation by Ion Fluxes. Trends Immunol. 2018, 39, 393–406. [Google Scholar] [CrossRef]
- Sharif, H.; Wang, L.; Wang, W.L.; Magupalli, V.G.; Andreeva, L.; Qiao, Q.; Hauenstein, A.V.; Wu, Z.; Núñez, G.; Mao, Y.; et al. Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome. Nature 2019, 570, 338–343. [Google Scholar] [CrossRef]
- Kuwar, R.; Rolfe, A.; Di, L.; Blevins, H.; Xu, Y.; Sun, X.; Bloom, G.S.; Zhang, S.; Sun, D. A novel inhibitor targeting NLRP3 inflammasome reduces neuropathology and improves cognitive function in alzheimer’s disease transgenic mice. J. Alzheimer’s Dis. 2021, 82, 1769–1783. [Google Scholar] [CrossRef]
- Liang, Z.; Damianou, A.; Di Daniel, E.; Kessler, B.M. Inflammasome activation controlled by the interplay between post-translational modifications: Emerging drug target opportunities. Cell Commun. Signal. 2021, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; Assanga, S.B.I.; Luján, L.L.; O’keefe, J.H.; Dinicolantonio, J.J. Nutraceutical strategies for suppressing nlrp3 inflammasome activation: Pertinence to the management of covid-19 and beyond. Nutrients 2021, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P.Y. The NLRP3 Inflammasome Mediates In Vivo Innate Immunity to Influenza A Virus through Recognition of Viral RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Son, S.; Hwang, I.; Hyeokhan, S.; Shin, J.S.; Shin, O.S.; Yu, J.W. Advanced glycation end products impair NLRP3 inflammasome-mediated innate immune responses in macrophages. J. Biol. Chem. 2017, 292, 20437–20448. [Google Scholar] [CrossRef] [Green Version]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.H.; Brickey, W.J.; Ting, J.P.Y. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, L.; Cardozo, L.F.M.F.; Borges, N.A.; Lindholm, B.; Stenvinkel, P.; Shiels, P.G.; Fouque, D.; Mafra, D. Can nutritional interventions modulate the activation of the NLRP3 inflammasome in chronic kidney disease? Food Res. Int. 2020, 136, 109306. [Google Scholar] [CrossRef]
- Drobny, E.C.; Abramson, E.C.; Baumann, G. Insulin receptors in acute infection: A study of factors conferring insulin resistance. J. Clin. Endocrinol. Metab. 1984, 58, 710–716. [Google Scholar] [CrossRef]
- Caputo, T.; Gilardi, F.; Desvergne, B. From chronic overnutrition to metaflammation and insulin resistance: Adipose tissue and liver contributions. FEBS Lett. 2017, 591, 3061–3088. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S1–S78. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbaresko, J.; Koch, M.; Schulze, M.B.; Nöthlings, U. Dietary pattern analysis and biomarkers of low-grade inflammation: A systematic literature review. Nutr. Rev. 2013, 71, 511–527. [Google Scholar] [CrossRef] [PubMed]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Tack, C.J.; Stienstra, R.; Joosten, L.A.B.; Netea, M.G. Inflammation links excess fat to insulin resistance: The role of the interleukin-1 family. Immunol. Rev. 2012, 249, 239–252. [Google Scholar] [CrossRef]
- Lagathu, C.; Yvan-Charvet, L.; Bastard, J.P.; Maachi, M.; Quignard-Boulangé, A.; Capeau, J.; Caron, M. Long-term treatment with interleukin-1β induces insulin resistance in murine and human adipocytes. Diabetologia 2006, 49, 2162–2173. [Google Scholar] [CrossRef] [Green Version]
- Stienstra, R.; Van Diepen, J.A.; Tack, C.J.; Zaki, M.H.; Van De Veerdonk, F.L.; Perera, D.; Neale, G.A.; Hooiveld, G.J.; Hijmans, A.; Vroegrijk, I.; et al. Inflammasome is a central player in the induction of obesity and insulin resistance. Proc. Natl. Acad. Sci. USA 2011, 108, 15324–15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, H.M. Dietary modulation of energy homoeostasis and metabolic-inflammation. Proc. Nutr. Soc. 2019, 78, 313–318. [Google Scholar] [CrossRef]
- McGillicuddy, F.C.; Harford, K.A.; Reynolds, C.M.; Oliver, E.; Claessens, M.; Mills, K.H.G.; Roche, H.M. Lack of interleukin-1 receptor I (IL-1RI) protects mice from high-fat diet-induced adipose tissue inflammation coincident with improved glucose homeostasis. Diabetes 2011, 60, 1688–1698. [Google Scholar] [CrossRef] [Green Version]
- Kirwan, A.M.; Lenighan, Y.M.; O’Reilly, M.E.; McGillicuddy, F.C.; Roche, H.M. Nutritional modulation of metabolic inflammation. Biochem. Soc. Trans. 2017, 45, 979–985. [Google Scholar] [CrossRef]
- Ralston, J.C.; Lyons, C.L.; Kennedy, E.B.; Kirwan, A.M.; Roche, H.M. Fatty Acids and NLRP3 Inflammasome-Mediated Inflammation in Metabolic Tissues. Annu. Rev. Nutr. 2017, 37, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Dewhurst-Trigg, R.; Hulston, C.J.; Markey, O. The effect of quantity and quality of dietary fat intake on subcutaneous white adipose tissue inflammatory responses. Proc. Nutr. Soc. 2020, 79, 542–556. [Google Scholar] [CrossRef]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncero-Ramos, I.; Rangel-Zuñiga, O.A.; Lopez-Moreno, J.; Alcala-Diaz, J.F.; Perez-Martinez, P.; Jimenez-Lucena, R.; Castaño, J.P.; Roche, H.M.; Delgado-Lista, J.; Ordovas, J.M.; et al. Mediterranean Diet, Glucose Homeostasis, and Inflammasome Genetic Variants: The CORDIOPREV Study. Mol. Nutr. Food Res. 2018, 62, 1700960. [Google Scholar] [CrossRef] [PubMed]
- Galland, L. Diet and inflammation. Nutr. Clin. Pract. 2010, 25, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Margioris, A.N. Fatty acids and postprandial inflammation. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 129–137. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Aro, A.; Willett, W.C. Health effects of trans-fatty acids: Experimental and observational evidence. Eur. J. Clin. Nutr. 2009, 63, S5–S21. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization Nutrition: Trans Fat. Available online: https://www.who.int/news-room/questions-and-answers/item/nutrition-trans-fat (accessed on 29 November 2021).
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A Systemic Review of the Roles of n-3 Fatty Acids in Health and Disease. J. Am. Diet. Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Jiang, W.; Spinetti, T.; Tardivel, A.; Castillo, R.; Bourquin, C.; Guarda, G.; Tian, Z.; Tschopp, J.; Zhou, R. Omega-3 Fatty Acids Prevent Inflammation and Metabolic Disorder through Inhibition of NLRP3 Inflammasome Activation. Immunity 2013, 38, 1154–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.R.; Midgette, Y.; Shah, R. Fish oil derived omega 3 fatty acids suppress adipose NLRP3 inflammasome signaling in human obesity. J. Endocr. Soc. 2019, 3, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, B. Anti-inflammatory diets for obesity and diabetes. J. Am. Coll. Nutr. 2009, 28, 482S–491S. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Devaraj, S.; Jialal, I. Dietary factors that promote or retard inflammation. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; Mcallan, L.; Nilaweera, K.N.; et al. Monounsaturated fatty acid-enriched high-fat diets impede adipose NLRP3 inflammasome-mediated IL-1β secretion and insulin resistance despite obesity. Diabetes 2015, 64, 2116–2128. [Google Scholar] [CrossRef] [Green Version]
- Levitan, E.B.; Cook, N.R.; Stampfer, M.J.; Ridker, P.M.; Rexrode, K.M.; Buring, J.E.; Manson, J.A.E.; Liu, S. Dietary glycemic index, dietary glycemic load, blood lipids, and C-reactive protein. Metabolism 2008, 57, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–226. [Google Scholar] [CrossRef]
- Pavillard, L.E.; Marín-Aguilar, F.; Bullon, P.; Cordero, M.D. Cardiovascular diseases, NLRP3 inflammasome, and western dietary patterns. Pharmacol. Res. 2018, 131, 44–50. [Google Scholar] [CrossRef]
- Lee, H.M.; Kim, J.J.; Kim, H.J.; Shong, M.; Ku, B.J.; Jo, E.K. Upregulated NLRP3 inflammasome activation in patients with type 2 diabetes. Diabetes 2013, 62, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef]
- Martinon, F. Signaling by ROS drives inflammasome activation. Eur. J. Immunol. 2010, 40, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Jhang, J.J.; Lin, J.H.; Yen, G.C. Beneficial Properties of Phytochemicals on NLRP3 Inflammasome-Mediated Gout and Complication. J. Agric. Food Chem. 2018, 66, 765–772. [Google Scholar] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H. Bin Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Qian, B.; Wang, C.; Zeng, Z.; Ren, Y.; Li, D.; Song, J. Le Ameliorative effect of sinapic acid on dextran sodium sulfate-(DSS-) induced ulcerative colitis in kunming (km) mice. Oxid. Med. Cell. Longev. 2020, 2020, 8393504. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hussein, O.E.; Abd El-Twab, S.M.; Hozayen, W.G. Ferulic acid protects against methotrexate nephrotoxicity: Via activation of Nrf2/ARE/HO-1 signaling and PPARγ, and suppression of NF-κB/NLRP3 inflammasome axis. Food Funct. 2019, 10, 4593–4607. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, D.; Wan, X.; Bai, Y.; Yuan, C.; Wang, T.; Yuan, D.; Zhang, C.; Liu, C. Chlorogenic Acid Suppresses miR-155 and Ameliorates Ulcerative Colitis through the NF-κB/NLRP3 Inflammasome Pathway. Mol. Nutr. Food Res. 2020, 64, e2000452. [Google Scholar] [CrossRef]
