
Antimicrobial Resistance and Molecular Epidemiology of Staphylococcus aureus from Hunters and Hunting Dogs

 ,
,  , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Samples and Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing and Resistance Genes
2.3. Molecular Typing in S. aureus Isolates
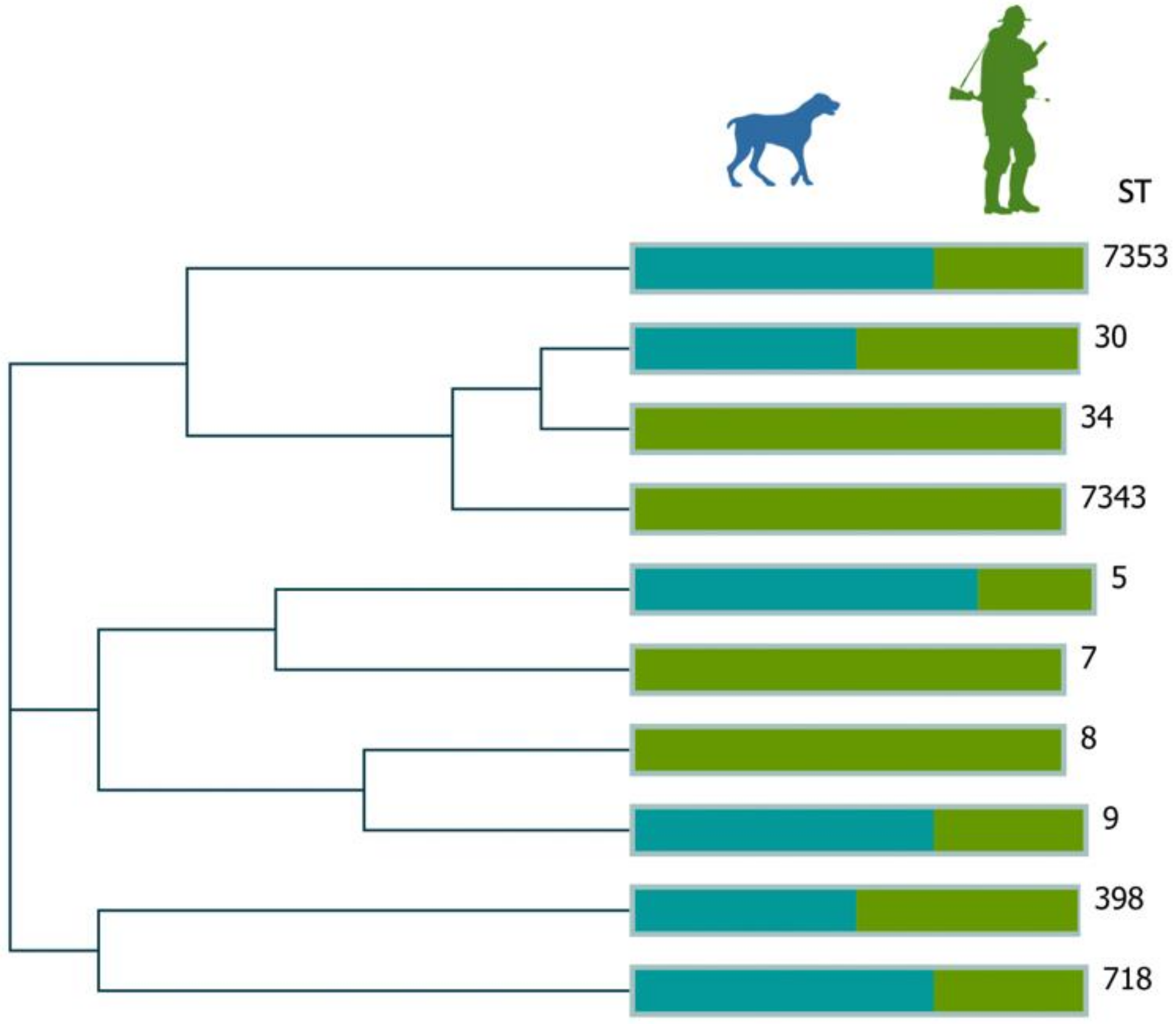
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.C.; Kananurak, A.; Tran, M.T.; Connolly, P.A.; Polage, C.R.; Iwase, T.; Bevins, C.L.; Underwood, M.A. Bacterial Colonization of the Hospitalized Newborn: Competition between Staphylococcus aureus and Staphylococcus epidermidis. Pediatr. Infect. Dis. J. 2019, 38, 682–686. [Google Scholar] [CrossRef] [PubMed]
- González-García, S.; Hamdan-Partida, A.; Bustos-Hamdan, A.; Bustos-Martínez, J. Factors of Nasopharynx that Favor the Colonization and Persistence of Staphylococcus aureus. In Pharynx-Diagnosis and Treatment; IntechOpen: Rijeka, Croatia, 2021; ISBN 1789856094. [Google Scholar]
- Sakr, A.; Brégeon, F.; Mège, J.-L.; Rolain, J.-M.; Blin, O. Staphylococcus aureus Nasal Colonization: An Update on Mechanisms, Epidemiology, Risk Factors, and Subsequent Infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Ahmad-Mansour, N.; Loubet, P.; Pouget, C.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.-P.; Molle, V. Staphylococcus aureus toxins: An update on their pathogenic properties and potential treatments. Toxins 2021, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Jevons, M.P. “Celbenin”-resistant staphylococci. Br. Med. J. 1961, 1, 124. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial resistance in the EU/EEA (EARS-Net) - Annual Epidemiological Report for 2019; European Centre for Disease Prevention and Control (ECDC): Stockholm, Sweden, 2020. [Google Scholar]
- WHO Regional Office for Europe and European Centre for Disease Prevention and Control. Surveillance of Antimicrobial Resistance in Europe, 2020 Data; WHO Regional Office for Europe and European Centre for Disease Prevention and Control: Copenhagen, Denmark, 2021. [Google Scholar]
- Shalaby, M.-A.W.; Dokla, E.M.E.; Serya, R.A.T.; Abouzid, K.A.M. Penicillin binding protein 2a: An overview and a medicinal chemistry perspective. Eur. J. Med. Chem. 2020, 199, 112312. [Google Scholar] [CrossRef]
- Schwendener, S.; Perreten, V. The bla and mec families of β-lactam resistance genes in the genera Macrococcus, Mammaliicoccus and Staphylococcus: An in-depth analysis with emphasis on Macrococcus. J. Antimicrob. Chemother. 2022, dkac107. [Google Scholar] [CrossRef]
- Boswihi, S.S.; Udo, E.E. Methicillin-resistant Staphylococcus aureus: An update on the epidemiology, treatment options and infection control. Curr. Med. Res. Pract. 2018, 8, 18–24. [Google Scholar] [CrossRef]
- Crespo-Piazuelo, D.; Lawlor, P.G. Livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) prevalence in humans in close contact with animals and measures to reduce on-farm colonisation. Ir. Vet. J. 2021, 74, 21. [Google Scholar] [CrossRef]
