1. Introduction
Climate change modifies mosquito species distributions around the world, enabling the establishment of newly introduced species. Invasive mosquito species (IMS) are newly introduced species in an area, where they tend to spread, potentially having an impact on native species and ecosystems or on human activities (agriculture, conservation, and tourism) [
1,
2]. Mosquito species invasion is a global concern. In Europe, IMS represent a public health threat in an increasing number of countries [
3,
4], while in the Americas, the West Nile virus (WNV) epidemic, which started in 1999 in the state of New York [
5], appeared to be transmitted by an invasive species,
Culex pipiens (Linnaeus, 1758).
The 2016 Zika epidemics renewed concerns in North America about the presence of locally established vector populations, namely of the yellow fever mosquito
Aedes (
Stegomyia)
aegypti (Linnaeus, 1762) and the Asian tiger mosquito
Aedes (
Stegomyia)
albopictus (Skuse, 1894) (Diptera: Culicidae). Among the IMS currently present in North America, these two species represent specific threats for Canada. Prior to 2016,
Ae. albopictus had been detected only on a rare and episodic basis in Canada by the WNV surveillance program, near the Montréal international airport in Québec [
6,
7] and in southern communities of Ontario [
8]. In 2017, the Centers for Disease Control and Prevention (CDC) of the United States of America estimated that
Ae. albopictus’ potential distribution in the United States could extend to the northernmost states of the east coast, which share borders with Canada: New York, Vermont, and New Hampshire [
9,
10,
11]. Enhanced surveillance in Windsor-Essex County (in Ontario, Canada) detected the presence of both species at the larval stage in 2016. This was the first time
Ae. aegypti was documented in Canada, and the first official detection of juvenile stages for either species north of the border [
8]. Diapause is a key adaptation for winter survival of mosquitoes [
12]. While both species are highly invasive and competitive anthropophilic container-inhabiting species, neither
Ae. albopictus nor
Ae. aegypti are particularly good at overwintering. While the former is capable of ecological plasticity to some extent, it has limited egg diapause and cold-hardiness in North America [
13,
14], which has restricted its northward range expansion during the last 35 years to latitudes well south of Québec [
11].
The spread of these invasive species is a major public health concern, mainly because of their ability to transmit a variety of arboviruses. Indeed, both species are competent vectors of the chikungunya virus, all four dengue virus serotypes, yellow fever virus, and Zika virus [
10,
13,
15,
16,
17]. Additionally,
Ae. albopictus has been shown, under experimental conditions, to be a competent vector for at least 22 arboviruses alone, including some that are endemic to Canada, such as WNV, Cache Valley, eastern equine encephalitis, and Jamestown Canyon viruses [
18,
19,
20], thus favoring a potential resurgence of mosquito-borne diseases native to North America [
21,
22,
23]. In addition,
Ae. albopictus is known to thrive around human habitation and urbanized environments, and to be a significant biting nuisance, feeding on a wide range of hosts [
24].
Other medically-relevant mosquito species ecologically-related to invasive
Aedes spp. are already present in Canadian territory, namely
Ochlerotatus japonicus [
25], sometimes referred to as
Aedes japonicus, a highly invasive species in North America, which was first detected in Quebec in 2002, and a competent vector of the WNV and Cache Valley virus [
26,
27,
28,
29].
Ochlerotatus triseriatus is endemic to the eastern part of North America, and was detected in Manitoba, Canada [
30]. It is a primary vector of La Crosse encephalitis virus (LACV), and is competent for other arboviruses, including WNV [
31,
32,
33,
34]. Along with
Ae. albopictus,
Oc. hendersoni, and
Oc. atropalpus, they form the
Aedine Multivoltine group, Oc. triseriatus Type (AMOT) ecological group, characterized by their use of naturally-occurring or artificial water containers for egg laying [
34]. Eggs of
Oc. triseriatus and
Oc. japonicus can undergo diapause and, thus, overwinter in temperate climates, despite adults being unable to survive through this period [
35,
36]. While there are temporal differences among these species’ life cycles, they are far from distinct and they frequently co-occur, cohabitating in an “
Aedes/
Ochlerotatus community” [
13,
26,
31,
34,
37,
38,
39].
To the best of our knowledge, no specific surveillance or study has been conducted on the AMOT species and their role in Québec’s arbovirus circulation, although specimens were detected at low abundance through WNV surveillance [
6]. Québec’s mosquito surveillance was historically designed for monitoring WNV and targets its most abundant competent vector species, namely the
Culex pipiens complex and
Ae. Vexans (Meigen, 1830).
