
Inflammatory Markers in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D Metabolites


Abstract
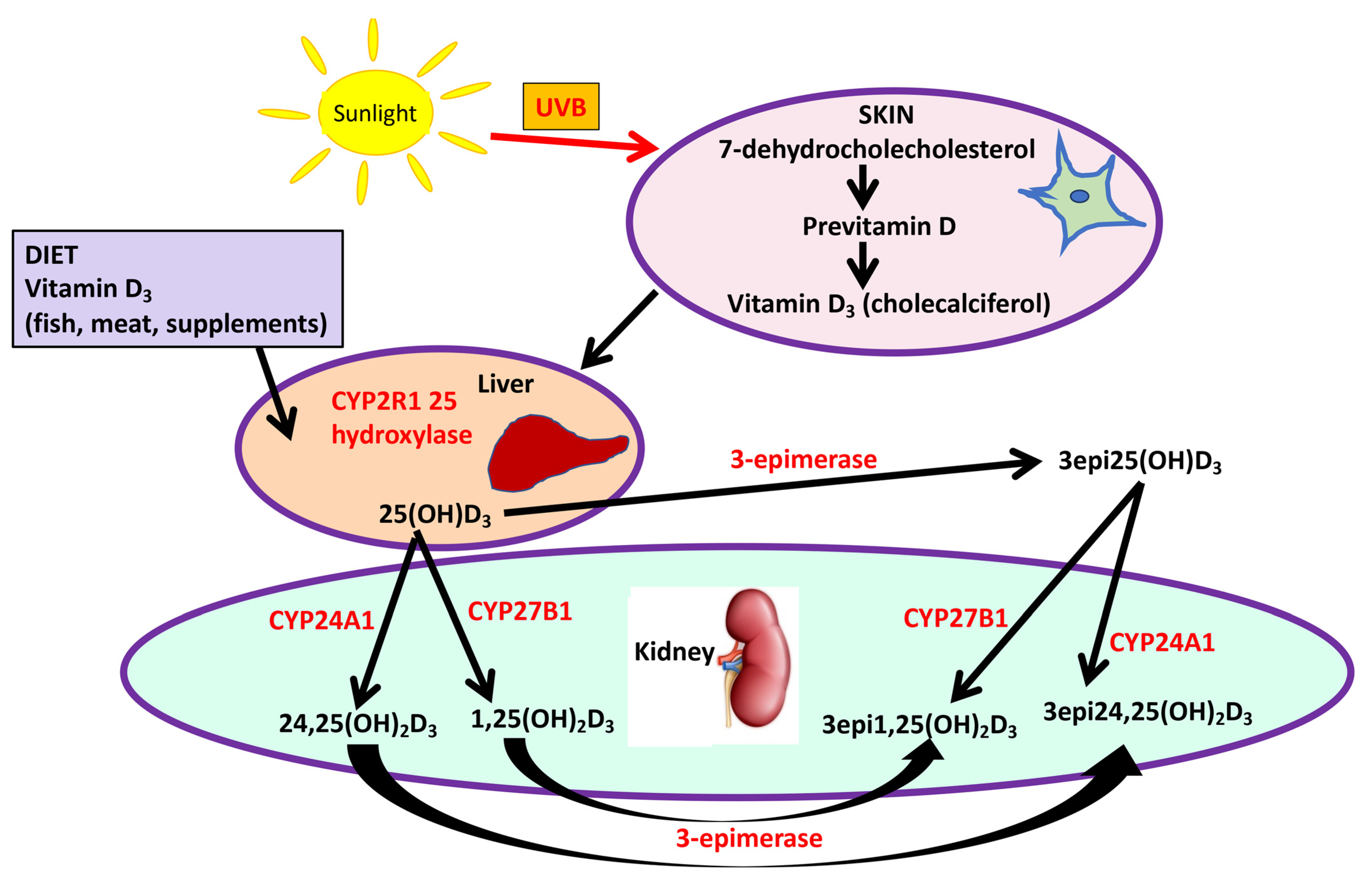
:1. Introduction
2. Materials and Methods
3. Statistics
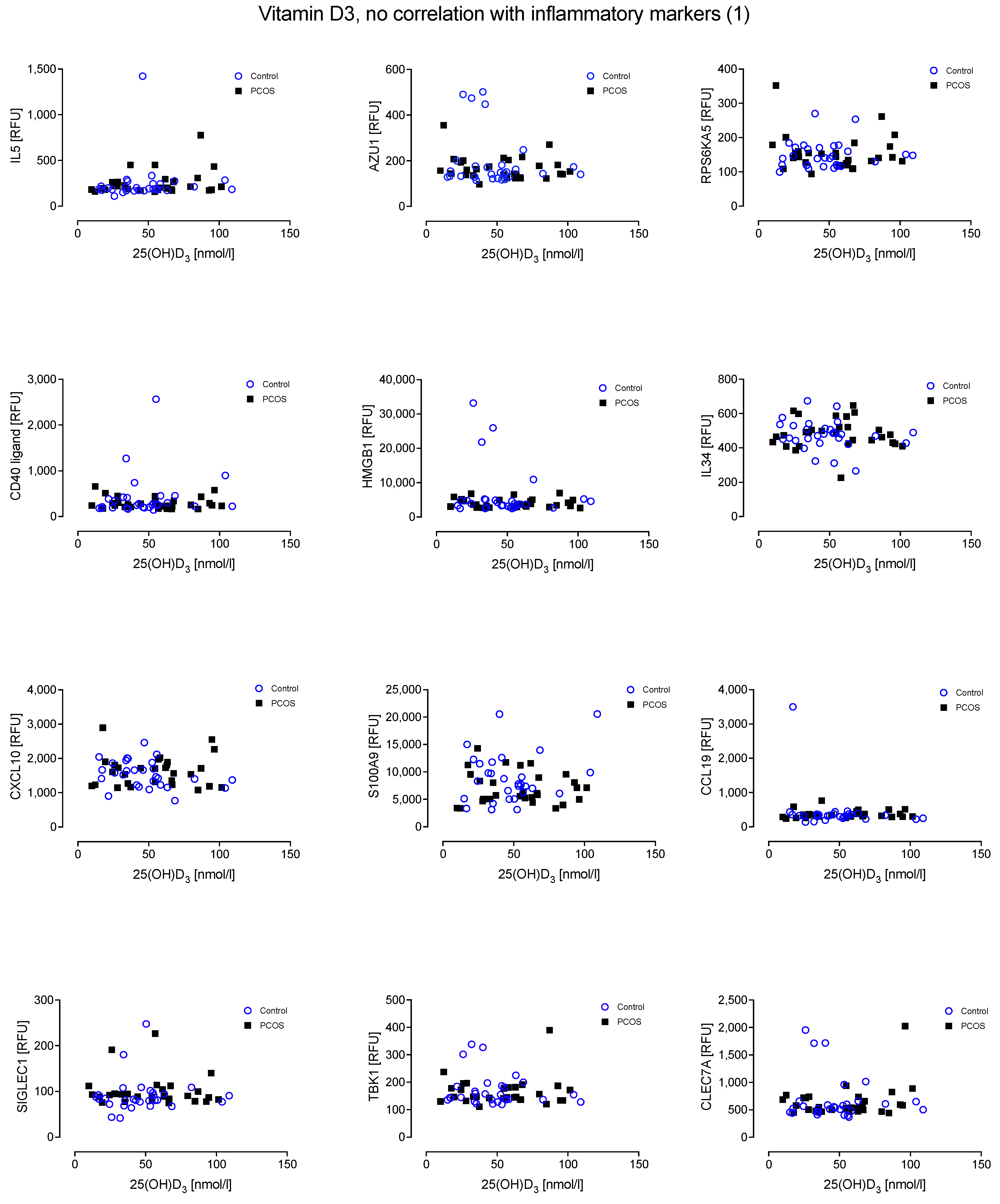
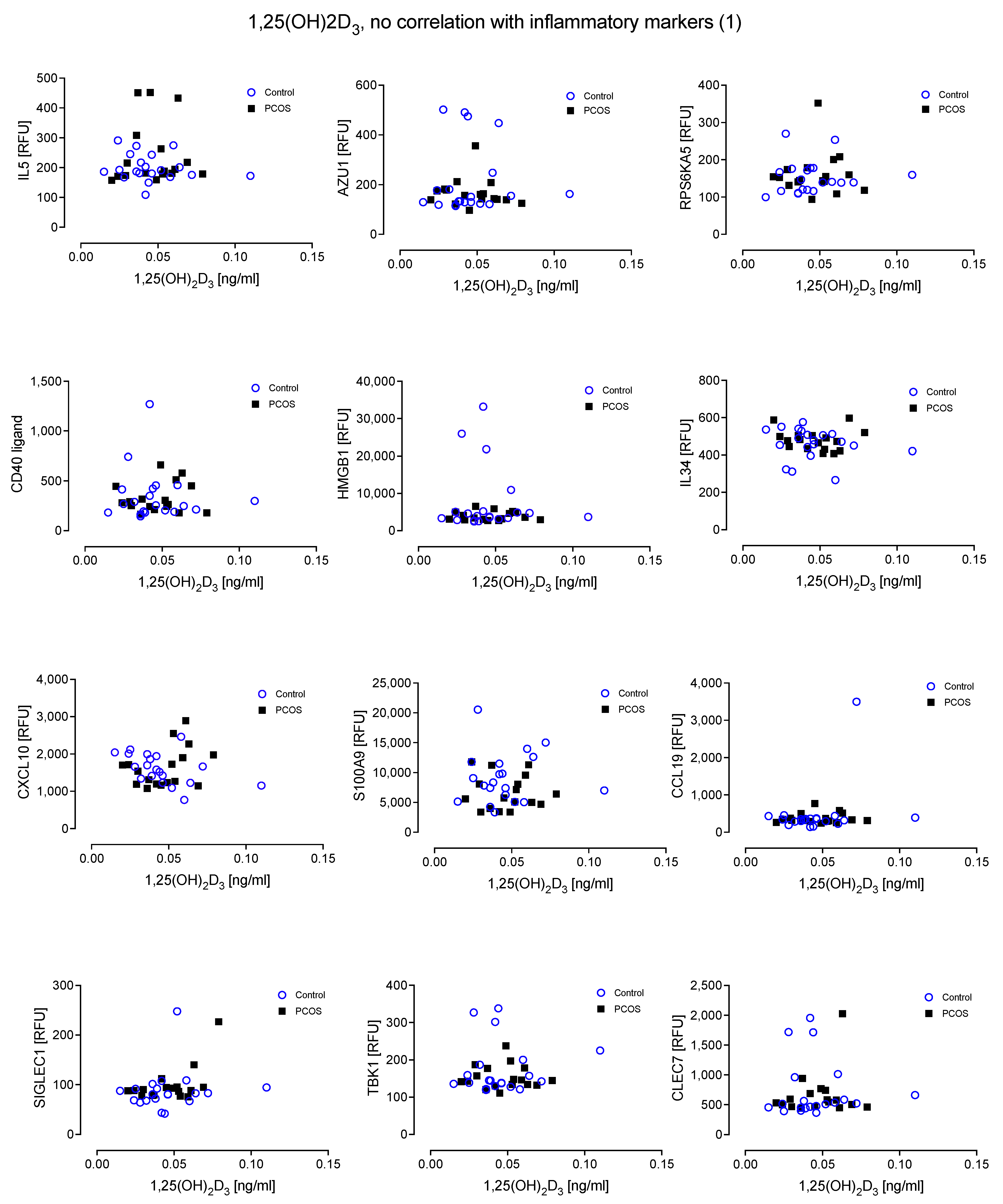
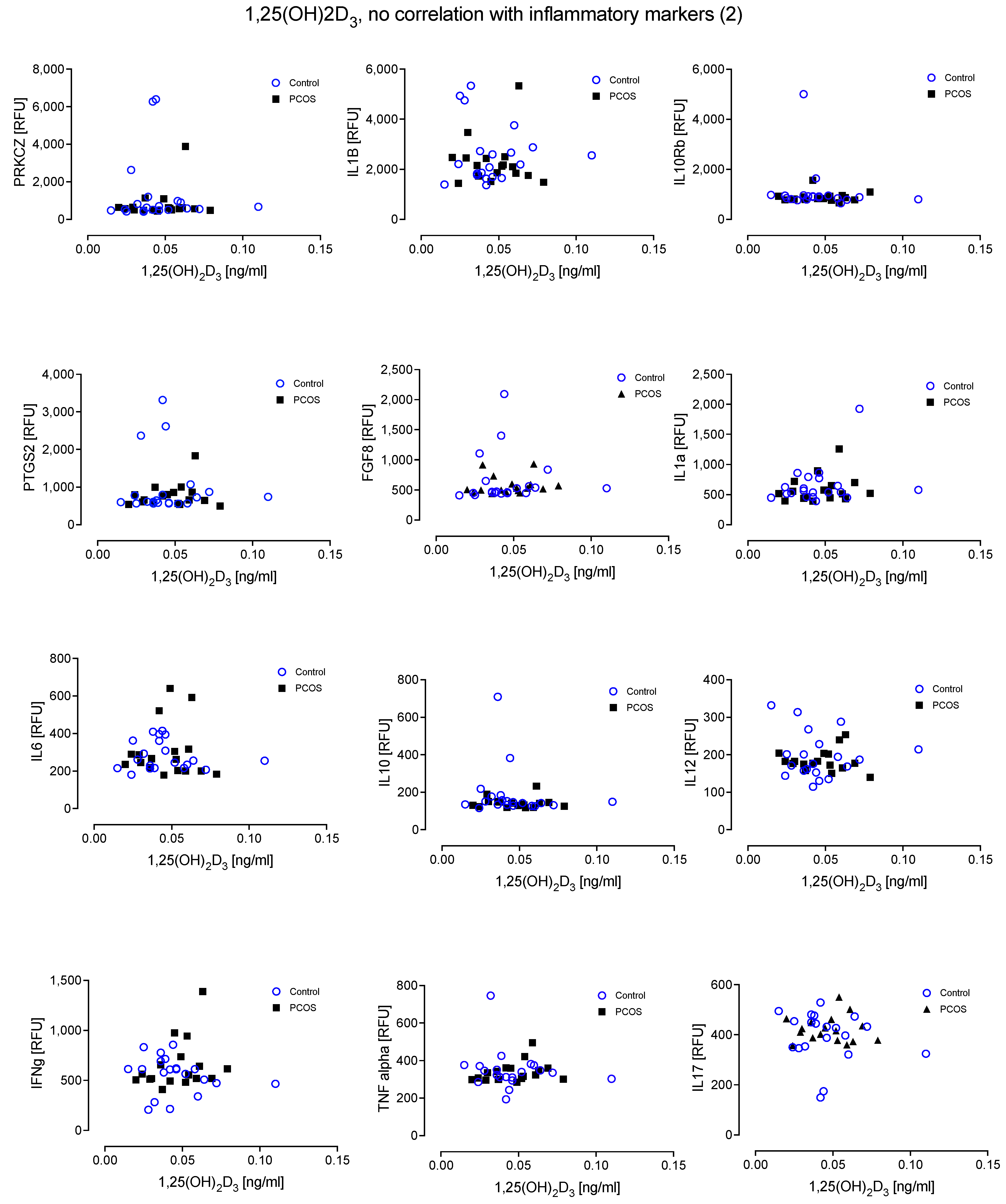
4. Results
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sathyapalan, T.; Atkin, S.L. Recent advances in cardiovascular aspects of polycystic ovary syndrome. Eur. J. Endocrinol. Eur. Fed. Endocr. Soc. 2012, 166, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.M. Complement activation: An emerging player in the pathogenesis of cardiovascular disease. Scientifica 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Morreale, H.F.; Luque-Ramirez, M.; Gonzalez, F. Circulating inflammatory markers in polycystic ovary syndrome: A systematic review and metaanalysis. Fertil. Steril. 2011, 95, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Sathyapalan, T.; Atkin, S.L. Mediators of inflammation in polycystic ovary syndrome in relation to adiposity. Mediat. Inflamm. 2010, 2010, 758656. [Google Scholar] [CrossRef]
- Nehir Aytan, A.; Bastu, E.; Demiral, I.; Bulut, H.; Dogan, M.; Buyru, F. Relationship between hyperandrogenism, obesity, inflammation and polycystic ovary syndrome. Gynecol. Endocrinol. 2016, 32, 709–713. [Google Scholar] [CrossRef]
- Boulman, N.; Levy, Y.; Leiba, R.; Shachar, S.; Linn, R.; Zinder, O.; Blumenfeld, Z. Increased C-reactive protein levels in the polycystic ovary syndrome: A marker of cardiovascular disease. J. Clin. Endocrinol. Metab. 2004, 89, 2160–2165. [Google Scholar] [CrossRef]
- Shen, S.H.; Shen, S.Y.; Liou, T.H.; Hsu, M.I.; Chang, Y.C.; Cheng, C.Y.; Hsu, C.S.; Tzeng, C.R. Obesity and inflammatory biomarkers in women with polycystic ovary syndrome. Eur. J. Obs. Gynecol. Reprod. Biol. 2015, 192, 66–71. [Google Scholar] [CrossRef]
- Un, B.; Dolapcioglu, K.S.; Guler Okyay, A.; Sahin, H.; Beyazit, A. Evaluation of hs-CRP and visseral adiposity index in patients with policystic ovary syndrome by clinical and laboratory findings. Eur. J. Obs. Gynecol. Reprod. Biol. 2016, 204, 16–20. [Google Scholar] [CrossRef]
