
Bacterial Pathogens and Symbionts Harboured by Ixodes ricinus Ticks Parasitising Red Squirrels in the United Kingdom

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
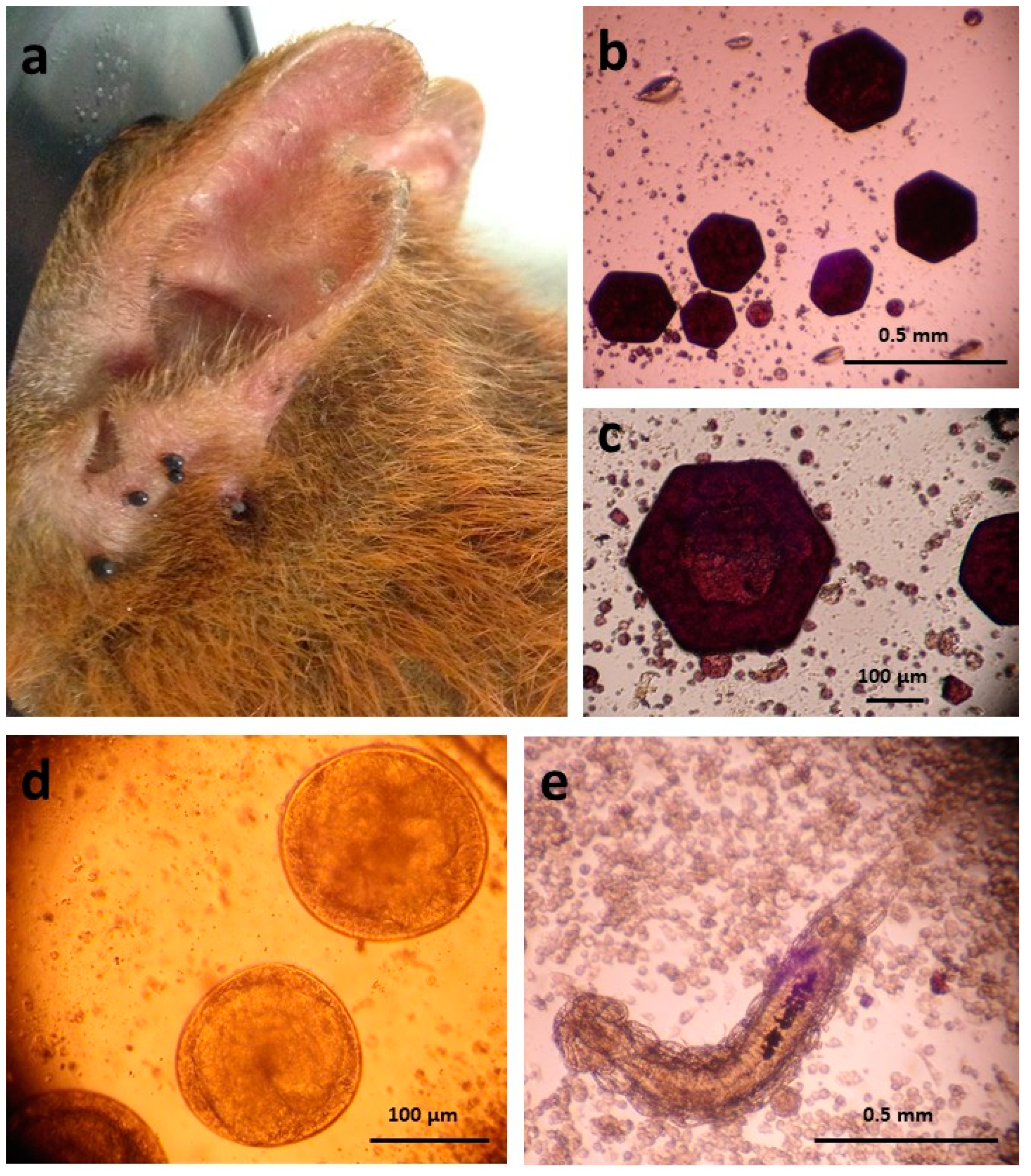
2.1. Ticks
2.2. Tick-Borne Bacteria
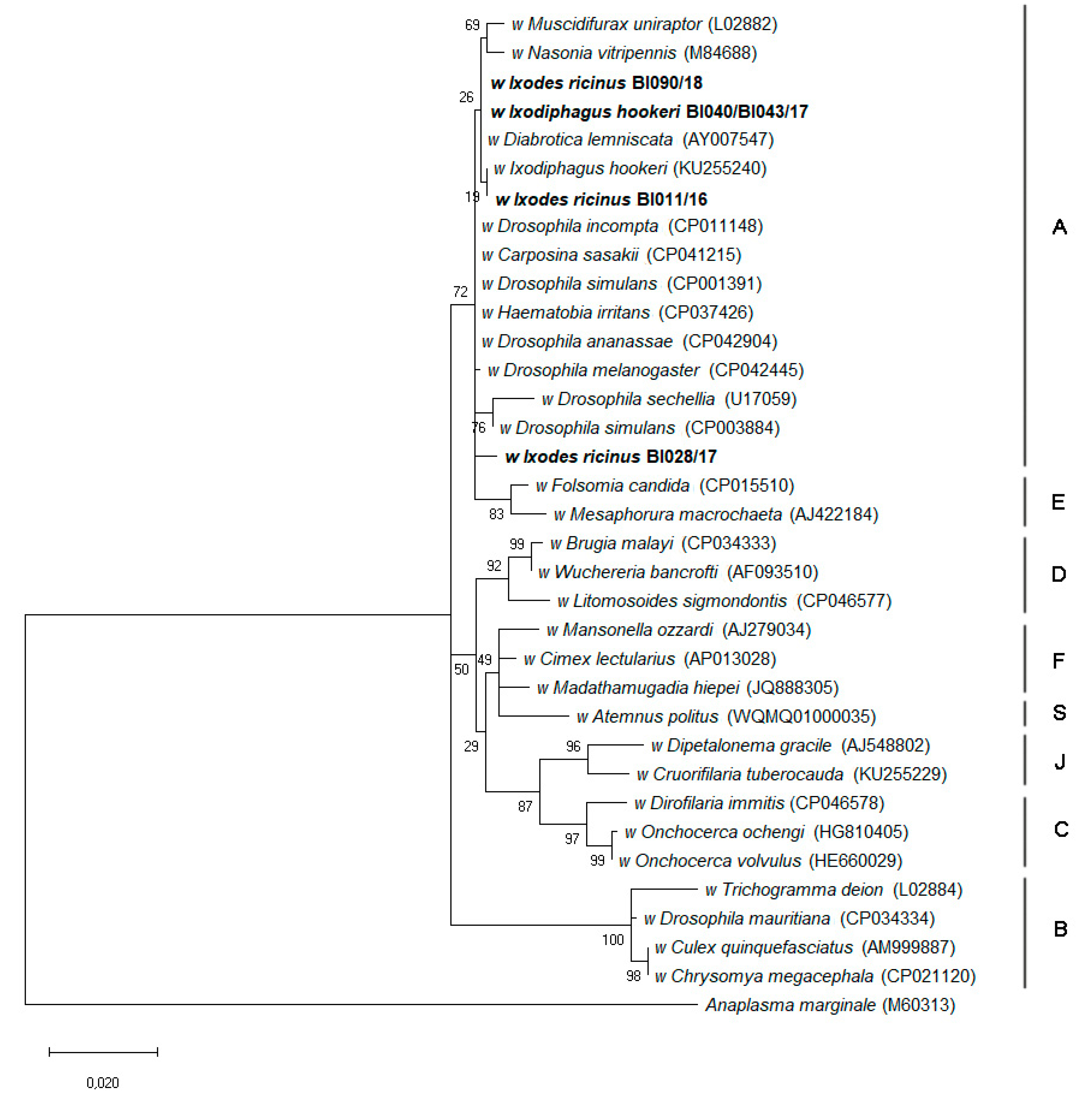
2.3. Ixodiphagus and Wolbachia
2.4. Sequences Submitted to a Public Database
3. Discussion
4. Materials and Methods
4.1. Tick Sampling
4.2. Tick Dissection
4.3. DNA Extraction and PCR Analysis
4.4. Sequence and Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN Red List. Available online: https://www.iucnredlist.org/species/20025/115155900 (accessed on 30 November 2020).