For targeted surveillance of
Ae. albopictus and
Ae. aegypti, we aimed to develop a sampling design that focused on the main introduction pathways for these exotic mosquitoes. Vehicular movements along road networks have been identified as the main route by which adult mosquitoes move to new regions in Europe [
40,
41,
42,
43]. The European Centre for Disease Prevention and Control recommends different surveillance scenarios to improve monitoring approaches and to limit the accidental mechanical introduction of IMS. Scenario 1 (unreported and unestablished IMS, with non-negligible introduction and establishment potential) corresponds to the situation in Québec relative to invasive
Aedes spp. and involves implementing surveillance aimed at detecting possible introduction and establishment of IMS at points of entry (sites where these mosquitoes could be introduced either by plane, boat, train or vehicle) [
42]. Similarly, the World Health Organization’s guiding principles for entomological surveillance of
Aedes spp. in the context of Zika virus epidemics recommend the enhancement of mosquito surveillance at border areas, and the implementation of vector surveillance and control at points of entry as per International Health Regulations [
44,
45].
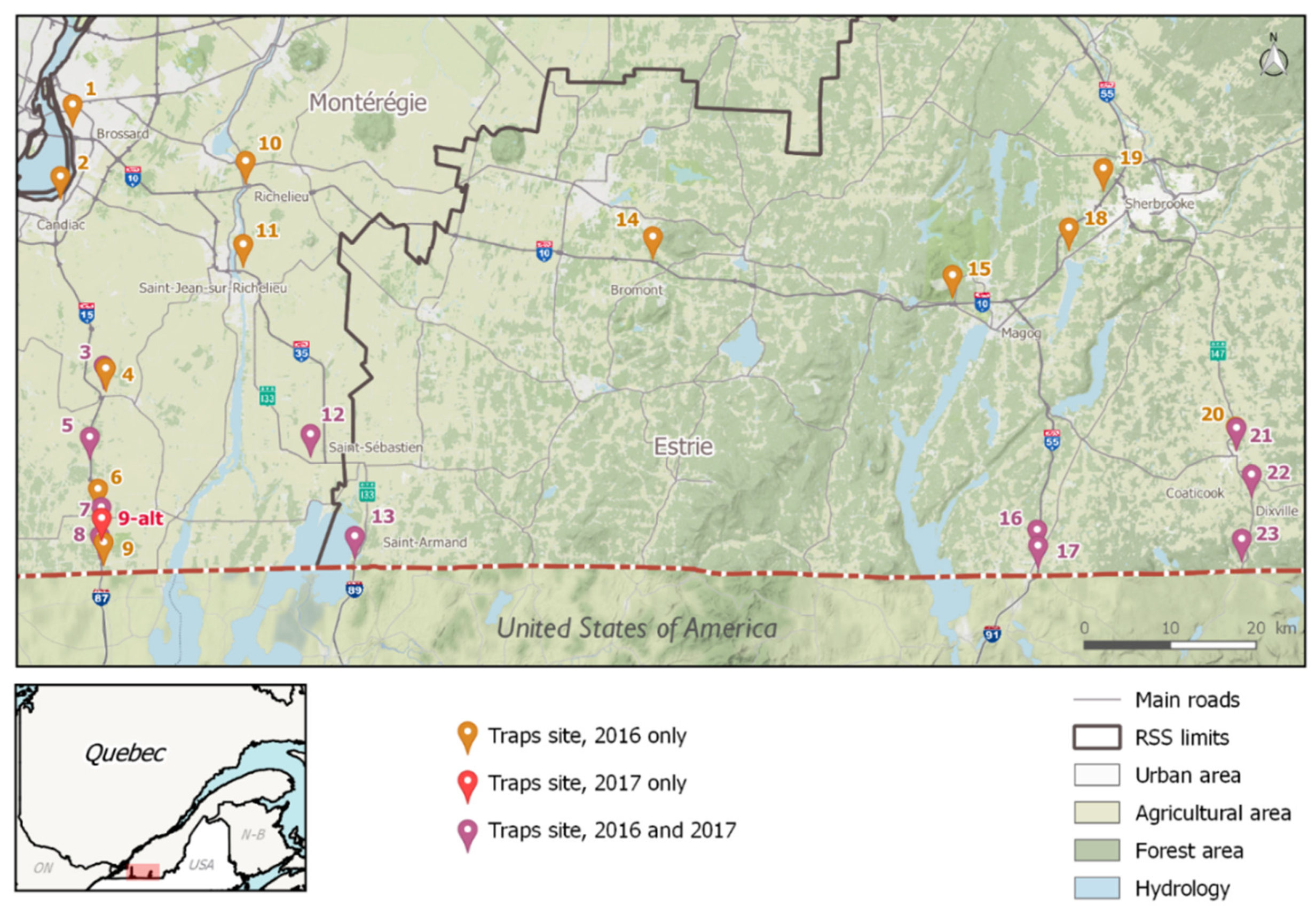
Given the current situation in neighboring Ontario, and knowing that the mechanical introduction of IMS through ground transportation is well documented even over long distances, the objective of this work was to assess, using an appropriate surveillance design, whether invasive Aedes spp. (Ae. albopictus and Ae. aegypti), could be or have been introduced mechanically in southern Québec. We also aimed to characterize the presence and abundance of other species already established in this region, including Oc. triseriatus and Oc. japonicus.