- Aboeldalyl, S.; James, C.; Seyam, E.; Ibrahim, E.M.; Shawki, H.E.; Amer, S. The Role of Chronic Inflammation in Polycystic Ovarian Syndrome-A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2021, 22, 2734. [Google Scholar] [CrossRef]
- Gonzalez, F.; Thusu, K.; Abdel-Rahman, E.; Prabhala, A.; Tomani, M.; Dandona, P. Elevated serum levels of tumor necrosis factor alpha in normal-weight women with polycystic ovary syndrome. Metabolism 1999, 48, 437–441. [Google Scholar] [CrossRef]
- Ranjbaran, J.; Farimani, M.; Tavilani, H.; Ghorbani, M.; Karimi, J.; Poormonsefi, F.; Khodadadi, I. Matrix metalloproteinases 2 and 9 and MMP9/NGAL complex activity in women with PCOS. Reproduction 2016, 151, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.; Shalev, E. MMPS and TIMPS in ovarian physiology and pathophysiology. Front. Biosci. 2004, 9, 2474–2483. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.A.; Vieira, C.S.; Jacob-Ferreira, A.L.; Belo, V.A.; Soares, G.M.; Fernandes, J.B.; Ferriani, R.A.; Tanus-Santos, J.E. Imbalanced circulating matrix metalloproteinases in polycystic ovary syndrome. Mol. Cell. Biochem. 2011, 353, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, K.C.; Komorowski, J.; O’Callaghan, C.J.; Tan, B.K.; Chen, J.; Prelevic, G.M.; Randeva, H.S. Increased circulating levels of matrix metalloproteinase-2 and -9 in women with the polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 1173–1177. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Guan, Y.M.; Zheng, J.H. Elevated serum levels of matrix metalloproteinase-2 in women with polycystic ovarian syndrome. Int. J. Gynaecol. Obs. 2007, 96, 204–205. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cai, L.Y.; Lv, H.M.; Xia, L.; Zhang, Y.J.; Zhang, H.X.; Guan, Y.M. Raised serum levels of matrix metalloproteinase-9 in women with polycystic ovary syndrome and its association with insulin-like growth factor binding protein-1. Gynecol. Endocrinol. 2008, 24, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Livadas, S.; Kandarakis, S.A.; Margeli, A.; Papassotiriou, I. Serum concentrations of atherogenic proteins neutrophil gelatinase-associated lipocalin and its complex with matrix metalloproteinase-9 are significantly lower in women with polycystic ovary syndrome: Hint of a protective mechanism? Eur. J. Endocrinol. 