- Voss, A.; Loeffen, F.; Bakker, J.; Klaassen, C.; Wulf, M. Methicillin-resistant Staphylococcus aureus in Pig Farming. Emerg. Infect. Dis. 2005, 11, 1965–1966. [Google Scholar] [CrossRef]
- Loeffler, A.; Pfeiffer, D.U.; Lindsay, J.A.; Soares-Magalhaes, R.; Lloyd, D.H. Lack of transmission of methicillin-resistant Staphylococcus aureus (MRSA) between apparently healthy dogs in a rescue kennel. Vet. Microbiol. 2010, 141, 178–181. [Google Scholar] [CrossRef]
- Van Duijkeren, E.; Hengeveld, P.; Zomer, T.P.; Landman, F.; Bosch, T.; Haenen, A.; van de Giessen, A. Transmission of MRSA between humans and animals on duck and turkey farms. J. Antimicrob. Chemother. 2016, 71, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahibzada, S.; Abraham, S.; Coombs, G.W.; Pang, S.; Hernández-Jover, M.; Jordan, D.; Heller, J. Transmission of highly virulent community-associated MRSA ST93 and livestock-associated MRSA ST398 between humans and pigs in Australia. Sci. Rep. 2017, 7, 5273. [Google Scholar] [CrossRef] [PubMed]
- Van Balen, J.C.; Landers, T.; Nutt, E.; Dent, A.; Hoet, A.E. Molecular epidemiological analysis to assess the influence of pet-ownership in the biodiversity of Staphylococcus aureus and MRSA in dog- and non-dog-owning healthy households. Epidemiol. Infect. 2017, 145, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, T.; Kinoshita, Y.; Niwa, H.; Shinzaki, Y.; Tamura, N.; Hobo, S.; Kuwano, A. Meticillin-resistant Staphylococcus aureus colonisation and infection in Thoroughbred racehorses and veterinarians in Japan. Vet. Rec. 2016, 178, 473. [Google Scholar] [CrossRef]
- Bramble, M.; Morris, D.; Tolomeo, P.; Lautenbach, E. Potential Role of Pet Animals in Household Transmission of Methicillin-Resistant Staphylococcus aureus: A Narrative Review. Vector-Borne Zoonotic Dis. 2010, 11, 617–620. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Sanz, E.; Torres, C.; Ceballos, S.; Lozano, C.; Zarazaga, M. Clonal Dynamics of Nasal Staphylococcus aureus and Staphylococcus pseudintermedius in Dog-Owning Household Members. Detection of MSSA ST398. PLoS ONE 2013, 8, e69337. [Google Scholar] [CrossRef] [Green Version]
- Lozano, C.; Rezusta, A.; Ferrer, I.; Pérez-Laguna, V.; Zarazaga, M.; Ruiz-Ripa, L.; Revillo, M.J.; Torres, C. Staphylococcus pseudintermedius Human Infection Cases in Spain: Dog-to-Human Transmission. Vector-Borne Zoonotic Dis. 2017, 17, 268–270. [Google Scholar] [CrossRef]
- Boost, M.V.; O’Donoghue, M.M.; James, A. Prevalence of Staphylococcus aureus carriage among dogs and their owners. Epidemiol. Infect. 2008, 136, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Daley, P.; Bajgai, J.; Penney, C.; Williams, K.; Whitney, H.; Golding, G.R.; Weese, S. A cross sectional study of animal and human colonization with Methicillin-Resistant Staphylococcus aureus (MRSA) in an Aboriginal community. BMC Public Health 2016, 16, 595. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.C.; Worthing, K.A.; Gottlieb, T.; Ward, M.P.; Norris, J.M. Molecular characterization of community-associated methicillin-resistant Staphylococcus aureus from pet dogs. Zoonoses Public Health 2020, 67, 222–230. [Google Scholar] [CrossRef]