- Lv, Y.; Gao, X.; Luo, Y.; Fan, W.; Shen, T.; Ding, C.; Yao, M.; Song, S.; Yan, L. Apigenin ameliorates HFD-induced NAFLD through regulation of the XO/NLRP3 pathways. J. Nutr. Biochem. 2019, 71, 110–121. [Google Scholar] [CrossRef]
- Yamagata, K.; Hashiguchi, K.; Yamamoto, H.; Tagami, M. Dietary Apigenin Reduces Induction of LOX-1 and NLRP3 Expression, Leukocyte Adhesion, and Acetylated Low-Density Lipoprotein Uptake in Human Endothelial Cells Exposed to Trimethylamine-N-Oxide. J. Cardiovasc. Pharmacol. 2019, 74, 558–565. [Google Scholar] [CrossRef]
- An, M.F.; Wang, M.Y.; Shen, C.; Sun, Z.R.; Zhao, Y.L.; Wang, X.J.; Sheng, J. Isoorientin exerts a urate-lowering effect through inhibition of xanthine oxidase and regulation of the TLR4-NLRP3 inflammasome signaling pathway. J. Nat. Med. 2021, 75, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Chiang, Y.F.; Chen, H.Y.; Huang, Y.J.; Wang, K.L.; Hong, Y.H.; Ali, M.; Shieh, T.M.; Hsia, S.M. Anti-inflammatory and anti-hyperuricemic effects of chrysin on a high fructose corn syrup-induced hyperuricemia rat model via the amelioration of urate transporters and inhibition of nlrp3 inflammasome signaling pathway. Antioxidants 2021, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Sun, H.; Zhang, Y.; Xu, W.; Wang, C.; Fang, Y.; Zhao, J. Neuroprotective Effects of Luteolin Against Spinal Cord Ischemia-Reperfusion Injury by Attenuation of Oxidative Stress, Inflammation, and Apoptosis. J. Med. Food 2018, 21, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Miyazawa, K.W.; Pinho-Ribeiro, F.A.; Borghi, S.M.; Staurengo-Ferrari, L.; Fattori, V.; Amaral, F.A.; Teixeira, M.M.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; et al. Hesperidin Methylchalcone Suppresses Experimental Gout Arthritis in Mice by Inhibiting NF-κB Activation. J. Agric. Food Chem. 2018, 66, 6269–6280. [Google Scholar] [CrossRef]
- Cao, H.; Liu, J.; Shen, P.; Cai, J.; Han, Y.; Zhu, K.; Fu, Y.; Zhang, N.; Zhang, Z.; Cao, Y. Protective Effect of Naringin on DSS-Induced Ulcerative Colitis in Mice. J. Agric. Food Chem. 2018, 66, 13133–13140. [Google Scholar] [CrossRef]
- Liu, S.; Tian, L.; Chai, G.; Wen, B.; Wang, B. Targeting heme oxygenase-1 by quercetin ameliorates alcohol-induced acute liver injury via inhibiting NLRP3 inflammasome activation. Food Funct. 2018, 9, 4184–4193. [Google Scholar] [CrossRef]
- Li, H.; Xiao, L.; He, H.; Zeng, H.; Liu, J.; Jiang, C.; Mei, G.; Yu, J.; Chen, H.; Yao, P.; et al. Quercetin Attenuates Atherosclerotic Inflammation by Inhibiting Galectin-3-NLRP3 Signaling Pathway. Mol. Nutr. Food Res. 2021, 65, 2000746. [Google Scholar] [CrossRef]
- Li, H.; Chen, F.J.; Yang, W.L.; Qiao, H.Z.; Zhang, S.J. Quercetin improves cognitive disorder in aging mice by inhibiting NLRP3 inflammasome activation. Food Funct. 2021, 12, 717–725. [Google Scholar] [CrossRef]
- Xue, Y.; Du, M.; Zhu, M.J. Quercetin suppresses NLRP3 inflammasome activation in epithelial cells triggered by Escherichia coli O157:H7. Free Radic. Biol. Med. 2017, 108, 760–769. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, Q.; Li, X.; Jiang, M.; Cui, B.W.; Xia, K.L.; Wu, Y.L.; Lian, L.H.; Nan, J.X. Amelioration of Alcoholic Liver Steatosis by Dihydroquercetin through the Modulation of AMPK-Dependent Lipogenesis Mediated by P2X7R-NLRP3-Inflammasome Activation. J. Agric. Food Chem. 2018, 66, 4862–4871. [Google Scholar] [CrossRef]
- Du, Y.C.; Lai, L.; Zhang, H.; Zhong, F.R.; Cheng, H.L.; Qian, B.L.; Tan, P.; Xia, X.M.; Fu, W.G. Kaempferol from: Penthorum chinense Pursh suppresses HMGB1/TLR4/NF-?B signaling and NLRP3 inflammasome activation in acetaminophen-induced hepatotoxicity. Food Funct. 2020, 11, 7925–7934. [Google Scholar] [CrossRef]
- Lalitha, N.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. Anti-hyperglycemic activity of myricetin, through inhibition of DPP-4 and enhanced GLP-1 levels, is attenuated by co-ingestion with lectin-rich protein. PLoS ONE 2020, 15, e0231543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.-Y.; Guo, Y.-J.; Han, W.-X.; Yang, M.-Q.; Wen, L.-P.; Wang, K.-Y.; Jiang, P. Curcumin relieves depressive-like behaviors via inhibition of the NLRP3 inflammasome and kynurenine pathway in rats suffering from chronic unpredictable mild stress. Int. Immunopharmacol. 2019, 67, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, L.; Salarolli, R.; Cardozo, L.F.M.F.; Santos, R.S.; de Brito, J.S.; Kemp, J.A.; Reis, D.; de Paiva, B.R.; Stenvinkel, P.; Lindholm, B.; et al. Impact of curcumin supplementation on expression of inflammatory transcription factors in hemodialysis patients: A pilot randomized, double-blind, controlled study. Clin. Nutr. 2020, 39, 3594–3600. [Google Scholar] [CrossRef]
- Chen, T.C.; Yen, C.K.; Lu, Y.C.; Shi, C.S.; Hsieh, R.Z.; Chang, S.F.; Chen, C.N. The antagonism of 6-shogaol in high-glucose-activated NLRP3 inflammasome and consequent calcification of human artery smooth muscle cells. Cell Biosci. 2020, 10, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.C.; Chang, Y.H. Comparison of inhibitory capacities of 6-, 8- and 10-gingerols/shogaols on the canonical NLRP3 inflammasome-mediated IL-1β secretion. Molecules 2018, 23, 466. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Huang, J.; Wang, K.; Yu, Q.; Zhu, C.; Ren, H. Pterostilbene Protects against Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Failure by Upregulating the Nrf2 Pathway and Inhibiting NF-κB, MAPK, and NLRP3 Inflammasome Activation. J. Med. Food 2020, 23, 952–960. [Google Scholar] [CrossRef]
- Wang, B.J.; Chiu, H.W.; Lee, Y.L.; Li, C.Y.; Wang, Y.J.; Lee, Y.H. Pterostilbene attenuates hexavalent chromium-induced allergic contact dermatitis by preventing cell apoptosis and inhibiting IL-1β-related NLRP3 inflammasome activation. J. Clin. Med. 2018, 7, 489. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Chen, Y.Y.; Hsiao, C.M.; Pan, M.H.; Wang, B.J.; Chen, Y.C.; Ho, C.T.; Huang, K.C.; Chen, R.J. Induction of Autophagy by Pterostilbene Contributes to the Prevention of Renal Fibrosis via Attenuating NLRP3 Inflammasome Activation and Epithelial-Mesenchymal Transition. Front. Cell Dev. Biol. 2020, 8, 436. [Google Scholar] [CrossRef]
- Chen, L.; Lan, Z. Polydatin attenuates potassium oxonate-induced hyperuricemia and kidney inflammation by inhibiting NF-κB/NLRP3 inflammasome activation via the AMPK/SIRT1 pathway. Food Funct. 2017, 8, 1785–1792. [Google Scholar] [CrossRef]
- Park, B.; Jo, K.; Lee, T.G.; Hyun, S.W.; Kim, J.S.; Kim, C.S. Polydatin inhibits NLRP3 inflammasome in dry eye disease by attenuating oxidative stress and inhibiting the nf-κb pathway. Nutrients 2019, 11, 2792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Wang, C.; Wu, W.; Liu, T.; Ji, B.; Zhou, F. Cyanidin-3-glucoside alleviates 4-Hydroxyhexenal-induced NLRP3 inflammasome activation via JNK-c-Jun/AP-1 pathway in human retinal pigment epithelial cells. J. Immunol. Res. 2018, 2018, 5604610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, S.; Wan, T.; Huang, Y.; Pang, N.; Jiang, X.; Gu, Y.; Zhang, Z.; Luo, J.; Yang, L. Cyanidin-3-O-β-glucoside inactivates NLRP3 inflammasome and alleviates alcoholic steatohepatitis via SirT1/NF-κB signaling pathway. Free Radic. Biol. Med. 2020, 160, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.L.; Sun, H.Y.; Wu, X.B.; Cheng, L.; Ren, J.D. Epigallocatechin-3-gallate attenuates acute pancreatitis induced lung injury by targeting mitochondrial reactive oxygen species triggered NLRP3 inflammasome activation. Food Funct. 2021, 12, 5658–5667. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Liu, M.; Yao, W.; Du, K.; He, M.; Jin, X.; Jiao, L.; Ma, G.; Wei, B.; Wei, M. Epigallocatechin-3-Gallate Attenuates Microglial Inflammation and Neurotoxicity by Suppressing the Activation of Canonical and Noncanonical Inflammasome via TLR4/NF-κB Pathway. Mol. Nutr. Food Res. 2019, 63, 1801230. [Google Scholar] [CrossRef]
- Lin, Y.F.; Lee, Y.H.; Hsu, Y.H.; Chen, Y.J.; Lin, Y.F.; Cheng, F.Y.; Chiu, H.W. Resveratrol-loaded nanoparticles conjugated with kidney injury molecule-1 as a drug delivery system for potential use in chronic kidney disease. Nanomedicine 2017, 12, 2741–2756. [Google Scholar] [CrossRef]
- Luzardo-Ocampo, I.; Loarca-Piña, G.; Gonzalez de Mejia, E. Gallic and butyric acids modulated NLRP3 inflammasome markers in a co-culture model of intestinal inflammation. Food Chem. Toxicol. 2020, 146, 111835. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Q.; Wang, T.; Kan, Z.; Li, X.; Hu, L.; Peng, C.-Y.; Qian, F.; Wang, Y.; Granato, D. Green tea polyphenols and epigallocatechin-3-gallate protect against perfluorodecanoic acid induced liver damage and inflammation in mice by inhibiting NLRP3 inflammasome activation. Food Res. Int. 2020, 127, 108628. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, M.; Wang, T.; Cai, M.; Qian, F.; Sun, Y.; Wang, Y. Green tea polyphenols prevent lipopolysaccharide-induced inflammatory liver injury in mice by inhibiting NLRP3 inflammasome activation. Food Funct. 2019, 10, 3898–3908. [Google Scholar] [CrossRef]