2008, 158, 525–531. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Sathyapalan, T.; Butler, A.E.; Atkin, S.L. Vitamin D Association with Macrophage-Derived Cytokines in Polycystic Ovary Syndrome: An Enhanced Risk of COVID-19 Infection? Front. Endocrinol. 2021, 12, 638621. [Google Scholar] [CrossRef]
- Khokha, R.; Murthy, A.; Weiss, A. Metalloproteinases and their natural inhibitors in inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 649–665. [Google Scholar] [CrossRef]
- Rudnicka, E.; Suchta, K.; Grymowicz, M.; Calik-Ksepka, A.; Smolarczyk, K.; Duszewska, A.M.; Smolarczyk, R.; Meczekalski, B. Chronic low grade inflammation in pathogenesis of PCOS. Int. J. Mol. Sci. 2021, 22, 3789. [Google Scholar] [CrossRef]
- Hahn, S.; Haselhorst, U.; Tan, S.; Quadbeck, B.; Schmidt, M.; Roesler, S.; Kimmig, R.; Mann, K.; Janssen, O.E. Low serum 25-hydroxyvitamin D concentrations are associated with insulin resistance and obesity in women with polycystic ovary syndrome. Exp. Clin. Endocrinol. Diabetes 2006, 114, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Li, H.W.; Brereton, R.E.; Anderson, R.A.; Wallace, A.M.; Ho, C.K. Vitamin D deficiency is common and associated with metabolic risk factors in patients with polycystic ovary syndrome. Metab. Clin. Exp. 2011, 60, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Bjelakovic, G.; Gluud, L.L.; Nikolova, D.; Whitfield, K.; Wetterslev, J.; Simonetti, R.G.; Bjelakovic, M.; Gluud, C. Vitamin D supplementation for prevention of mortality in adults. Cochrane Database Syst. Rev. 2014, Jan 10, Cd007470. [Google Scholar] [CrossRef]
- Osorio Landa, H.K.; Perez Diaz, I.; Laguna Barcenas, S.D.C.; Lopez Navarro, J.M.; Abella Roa, M.F.; Corral Orozco, M.; Mancilla Ortega, J.P.; Martinez Duarte, D.A.; Morales Montalvo, S.I.; Muzquiz Aguirre, S.; et al. Association of serum vitamin D levels with chronic disease and mortality. Nutr. Hosp. 2020, 37, 335–342. [Google Scholar] [CrossRef]