- Penna, B.; Silva, M.B.; Soares, A.E.R.; Vasconcelos, A.T.R.; Ramundo, M.S.; Ferreira, F.A.; Silva-Carvalho, M.C.; de Sousa, V.S.; Rabello, R.F.; Bandeira, P.T.; et al. Comparative genomics of MRSA strains from human and canine origins reveals similar virulence gene repertoire. Sci. Rep. 2021, 11, 4724. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, Y.; Koide, S.; Maeyama, Y.; Tamai, K.; Hayashi, W.; Tanaka, H.; Iimura, M.; Suzuki, M.; Nagano, Y.; Arakawa, Y.; et al. Predominance of methicillin-resistant Staphylococcus aureus SCCmec type II-CC5 and SCCmec type IV-CC1/CC8 among companion animal clinical isolates in Japan: Findings from phylogenetic comparison with human clinical isolates. J. Glob. Antimicrob. Resist. 2020, 20, 253–259. [Google Scholar] [CrossRef] [PubMed]
- CLSI Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 8.0.; EUCAST: Växjö, Sweden, 2018. [Google Scholar]
- Silva, V.; Caniça, M.; Ferreira, E.; Vieira-Pinto, M.; Saraiva, C.; Pereira, J.E.; Capelo, J.L.; Igrejas, G.; Poeta, P. Multidrug-Resistant Methicillin-Resistant Coagulase-Negative Staphylococci in Healthy Poultry Slaughtered for Human Consumption. Antibiotics 2022, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Etienne, J.; Vandenesch, F.; Nesme, X. Relationships between Staphylococcus aureus Genetic Background, Virulence Factors, agr Groups (Alleles), and Human Disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.-O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine Leukocidin—Producing Staphylococcus aureus in Primary Skin Infections and Pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nishifuji, K.; Sasaki, M.; Fudaba, Y.; Aepfelbacher, M.; Takata, T.; Ohara, M.; Komatsuzawa, H.; Amagai, M.; Sugai, M. Identification of the Staphylococcus aureus etd pathogenicity island which encodes a novel exfoliative toxin, ETD, and EDIN-B. Infect. Immun. 2002, 70, 5835–5845. [Google Scholar] [CrossRef] [Green Version]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Iyori, K.; Futagawa-Saito, K.; Hisatsune, J.; Yamamoto, M.; Sekiguchi, M.; Ide, K.; Son, W.; Olivry, T.; Sugai, M.; Fukuyasu, T. Staphylococcus pseudintermedius exfoliative toxin EXI selectively digests canine desmoglein 1 and causes subcorneal clefts in canine epidermis. Vet. Dermatol. 2011, 22, 319–326. [Google Scholar] [CrossRef]