- Gao, X.; Fan, W.; Tan, L.; Shi, Y.; Ding, C.; Liu, S.; Miao, Y.; Luo, Y.; Shi, X.; DeSaeger, S.; et al. Soy isoflavones ameliorate experimental colitis by targeting ERα/NLRP3 inflammasome pathways. J. Nutr. Biochem. 2020, 83, 108438. [Google Scholar] [CrossRef]
- Fan, R.; You, M.; Toney, A.M.; Kim, J.; Giraud, D.; Xian, Y.; Ye, F.; Gu, L.; Ramer-Tait, A.E.; Chung, S. Red Raspberry Polyphenols Attenuate High-Fat Diet–Driven Activation of NLRP3 Inflammasome and its Paracrine Suppression of Adipogenesis via Histone Modifications. Mol. Nutr. Food Res. 2020, 64, e1900995. [Google Scholar] [CrossRef] [PubMed]
- Nan, B.; Yang, C.; Li, L.; Ye, H.; Yan, H.; Wang, M.; Yuan, Y. Allicin alleviated acrylamide-induced NLRP3 inflammasome activation via oxidative stress and endoplasmic reticulum stress in Kupffer cells and SD rats liver. Food Chem. Toxicol. 2021, 148, 111937. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Yen, C.C.; Kuo, L.L.; Lo, C.W.; Huang, C.S.; Chen, C.C.; Lii, C.K. Benzyl isothiocyanate ameliorates high-fat/cholesterol/cholic acid diet-induced nonalcoholic steatohepatitis through inhibiting cholesterol crystal-activated NLRP3 inflammasome in Kupffer cells. Toxicol. Appl. Pharmacol. 2020, 393, 114941. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Shang, H.; Chen, Y.Q.; Pan, L.L.; Bhatia, M.; Sun, J. Sulforaphane Protects Pancreatic Acinar Cell Injury by Modulating Nrf2-Mediated Oxidative Stress and NLRP3 Inflammatory Pathway. Oxid. Med. Cell. Longev. 2016, 2016, 4916497. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.; Kim, J.; Lee, M.J.; Kim, Y.J.; Cho, Y.W.; Lee, G.S. Methylsulfonylmethane inhibits NLRP3 inflammasome activation. Cytokine 2015, 71, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Xia, Z.; Shao, N.; Li, B.; Xue, L.; Peng, Y.; Zhi, F.; Yang, Y. Carnosic acid prevents dextran sulfate sodium-induced acute colitis associated with the regulation of the Keap1/Nrf2 pathway. Sci. Rep. 2017, 7, 12117. [Google Scholar] [CrossRef] [PubMed]
- Marcuzzi, A.; Piscianz, E.; Zweyer, M.; Bortul, R.; Loganes, C.; Girardelli, M.; Baj, G.; Monasta, L.; Celeghini, C. Geranylgeraniol and neurological impairment: Involvement of apoptosis and mitochondrial morphology. Int. J. Mol. Sci. 2016, 17, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.M.; Panis, C.; Da Silva, S.S.; Macri, J.A.; Kawakami, N.Y.; Hayashida, T.H.; Madeira, T.B.; Acquaro, V.R.; Nixdorf, S.L.; Pizzatti, L.; et al. Kaurenoic acid possesses leishmanicidal activity by triggering a NLRP12/IL-1 β/cNOS/NO Pathway. Mediat. Inflamm. 2015, 2015, 392918. [Google Scholar] [CrossRef] [Green Version]
- Robblee, M.M.; Kim, C.C.; Abate, J.P.; Valdearcos, M.; Sandlund, K.L.M.; Shenoy, M.K.; Volmer, R.; Iwawaki, T.; Koliwad, S.K. Saturated Fatty Acids Engage an IRE1α-Dependent Pathway to Activate the NLRP3 Inflammasome in Myeloid Cells. Cell Rep. 2016, 14, 2611–2623. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, F.; Khazaei, M.; Komaki, A.; Amiri, I.; Jalili, C. Virgin coconut oil (VCO) by normalizing NLRP3 inflammasome showed potential neuroprotective effects in Amyloid-β induced toxicity and high-fat diet fed rat. Food Chem. Toxicol. 2018, 118, 68–83. [Google Scholar] [CrossRef]
- López-Tenorio, I.I.; Domínguez-López, A.; Miliar-García, Á.; Escalona-Cardoso, G.N.; Real-Sandoval, S.A.; Gómez-Alcalá, A.; Jaramillo-Flores, M.E. Modulation of the mRNA of the Nlrp3 inflammasome by Morin and PUFAs in an obesity model induced by a high-fat diet. Food Res. Int. 2020, 137, 109706. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Wang, T.; Wang, F.; Yang, Y.; He, C.; Yang, W.; Zhang, J.; Zou, Z. High n-3 fatty acids counteract hyperglycemia-induced insulin resistance in fat-1 mice: Via pre-adipocyte NLRP3 inflammasome inhibition. Food Funct. 2021, 12, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Shan, C.; Ma, T.; Geng, S.; Ning, D. Walnut oil alleviates DSS–induced colitis in mice by inhibiting NLRP3 inflammasome activation and regulating gut microbiota. Microb. Pathog. 2021, 154, 104866. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.; Li, Y.; Chen, J.; Zong, Y.; Yang, X. DHA/AA alleviates LPS-induced Kupffer cells pyroptosis via GPR120 interaction with NLRP3 to inhibit inflammasome complexes assembly. Cell Death Dis. 2021, 12, 73. [Google Scholar] [CrossRef]
- Dang, R.; Zhou, X.; Tang, M.; Xu, P.; Gong, X.; Liu, Y.; Jiao, H.; Jiang, P. Fish oil supplementation attenuates neuroinflammation and alleviates depressive-like behavior in rats submitted to repeated lipopolysaccharide. Eur. J. Nutr. 2018, 57, 893–906. [Google Scholar] [CrossRef]
- Liu, N.; Meng, B.; Zeng, L.; Yin, S.; Hu, Y.; Li, S.; Fu, Y.; Zhang, X.; Xie, C.; Shu, L.; et al. Discovery of a novel rice-derived peptide with significant anti-gout potency. Food Funct. 2020, 11, 10542–10553. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, Y.; Zeng, L.; Yin, S.; Hu, Y.; Li, S.; Fu, Y.; Zhang, X.; Xie, C.; Shu, L.; et al. RDP3, A Novel Antigout Peptide Derived from Water Extract of Rice. J. Agric. Food Chem. 2020, 68, 7143–7151. [Google Scholar] [CrossRef]
- Han, J.; Wang, X.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Li, Y.; Ming, T.; Wang, Z.J.; Su, X. Protective effects of tuna meat oligopeptides (TMOP) supplementation on hyperuricemia and associated renal inflammation mediated by gut microbiota. FASEB J. 2020, 34, 5061–5076. [Google Scholar] [CrossRef] [Green Version]
- Bitzer, Z.T.; Wopperer, A.L.; Chrisfield, B.J.; Tao, L.; Cooper, T.K.; Vanamala, J.; Elias, R.J.; Hayes, J.E.; Lambert, J.D. Soy protein concentrate mitigates markers of colonic inflammation and loss of gut barrier function in vitro and in vivo. J. Nutr. Biochem. 2017, 40, 201–208. [Google Scholar] [CrossRef]
- Gómez Castro, M.F.; Miculán, E.; Herrera, M.G.; Ruera, C.; Perez, F.; Prieto, E.D.; Barrera, E.; Pantano, S.; Carasi, P.; Chirdo, F.G. P31-43 gliadin peptide forms oligomers and induces NLRP3 inflammasome/caspase 1-dependent mucosal damage in small intestine. Front. Immunol. 2019, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Xu, X.; Sho, T.; Luo, W.; Zhang, J.; Xu, W.; Yao, J.; Xu, J. Effects of n-acetyl-cysteine supplementation in late gestational diet on maternal-placental redox status, placental NLRP3 inflammasome, and fecal microbiota in sows. J. Anim. Sci. 2019, 97, 1757–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, X.; Jiang, D.; Chen, J.; Jia, H.; Wu, Z.; Kim, I.H.; Yang, Y. Glycine attenuates lipopolysaccharide-induced acute lung injury by regulating NLRP3 inflammasome and NRF2 signaling. Nutrients 2020, 12, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Lu, X.Y.; Zhu, W.L.; Liu, X.Q.; Li, B.Y.; Song, L.; Liu, H.F.; Cai, W.W.; Deng, Y.X.; Xu, T.T.; et al. Carnosine ameliorates age-related dementia: Via improving mitochondrial dysfunction in SAMP8 mice. Food Funct. 2020, 11, 2489–2497. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, T.; Yu, D.; Fang, X.; Fan, H.; Liu, Q.; Yi, G.; Yi, X.; Liu, Q. L-Homocarnosine attenuates inflammation in cerebral ischemia–reperfusion injury through inhibition of nod-like receptor protein 3 inflammasome. Int. J. Biol. Macromol. 2018, 118, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guan, X.; Chen, X.; Cai, Y.; Ma, Y.; Ma, J.; Zhang, Q.; Dai, L.; Fan, X.; Bai, Y. Choline Supplementation Ameliorates Behavioral Deficits and Alzheimer’s Disease-Like Pathology in Transgenic APP/PS1 Mice. Mol. Nutr. Food Res. 2019, 63, 1801407. [Google Scholar] [CrossRef]
- Han, X.; Song, J.; Lian, L.H.; Yao, Y.L.; Shao, D.Y.; Fan, Y.; Hou, L.S.; Wang, G.; Zheng, S.; Wu, Y.L.; et al. Ginsenoside 25-OCH3-PPD Promotes Activity of LXRs to Ameliorate P2X7R-Mediated NLRP3 Inflammasome in the Development of Hepatic Fibrosis. J. Agric. Food Chem. 2018, 66, 7023–7035. [Google Scholar] [CrossRef]
- Wang, F.; Park, J.S.; Ma, Y.; Ma, H.; Lee, Y.J.; Lee, G.R.; Yoo, H.S.; Roh, Y.S.; Hong, J.T. Ginseng saponin enriched in rh1 and rg2 ameliorates nonalcoholic fatty liver disease by inhibiting inflammasome activation. Nutrients 2021, 13, 856. [Google Scholar] [CrossRef]
- Li, C.W.; Deng, M.Z.; Gao, Z.J.; Dang, Y.Y.; Zheng, G.D.; Yang, X.J.; Chao, Y.X.; Cai, Y.F.; Wu, X.L. Effects of compound K, a metabolite of ginsenosides, on memory and cognitive dysfunction in db/db mice involve the inhibition of ER stress and the NLRP3 inflammasome pathway. Food Funct. 2020, 11, 4416–4427. [Google Scholar] [CrossRef]