- Butler, A.E.; Dargham, S.R.; Abouseif, A.; El Shewehy, A.; Atkin, S.L. Vitamin D deficiency effects on cardiovascular parameters in women with polycystic ovary syndrome: A retrospective, cross-sectional study. J. Steroid Biochem. Mol. Biol. 2021, 211, 105892. [Google Scholar] [CrossRef]
- Lerchbaum, E.; Obermayer-Pietsch, B. Vitamin D and fertility: A systematic review. Eur. J. Endocrinol. Eur. Fed. Endocr. Soc. 2012, 166, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Selimoglu, H.; Duran, C.; Kiyici, S.; Ersoy, C.; Guclu, M.; Ozkaya, G.; Tuncel, E.; Erturk, E.; Imamoglu, S. The effect of vitamin D replacement therapy on insulin resistance and androgen levels in women with polycystic ovary syndrome. J. Endocrinol. Investig. 2010, 33, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Krul-Poel, Y.; Snackey, C.; Louwers, Y.; Lips, P.; Lambalk, C.; Laven, J.; Simsek, S. The role of vitamin D in metabolic disturbances in polycystic ovary syndrome (PCOS): A systematic review. Eur. J. Endocrinol. Eur. Fed. Endocr. Soc. 2013, 169, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Mangin, M.; Sinha, R.; Fincher, K. Inflammation and vitamin D: The infection connection. Inflamm. Res. 2014, 63, 803–819. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D metabolism, mechanism of action, and clinical applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Sakaki, T.; Kagawa, N.; Yamamoto, K.; Inouye, K. Metabolism of vitamin D3 by cytochromes P450. Front. Biosci. 2005, 10, 119–134. [Google Scholar] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed]
- Kamao, M.; Tatematsu, S.; Hatakeyama, S.; Sakaki, T.; Sawada, N.; Inouye, K.; Ozono, K.; Kubodera, N.; Reddy, G.S.; Okano, T. C-3 epimerization of vitamin D3 metabolites and further metabolism of C-3 epimers: 25-hydroxyvitamin D3 is metabolized to 3-epi-25-hydroxyvitamin D3 and subsequently metabolized through C-1alpha or C-24 hydroxylation. J. Biol. Chem. 2004, 279, 15897–15907. [Google Scholar] [CrossRef]
- Schleicher, R.L.; Encisco, S.E.; Chaudhary-Webb, M.; Paliakov, E.; McCoy, L.F.; Pfeiffer, C.M. Isotope dilution ultra performance liquid chromatography-tandem mass spectrometry method for simultaneous measurement of 25-hydroxyvitamin D2, 25-hydroxyvitamin D3 and 3-epi-25-hydroxyvitamin D3 in human serum. Clin. Chim. Acta 2011, 412, 1594–1599. [Google Scholar] [CrossRef]