- Futagawa-Saito, K.; Sugiyama, T.; Karube, S.; Sakurai, N.; Ba-Thein, W.; Fukuyasu, T. Prevalence and Characterization of Leukotoxin-Producing Staphylococcus intermedius in Isolates from Dogs and Pigeons. J. Clin. Microbiol. 2004, 42, 5324–5326. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shopsin, B.; Mathema, B.; Alcabes, P.; Said-Salim, B.; Lina, G.; Matsuka, A.; Martinez, J.; Kreiswirth, B.N. Prevalence of agr Sprecificity Groups amoung Staphylococcus aureus Strains Colonizing Children and Their Guardians. J. Clin. Microbiol. 2003, 57, 456–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; McClure, J.-A.; Elsayed, S.; Louie, T.; Conly, J.M. Novel Multiplex PCR Assay for Characterization and Concomitant Subtyping of Staphylococcal Cassette Chromosome mecTypes I to V in Methicillin-Resistant Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 5026–5033. [Google Scholar] [CrossRef] [Green Version]
- Laux, C.; Peschel, A.; Krismer, B. Staphylococcus aureus Colonization of the Human Nose and Interaction with Other Microbiome Members. Microbiol. Spectr. 2019, 7, 2. [Google Scholar] [CrossRef]
- Velasco, V.; Buyukcangaz, E.; Sherwood, J.S.; Stepan, R.M.; Koslofsky, R.J.; Logue, C.M. Characterization of Staphylococcus aureus from Humans and a Comparison with İsolates of Animal Origin, in North Dakota, United States. PLoS ONE 2015, 10, e0140497. [Google Scholar] [CrossRef] [Green Version]
- Bektas, S.; Obradovic, A.; Aljicevic, M.; Numanovic, F.; Hodzic, D.; Sporisevic, L. The frequency of community-acquired methicillin-resistant Staphylococcus aureus (CA-mrsa) among samples in institute for public health in canton Sarajevo. Mater. Sociomed. 2016, 28, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Kasela, M.; Grzegorczyk, A.; Nowakowicz-Dębek, B.; Malm, A. The Prevalence of Virulence Determinants and Antibiotic Resistance Patterns in Methicillin—Resistant Staphylococcus aureus in a Nursing Home in Poland. Pathogens 2021, 10, 427. [Google Scholar] [CrossRef]
- János, D.; Viorel, H.; Ionica, I.; Corina, P.; Tiana, F.; Roxana, D. Carriage of Multidrug Resistance Staphylococci in Shelter Dogs in Timisoara, Romania. Antibiotics 2021, 10, 801. [Google Scholar] [CrossRef]
- Ma, G.C.; Worthing, K.A.; Ward, M.P.; Norris, J.M. Commensal Staphylococci Including Methicillin-Resistant Staphylococcus aureus from Dogs and Cats in Remote New South Wales, Australia. Microb. Ecol. 2020, 79, 164–174. [Google Scholar] [CrossRef]
- Huang, T.; Chou, C. Methicillin-sensitive and methicillin-resistant Staphylococcus aureus strains and their toxin genes in the nostrils of dogs and workers at an animal shelter. J. Appl. Microbiol. 2019, 126, 1899–1909. [Google Scholar] [CrossRef]
- Coelho, C.; Torres, C.; Radhouani, H.; Pinto, L.; Lozano, C.; Gómez-Sanz, E.; Zaragaza, M.; Igrejas, G.; Poeta, P. Molecular Detection and Characterization of Methicillin-Resistant Staphylococcus aureus (MRSA) Isolates from Dogs in Portugal. Microb. Drug Resist. 2011, 17, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, M.; Bukar-Kolo, Y.M.; Geidam, Y.A.; Gulani, I.A. Phenotypic and genotypic detection of methicillin-resistant Staphylococcus aureus in hunting dogs in Maiduguri metropolitan, Borno State, Nigeria. Vet. World 2016, 9, 501–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabri, F.V.; Pinto, N.B.; de Mattos, M.d.S.F.; Rodrigues, R.F.; Shinohara, D.R.; Pereira, P.M.; Nishiyama, S.A.B.; Tognim, M.C.B. First report of oxacillin-susceptible mecA-positive Staphylococcus aureus in healthy dogs and their owners in southern Brazil. Prev. Vet. Med. 2021, 189, 105286. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.P.; Anderson, K.L.; Correa, M.T.; Lyman, R.; Ruffin, F.; Reller, L.B.; Fowler, V.G., Jr. Transmission of MRSA between Companion Animals and Infected Human Patients Presenting to Outpatient Medical Care Facilities. PLoS ONE 2011, 6, e26978. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.F.; Misic, A.M.; Morris, D.O.; Moss, J.T.; Tolomeo, P.; Beiting, D.P.; Nachamkin, I.; Lautenbach, E.; Rankin, S.C. Genome sequencing reveals strain dynamics of methicillin-resistant Staphylococcus aureus in the same household in the context of clinical disease in a person and a dog. Vet. Microbiol. 2015, 180, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-Y.; Chae, J.-C.; Han, J.-I.; Song, W.-K.; Lee, C.-M.; Park, H.-M. Distribution and epidemiological relatedness of methicillin-resistant Staphylococcus aureus isolated from companion dogs, owners, and environments. J. Vet. Med. Sci. 2020, 82, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Sahin-Tóth, J.; Kovács, E.; Tóthpál, A.; Juhász, J.; Forró, B.; Bányai, K.; Havril, K.; Horváth, A.; Ghidán, Á.; Dobay, O. Whole genome sequencing of coagulase positive staphylococci from a dog-and-owner screening survey. PLoS ONE 2021, 16, e0245351. [Google Scholar] [CrossRef]
- Effelsberg, N.; Stegger, M.; Peitzmann, L.; Altinok, O.; Coombs, G.W.; Pichon, B.; Kearns, A.; Randad, P.R.; Heaney, C.D.; Bletz, S. Global Epidemiology and Evolutionary History of Staphylococcus aureus ST45. J. Clin. Microbiol. 2022, 59, e02198-20. [Google Scholar] [CrossRef]
- Asanin, J.; Misic, D.; Aksentijevic, K.; Tambur, Z.; Rakonjac, B.; Kovacevic, I.; Spergser, J.; Loncaric, I. Genetic Profiling and Comparison of Human and Animal Methicillin-Resistant Staphylococcus aureus (MRSA) Isolates from Serbia. Antibiotics 2019, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Silva, V.; Ferreira, E.; Manageiro, V.; Reis, L.; Tejedor-Junco, M.T.; Sampaio, A.; Capelo, J.L.; Caniça, M.; Igrejas, G.; Poeta, P. Distribution and Clonal Diversity of Staphylococcus aureus and Other Staphylococci in Surface Waters: Detection of ST425-t742 and ST130-t843 mecC-Positive MRSA Strains. Antibiotics 2021, 10, 1416. [Google Scholar] [CrossRef]
- Luzzago, C.; Locatelli, C.; Franco, A.; Scaccabarozzi, L.; Gualdi, V.; Viganò, R.; Sironi, G.; Besozzi, M.; Castiglioni, B.; Lanfranchi, P.; et al. Clonal diversity, virulence-associated genes and antimicrobial resistance profile of Staphylococcus aureus isolates from nasal cavities and soft tissue infections in wild ruminants in Italian Alps. Vet. Microbiol. 2014, 170, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.; Matuszewska, M.; Jia, M.; Zhou, J.; Ba, X.; Duan, J.; Zhang, C.; Zhao, J.; Tao, M.; Fan, J. A Survey of Chinese Pig Farms and Human Healthcare Isolates Reveals Separate Human and Animal Methicillin-Resistant Staphylococcus aureus Populations. Adv. Sci. 2021, 9, 2103388. [Google Scholar] [CrossRef] [PubMed]