- Zhao, X.J.; Yang, Y.Z.; Zheng, Y.J.; Wang, S.C.; Gu, H.M.; Pan, Y.; Wang, S.J.; Xu, H.J.; Kong, L.D. Dataset on assessment of magnesium isoglycyrrhizinate injection for dairy diet and body weight in fructose-induced metabolic syndrome of rats. Data Br. 2018, 18, 69–75. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, N.N.; Hu, X.; Zheng, Y. Anti-colitic effects of Physalin B on dextran sodium sulfate-induced BALB/c mice by suppressing multiple inflammatory signaling pathways. J. Ethnopharmacol. 2020, 259, 112956. [Google Scholar] [CrossRef]
- Liao, P.C.; Lai, M.H.; Hsu, K.P.; Kuo, Y.H.; Chen, J.; Tsai, M.C.; Li, C.X.; Yin, X.J.; Jeyashoke, N.; Chao, L.K.P. Identification of β-Sitosterol as in Vitro Anti-Inflammatory Constituent in Moringa oleifera. J. Agric. Food Chem. 2018, 66, 10748–10759. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fan, J.; Chen, H.W.; Liu, E.Q. Trametes orientalis polysaccharide alleviates PM2.5-induced lung injury in mice through its antioxidant and anti-inflammatory activities. Food Funct. 2019, 10, 8005–8015. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, S.; Chen, J.; Lin, J.; Xiong, Q.; Yang, Y.; Yuan, J.; Zhou, L.; He, L.; Hou, S.; et al. Therapeutic roles of polysaccharides from Dendrobium Officinaleon colitis and its underlying mechanisms. Carbohydr. Polym. 2018, 185, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xiao, N.; Zeng, L.; Xiao, J.; Huang, J.; Xu, Y.; Chen, Y.; Ren, Y.; Du, B. Structural characteristics of a mannoglucan isolated from Chinese yam and its treatment effects against gut microbiota dysbiosis and DSS-induced colitis in mice. Carbohydr. Polym. 2020, 250, 116958. [Google Scholar] [CrossRef]
- Chen, Y.S.; Chen, Q.Z.; Wang, Z.J.; Hua, C. Anti-inflammatory and hepatoprotective effects of Ganoderma lucidum polysaccharides against carbon tetrachloride-induced liver injury in Kunming Mice. Pharmacology 2019, 103, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Li, Y.; Cai, J.; Ji, J.; Wang, Y.; Zhang, W.; Pan, W.; Chen, Y. Polysaccharides from: Armillariella tabescens mycelia ameliorate insulin resistance in type 2 diabetic mice. Food Funct. 2020, 11, 9675–9685. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Pan, L.L.; Niu, W.; Fang, X.; Liang, W.; Li, J.; Li, H.; Pan, X.; Chen, W.; Zhang, H.; et al. Modulation of Gut Microbiota by Low Methoxyl Pectin Attenuates Type 1 Diabetes in Non-obese Diabetic Mice. Front. Immunol. 2019, 10, 1733. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Pan, L.L.; Luo, Y.; Niu, W.; Fang, X.; Liang, W.; Li, J.; Li, H.; Pan, X.; Yang, G.; et al. Low Methoxyl Pectin Protects against Autoimmune Diabetes and Associated Caecal Dysfunction. Mol. Nutr. Food Res. 2019, 63, e1900307. [Google Scholar] [CrossRef]
- Castro-Alves, V.C.; Shiga, T.M.; Nascimento, J.R.O. do Polysaccharides from chayote enhance lipid efflux and regulate NLRP3 inflammasome priming in macrophage-like THP-1 cells exposed to cholesterol crystals. Int. J. Biol. Macromol. 2019, 127, 502–510. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, X.; Wang, Y.; Jin, W.; Guo, Y. The immunoenhancement effects of starfish: Asterias rollestoni polysaccharides in macrophages and cyclophosphamide-induced immunosuppression mouse models. Food Funct. 2020, 11, 10700–10708. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Chen, C. Sinapic acid and its derivatives as medicine in oxidative stress-induced diseases and aging. Oxid. Med. Cell. Longev. 2016, 2016, 3571614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosazza, J.P.N.; Huang, Z.; Dostal, L.; Volm, T.; Rousseau, B. Review: Biocatalytic transformations of ferulic acid: An abundant aromatic natural product. J. Ind. Microbiol. 1995, 15, 457–471. [Google Scholar] [CrossRef]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic Acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.C.; Chang, Y.H.; Chang, K.S. Structural moieties required for cinnamaldehyde-related compounds to inhibit canonical IL-1β secretion. Molecules 2018, 23, 3241. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wang, J.; Wen, W.; Pan, T.; Chen, H.; Fu, Y.; Wang, F.; Huang, J.H.; Xu, S. Cinnamaldehyde suppresses NLRP3 derived IL-1β via activating succinate/HIF-1 in rheumatoid arthritis rats. Int. Immunopharmacol. 2020, 84, 106570. [Google Scholar] [CrossRef]
- Hassanzadeh, S.; Read, M.I.; Bland, A.R.; Majeed, M.; Jamialahmadi, T.; Sahebkar, A. Curcumin: An inflammasome silencer. Pharmacol. Res. 2020, 159, 104921. [Google Scholar] [CrossRef]
- Netea-Maier, R.T.; Plantinga, T.S.; van de Veerdonk, F.L.; Smit, J.W.; Netea, M.G. Modulation of inflammation by autophagy: Consequences for human disease. Autophagy 2016, 12, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Meckling, K.A.; Marcone, M.F.; Kakuda, Y.; Tsao, R. Synergistic, additive, and antagonistic effects of food mixtures on total antioxidant capacities. J. Agric. Food Chem. 2011, 59, 960–968. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Roberts, L.A.; Ma, S.; Suzuki, K. Organosulfur Compounds: A Review of Their Anti-inflammatory Effects in Human Health. Front. Nutr. 2020, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Osipova, V.; Polovinkina, M.; Gracheva, Y.; Shpakovsky, D.; Osipova, A.; Berberova, N. Antioxidant activity of some organosulfur compounds in vitro. Arab. J. Chem. 2021, 14, 103068. [Google Scholar] [CrossRef]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Bursać Kovačević, D. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Chen, X.; Li, Y.; Guo, S.; Wang, Z.; Yu, X. Advances in Pharmacological Activities of Terpenoids. Nat. Prod. Commun. 2020, 15, 1934578X20903555. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Bardaweel, S.K.; Mubarak, M.S.; Koch, W.; Gaweł-Beben, K.; Antosiewicz, B.; Sharifi-Rad, J. Immunomodulatory Effects of Diterpenes and Their Derivatives Through NLRP3 Inflammasome Pathway: A Review. Front. Immunol. 2020, 11, 572136. [Google Scholar] [CrossRef]
- Zhao, X.; Pu, D.; Zhao, Z.; Zhu, H.; Li, H.; Shen, Y.; Zhang, X.; Zhang, R.; Shen, J.; Xiao, W.; et al. Teuvincenone F suppresses LPS-induced inflammation and NLRP3 inflammasome activation by attenuating NEMO ubiquitination. Front. Pharmacol. 2017, 8, 565. [Google Scholar] [CrossRef] [Green Version]
- Aachoui, Y.; Chowdhury, R.R.; Fitch, R.W.; Ghosh, S.K. Molecular signatures of phytol-derived immunostimulants in the context of chemokine-cytokine microenvironment and enhanced immune response. Cell. Immunol. 2011, 271, 227–238. [Google Scholar] [CrossRef]
- Liu, D.; Qin, H.; Yang, B.; Du, B.; Yun, X. Oridonin ameliorates carbon tetrachloride-induced liver fibrosis in mice through inhibition of the NLRP3 inflammasome. Drug Dev. Res. 2020, 81, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.X.; Yuan, Y.; Liu, C.; Cai, Z.; Xie, Q.; Hu, T.; Tang, Q.; Wu, Q.Q. Indigo fruits ingredient, aucubin, protects against LPS-induced cardiac dysfunction in mice. J. Pharmacol. Exp. Ther. 2019, 371, 348–359. [Google Scholar] [CrossRef]
- Zhao, C.C.; Xu, J.; Xie, Q.M.; Zhang, H.Y.; Fei, G.H.; Wu, H.M. Abscisic acid suppresses the activation of NLRP3 inflammasome and oxidative stress in murine allergic airway inflammation. Phyther. Res. 2021, 35, 3298–3309. [Google Scholar] [CrossRef]
- Huang, Y.; Wan, T.; Pang, N.; Zhou, Y.; Jiang, X.; Li, B.; Gu, Y.; Huang, Y.; Ye, X.; Lian, H.; et al. Cannabidiol protects livers against nonalcoholic steatohepatitis induced by high-fat high cholesterol diet via regulating NF-κB and NLRP3 inflammasome pathway. J. Cell. Physiol. 2019, 234, 21224–21234. [Google Scholar] [CrossRef] [PubMed]
- Nagoor Meeran, M.F.; Azimullah, S.; Laham, F.; Tariq, S.; Goyal, S.N.; Adeghate, E.; Ojha, S. α-Bisabolol protects against β-adrenergic agonist-induced myocardial infarction in rats by attenuating inflammation, lysosomal dysfunction, NLRP3 inflammasome activation and modulating autophagic flux. Food Funct. 2020, 11, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, F.; Li, L.; Lynch, E.C.; Xie, L.; Zhao, Y.; Fang, K.; Li, J.; Luo, J.; Xu, L.; et al. Celastrol alleviates metabolic disturbance in high-fat diet-induced obese mice through increasing energy expenditure by ameliorating metabolic inflammation. Phyther. Res. 2021, 35, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Wang, X.; Zhong, X.; Ji, L.; Guo, Z.; Liu, Y.; Shang, X. Sesquiterpenoids and their anti-inflammatory activity: Evaluation of Ainsliaea yunnanensis. Molecules 2019, 24, 1701. [Google Scholar] [CrossRef] [Green Version]
- Gianfrancesco, M.A.; Dehairs, J.; L’homme, L.; Herinckx, G.; Esser, N.; Jansen, O.; Habraken, Y.; Lassence, C.; Swinnen, J.V.; Rider, M.H.; et al. Saturated fatty acids induce NLRP3 activation in human macrophages through K + efflux resulting from phospholipid saturation and Na, K-ATPase disruption. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2019, 1864, 1017–1030. [Google Scholar] [CrossRef]