- Al-Zohily, B.; Al-Menhali, A.; Gariballa, S.; Haq, A.; Shah, I. Epimers of Vitamin D: A Review. Int. J. Mol. Sci. 2020, 21, 470. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Dargham, S.R.; Latif, A.; Mokhtar, H.R.; Robay, A.; Chidiac, O.M.; Jayyousi, A.; Al Suwaidi, J.; Crystal, R.G.; Abi Khalil, C.; et al. Association of vitamin D(3) and its metabolites in patients with and without type 2 diabetes and their relationship to diabetes complications. Ther. Adv. Chronic Dis. 2020, 11, 2040622320924159. [Google Scholar] [CrossRef]
- Molnar, F.; Sigueiro, R.; Sato, Y.; Araujo, C.; Schuster, I.; Antony, P.; Peluso, J.; Muller, C.; Mourino, A.; Moras, D.; et al. 1alpha,25(OH)2-3-epi-vitamin D3, a natural physiological metabolite of vitamin D3: Its synthesis, biological activity and crystal structure with its receptor. PLoS ONE 2011, 6, e18124. [Google Scholar] [CrossRef]
- Wortsman, J.; Matsuoka, L.Y.; Chen, T.C.; Lu, Z.; Holick, M.F. Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 2000, 72, 690–693. [Google Scholar] [CrossRef]
- Cunningham, T.K.; Allgar, V.; Dargham, S.R.; Kilpatrick, E.; Sathyapalan, T.; Maguiness, S.; Mokhtar Rudin, H.R.; Abdul Ghani, N.M.; Latiff, A.; Atkin, S.L. Association of Vitamin D Metabolites With Embryo Development and Fertilization in Women With and Without PCOS Undergoing Subfertility Treatment. Front. Endocrinol. 2019, 10, 13. [Google Scholar] [CrossRef]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.R.; Kazantzidis, A.; Kift, R.C.; Farrar, M.D.; Wilkinson, J.; Rhodes, L.E. Meeting Vitamin D Requirements in White Caucasians at UK Latitudes: Providing a Choice. Nutrients 2018, 10, 497. [Google Scholar] [CrossRef] [PubMed]
- Kahal, H.; Halama, A.; Aburima, A.; Bhagwat, A.M.; Butler, A.E.; Grauman, J.; Suhre, K.; Sathyapalan, T.; Atkin, S.L. Effect of induced hypoglycemia on inflammation and oxidative stress in type 2 diabetes and control subjects. Sci. Rep. 2020, 10, 4750. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.; Vaught, J.D.; Bock, C.; Gold, L.; Katilius, E.; Keeney, T.R.; Kim, N.; Saccomano, N.A.; Wilcox, S.K.; Zichi, D.; et al. From SOMAmer-based biomarker discovery to diagnostic and clinical applications: A SOMAmer-based, streamlined multiplex proteomic assay. PLoS ONE 2011, 6, e26332. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.C.; Dunaif, A. Should all women with PCOS be treated for insulin resistance? Fertil. Steril. 2012, 97, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.-L.; Liang, X.-Y.; Yang, X.; Li, Y.; Wei, L.-N. Low-grade chronic inflammation in the peripheral blood and ovaries of women with polycystic ovarian syndrome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 148–150. [Google Scholar] [CrossRef]
- González, F.; Considine, R.V.; Abdelhadi, O.A.; Acton, A.J. Inflammation triggered by saturated fat ingestion is linked to insulin resistance and hyperandrogenism in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2020, 105, e2152–e2167. [Google Scholar] [CrossRef]
- Stocco, C. Tissue physiology and pathology of aromatase. Steroids 2012, 77, 27–35. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2020, Jan 29, 1607. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef]
- Jiang, J.; Cai, X.; Pan, Y.; Du, X.; Zhu, H.; Yang, X.; Zheng, D.; Gaisano, H.; Wei, T.; He, Y. Relationship of obesity to adipose tissue insulin resistance. BMJ Open Diabetes Res. Care 2020, 8, e000741. [Google Scholar] [CrossRef] [Green Version]
- Legro, R.S. Obesity and PCOS: Implications for diagnosis and treatment. In Seminars in Reproductive Medicine; Thieme Medical Publishers: New York, NY, USA, 2012; pp. 496–506. [Google Scholar]
- Moin, A.S.M.; Sathyapalan, T.; Diboun, I.; Elrayess, M.A.; Butler, A.E.; Atkin, S.L. Metabolic consequences of obesity on the hypercoagulable state of polycystic ovary syndrome. Sci. Rep. 2021, 11, 5320. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Abouseif, A.; Dargham, S.R.; Sathyapalan, T.; Atkin, S.L. Metabolic comparison of polycystic ovarian syndrome and control women in Middle Eastern and UK Caucasian populations. Sci. Rep. 2020, 10, 18895. [Google Scholar] [CrossRef] [PubMed]