2. Results
2.1. Mosquito Captures
In 2016, eggs were found in 10 ovitraps (OVI, designed to collect mosquito eggs, consist of a wood piece, as an oviposition substrate, placed in a black plastic bucket half-filled with a fermented herb solution, as a natural attractant) (OVI positivity index, OPI = 4%) and there was a positive OVI found at 7 of the 23 sampling sites (30.4%). A total of 472 eggs were retrieved from the OVI, of which 205 hatched, reached fourth instar larvae stage (43.4%), and were successfully identified to the species level. Four adult mosquitoes were accidentally captured in OVI and subsequently identified to the species level (
Table 1). In 2017, eggs were found in 74 OVI (OPI = 22.8%), distributed over 11 of 12 sites (91.7%). A total of 7654 eggs were collected, of which 2206 hatched into larvae (28.8%); 1063 reached the adult stage (13.9% of the total eggs), and were identified by the species. The number of eggs collected daily peaked during mid-July to mid-August 2017, reaching a maximum of 3150 eggs on 31 July. Location-wise, the overall OPI was 62.5% (15 of 24 sites collected eggs during the two-year sampling period).
In 2016, out of the 46 Biogent Sentinel 2 (BGS2) trap-nights, 21 were adult-positive (adult trap positivity index, ATPI = 45.7%) and 30 of 276 of the Gravid Aedes traps (GAT) trap-nights were adult positive (ATPI = 10.9%). In 2017, out of the 108 BGS2 trap-nights, 55 were adult-positive (ATPI = 50.9%), and 42 of 324 of the GAT trap-nights were adult positive (ATPI = 13.0%). The number of adults captured peaked during the mid-July to mid-August 2017 period. All adults were identified to the species level in 2016 and five were not identified in 2017 because of sample deterioration. No mosquito (egg or adult) were found at sites one and two, located near Montréal, over the sampling period (2016–2017).
2.2. Molecular Identification of Eggs
A total of 184 unhatched mosquito eggs (n = 85 in 2016 and n = 278 in 2017) were identified at the national microbiology laboratory (NML) based on analysis of the CO1 gene:
Oc. japonicus (n = 41 in 2017),
Oc. triseriatus (n = 84 in 2016 and n = 57 in 2017),
Oc. punctor (n = 1 in 2016), and one invasive
Aedes spp. (n = 1 in 2017, with 100% identity with
Ae. aegypti, 99% identity with
Ae. albopictus) (
Table 1). The PCR and sequencing results of the CO1 gene on the
Aedes mosquito were reproducible, with the PCR and sequencing performed twice, obtaining the same results. In addition, sequence data arising from the amplification of the ITS1 and ITS2 regions, and the results of a real-time PCR assay were most consistent with
Ae. aegypti.
The Ae. aegypti egg came from an OVI deployed on 17 July 2017, at site no. 13, which was located in Saint-Armand (in Estrie, GPS coordinates: 45.0167, -73.0836) near the border between Québec and the state of Vermont. The OVI was on plot 3, located at the edge of a forested area, about 30 m away from the commercial road coming into Canada from the U.S., where trucks stop to pass border control. Overall, 85 eggs were captured during this OVI-night; five hatched and, among those, one could be identified by PCR as Oc. triseriatus (data not shown). Of the remaining 80 unhatched eggs sent to the national microbiology laboratory, 43 were too damaged to produce an amplification product, 36 were identified as Oc. triseriatus, and one was Ae. aegypti.
2.3. Mosquito Species Richness
Mosquito species richness indices (MSRI) were calculated yearly and monthly using egg and adult presence data: in total, 12 species were collected in 2016, 18 in 2017, and 24 for both 2016–2017 combined. Total MSRI were also calculated for each type of trap. BGS2 had a richness of eight in 2016, 17 in 2017, and 20 for both years combined. GAT had a richness of seven in 2016, five in 2017, and nine for both years combined. OVI had a richness of three in 2016, three in 2017, and four for both years combined (excluding species identified from occasional adult captures). The highest monthly MSRI were obtained in July 2017 (MSRI = 12) for BGS2, in August 2016 (MSRI = 6) for GAT, and in July 2017 (MSRI = 4) for OVI.