- Witcher, K.J.; Novick, R.P.; Schlievert, P.M. Modulation of immune cell proliferation by glycerol monolaurate. Clin. Diagn. Lab. Immunol. 1996, 3, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Yang, Y.; Ou, T.; Key, C.C.C.; Tong, S.H.; Sequeira, R.C.; Nelson, J.M.; Nie, Y.; Wang, Z.; Boudyguina, E.; et al. Dietary PUFAs attenuate NLRP3 inflammasome activation via enhancing macrophage autophagy. J. Lipid Res. 2017, 58, 1808–1821. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Lv, Y.; Liu, Y.; Li, J.; Wang, X.; Zhou, Z.; Tipoe, G.L.; Ouyang, S.; Guo, Y.; Zhang, J.; et al. Wolfberry-Derived Zeaxanthin Dipalmitate Attenuates Ethanol-Induced Hepatic Damage. Mol. Nutr. Food Res. 2019, 63, e1801339. [Google Scholar] [CrossRef]
- Wu, L.; Lyu, Y.; Srinivasagan, R.; Wu, J.; Ojo, B.; Tang, M.; El-Rassi, G.D.; Metzinger, K.; Smith, B.J.; Lucas, E.A.; et al. Astaxanthin-Shifted gut microbiota is associated with inflammation and metabolic homeostasis in mice. J. Nutr. 2020, 150, 2687–2698. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef]
- Dolinay, T.; Kim, Y.S.; Howrylak, J.; Hunninghake, G.M.; An, C.H.; Fredenburgh, L.; Massaro, A.F.; Rogers, A.; Gazourian, L.; Nakahira, K.; et al. Inflammasome-regulated cytokines are critical mediators of acute lung injury. Am. J. Respir. Crit. Care Med. 2012, 185, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Bauer, K. Carnosine and homocarnosine, the forgotten, enigmatic peptides of the brain. Neurochem. Res. 2005, 30, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Lin, C.Y.; Huang, S.F.; Lin, H.C.; Chang, W.L.; Chang, T.C. Effect and mechanism of ginsenosides CK and Rg1 on stimulation of glucose uptake in 3T3-L1 adipocytes. J. Agric. Food Chem. 2010, 58, 6039–6047. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, J.N.; Yan, M.H.; Xing, J.J.; Liu, W.C.; Li, W. Caspase-Mediated Anti-Apoptotic Effect of Ginsenoside Rg5, a Main Rare Ginsenoside, on Acetaminophen-Induced Hepatotoxicity in Mice. J. Agric. Food Chem. 2017, 65, 9226–9236. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Wan, Y.; Jin, X.J.; Ouyang, B.Q.; Bai, T.; Zhao, Y.Q.; Nan, J.X. 25-OCH3-PPD induces the apoptosis of activated t-HSC/Cl-6 cells via c-FLIP-mediated NF-κB activation. Chem. Biol. Interact. 2011, 194, 106–112. [Google Scholar] [CrossRef]
- Han, G.C.; Ko, S.K.; Sung, J.H.; Chung, S.H. Compound K enhances insulin secretion with beneficial metabolic effects in db/db mice. J. Agric. Food Chem. 2007, 55, 10641–10648. [Google Scholar] [CrossRef]
- Song, W.; Wei, L.; Du, Y.; Wang, Y.; Jiang, S. Protective effect of ginsenoside metabolite compound K against diabetic nephropathy by inhibiting NLRP3 inflammasome activation and NF-κB/p38 signaling pathway in high-fat diet/streptozotocin-induced diabetic mice. Int. Immunopharmacol. 2018, 63, 227–238. [Google Scholar] [CrossRef]
- Mao, Y.M.; Zeng, M.D.; Chen, Y.; Chen, C.W.; Fu, Q.C.; Cai, X.; Wu, S.M.; Chen, Y.G.; Sun, Y.; Li, J.; et al. Magnesium isoglycyrrhizinate in the treatment of chronic liver diseases: A randomized, double-blind, multi-doses, active drug controlled, multi-center study. Chin. J. Hepatol. 2009, 17, 847–851. [Google Scholar]
- Zhao, X.J.; Yang, Y.Z.; Zheng, Y.J.; Wang, S.C.; Gu, H.M.; Pan, Y.; Wang, S.J.; Xu, H.J.; Kong, L.D. Magnesium isoglycyrrhizinate blocks fructose-induced hepatic NF-κB/NLRP3 inflammasome activation and lipid metabolism disorder. Eur. J. Pharmacol. 2017, 809, 141–150. [Google Scholar] [CrossRef]
- Vieira, A.T.; Pinho, V.; Lepsch, L.B.; Scavone, C.; Ribeiro, I.M.; Tomassini, T.; Ribeiro-Dos-Santos, R.; Soares, M.B.P.; Teixeira, M.M.; Souza, D.G. Mechanisms of the anti-inflammatory effects of the natural secosteroids physalins in a model of intestinal ischaemia and reperfusion injury. Br. J. Pharmacol. 2005, 146, 244–251. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Q.; Li, Z.; Gao, Y.; Pang, Z.; Wu, Y.; Li, G.; Lu, D.; Zhang, L.; Li, D. Astragalus Polysaccharides Attenuate Ovalbumin-Induced Allergic Rhinitis in Rats by Inhibiting NLRP3 Inflammasome Activation and NOD2-Mediated NF-?B Activation. J. Med. Food 2021, 24, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zong, S.; Ye, Z.; Zhang, X.; Chen, H.; Ye, M. Protective effect of: Lachnum polysaccharide on dextran sulfate sodium-induced colitis in mice. Food Funct. 2020, 11, 846–859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Han, S.; Zhang, Z.; Zhang, W.; Yang, J.; Wan, Z.; Qin, L. Cereal Fiber Ameliorates High-Fat/Cholesterol-Diet-Induced Atherosclerosis by Modulating the NLRP3 Inflammasome Pathway in ApoE-/- Mice. J. Agric. Food Chem. 2018, 66, 4827–4834. [Google Scholar] [CrossRef]
- Wallert, M.; Börmel, L.; Lorkowski, S. Inflammatory Diseases and Vitamin E—What Do We Know and Where Do We Go? Mol. Nutr. Food Res. 2021, 65, 2000097. [Google Scholar] [CrossRef] [PubMed]
- Tapia, G.; Silva, D.; Romero, N.; Pettinelli, P.; Dossi, C.G.; de Miguel, M.; González-Mañán, D. Role of dietary α- and γ-tocopherol from Rosa mosqueta oil in the prevention of alterations induced by high-fat diet in a murine model. Nutrition 2018, 53, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lim, Y. Gamma-tocopherol ameliorates hyperglycemia-induced hepatic inflammation associated with NLRP3 inflammasome in alloxan-induced diabetic mice. Nutr. Res. Pract. 2019, 13, 377–383. [Google Scholar] [CrossRef]
- Mullin, G.E.; Limektkai, B.; Wang, L.; Hanaway, P.; Marks, L.; Giovannucci, E. Dietary Supplements for COVID-19. In Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2021; Volume 1318, pp. 499–515. [Google Scholar]
- Sheng, K.; He, S.; Sun, M.; Zhang, G.; Kong, X.; Wang, J.; Wang, Y. Synbiotic supplementation containing: Bifidobacterium infantis and xylooligosaccharides alleviates dextran sulfate sodium-induced ulcerative colitis. Food Funct. 2020, 11, 3964–3974. [Google Scholar] [CrossRef]
- Suzuki, H.; Yamazaki, T.; Ohshio, K.; Sugamata, M.; Yoshikawa, M.; Kanauchi, O.; Morita, Y. A Specific Strain of Lactic Acid Bacteria, Lactobacillus paracasei, Inhibits Inflammasome Activation In Vitro and Prevents Inflammation-Related Disorders. J. Immunol. 2020, 205, 811–821. [Google Scholar] [CrossRef]
- Pan, X.; Fang, X.; Wang, F.; Li, H.; Niu, W.; Liang, W.; Wu, C.; Li, J.; Tu, X.; Pan, L.L.; et al. Butyrate ameliorates caerulein-induced acute pancreatitis and associated intestinal injury by tissue-specific mechanisms. Br. J. Pharmacol. 2019, 176, 4446–4461. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.C.; Ouyang, C.N.; Yuan, S.N.; Lin, H.C.; Huang, K.Y.; Wu, P.S.; Liu, C.Y.; Tsai, K.J.; Loi, L.K.; Chen, Y.J.; et al. Pretreatment with a heat-killed probiotic modulates the NLRP3 inflammasome and attenuates colitis-associated colorectal cancer in mice. Nutrients 2019, 11, 516. [Google Scholar] [CrossRef] [Green Version]
- Loss, H.; Aschenbach, J.R.; Ebner, F.; Tedin, K.; Lodemann, U. Effects of a pathogenic ETEC strain and a probiotic Enterococcus faecium strain on the inflammasome response in porcine dendritic cells. Vet. Immunol. Immunopathol. 2018, 203, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.J.; Xu, J.J.; Wang, X.; Zhu, Y.H.; Wu, Q.; Wang, J.F. Lactobacillus johnsonii l531 ameliorates Escherichia coli-induced cell damage via inhibiting NLRP3 inflammasome activity and promoting ATG5/ATG16L1-mediated autophagy in porcine mammary epithelial cells. Vet. Sci. 2020, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, Y.; Chu, B.; Yuan, L.; Liu, N.; Zhu, Y.; Wang, J. Lactobacillus johnsonii l531 alleviates the damage caused by Salmonella typhimurium via inhibiting tlr4, nf-κb, and nlrp3 inflammasome signaling pathways. Microorganisms 2021, 9, 1983. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, Y.; Chu, B.; Liu, N.; Chen, S.; Wang, J. Lactobacillus rhamnosus GR-1 Prevents Escherichia coli-Induced Apoptosis Through PINK1/Parkin-Mediated Mitophagy in Bovine Mastitis. Front. Immunol. 2021, 12, 3743. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive compounds: Definition and assessment of activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, F.; Wang, M. Bioactive Substances of Animal Origin. In Handbook of Food Chemistry; Springer: Berlin, Germany, 2015; pp. 1009–1033. ISBN 9783642366055. [Google Scholar]
- Zhao, Y.; Wu, Y.Z.; Wang, M. Bioactive Substances of Plant Origin. In Handbook of Food Chemistry; Springer: Berlin, Germany, 2015; pp. 967–1008. ISBN 9783642366055. [Google Scholar]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Ditu, L.M.; Grigore, M.E.; Camen-Comanescu, P.; Holban, A.M. Introduction in Nutraceutical and Medicinal Foods. In Therapeutic, Probiotic, and Unconventional Foods; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–12. ISBN 9780128146262. [Google Scholar]
- Hung, W.L.; Ho, C.T.; Pan, M.H. Targeting the NLRP3 Inflammasome in Neuroinflammation: Health Promoting Effects of Dietary Phytochemicals in Neurological Disorders. Mol. Nutr. Food Res. 2020, 64, 1900550. [Google Scholar] [CrossRef]