- Hardouin, E.A.; Baltazar-Soares, M.; Schilling, A.-K.; Butler, H.; García-Rodríguez, O.; Crowley, E.; Liang, W.-J.; Meredith, A.; Lurz, P.W.W.; Forster, J.; et al. Conservation of genetic uniqueness in remaining populations of red squirrels (Sciurus vulgaris L.) in the South of England. Ecol. Evol. 2019, 9, 6547–6558. [Google Scholar] [CrossRef] [Green Version]
- Sainsbury, A.W.; Deaville, R.; Lawson, B.; Cooley, W.A.; Farelly, S.S.J.; Stack, M.J.; Duff, P.; McInnes, C.J.; Gurnell, J.; Russell, P.H.; et al. Poxviral disease in red squirrels Sciurus vulgaris in the UK: Spatial and temporal trends of an emerging threat. EcoHealth 2008, 5, 305. [Google Scholar] [CrossRef] [PubMed]
- Avanzi, C.; Del-Pozo, J.; Benjak, A.; Stevenson, K.; Simpson, V.R.; Busso, P.; McLuckie, J.; Loiseau, C.; Lawton, C.; Schoening, J.; et al. Red squirrels in the British Isles are infected with leprosy bacilli. Science 2016, 354, 744–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schilling, A.-K.; Lurz, P.W.W.; Stevenson, K.; Avanzi, C.; Shuttleworth, C.M.; Cole, S.T.; Meredith, A.L. Leprosy in red squirrels in the UK. Vet. Rec. 2019, 184, 416. [Google Scholar] [CrossRef] [PubMed]
- Humair, P.-F.; Gern, L. Relationship between Borrelia burgdorferi sensu lato species, red squirrels (Sciurus vulgaris) and Ixodes ricinus in enzootic areas in Switzerland. Acta Trop. 1998, 69, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Paulauskas, A.; Ambrasiene, D.; Radzijevskaja, J.; Rosef, O.; Turcinaviciene, J. Diversity in prevalence and genospecies of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks and rodents in Lithuania and Norway. Int. J. Med. Microbiol. 2008, 298 (Suppl. S1), 180–187. [Google Scholar] [CrossRef] [Green Version]
- Romeo, C.; Pisanu, B.; Ferrari, N.; Basset, F.; Tillon, L.; Wauters, L.A.; Martinoli, A.; Saino, N.; Chapuis, J.-L. Macroparasite community of the Eurasian red squirrel (Sciurus vulgaris): Poor species richness and diversity. Parasitol. Res. 2013, 112, 3527–3536. [Google Scholar] [CrossRef] [PubMed]
- Simpson, V.R.; Hargreaves, J.; Butler, H.M.; Davison, N.J.; Everest, D.J. Causes of mortality and pathological lesions observed post-mortem in red squirrels (Sciurus vulgaris) in Great Britain. BMC Vet. Res. 2013, 9, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisanu, B.; Chapuis, J.-L.; Dozières, A.; Basset, F.; Poux, V.; Vourc’h, G. High prevalence of Borrelia burgdorferi s.l. in the European red squirrel Sciurus vulgaris in France. Ticks Tick-Borne Dis. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Ruyts, S.C.; Frazer-Mendelewska, E.; Van Den Berge, K.; Verheyen, K.; Sprong, H. Molecular detection of tick-borne pathogens Borrelia afzelii, Borrelia miyamotoi and Anaplasma phagocytophilum in Eurasian red squirrels (Sciurus vulgaris). Eur. J. Wildl. Res. 2017, 63, 43. [Google Scholar] [CrossRef]
- Mysterud, A.; Stigum, V.M.; Jaarsma, R.I.; Sprong, H. Genospecies of Borrelia burgdorferi sensu lato detected in 16 mammal species and questing ticks from northern Europe. Sci. Rep. 2019, 9, 5088. [Google Scholar] [CrossRef]
- Lipatova, I.; Razanske, I.; Jurgelevicius, V.; Paulauskas, A. Bartonella washoensis infection in red squirrels (Sciurus vulgaris) and their ectoparasites in Lithuania. Comp. Immunol. Microbiol. Infect. Dis. 2020, 68, 101391. [Google Scholar] [CrossRef] [PubMed]
- Majerová, K.; Hönig, V.; Houda, M.; Papežík, P.; Fonville, M.; Sprong, H.; Rudenko, N.; Golovchenko, M.; Černá Bolfíková, B.; Hulva, P.; et al. Hedgehogs, squirrels, and blackbirds as sentinel hosts for active surveillance of Borrelia miyamotoi and Borrelia burgdorferi complex in urban and rural environments. Microorganisms 2020, 8, 1908. [Google Scholar] [CrossRef]