- Wobke, T.K.; Sorg, B.L.; Steinhilber, D. Vitamin D in inflammatory diseases. Front. Physiol. 2014, 5, 244. [Google Scholar] [CrossRef]
- Zhang, C.; Fu, S.; Zhao, M.; Liu, D.; Zhao, Y.; Yao, Y. Associations Between Complement Components and Vitamin D and the Physical Activities of Daily Living Among a Longevous Population in Hainan, China. Front. Immunol. 2020, 11, 1543. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.F.; Li, B.X.; Zhang, Q. Vitamin D improves levels of hormonal, oxidative stress and inflammatory parameters in polycystic ovary syndrome: A meta-analysis study. Ann. Palliat. Med. 2021, 10, 169–183. [Google Scholar] [CrossRef]
- Ostadmohammadi, V.; Jamilian, M.; Bahmani, F.; Asemi, Z. Vitamin D and probiotic co-supplementation affects mental health, hormonal, inflammatory and oxidative stress parameters in women with polycystic ovary syndrome. J. Ovarian Res. 2019, 12, 5. [Google Scholar] [CrossRef]
- Moin, A.S.M.; Al-Qaissi, A.; Sathyapalan, T.; Atkin, S.L.; Butler, A.E. Renin-Angiotensin System Overactivation in Type 2 Diabetes: A Risk for SARS-CoV-2 Infection? Diabetes Care 2020, 43, e131–e133. [Google Scholar] [CrossRef]
- Lewis, R.D.; Narayanaswamy, A.K.; Farewell, D.; Rees, D.A. Complement activation in polycystic ovary syndrome occurs in the postprandial and fasted state and is influenced by obesity and insulin sensitivity. Clin. Endocrinol. 2021, 94, 74–84. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 24) | PCOS (n = 24) | |
|---|---|---|
| Age (years) | 32.5 ± 4.1 | 31 ± 6.4 |
| BMI (kg/m2) | 24.8 ± 1.1 | 25.9 ± 1.8 |
| Fasting glucose (mmol/L) | 4.9 ± 0.4 | 4.7 ± 0.8 |
| HbA1C (mmol/mol) | 30.9 ± 6.5 | 31.8 ± 3.0 |
| HOMA-IR | 1.8 ± 1.0 | 1.9 ± 1.6 |
| SHBG (nmol/L) | 104.2 ± 80.3 | 71.7 ± 62.2 |
| Free androgen index (FAI) | 1.3 ± 0.5 | 4.1 ± 2.9 ** |
| CRP (mg L−1) | 2.34 ± 2.34 | 2.77 ± 2.57 |
| AMH (ng/mL) | 24 ± 13 | 57 ± 14 ** |
| 25 hydroxy vitamin D3 (nmol/l) | 46.2± 23.5 | 54.0 ± 27.4 |
| 25-hydroxy-3epi-vitamin D3 | 0.97 ± 0.64 | 1.69 ± 1.36 |
| 1,25 dihydroxy vitamin D3 (ng/mL) | 0.03 ± 0.02 | 0.04 ± 0.2 |
| Control | PCOS | p Value | |
|---|---|---|---|
| IL5 | 247 (231) | 252 (130) | 0.92 |
| AZU1 | 194 (120) | 168 (52) | 0.29 |