2.4. Distribution of AMOT Species
2.4.1. Spatial Distribution
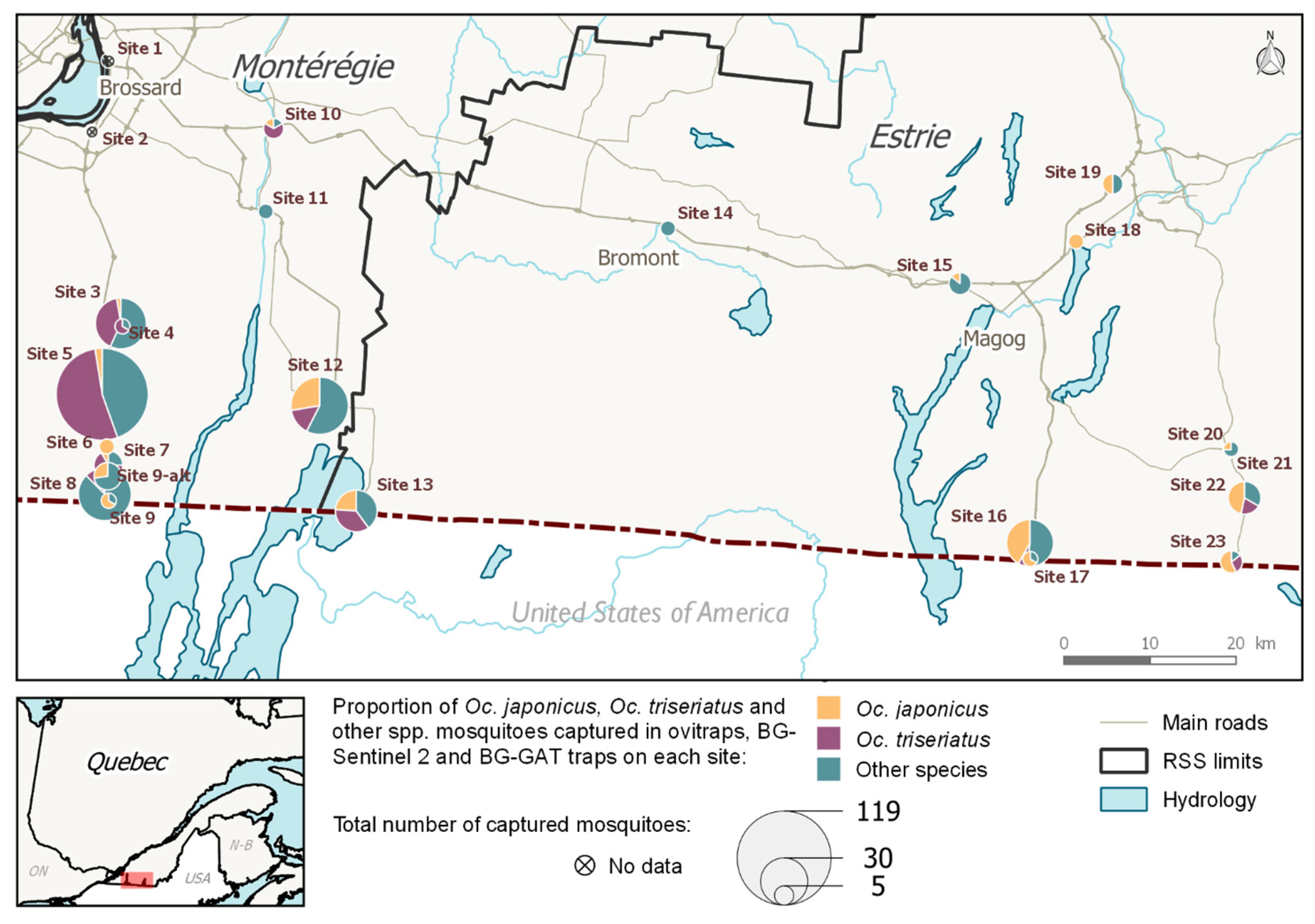
Of the 11 sites located along the eastern roads running through Estrie (sites nos. 13–23), three were
Oc. triseriatus dominant, and eight were
Oc. japonicus dominant.