- Cisak, E.; Wójcik-Fatla, A.; Zając, V.; Sawczyn, A.; Sroka, J.; Dutkiewicz, J. Spiroplasma—An emerging arthropod-borne pathogen? Ann. Agric. Environ. Med. 2015, 22, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Palomar, A.M.; Premchand-Branker, S.; Alberdi, P.; Belova, O.; Moniuszko-Malinowska, A.; Kahl, O.; Bell-Sakyi, L. Isolation of known and potentially pathogenic tick-borne microorganisms from European ixodid ticks using tick cell lines. Ticks Tick-Borne Dis. 2019, 10, 628–638. [Google Scholar] [CrossRef]
- Schilling, A.-K.; van Hooij, A.; Lurz, P.W.W.; DelPozo, J.; Stevenson, K.; Meredith, A.; Geluk, A. Detection of humoral immunity to mycobacteria causing leprosy in Eurasian red squirrels (Sciurus vulgaris) using a quantitative rapid test. Eur. J. Wildl. Res. 2019, 65, 49. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, A.I.; Bakker, J.W.; Koenraadt, J.M.; Fonville, M.; Takumi, K.; Sprong, H.; Demir, S. Tripartite interactions among Ixodiphagus hookeri, Ixodes ricinus and deer: Differential interference with transmission cycles of tick-borne pathogens. Pathogens 2020, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasit. Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaRose, J.P.; Meredith, A.L.; Everest, D.J.; Fiegna, C.; McInnes, C.J.; Shaw, D.J.; Milne, E.M. Epidemiological and postmortem findings in 262 red squirrels (Sciurus vulgaris) in Scotland, 2005 to 2009. Vet. Rec. 2010, 167, 297–302. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Nava, S.; Petney, T. Description of all the stages of Ixodes inopinatus n. sp. (Acari: Ixodidae). Ticks Tick-Borne Dis. 2014, 5, 734–743. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Rieß, R.; Kahl, O.; Wölfel, S.; Dobler, G.; Nava, S.; Estrada-Peña, A. Ixodes inopinatus—Occurring also outside the Mediterranean region. Ticks Tick-Borne Dis. 2018, 9, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Hauck, D.; Springer, A.; Pachnicke, S.; Schunack, B.; Fingerle, V.; Strube, C. Ixodes inopinatus in northern Germany: Occurrence and potential vector role for Borrelia spp., Rickettsia spp., and Anaplasma phagocytophilum in comparison with Ixodes ricinus. Parasitol. Res. 2019, 118, 3205–3216. [Google Scholar] [CrossRef] [PubMed]
- Younsi, H.; Fares, W.; Cherni, S.; Dachraoui, K.; Barhoumi, W.; Najjar, C.; Zhioua, E. Ixodes inopinatus and Ixodes ricinus (Acari: Ixodidae) are sympatric ticks in North Africa. J. Med. Entomol. 2020, 57, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Millins, C.; Gilbert, L.; Johnson, P.; James, M.; Kilbride, E.; Birtles, R.; Biek, R. Heterogeneity in the abundance and distribution of Ixodes ricinus and Borrelia burgdorferi (sensu lato) in Scotland: Implications for risk prediction. Parasit. Vectors 2016, 9, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millins, C.; Leo, W.; MacInnes, I.; Ferguson, J.; Charlesworth, G.; Nayar, D.; Davison, R.; Yardley, J.; Kilbride, E.; Huntley, S.; et al. Emergence of Lyme disease on treeless islands, Scotland, United Kingdom. Emerg. Infect. Dis. 2021, 27, 538–546. [Google Scholar] [CrossRef]
- Layzell, S.J.; Bailey, D.; Peacey, M.; Nuttall, P.A. Prevalence of Borrelia burgdorferi and Borrelia miyamotoi in questing Ixodes ricinus ticks from four sites in the UK. Ticks Tick-Borne Dis. 2018, 9, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Hansford, K.M.; Fonville, M.; Gillingham, E.L.; Coipan, E.C.; Pietzsch, M.E.; Krawczyk, A.I.; Vaux, A.G.C.; Cull, B.; Sprong, H.; Medlock, J.M. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick-Borne Dis. 2017, 8, 353–361. [Google Scholar] [CrossRef]