- Bagherniya, M.; Khedmatgozar, H.; Fakheran, O.; Xu, S.; Johnston, T.P.; Sahebkar, A. Medicinal plants and bioactive natural products as inhibitors of NLRP3 inflammasome. Phyther. Res. 2021, 35, 4804–4833. [Google Scholar] [CrossRef]
- Oliviero, F.; Scanu, A.; Zamudio-Cuevas, Y.; Punzi, L.; Spinella, P. Anti-inflammatory effects of polyphenols in arthritis. J. Sci. Food Agric. 2018, 98, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.Q.; Pan, R.; Tang, Y.; Zhou, X.G.; Wu, J.M.; Yu, L.; Law, B.Y.K.; Ai, W.; Yu, C.L.; Qin, D.L.; et al. Lychee seed polyphenol inhibits Aβ-induced activation of NLRP3 inflammasome via the LRP1/AMPK mediated autophagy induction. Biomed. Pharmacother. 2020, 130, 110575. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth-Walter, F.; Moskovskich, A.; Gomez-Casado, C.; Diaz-Perales, A.; Oida, K.; Singer, J.; Kinaciyan, T.; Fuchs, H.C.; Jensen-Jarolim, E. Immune suppressive effect of cinnamaldehyde due to inhibition of proliferation and induction of apoptosis in immune cells: Implications in cancer. PLoS ONE 2014, 9, e108402. [Google Scholar] [CrossRef] [PubMed]
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic compounds in fruits—An overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Hollman, P.C.H.; Katan, M.B. Dietary flavonoids: Intake, health effects and bioavailability. Food Chem. Toxicol. 1999, 37, 937–942. [Google Scholar] [CrossRef]
- Ottaviani, J.I.; Momma, T.Y.; Heiss, C.; Kwik-Uribe, C.; Schroeter, H.; Keen, C.L. The stereochemical configuration of flavanols influences the level and metabolism of flavanols in humans and their biological activity in vivo. Free Radic. Biol. Med. 2011, 50, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S.; Williamson, G. Interactions affecting the bioavailability of dietary polyphenols in vivo. Int. J. Vitam. Nutr. Res. 2007, 77, 224–235. [Google Scholar] [CrossRef]
- Di, L.; Kerns, E.H. Drug-Like Properties: Concepts, Structure Design and Methods from ADME to Toxicity Optimization; Academic Press: Cambridge, MA, USA, 2016; ISBN 9780128010761. [Google Scholar]
- Giada, M.D.L.R. Food Phenolic Compounds: Main Classes, Sources and Their Antioxidant Power. In Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Ruiz-Rodriguez, A.; Marín, F.R.; Ocaña, A.; Soler-Rivas, C. Effect of domestic processing on bioactive compounds. Phytochem. Rev. 2008, 7, 345–384. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Medina-Remón, A.; Pérez-Jiménez, J.; Neveu, V.; Knaze, V.; Slimani, N.; Scalbert, A. Effects of food processing on polyphenol contents: A systematic analysis using Phenol-Explorer data. Mol. Nutr. Food Res. 2015, 59, 160–170. [Google Scholar] [CrossRef]
- Al-juhaimi, F.; Ghafoor, K.; Özcan, M.M.; Jahurul, M.H.A.; Babiker, E.E.; Jinap, S.; Sahena, F.; Sharifudin, M.S.; Zaidul, I.S.M. Effect of various food processing and handling methods on preservation of natural antioxidants in fruits and vegetables. J. Food Sci. Technol. 2018, 55, 3872–3880. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The total antioxidant content of more than 3100 foods, beverages, spices, herbs and supplements used worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Gul, K.; Singh, A.K.; Jabeen, R. Nutraceuticals and Functional Foods: The Foods for the Future World. Crit. Rev. Food Sci. Nutr. 2016, 56, 2617–2627. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, D.; von Brugger, J.; Bialow, S. Functional food science: Differences and similarities with food science. Funct. Foods Health Dis. 2021, 11, 408–430. [Google Scholar] [CrossRef]
- Vettorazzi, A.; de Cerain, A.L.; Sanz-Serrano, J.; Gil, A.G.; Azqueta, A. European regulatory framework and safety assessment of food-related bioactive compounds. Nutrients 2020, 12, 613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellumori, M.; Cecchi, L.; Innocenti, M.; Clodoveo, M.L.; Corbo, F.; Mulinacci, N. The EFSA health claim on olive oil polyphenols: Acid hydrolysis validation and total hydroxytyrosol and tyrosol determination in Italian virgin olive oils. Molecules 2019, 24, 2179. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.W.; Li, C.I.; Huang, K.C.; Liu, C.S.; Chen, H.L.; Lee, C.C.; Chiou, Y.Y.; Chen, R.J. 3-MCPD and glycidol coexposure induces systemic toxicity and synergistic nephrotoxicity via NLRP3 inflammasome activation, necroptosis, and autophagic cell death. J. Hazard. Mater. 2021, 405, 124241. [Google Scholar] [CrossRef]
- Jia, X.; Qiu, T.; Yao, X.; Jiang, L.; Wang, N.; Wei, S.; Tao, Y.; Pei, P.; Wang, Z.; Zhang, J.; et al. Arsenic induces hepatic insulin resistance via mtROS-NLRP3 inflammasome pathway. J. Hazard. Mater. 2020, 399, 123034. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Cai, D.; Li, P.; Jin, J.; Jiang, X.; Li, Z.; Tian, L.; Chen, G.; Sun, J.; et al. Chronic oral exposure to cadmium causes liver inflammation by NLRP3 inflammasome activation in pubertal mice. Food Chem. Toxicol. 2021, 148, 111944. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Yan, D.; Wang, Y.; Wang, N.; Liu, Y.; Tan, A.; Chen, X.; Yan, H. Chronic acrylamide exposure induced glia cell activation, NLRP3 inflammasome upregulation and cognitive impairment. Toxicol. Appl. Pharmacol. 2020, 393, 114949. [Google Scholar] [CrossRef]
- Bo, N.; Yilin, H.; Chaoyue, Y.; Lu, L.; Yuan, Y. Acrylamide induces NLRP3 inflammasome activation via oxidative stress- and endoplasmic reticulum stress-mediated MAPK pathway in HepG2 cells. Food Chem. Toxicol. 2020, 145, 111679. [Google Scholar] [CrossRef]
- Lee, P.Y.; Liu, C.C.; Wang, S.C.; Chen, K.Y.; Lin, T.C.; Liu, P.L.; Chiu, C.C.; Chen, I.C.; Lai, Y.H.; Cheng, W.C.; et al. Mycotoxin zearalenone attenuates innate immune responses and suppresses nlrp3 inflammasome activation in lps-activated macrophages. Toxins 2021, 13, 593. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Shi, L.; Yao, K.; Sun, X.; Yang, G.; Jiang, L.; Zhang, C.; Wang, N.; Zhang, H.; et al. Zearalenone induces NLRP3-dependent pyroptosis via activation of NF-κB modulated by autophagy in INS-1 cells. Toxicology 2019, 428, 152304. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Wang, S.; Jiang, L.; Jiao, Y.; Sun, X.; Li, J.; Yang, L.; Hou, Y.; Wang, N.; Yao, X.; et al. Patulin induces pyroptosis through the autophagic-inflammasomal pathway in liver. Food Chem. Toxicol. 2021, 147, 111867. [Google Scholar] [CrossRef] [PubMed]
- Molagoda, I.M.N.; Lee, S.; Jayasooriya, R.G.P.T.; Jin, C.Y.; Choi, Y.H.; Kim, G.Y. Deoxynivalenol enhances IL-1ß expression in bv 2. Microglial cells through activation of the NF-ΚB pathway and the ASC/NLRP 3. inflammasome. EXCLI J. 2019, 18, 356–369. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Chemical Class | Compound | Food Source | Experimental Model | Dose | Treatment (Duration) | Main Outcome | References |
|---|---|---|---|---|---|---|---|
| Phenolic acids | Sinapic acid | Vegetables, spices, fruits, cereals, oilseed wine, vinegar | Male Kunming mice (induced colitis) | 10, 50 mg/kg | 7 days | Dose-dependent reduction in the NLRP3 inflammasome proteins expression. | [57] |
| Ferulic acid | Fruits, vegetables, grains, beans, seeds, nuts, grasses, corn, wheat, turmeric | Male Wistar rat (induced kidney injury) | 25, 50 mg/kg | 15 days | Both doses of ferulic acid administration have produced an amelioration of NLRP3 and caspase-1 proteins expression. | [58] | |
| Chlorogenic acid | Fruit, coffee beans, eggplants | 1. Male and female BALB/c mice (colon tissue) | 1. 20, 40 mg/kg | 1. 7 days | 1. Decrease in NLRP3, ASC, caspase-1 p45, and caspase-1 p20 protein levels. | [59] | |
| 2. RAW264.7 cells | 2. Many concentrations of chlorogenic acid | 2. Not reported | 2. Decrease secretion of IL-1𝛽 and IL-18. | ||||
| Flavones and flavonones | Apigenin | Parsley, onions, oranges, tea | 1. Male C57BL/6 J mice (induced NAFLD) | 1. 50 mg/kg of BW per day | 1. 7 days | 1., 2.: Reduction in NLRP3, ASC, pro-caspase-1, caspase-1, together with a reduction in ROS production. 3. Reduced NLRP3 protein expression. | 1 and 2. [60] 3. [61] |
| 2. Hepa1–6 cells | 2. 16 and 32 μM | 2. 24 h | |||||
| 3. ISO-HAS human endothelial cells | 3. 30 and 50 μM | 3. 24 h | |||||
| Isoorientin | Gentiana | 1. Male ICR mice (induced hyperuricemia) | 1. 5 and 10 mg/kg of BW | 1. Single dose | Dose-dependent inhibition of xanthine oxidase activity and interleukin release. | [62] | |
| 2. MXC207 cells | 2. 25, 50, 100, 200, and 400 µM | 2. 24 h | |||||
| Chrysin | Honey, propolis, carrots, chamomile, fruits, mushrooms | Male Sprague Dawley rats (induced hyperuricemia) | 50, 100, and 150 mg/kg of BW | 4 weeks | Reduction in IL-1β expression and ROS activity. | [63] | |
| Luteolin | Chamomile, carrots, olive oil, species | Male Sprague Dawley rats (induced SCII) | 50 and 100 mg/kg of BW | 14 days | Reduction in NLRP3, IL-1β, and IL-18 expression. | [64] | |
| Hesperidin methylchalcone | Citrus | Swiss mice (induced gout arthritis) | 30 mg/kg of BW | Single dose | Reduction in NLRP3, ASC, pro-caspase-1, and pro-IL-1β mRNA expression. | [65] | |