- Driebe, E.M.; Sahl, J.W.; Roe, C.; Bowers, J.R.; Schupp, J.M.; Gillece, J.D.; Kelley, E.; Price, L.B.; Pearson, T.R.; Hepp, C.M. Using whole genome analysis to examine recombination across diverse sequence types of Staphylococcus aureus. PLoS ONE 2015, 10, e0130955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, B.; Appelen, L.; Franck, B.; den Heijer, C.D.J.; Bartholomeeusen, S.; Coenen, S. Staphylococcus aureus, including meticillin-resistant Staphylococcus aureus, among general practitioners and their patients: A cross-sectional study. PLoS ONE 2015, 10, e0140045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurita, J.; Barba, P.; Ortega-Paredes, D.; Mora, M.; Rivadeneira, S. Local circulating clones of Staphylococcus aureus in Ecuador. Braz. J. Infect. Dis. 2016, 20, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Goudarzi, H.; Goudarzi, M.; Sabzehali, F.; Fazeli, M.; Salimi Chirani, A. Genetic analysis of methicillin-susceptible Staphylococcus aureus clinical isolates: High prevalence of multidrug-resistant ST239 with strong biofilm-production ability. J. Clin. Lab. Anal. 2020, 34, e23494. [Google Scholar] [CrossRef]
- Silva, V.; Lopes, A.F.; Soeiro, V.; Caniça, M.; Manageiro, V.; Pereira, J.E.; Maltez, L.; Capelo, J.L.; Igrejas, G.; Poeta, P. Nocturnal Birds of Prey as Carriers of Staphylococcus aureus and Other Staphylococci: Diversity, Antimicrobial Resistance and Clonal Lineages. Antibiotics 2022, 11, 240. [Google Scholar] [CrossRef]
- Silva, V.; Almeida, F.; Carvalho, J.A.; Castro, A.P.; Ferreira, E.; Manageiro, V.; Tejedor-Junco, M.T.; Caniça, M.; Igrejas, G.; Poeta, P. Emergence of community-acquired methicillin-resistant Staphylococcus aureus EMRSA-15 clone as the predominant cause of diabetic foot ulcer infections in Portugal. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 179–186. [Google Scholar] [CrossRef]
- Silva, V.; Alfarela, C.; Caniça, M.; Manageiro, V.; Nóvoa, M.; Leiva, B.; Kress, M.; Capelo, J.L.; Poeta, P.; Igrejas, G. A One Health Approach Molecular Analysis of Staphylococcus aureus Reveals Distinct Lineages in Isolates from Miranda Donkeys (Equus asinus) and Their Handlers. Antibiotics 2022, 11, 374. [Google Scholar] [CrossRef]
- Papadimitriou-Olivgeris, M.; Drougka, E.; Fligou, F.; Dodou, V.; Kolonitsiou, F.; Filos, K.S.; Anastassiou, E.D.; Petinaki, E.; Marangos, M.; Spiliopoulou, I. Spread of Tst–Positive Staphylococcus aureus Strains Belonging to ST30 Clone among Patients and Healthcare Workers in Two Intensive Care Units. Toxins 2017, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.-Y.; Huang, Y.