- Cull, B.; Hansford, K.M.; McGinley, L.; Gillingham, E.L.; Vaux, A.G.C.; Smith, R.; Medlock, J.M. A nationwide study on Borrelia burgdorferi s.l. infection rates in questing Ixodes ricinus: A six-year snapshot study in protected recreational areas in England and Wales. Med. Vet. Entomol. 2021. [Google Scholar] [CrossRef]
- Hansford, K.M.; Fonville, M.; Jahfari, S.; Sprong, H.; Medlock, J.M. Borrelia miyamotoi in host-seeking Ixodes ricinus ticks in England. Epidemiol. Infect. 2015, 143, 1079–1087. [Google Scholar] [CrossRef]
- Hall, J.L.; Alpers, K.; Bown, K.J.; Martin, S.J.; Birtles, R.J. Use of mass-participation outdoor events to assess human exposure to tickborne pathogens. Emerg. Infect. Dis. 2017, 23, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Kubiak, K.; Szczotko, M.; Dmitryjuk, M. Borrelia miyamotoi—An emerging human tick-borne pathogen in Europe. Microorganisms 2021, 9, E154. [Google Scholar] [CrossRef] [PubMed]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Alberdi, M.P.; Walker, A.R.; Paxton, E.A.; Sumption, K.J. Natural prevalence of infection with Ehrlichia (Cytoecetes) phagocytophila of Ixodes ricinus ticks in Scotland. Vet. Parasitol. 1998, 78, 203–213. [Google Scholar] [CrossRef]
- Smith, F.D.; Ellse, L.; Wall, R. Prevalence of Babesia and Anaplasma in ticks infesting dogs in Great Britain. Vet. Parasitol. 2013, 198, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Duplan, F.; Davies, S.; Filler, S.; Abdullah, S.; Keyte, S.; Newbury, H.; Helps, C.R.; Wall, R.; Tasker, S. Anaplasma phagocytophilum, Bartonella spp., haemoplasma species and Hepatozoon spp. in ticks infesting cats: A large-scale survey. Parasit. Vectors 2018, 11, 201. [Google Scholar] [CrossRef]
- Rikihisa, Y.; Zhi, N.; Wormser, G.P.; Wen, B.; Horowitz, H.W.; Hechemy, K.E. Ultrastructural and antigenic characterization of a granulocytic ehrlichiosis agent directly isolated and stably cultivated from a patient in New York State. J. Infect. Dis. 1997, 175, 210–213. [Google Scholar] [CrossRef] [Green Version]
- Tijsse-Klasen, E.; Jameson, L.J.; Fonville, M.; Leach, S.; Sprong, H.; Medlock, J.M. First detection of spotted fever group rickettsiae in Ixodes ricinus and Dermacentor reticulatus ticks in the UK. Epidemiol. Infect. 2011, 139, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Jahfari, S.; Fonville, M.; Hengeveld, P.; Reusken, C.; Scholte, E.-J.; Takken, W.; Heyman, P.; Medlock, J.M.; Heylen, D.; Kleve, J.; et al. Prevalence of Neoehrlichia mikurensis in ticks and rodents from North-west Europe. Parasites Vectors 2012, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Hartelt, K.; Oehme, R.; Frank, H.; Brockmann, S.O.; Hassler, D.; Kimmig, P. Pathogens and symbionts in ticks: Prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp. in Southern Germany. Int. J. Med. Microbiol. 2004, 293 (Suppl. S37), 86–92. [Google Scholar] [CrossRef]
- Sarih, M.; M’Ghirbi, Y.; Bouattour, A.; Gern, L.; Baranton, G.; Postic, D. Detection and identification of Ehrlichia spp. in ticks collected in Tunisia and Morocco. J. Clin. Microbiol. 2005, 43, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- van Overbeek, L.; Gassner, L.; van der Plas, C.L.; Kastelein, P.; Nunes-da Rocha, U.; Takken, W. Diversity of Ixodes ricinus tick-associated bacterial communities from different forests. FEMS Microbiol. Ecol. 2008, 66, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Carpi, G.; Cagnacci, F.; Wittenkindt, N.E.; Zhao, F.; Qi, J.; Tomsho, L.P.; Drautz, D.I.; Rizzoli, A.; Schuster, S.C. Metagenomic profile of the bacterial communities associated with Ixodes ricinus ticks. PLoS ONE 2011, 6, e25604. [Google Scholar] [CrossRef] [PubMed]