Oc. japonicus was in a higher proportion at these four sites (
Figure 1). The proportion of adult mosquitoes collected by species varied from site to site (aggregated by trap and by site). Of the 13 sites located along the two western roads running through Montérégie (sites nos. 1–12, including 9 and 9-alt), both
Oc. triseriatus and
Oc. japonicus were present (D > 5%) in eight sites.
Oc. triseriatus occurred in higher proportion in four of the sites, while
Oc. japonicus occurred in higher proportion in two of the sites (
Figure 2).
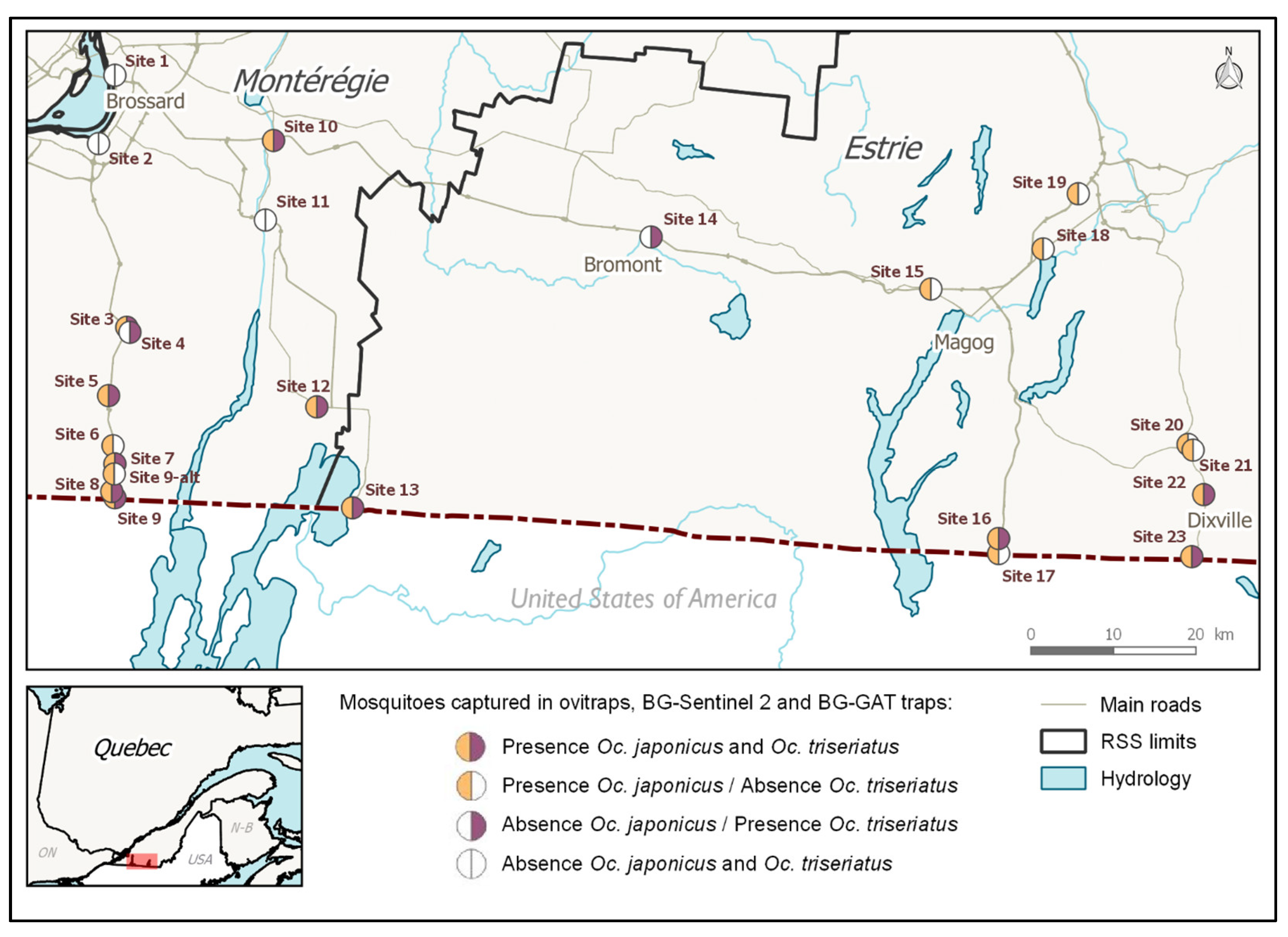
AMOT species presence distribution at 24 sites were estimated using combined egg and adult samples (
Figure 2).
Oc. japonicus’ was widespread: it was present in 19/24 or 79.2% of the sites sampled in 2016 and 2017, while
Oc. triseriatus had a moderate presence (13/24, 54.2%).