| RPS6KA5 | 150 (39) | 155 (52) | 0.68 |
| CD40LG | 420 (480) | 296 (128) | 0.18 |
| HMGB1 | 6397 (7451) | 4024 (1283) | 0.10 |
| IL34 | 475 (86) | 482 (86) | 0.77 |
| CXCL10 | 1546 (392) | 1632 (438) | 0.44 |
| S100A9 | 8943 (4519) | 7103 (2911) | 0.07 |
| CCL19 | 424 (597) | 365 (114) | 0.61 |
| SIGLEC1 | 91 (39) | 101 (33) | 0.33 |
| TBK1 | 170 (59) | 166 (51) | 0.77 |
| CLEC7A | 683 (413) | 654 (298) | 0.76 |
| PRKCZ | 1079 (1513) | 742 (627) | 0.27 |
| IL1B | 2492 (1057) | 2316 (785) | 0.48 |
| IL10RB | 1414 (2157) | 924 (188) | 0.23 |
| PTGS2 | 885 (674) | 760 (273) | 0.36 |
| FGF8 | 621 (357) | 584 (140) | 0.60 |
| IL1A | 636 (280) | 584 (173) | 0.40 |
| IL6 | 269 (69) | 298 (127) | 0.30 |
| IL10 | 290 (589) | 146 (26) | 0.19 |
| IL12 | 198 (53) | 213 (105) | 0.49 |
| IFN-g | 552 (156) | 804 (906) | 0.15 |
| TNF-a | 344 (89) | 352 (58) | 0.67 |
| IL17 | 412 (91) | 462 (205) | 0.23 |
| MMP9 | 32,886 (19,126) | 29,041 (15,122) | 0.40 |
| MMP3 | 428 (117) | 720 (1264) | 0.22 |
| MMP7 | 1027 (299) | 939 (284) | 0.25 |
| MMP17 | 959 (817) | 698 (203) | 0.10 |
| MMP2 | 6508 (1628) | 6492 (1399) | 0.97 |
| MMP12 | 883 (405) | 833 (312) | 0.60 |
| MMP1 | 975 (756) | 859 (452) | 0.48 |
| MMP13 | 684 (696) | 527 (184) | 0.25 |
| MMP14 | 1404 (1802) | 1007 (373) | 0.25 |
| MMP16 | 671 (301) | 575 (73) | 0.10 |
| MMP10 | 750 (325) | 823 (336) | 0.41 |
| MMP8 | 3439 (2877) | 2500 (1175) | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moin, A.S.M.; Sathyapalan, T.; Atkin, S.L.; Butler, A.E. Inflammatory Markers in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D Metabolites. Nutrients 2022, 14, 3540. https://doi.org/10.3390/nu14173540
Moin ASM, Sathyapalan T, Atkin SL, Butler AE. Inflammatory Markers in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D Metabolites. Nutrients. 2022; 14(17):3540. https://doi.org/10.3390/nu14173540
Chicago/Turabian StyleMoin, Abu Saleh Md, Thozhukat Sathyapalan, Stephen L. Atkin, and Alexandra E. Butler. 2022. "Inflammatory Markers in Non-Obese Women with Polycystic Ovary Syndrome Are Not Elevated and Show No Correlation with Vitamin D Metabolites" Nutrients 14, no. 17: 3540. https://doi.org/10.3390/nu14173540