- Tijsse-Klasen, E.; Braks, M.; Scholte, E.-J.; Sprong, H. Parasites of vectors—Ixodiphagus hookeri and its Wolbachia symbionts in ticks in the Netherlands. Parasit. Vectors 2011, 4, 228. [Google Scholar] [CrossRef] [Green Version]
- Plantard, O.; Bouju-Albert, A.; Malard, M.-A.; Hermouet, A.; Capron, G.; Verheyden, H. Detection of Wolbachia in the tick Ixodes ricinus is due to the presence of the Hymenoptera endoparasitoid Ixodiphagus hookeri. PLoS ONE 2012, 7, e30692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebbak, A.; Dahmana, H.; Almeeras, L.; Raoult, D.; Boulanger, N.; Jaulhac, B.; Mediannikov, O.; Parola, P. Co-infection of bacteria and protozoan parasites in Ixodes ricinus nymphs collected in the Alsace region, France. Ticks Tick-Borne Dis. 2019, 10, 101241. [Google Scholar] [CrossRef]
- Khoo, J.J.; Kurtti, T.J.; Husin, N.A.; Beliavskaia, A.; Lim, F.S.; Zulkifli, M.M.S.; Al-Khafaji, A.M.; Hartley, C.; Darby, A.C.; Hughes, G.L.; et al. Isolation and propagation of laboratory strains and a novel flea-derived field strain of Wolbachia in tick cell lines. Microorganisms 2020, 8, 988. [Google Scholar] [CrossRef]
- Burtt, E.T. The occurrence of the tick parasite Ixodiphagus caucurtei du Buysson (Hymenoptera; Chalcidoidea; fam. Encyrtidae) in Great Britain. Proc. R. Entomol. Soc. Lond. (Ser. A) 1943, 18, 28–29. [Google Scholar] [CrossRef]
- Lejal, E.; Moutailler, S.; Šimo, L.; Vayssier-Taussat, M.; Pollet, T. Tick-borne pathogen detection in midgut and salivary glands of adult Ixodes ricinus. Parasit. Vectors 2019, 12, 152. [Google Scholar] [CrossRef] [Green Version]
- Moutailler, S.; Moro, C.V.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.-F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-infection of ticks: The rule rather than the exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasit. Vectors 2018, 11, 12. [Google Scholar] [CrossRef]
- Smit, J.D.G.; Grandjean, O.; Guggenheim, R.; Winterhalter, K.H. Haemoglobin crystals in the midgut of the tick Ornithodorus moubata Murray. Nature 1977, 266, 536–538. [Google Scholar] [CrossRef]
- Smit, J.D.G.; Guggenheim, R.; Bauer, P.G. Crystallized hemoglobin in Rhodnius prolixus after a blood meal on guinea-pig. Experientia 1983, 39, 1335–1338. [Google Scholar] [CrossRef]
- Bland, D.M.; Jarrett, C.O.; Bosio, C.F.; Hinnebusch, B.J. Infectious blood source alters early foregut infection and regurgitative transmission of Yersinia pestis by rodent fleas. PLoS Pathog. 2018, 14, e1006859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliburton, W.D. On the haemoglobin crystals of rodents’ blood. Quart. J. Microscop. Sci. 1887, s2-28, 181–199. [Google Scholar]
- Schilling, A.-K.; Lurz, P.W.W.; Meredith, A.L. Field anaesthesia of Eurasian red squirrels (Sciurus vulgaris). Vet. Anaesth. Analg. 2020, 47, 736–738. [Google Scholar] [CrossRef]
- Hillyard, P. Ticks of Northwest Europe. Synopses of the British Fauna (New Series); The Natural History Museum: London, UK, 1996; Volume 52. [Google Scholar]
- Black, W.C.; Piesman, J. Phylogeny of hard and soft tick taxa (Acari:Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.J.B.; Happ, C.M.; Mayer, L.W.; Piesman, J. Detection of Borrelia burgdorferi in ticks by species-specific amplification of the flagellin gene. Am. J. Trop. Med. Hyg. 1992, 47, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Hendricks, A.; Burge, D. Molecular identification and analysis of Borrelia burgdorferi sensu lato in lizards in the Southeastern United States. Appl. Environ. Microbiol. 2005, 71, 2616–2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovius, J.W.; de Wever, B.; Sohne, M.; Brouwer, M.C.; Coumou, J.; Wagemakers, A.; Oei, A.; Knol, H.; Narasimhan, S.; Hodiamont, C.J.; et al. A case of meningoencephalitis by the relapsing fever spirochaete Borrelia miyamotoi in Europe. Lancet 2013, 382, 658. [Google Scholar] [CrossRef] [Green Version]