Oc. japonicus was present in 10/13 sites (76.9%) located along the two western roads running through Montérégie, while
Oc. triseriatus was present in nine of these sites (69.2%).
Oc. japonicus was present in 9/11 sites (81.8%) located along the eastern roads running through Estrie, and
Oc. triseriatus was present in four (36.4%) of the eastern sites. The co-occurrence of both species was observed in 8/13 sites (61.5%) located along the roads running through Montérégie and on 3 of the 11 sites (27.3%) located along the roads running through Estrie. This co-occurrence of
Oc. japonicus and
Oc. triseriatus was more common in the western part of our study region than in the eastern part.
2.4.2. Temporal Distribution
The monthly average abundance of adult mosquito peaked in August for both
Oc. triseriatus and
Oc. japonicus (
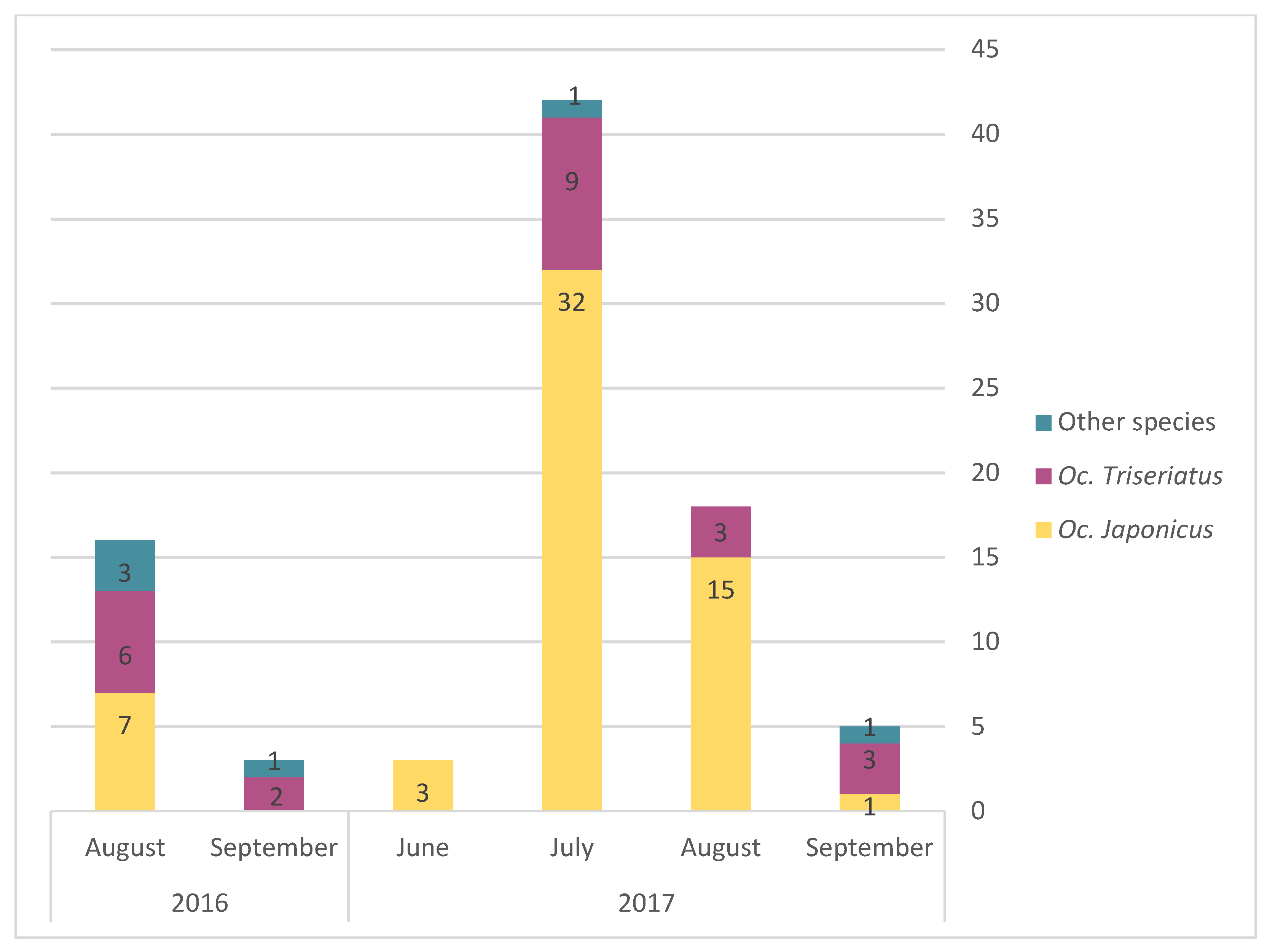
Figure 3). Egg presence for the different AMOT species varied following similar temporal trends, but with a peak in July (
Figure 4). However, we observed a slight difference in terms of the relative frequency for
Oc. triseriatus and
Oc. japonicus between August and September: the majority of eggs captured in September were
Oc. triseriatus in 2016 and 2017, while the majority of eggs captured in June to August were
Oc. japonicus in 2016 and 2017 (
Figure 4). The relative frequency of eggs peaked one month earlier than adults for both species (
Figure 4).