| Naringin | Citrus | Male C57BL/6 mice (induced ulcerative colitis) | 25, 50, and 100 mg/kg of BW | 7 days | Dose-dependent reduction in NLRP3, ASC, caspase-1, and IL-1β expression. | [66] | |
| Flavonols | Quercetin | Fruits, vegetables, seeds, grains | 1. Male SPF-Wistar rats (induced liver injury) | 1. 100 mg/kg of BW | 1. 14 days | 1. Reduction in ROS, NF-κB, NLRP3, IL-1β, and IL-18 expression. | 1. [67] |
| 2. ApoE-/- mice (induced atherosclerotic inflammation) | 2. 100 mg/kg of BW | 2. 16 days | 2. Decrease in pro-IL-1β and IL-1β production. | 2. [68] | |||
| 3. Senescence accelerated mouse P8 and R1 | 3. 35 and 70 mg/kg of BW | 3. 4 weeks | 3. Increase in cognitive functions and reduction in NLRP3 activation factors expression. | 3. [69] | |||
| 4. Caco-2 cell triggered with E.coli | 4. 200 µM | 4. 12 h | 4. Reduction in NLRP3, caspase-1, and IL-1β expression, together with an enhanced ROS scavenger activity. | 4. [70] | |||
| Dihydroquercetin | Onions, milk thistle, Douglas fir bark | 1. Male C57BL/6 mice (induced liver steatosis) | 1. 1, 5, and 25 mg/kg of BW | 1. Single dose | Inhibition of NLRP3, caspase-1 cleavage, and IL-1β production. | [71] | |
| 2. Human hepatoma cells HepG2 | 2. 6.25, 25, and 100 µM | 2. 1 h | |||||
| Kaempferol | Tea, vegetables, oranges, wine | Male C57BL/6 mice (induced hepatotoxicity) | 30 and 60 mg/kg of BW | 7 days | Dose-dependent reduction in IL-1β, TNF-α, and IL-6 expression. | [72] | |
| Myricetin | Vegetables, fruits, nuts, berries, tea, red wine | Male Wistar rats (induced diabetes) | 20 mg/kg of BW | 4 weeks | Reduced NLRP3 inflammasome activation. | [73] | |
| Other phenolics | Curcumin | Turmeric | 1. Male Sprague Dawley rats (chronic unpredictable mild stress) | 1. 100 mg/kg of BW | 1. 4 weeks | 1. Reduction in IL-1β, IL-6 and TNF-α expression. | 1. [74] |
| 2. Hemodialysis patients | 2. 2.5 g of turmeric (95% curcumin) after dialysis | 2. 12 weeks | 2. Lower expression levels of NLRP3 inflammasome markers (NF-kB, NLRP3 and IL-1β. | 2. [75] | |||
| 6-shogaol | Ginger roots | 1. Human artery smooth muscle cells (induced calcification) | 1. Not indicated | 1. Not indicated | 1. Reduction in NLRP3, caspase-1 and IL-1β expression. | 1. [76] | |
| 2. Human THP-1 monocytes | 2. 5, 10, 20 and 40 µM | 2. 1 h | 2. Reduction in the canonical NLRP3 inflammasome-mediated IL-1β secretion. | 2. [77] | |||
| Pterostilbene | Grapes, blueberries | 1. Female BALB/c mice (induced acute liver failure) | 1. 50 mg/kg/12 h of BW | 1. 24 h | 1., 2.: Decrease in IL-1β, IL-6, caspase-1, TNF-α, and NLRP3 protein expression. 3. Increased autophagy, resulting in a decrease in NLRP3 and caspase-1. | 1. [78] | |
| 2. Female C57BL/6 mice (allergic contact dermatitis) | 2. 500 mg/kg of BW | 2. 5 weeks | 2. [79] | ||||
| 3. NRK-52E cells | 3. 2 µM | 3. 24, 48, and 72 h | 3. [80] | ||||
| Polydatin | Grapes juice | 1. Male Sprague Dawley rats (induced hyperuricemia) | 1. 25 and 50 mg/kg of BW | 1. 7 days | 1. Dose-dependent decrease in IL-1β, TNF-α, IL-6, NLRP3, and caspase-1. | 1. [81] 2 and 3. [82] | |
| 2. Male Wistar rats (induced dry-eye disease) | 2. 0.05 and 0.5% ocular solution | 2. 4 days | 2. Dose-dependent decrease in IL-1β, IFN-α, TNF-α, and IL-6. | ||||
| 3. Human conjunctival cell line HCC | 3. 0.1, 1 and 10 µM | 3. 8 h | 3. Dose-dependent decrease in NLRP3 and caspase-1. | ||||
| Cyanidin-3-O-β-glucoside | Red-violet fruits | 1. Human retinal pigment epithelium cells, ARPE-19 | 1. 50 and 100 µM | 1. 2 h | 1. Dose-dependent decrease in NLRP3, IL-18, IL-β and caspase-1. | 1. [83] | |
| 2. Male C57BL/6 J mice (induced hepatic inflammation) | 2. 200 mg/kg of BW | 2. 8 weeks | 2. Decrease in NLRP3, IL-18, IL-1β, and caspase-1 expression, together with the block of NF-κB signaling pathway. | 2. [84] | |||
| Epigallocatechin-3-gallate | Green tea | 1. Male Balb/C mice (induced acute pancreatitis) 2. Balb/C adherent cells 3. APP/PS1 transgenic mice 4. Mouse microglial cell line BV2 | 1. 5, 10, 20, 40, and 80 mg/kg of BW 2. 2.5, 5, and 10 µM 3. 2 mg/kg of BW 4. 10 µM | 1. 4 weeks 2. 24 h 3. 4 weeks 4. 1 h | 1. Dose-dependent decrease in IL-1β. 2. Dose-dependent decrease in caspase-1, IL-1β, and ROS. 3., 4.: Suppressed activation of NLRP3 inflammasome via TLR4/NF-κB pathway. | 1. and 2. [85] 3. and 4. [86] | |
| Resveratrol (encapsulated in poly(lactic-co-glycolic acid) nanoparticles) | Grapes, blueberries, raspberries, mulberries, peanuts | 1. Male C57BL/6 mice (induced kidney injury) 2. Epithelial cell line HK-2 | 1. 2 and 4 mg/kg 2. 25, 50, and 100 µM | 1. 2 weeks 2. 24 h | 1., 2.: Dose-dependent reduction in NLRP3, pro-caspase-1, cleaved-caspase-1, and IL-1β expression. | 1, and 2. [87] | |
| Polyphenols mixture | 1. Fermented non-digestible fraction of baked corn and common bean snacks (FNDF) 2. FNDF pure components (include gallic acid) | Zea mays L., Phaseolus vulgaris L. | Caco-2 cells, THP-1 cells | 1. 40, 200, and 300 µg/mL 2. Gallic acid 38.85 µM | 48 h | Inhibition of the NLRP3 assemblage, decreasing caspase-1 activity, IL-1β, and apoptosis. | [88] |
| 1. Green tea polyphenols (GTPs) 2. Epigallocatechin- 3-gallate (EGCG) | Green tea | Male ICR mice (liver tissue) | 1. 0.32% GTPs (w/v) 2. 0.32% EGCG (w/v) | 12 days | Downregulation of NLRP3, ASC, caspase-1, and IL-1β proteins expression. | [89] | |
| Green tea polyphenols (GTPs) | Green tea | Male ICR mice (liver tissue) | 100, 200 mg/kg of BW | 7 days | Inhibition of NLRP3, ASC, and caspase-1 (p20) expression in a dose-dependent manner. | [90] | |
| Soy isoflavones | Soybeans | Male C57BL/6 mice (colon tissue) | 50, 100 mg/kg of BW | 5 days | Reduction in NLRP3, Caspase-1 p20 and ASC protein levels and suppression of IL-1β and IL-18 secretion. | [91] | |
| Red raspberry polyphenols | Rubus idaeus L. | 1. Male C57BL/6 mice (adipose tissue) 2. C3H10T1/2 cells | 1. 120 mg/kg of BW per day 2. 10 μg mL−1 | 1. 16 weeks 2. 2 days | 1. Attenuation of NLRP3 inflammasome activation in adipose tissue macrophages and epididymal white adipose tissue. 2. Reducing of IL-1β, IL-18, and NLRP3 protein levels expression. | [92] | |
| Organosulfur compounds | Allicin | Garlic | 1. Male Sprague Dawley rats (acrylamide treated) 2. Kupffer cells (BNCC341160) | 1. 25 and 50 mg/kg of BW 2. 3.75, 7.5, and 15 μM | 1. 4 weeks 2. 2 h | 1., 2.: Reduced NLRP3 inflammasome activation, decreasing cleaved-caspase-1, IL-1β, IL-18, IL-6, and TNF-α secretion. | 1. and 2. [93] |
| Benzyl isothiocyanate | Cruciferous vegetables | 1. Male C57BL/6 J mice (induced nonalcoholic steatohepatitis) 2. Mice Kupffer cells | 1. 1 g/kg of BW 2. 2.5 and 5.0 μM | 1. 9 weeks 2. 4 h | 1., 2.: Reduction in NLRP3, p20 caspase-1, and IL1-β expression. | 1. and 2. [94] | |
| Sulforaphane | Cruciferous vegetables | BALB/c mice (induced pancreatic injury) | 5 mg/kg of BW | 3 days | Reduced expression of NLRP3, p20 caspase-1, and IL1-β. | [95] | |
| Methylsulfonylmethane | Garlic | Bone marrow-derived macrophages | 0.3, 0.5, 1.0, 2.0, 4.0, and 8.0% | 6 h | Blocking the NF-κB signaling and pro IL1-β expression. | [96] | |
| Terpenes and terpenoids | Carnosic acid | Rosmarinus and Salvia | Male Balb/c mice (induced acute colitis) | 50 or 100 mg/kg of BW | 10 days | Reduced release of caspase-2 and ROS. | [97] |
| Geranylgeraniol | Flax, sunflower, and olive oils | Daoy cells | 50 μM | 24 h | Inhibition of NLRP3 gene expression. | [98] | |
| Kaurenoic Acid | X. aethiopica fruits | BALB/c mice macrophages | 10, 30, 50, 70, and 90 μM | 24 h | Increased the production of NO and IL-1β. | [99] | |
| Fatty acids | Saturated fatty acids | Palmitate acid | LPS-primed bone marrow-derived macrophages generated from wild-type (WT), Nlrp3−/−, Pycard−/−, or Nlrc4−/− | 200 or 500 μM | 24 h | Inhibition of AMP-activated protein kinase followed by the accumulation of mitochondrial ROS, thus activating the NLRP3-ASC inflammasome and causing caspase-1, IL-1β, and IL-18 increased production. | [16] |
| Stearate acid | LPS-primed bone marrow-derived dendritic cells | 250 μM | 5, 20 h | NLRP3 inflammasome activation mediated by IRE1α activation (inositol-requiring enzyme 1-α) through the saturated phosphatidylcholine accumulation. | [100] | ||
| Virgin coconut oil | Male Wistar rats AD (receiving Amyloid-β) and high-fat diet (HFD) models both in vitro and in vivo | 8 and 10% | 8 weeks | Reduction in IL-1β protein, caspase-1, and NLRP3 genes expression. | [101] | ||
| PUFA | Fish oil | Obesity male Wistar rat models | Intragastrically 1 mL/kg per day | 8 weeks | Reduction in IL-1β protein, caspase-1, and NLRP3 genes expression. | [102] | |
| Safflower oil | A. Fat-1 transgenic mice generated onto a C57BL/6 background B. Wild-type mice | 10 g safflower oil (per 100 g of diet) | 80 days | Caspase-1, IL-1β, and IL-18 reduction. Blockade of high glucose-induced TXNIP via the PI3K/Akt pathway in pre-adipocytes. | [103] | ||