- Wagemakers, A.; Jahfari, S.; de Wever, B.; Spanjaard, L.; Starink, M.V.; de Vries, H.J.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi in vectors and hosts in The Netherlands. Ticks Tick-Borne Dis. 2017, 8, 370–374. [Google Scholar] [CrossRef]
- Liz, J.S.; Sumner, J.W.; Pfister, K.; Brossard, M. PCR detection and serological evidence of granulocytic ehrlichial infection in roe deer (Capreolus capreolus) and chamois (Rupicapra rupicapra). J. Clin. Microbiol. 2002, 40, 892–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezas-Cruz, A.C.; Zweygarth, E.; Ribeiro, M.F.; da Silveira, J.A.; de la Fuente, J.; Grubhoffer, L.; Valdés, J.J.; Passos, L.M. New species of Ehrlichia isolated from Rhipicephalus (Boophilus) microplus shows an ortholog of the E. canis major immunogenic glycoprotein gp36 with a new sequence of tandem repeats. Parasit. Vectors 2012, 5, 291. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.A.; Livanova, N.N.; Panov, V.V.; Doroschenko, E.K.; Pukhovskaya, N.M.; Vysochina, N.P.; Ivanov, L.I. Genetic diversity of Anaplasma and Ehrlichia in the Asian part of Russia. Ticks Tick-Borne Dis. 2010, 1, 57–65. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Fukatsu, T.; Nikoh, N. Endosymbiotic microbiota of the bamboo pseudococcid Antonina crawii (Insecta, Homoptera). Appl. Environ. Microbiol. 2000, 66, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Haselkorn, T.S.; Markow, T.A.; Moran, N.A. Multiple introductions of the Spiroplasma bacterial endosymbiont into Drosophila. Mol. Ecol. 2009, 18, 1294–1305. [Google Scholar] [CrossRef]
- Casiraghi, M.; Anderson, T.J.; Bandi, C.; Bazzocchi, C.; Genchi, C. A phylogenetic analysis of filarial nematodes: Comparison with the phylogeny of Wolbachia endosymbionts. Parasitology 2001, 122, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Lefoulon, E.; Gavotte, L.; Junker, K.; Barbuto, M.; Uni, S.; Landmann, F.; Laaksonen, S.; Saari, S.; Nikander, S.; de Souza Lima, S.; et al. A new type F Wolbachia from Splendidofilariinae (Onchocercidae) supports the recent emergence of this supergroup. Int. J. Parasitol. 2012, 42, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Woldehiwet, Z.; Horrocks, B.K. Antigenicity of ovine strains of Anaplasma phagocytophilum grown in tick cells and ovine granulocytes. J. Comp. Pathol. 2005, 132, 322–328. [Google Scholar] [CrossRef]
- Moniuszko, A.; Rueckert, C.; Alberdi, M.P.; Barry, G.; Stevenson, B.; Fazakerley, J.K.; Kohl, A.; Bell-Sakyi, L. Coinfection of tick cell lines has variable effects on replication of intracellular bacterial and viral pathogens. Ticks Tick-Borne Dis. 2014, 5, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Santibáñez, S.; Portillo, A.; Palomar, A.M.; Oteo, J.A. Isolation of Rickettsia amblyommatis in HUVEC line. New Microb. New Infect. 2017, 21, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Palomar, A.M.; Kazimirova, M. Isolation and propagation of a Spiroplasma sp. from Slovakian Ixodes ricinus ticks in Ixodes spp. cell lines. Ticks Tick-Borne Dis. 2015, 6, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Auto. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site/Year | Squirrels Trapped | Squirrels with Ticks (%) | Squirrels Whose Ticks Were Screened | Ticks Screened (Larvae, Nymphs) | Pools | Single Ticks |
|---|---|---|---|---|---|---|