The temporal consistency of the number of eggs of Oc. japonicus at the same site between 2016 and 2017 was assessed (Fisher’s exact test (FET), p = 0.005). The null hypothesis, according to which the abundance of eggs in certain sites is the same from one year to the other, was rejected. In other words, the abundance of Oc. japonicus eggs at the same sites appears to vary from one year to another. The same test, done for Oc. triseriatus, gave a p-value of 0.0667, suggesting that the presence of Oc. triseriatus’ eggs in certain sites is relatively constant from one year to another. The same comparison was done for adults’ presence and the results gave a p-value of 0.49 for Oc. japonicus and a p-value of 0.05 for Oc. triseriatus. This, again, showed that the adults of Oc. triseriatus seemed to be present in a relatively stable manner on the same sites one year after another, whereas adults of Oc. japonicus were not.
2.5. Impact of Land Use on Mosquito Species Richness and Abundance
Regression analysis of species richness within sampling sites versus the percentage of developed zones and road segments (RR = 0.658,
p-value < 0.001 and 0.440,
p = 0.001, respectively) showed that species richness increased with decreasing development or shorter road segments. Species richness of the 24 sites was also positively associated with the degree of wetlands cover (RR = 1.12,
p < 0.001) (
Table 3).
The number of
Oc. triseriatus adults captured using GAT was negatively associated with the percentage of developed zones (RR = 0.457,
p = 0.002) (
Table 4a); with a greater number of
Oc. triseriatus being found in less developed zones. The number of
Oc. japonicus adults was negatively associated with the road segments variable (RR = 0.386,
p = 0.0158) (
Table 4b), with a greater number of adults of this species being found where there were fewer road segments (or shorter total road segment length). The presence of
Oc. triseriatus adults, captured using GAT and OVI eggs data, was statistically associated with developed areas (OR = 0.932,
p < 0.001) and road segments (OR = 0.932,
p = 0.03) (
Table 5a). The presence of
Oc. japonicus was statistically associated with the percentage of wetlands (OR = 1.013,
p = 0.02), developed areas (OR = 0.973,
p = 0.007), and road segments (OR = 0.923,
p = 0.004) (
Table 5b). While statistically significant, the magnitude of the effects detected is weak, with the OR values being close to one.
5. Conclusions
Since the Zika virus epidemics in the Americas, the arrival of invasive
Aedes spp. in North America has raised concerns, including for Québec’s public health authorities. Models based on temperature, precipitation, and winter survival capacity predict that under the current climate, southern Québec could be favorable for the temperate
Ae. albopictus’ establishment (least conservative scenario) and a much greater range expansion could occur for 2011 to 2040 [
83]. This study highlights the importance of choosing the appropriate methodology in line with the initial monitoring objectives. To optimize IMS monitoring for the identification of invasive
Aedes species introduction in southern Québec, based on what has been learned from AMOT species, all three types of traps used in this study should be used minimally from mid-July to mid-August in lightly-dense forested areas with ground transportation activities. Sites could be chosen according to the presence of
Oc. japonicus. For optimal identification of captured eggs, the fourth-instar larvae method should be employed. Our finding of the presence of
Ae. aegypti is not a sign of elevated disease risk for humans, but confirms the possibility of a mechanical introduction of an invasive
Aedes spp. through ground transportation activities in Québec. The repetitive findings of
Ae. albopictus’ eggs in the neighboring state of Vermont in 2019 and 2020 [
84,
85] illustrates that the threat of this species is real, and is progressively making its way northward. It is not a question of “if” or “how,” but rather “when” and “what” regarding the impact it is going to have. Finally, the presence of
Oc. triseriatus and
Oc. japonicus in southern Québec is of public health significance, since they are competent vectors of a number of zoonotic pathogens. While their presence was known to health authorities, their abundance and spread, as shown in this assay, was unexpectedly high. This should be considered in the province’s risk assessment plan of arboviruses, namely for LACV. Using AMOT species as a surrogate in future projects could prove to be a novel way of approaching the question about the risk of the emergence of vector-borne diseases resulting from climate changes in Canada, given the absence of
Ae. albopictus occurrence in the province. By providing indicators of suitable habitat for introduction, such an innovative public health preparedness approach could optimize sampling strategies when conducting mosquito surveillance of IMS under climate change. The present study adds to the increasing number of studies reporting range expansions of IMS, and demonstrates that ongoing monitoring, using diversified capture techniques to target a wide range of species, may provide useful information to public health with respect to the growing risk of emerging mosquito-borne diseases in southern Canada.