| Walnut oil | DSS-induced colitis in Kunming (KM) male mice | 2.5 mL/kg⋅d Walnut oil | 27 days | inhibition of ROS production, mediation of NLRP3/ASC/caspase-1 signaling pathway, regulation of gut microbiota and SCFAs levels. | [104] | ||
| DHA | A. ASH-associated mouse LPS-induced Kupffer cell lines B. C57BL/6 J mice | 1. 50 μM 2. Intragastrally DHA 50 mg/kg once per day | 1. 4 h 2. 7 days | Mechanisms through which ω-3 PUFAs reduce metabolic inflammation may include the G protein-coupled receptor 120 (GPR120) and GPR40 which interact with NLRP3 and inhibited the NLRP3 inflammasome complex assembly. | [105] | ||
| Fish oil | Male Sprague Dawley rats (prefrontal cortex and hippocampus of rats) | 1.5 g/kg of BW | 3 weeks | Reduced MDA level and mRNA levels of pro-inflammatory cytokines 1 L-1β, IL-6. Mitigation of the LPS-induced P2X7R and NLRP3 activation, downregulated LPS-induced iNOS and NF-κB expression in both prefrontal cortex and hippocampus. | [106] | ||
| Proteins and amino acid derivatives | RDP2 | Oryza Sativa | Kunming mice (induced hyperuremia) | Allopurinolo:10 mg/kg of BW Benzbromaron: 8 mg/kg of BW RDP2: 5, 10, 100 μg/kg of BW | 7 days | 1. Reduction serum uric acid levels 2. Reduction in NLRP3, ASC, and caspase-1 expression in the kidneys. | [107] |
| RDP3 | Oryza Sativa | Kunming mice (induced hyperuremia) | Allopurinol:10 mg/kg of BWBenzbromaron: 8 mg/kg of BW RDP3: 100 μg/kg, 500 μg/kg and 1 mg/kg of BW | 7 days | 1. Serum uric acid concentrations in the RDP3 group were significantly lower than in the other treatments. 2. RDP3 reduced inflammation by inhibiting the expression of the NLRP3 inflammasome. | [108] | |
| TMOP (Tuna meat oligopeptides) | Tuna | ICR mice (induced hyperuremia) | 50 mg/kg and 300 mg/kg of BW | 8 weeks | Dose-dependent reduction in hyperuricaemia, due to the inhibition of NLRP3 inflammasome complex. | [109] | |
| Soy protein concentrate (SPC) | Soy | CF-1 mice (induced acute ulcerative colitis) | DSS (1.5% DSS in drinking fluid and AIN93G), DS6 (1.5% DSS and 6% dietary SPC), and DS12 (1.5% DSS and 12% dietary SPC) | 7 days | Prevent increased pro-inflammatory signaling and thereby moderate colitis severity. | [110] | |
| α-gliadin 31–43 | Gluten | C57BL/6 mice intestinal samples | 200 μl | 4/16 h | 1. Formation of oligomers that activate the inflammasome.2. Stimulation of IL-1β release. | [111] | |
| N-acetyl-cysteine (NAC) | Garlic, onions, and leeks | Raised and pregnant sows | 500 mg/kg of BW | From day 85 until delivery | Reduction maternal and placental inflammatory cytokines through inhibition of the NLRP3 inflammasome. | [112] | |
| Glycine | Fish, meat, spirulina algae, soy protein, egg whites | C57BL/6 male mice (induced lung injury) | 1000 mg in 5 mL of 0.9% saline | 7 days | Prevent mucin reduction and upregulation of pro-inflammatory cytokines. | [113] | |
| Carnosine | Fish and meat | SAMP8 mice | 100–200 mg/kg of BW | 6 weeks | Decreased levels of malondialdehyde and reactive oxygen species (ROS), increased activity of superoxide dismutase (SOD) and the level of adenosine triphosphate; NLRP3 inflammasome reduction. | [114] | |
| L-Homocarnosine | Meat | Ischemic albino Wistar rats | 0.5 mM and 1 mM | 45 days | Reduction in NLRP3 inflammasome levels to near normal levels. | [115] | |
| Choline | Eggs, meat, fish, legumes | APP/PS1 transgenic mice | 1.10 g/kg and 4.95 g/kg of BW | 9 mounths | Reduction Aβ deposition, microgliosis, and pro-inflammatory cytokine production. | [116] | |
| Saponins and sterols | 25-OCH3-PPD ginsenoside | Panax ginseng | C57BL/6 mice (induced TAA) | 5, 10, or 20 mg/kg of BW | 5 weeks | Reduction inflammation by regulating P2X7R-mediated NLRP-3 inflammasome. | [117] |
| Rh1 and Rg2 ginsenosides | Panax notoginseng | C57BL/6 mice (induced NAFLD) | 50 or 150 mg/kg of BW | 9 weeks | Inhibition NLRP3 inflammasome, promoting mitophagy, and reduction mtROS production. | [118] | |
| Ginsenoside compound K (CK) | Panax ginseng | Diabetic db/db and db/m mice | 10 mg/kg of BW | 12 weeks | Downregulation inflammatory cytokines and mediator production by suppressing the NLRP3 inflammasome pathway. | [119] | |
| Magnesium isoglycyrrhizinate | Glycyrrhiza glabra | Sprague Dawley mice | 10, 20, and 40 mg/kg of BW | 11 weeks | Inhibition the activation of the NF-κB/inflammasome NLRP3 and reduction the immunological–inflammatory response. | [120] | |
| Physalin B | Physalis alkekengi L. | BALB/c mice | 250 μL | 7 days | Reduction the pro-inflammatory cytokine levels, suppression the NF-κB cascade and pathway STAT3 and arrestin1 signaling, and inhibition NLRP3 inflammasome activation. | [121] | |
| β-sitosterol | Moringa oleifera | HaCaT keratinocytes and J774A.1 macrophages | 7.5 to 30 μM | 24 h | Significant reduction in NLRP3 expression, inhibition of caspase-1, and NF-KB activation in macrophages. | [122] | |
| Polysaccharides | Polysaccharide composed of galactose, glucose, mannose, and arabinose with molar ratios of 5.79:5.77:3.45:1.20 (average MW 63000) | Trametes orientalis | Male Kunming mice (induced lung injury) | Intragastrally 50, 100, 200 mg/kg of BW | 21 days | Suppression of IL-1β expression and blockage of NLRP3, ASC, and caspase-1 increases in a dose-dependent manner. | [123] |
| Polysaccharides extracted from Dendrobium officinale | Dendrobium officinale | 1. DSS male BalB/c mice (induced induced acute ulcerative colitis) 2. LPS-stimulated NCM460 cells | 1. 50, 100, and 200 mg/kg of BW 2. 50, 100, and 200 mg/mL | 1. 7 days 2. 24 h | Suppression of NLRP3, ASC, caspase-1, IL-1β, and IL-18 mRNA expression. | [124] | |
| CYP-1 mannoglucan | Chinese yam | 1. In vitro RAW 264.7 murine macrophage cells 2. In vivo DDS-induced colitis male C57BL/6 J mice | 1. 31.25, 62.5, 125, 250, and 500 μg/mL 2. Intragastrically 300 mg/kg of BW | 1. 24 h 2. 7 days | Suppress the expression of several key genes involved in colonic inflammatory signaling pathways (such as NF-κB and NLRP3). | [125] | |
| Ganoderma lucidum polysaccharides | Ganoderma lucidum | Male Kun-Ming mice | 50, 100, 150 mg/kg of BW | 7 days | Decreased protein expression levels of NLRP3, ASC, and caspase-1 in liver tissue. | [126] | |
| Mannose, arabinose, and fucose at a molar ratio of 1.6:1.0:2.7 | Armillariella tabescens | Male C57BL/6 J mice | 100, 200, and 400 mg/kg Per day | 4 weeks | Reduction in MDA, pro-inflammatory factors (TNF-α, IL-18, and IL-1β) and FAS, G6Pase, and PEPCK levels in a dose-dependent manner. Decreased TXNIP and NLRP3 expression levels. | [127] | |
| Low methoxyl pectin | 4-week-old female non-obese diabetic (NOD) mice | Diets with 5% (wt/wt) LMP | 1. 36 weeks 2. 18 weeks | Suppression of NLRP3 and associated proteins expression (NLRP3, caspase-1-p20, cleaved IL-1β, and cleaved IL-18) in cecum; increase in the SCFAs (short chain fatty acids) by gut microbiota. | 1. [128] 2. [129] | ||
| Non-digestible carbohydrates (NDCs) consisting of pectic homogalacturonan and highly branched rhamnogalacturonan-II, as well as hemicellulosic material including glucomannan, xyloglucan, and glucurono(arabino)xylan | Chayote fruit | Human THP-1 macrophage-like cells Human monocytic cell line THP-1 | 100, 200, and 400 μg/mL | 24 h | Inhibition of CC-induced active caspase-1 (400 μg/mL), reduction in ROS accumulation and IL-1β. mRNA expression of IL-1β and NLRP3 in macrophage-like cells. Inhibition of NLRP3 and IL-1β gene expression in both CC-pretreated macrophage-like cells LPS-induced cells. | [130] | |
| Mannoglucan sulfate SF-2 | Starfish (A. rollestoni) | 1. RAW 264.7 cells murine macrophages 2. Primary peritoneal macrophages isolated form male ICR mice 3. ICR mice | 1. and 2. 80 μg/mL 3. 30 and 60 mg/kg of BW | 1. and 2. 0 h, 0.5 h, 1 h, 3 h, 6 h, and 9 h. C. 14 days | Improved release of cytokines and NLRP3 expression by the elevated expression of NLRP3, cleaved caspase-1, and ASC proteins. | [131] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spano, M.; Di Matteo, G.; Ingallina, C.; Ambroselli, D.; Carradori, S.; Gallorini, M.; Giusti, A.M.; Salvo, A.; Grosso, M.; Mannina, L. Modulatory Properties of Food and Nutraceutical Components Targeting NLRP3 Inflammasome Activation. Nutrients 2022, 14, 490. https://doi.org/10.3390/nu14030490
Spano M, Di Matteo G, Ingallina C, Ambroselli D, Carradori S, Gallorini M, Giusti AM, Salvo A, Grosso M, Mannina L. Modulatory Properties of Food and Nutraceutical Components Targeting NLRP3 Inflammasome Activation. Nutrients. 2022; 14(3):490. https://doi.org/10.3390/nu14030490
Chicago/Turabian StyleSpano, Mattia, Giacomo Di Matteo, Cinzia Ingallina, Donatella Ambroselli, Simone Carradori, Marialucia Gallorini, Anna Maria Giusti, Andrea Salvo, Michela Grosso, and Luisa Mannina. 2022. "Modulatory Properties of Food and Nutraceutical Components Targeting NLRP3 Inflammasome Activation" Nutrients 14, no. 3: 490. https://doi.org/10.3390/nu14030490