| Brownsea/Autumn 2016 | 26 | 25 (96.2%) | 19 | 81 (33, 48) | 18 | 1 |
| Brownsea/Spring 2017 | 26 | 24 (92.3%) | 11 | 13 (0, 13) | 0 | 13 |
| Brownsea/Autumn 2017 | 20 | 20 (100%) | 20 | 211 (0, 211)1 | 29 | 39 |
| Brownsea/Spring 2018 | 25 | 15 (60.0%) | 15 | 43 (3, 40) | 9 | 19 |
| Brownsea/Autumn 2018 | 29 | 28 (96.6%) | 28 | 51 (0, 51) | 0 | 51 |
| Brownsea total | 126 | 112 (88.9%) | 93 | 399 (36, 363) 1 | 56 | 123 |
| Arran/Spring 2017 | 17 | 4 (23.5%) | 0 | |||
| Arran/Autumn 2017 | 6 | 6 (100%) | 6 | 21 (0, 21) | 4 | 4 |
| Arran/Spring 2018 | 14 | 1 (7.1%) | 0 | |||
| Arran/Autumn 2018 | 25 | 22 (88.0%) | 18 | 18 (0, 18) | 0 | 18 |
| Arran total | 62 | 33 (53.2%) | 24 | 39 (0, 39) | 4 | 22 |
| Site and Year | No. of Ticks Sampled 1 | No. Borrelia Positive 1 | Borrelia afzelii1 | Borrelia burgdorferi s.s. 1 | Borrelia garinii1 | Borrelia valaisiana1 | Borrelia miyamotoi1 |
|---|---|---|---|---|---|---|---|
| Brownsea Island | |||||||
| 2016 Autumn | 81 (18, 1) | 5 (5, 0) | 5 (5, 0) | 0 | 0 | 0 | 1 (1, 0) |
| 2017 Spring | 13 (0, 13) | 0 | 0 | 0 | 0 | 0 | 0 |
| 2017 Autumn | 209 (29, 37) | 22 (19, 3) 2 | 19 (16, 3) | 0 | 2 (2, 0) | 1 (1, 0) | ND |
| 2018 Spring | 43 (9, 19) | 11 (4, 7) 3 | 3 (1, 2) | 0 | 6 (1, 5) | 1 (1, 0) | 0 |
| 2018 Autumn | 51 (0, 51) | 9 (0, 9) 4 | 6 (0, 6) | 0 | 1 (0, 1) | 1 (0, 1) | ND |
| Total | 397 (56, 121) | 47 (28, 19) 3 | 33 (22, 11) | 0 | 9 (3, 6) | 3 (2, 1) | 1 |
| Arran | |||||||
| 2017 Autumn | 21 (4, 4) | 3 (2, 1) | 0 | 2 (1, 1) | 1 (1, 0) | 0 | ND |
| 2018 Autumn | 18 (0, 18) | 6 (0, 6) | 1 (0, 1) | 1 (0, 1) | 2 (0, 2) | 2 (0, 2) | ND |
| Total | 39 (4, 22) | 9 (2, 7) | 1 (0, 1) | 3 (1, 2) | 3 (1, 2) | 2 (0, 2) | ND |
| Overall minimum prevalence | 12.8% 56/436 | 7.8% 34/436 | 0.7% 3/436 | 2.8% 12/436 | 1.1% 5/436 | 0.7% 1/136 |
| Site and Year | No. of Ticks Sampled 1 | Anaplasma phagocytophilum1 | Ehrlichia sp. 1 | Spiroplasma sp. 1 | Wolbachia sp. 1 |
|---|---|---|---|---|---|
| Brownsea Island | |||||
| 2016 Autumn | 81 (18, 1) | 1 (1, 0) | 0 | 0 | 10 (9, 1) |
| 2017 Spring | 13 (0, 13) | 0 | 1 (0, 1) | 0 | 8 (0, 8) |
| 2017 Autumn | 209 (29, 37) | 3 (3, 0) 2 | 0 | 1 (1, 0) | 12 (6, 6) |
| 2018 Spring | 43 (9, 19) | 1 (0, 1) | 0 | 0 | 13 (5, 8) |
| 2018 Autumn | 51 (0, 51) | 0 3 | 0 | 0 3 | 24 (0, 24) |
| Total | 397 (56, 121) | 5 (4, 1) | 1 (0, 1) | 1 (1, 0) | 67 (20, 47) |
| Arran | |||||
| 2017 Autumn | 21 (4, 4) | 2 (2, 0) | 0 | 1 (1, 0) | 0 |
| 2018 Autumn | 18 (0, 18) | 0 3 | 0 | 0 | 0 |
| Total | 39 (4, 22) | 2 | 0 | 1 (1, 0) | 0 |
| Overall minimum prevalence | 1.6% (7/436) | 0.2% (1/436) | 0.5% (2/436) | 15.4% (67/436) 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luu, L.; Palomar, A.M.; Farrington, G.; Schilling, A.-K.; Premchand-Branker, S.; McGarry, J.; Makepeace, B.L.; Meredith, A.; Bell-Sakyi, L. Bacterial Pathogens and Symbionts Harboured by Ixodes ricinus Ticks Parasitising Red Squirrels in the United Kingdom. Pathogens 2021, 10, 458. https://doi.org/10.3390/pathogens10040458
Luu L, Palomar AM, Farrington G, Schilling A-K, Premchand-Branker S, McGarry J, Makepeace BL, Meredith A, Bell-Sakyi L. Bacterial Pathogens and Symbionts Harboured by Ixodes ricinus Ticks Parasitising Red Squirrels in the United Kingdom. Pathogens. 2021; 10(4):458. https://doi.org/10.3390/pathogens10040458
Chicago/Turabian StyleLuu, Lisa, Ana M. Palomar, Gemma Farrington, Anna-Katarina Schilling, Shonnette Premchand-Branker, John McGarry, Benjamin L. Makepeace, Anna Meredith, and Lesley Bell-Sakyi. 2021. "Bacterial Pathogens and Symbionts Harboured by Ixodes ricinus Ticks Parasitising Red Squirrels in the United Kingdom" Pathogens 10, no. 4: 458. https://doi.org/10.3390/pathogens10040458