 ,
, 





{kind=link}
{kind=link}
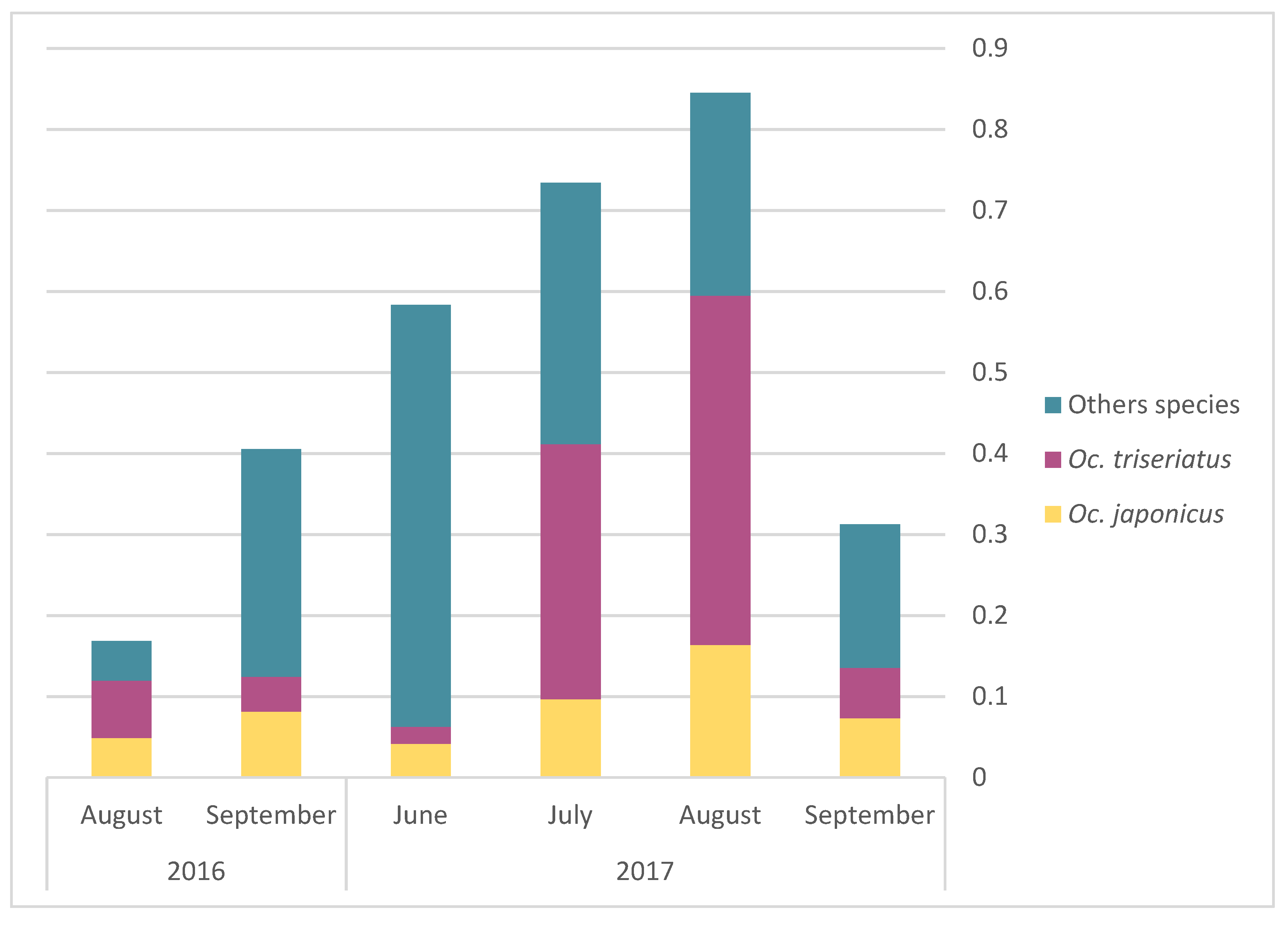{kind=link}
{kind=link}
{kind=link}