-C. Livestock-associated meticillin-resistant Staphylococcus aureus in Asia: An emerging issue? Int. J. Antimicrob. Agents 2015, 45, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Peeters, L.E.J.; Argudín, M.A.; Azadikhah, S.; Butaye, P. Antimicrobial resistance and population structure of Staphylococcus aureus recovered from pigs farms. Vet. Microbiol. 2015, 180, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yu, X.; Tao, X.; Zhang, J.; Zhang, B.; Dong, R.; Xue, C.; Grundmann, H.; Zhang, J. Staphylococcus aureus ST398 from slaughter pigs in northeast China. Int. J. Med. Microbiol. 2014, 304, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.P.; Wan, M.T.; Chen, M.M.; Su, H.Y.; Lauderdale, T.L.; Chou, C.C. Molecular characterization and clonal genetic diversity of methicillin-resistant Staphylococcus aureus of pig origin in Taiwan. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.; Fang, Y.-P.; Chang, Y.-F.; Wu, T.-H.; Yang, Y.-Y.; Huang, Y.-C. Comparison of molecular epidemiology of bloodstream methicillin-resistant Staphylococcus aureus isolates between a new and an old hospital in central Taiwan. Int. J. Infect. Dis. 2019, 79, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Sinlapasorn, S.; Lulitanond, A.; Angkititrakul, S.; Chanawong, A.; Wilailuckana, C.; Tavichakorntrakool, R.; Chindawong, K.; Seelaget, C.; Krasaesom, M.; Chartchai, S.; et al. SCCmec IX in meticillin-resistant Staphylococcus aureus and meticillin-resistant coagulase-negative staphylococci from pigs and workers at pig farms in Khon Kaen, Thailand. J. Med. Microbiol. 2015, 64, 1087–1093. [Google Scholar] [CrossRef] [Green Version]
- Windahl, U.; Ågren, J.; Holst, B.S.; Börjesson, S. Colonization with methicillin-resistant Staphylococcus pseudintermedius in multi-dog households: A longitudinal study using whole genome sequencing. Vet. Microbiol. 2016, 189, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Yarbrough, M.L.; Lainhart, W.; Burnham, C.A.D. Epidemiology, Clinical Characteristics, and Antimicrobial Susceptibility Profiles of Human Clinical Isolates of Staphylococcus intermedius Group. J. Clin. Microbiol. 2022, 56, e01788–e01817. [Google Scholar] [CrossRef] [Green Version]
- Cuny, C.; Layer, F.; Hansen, S.; Werner, G.; Witte, W. Nasal Colonization of Humans with Occupational Exposure to Raw Meat and to Raw Meat Products with Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus. Toxins 2019, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Tavares, A.; Miragaia, M.; Rolo, J.; Coelho, C.; de Lencastre, H. High prevalence of hospital-associated methicillin-resistant Staphylococcus aureus in the community in Portugal: Evidence for the blurring of community–hospital boundaries. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1269–1283. [Google Scholar] [CrossRef]
- Strauß, L.; Stegger, M.; Akpaka, P.E.; Alabi, A.; Breurec, S.; Coombs, G.; Egyir, B.; Larsen, A.R.; Laurent, F.; Monecke, S.; et al. Origin, evolution, and global transmission of community-acquired Staphylococcus aureus ST8. Proc. Natl. Acad. Sci. USA 2017, 114, E10596–E10604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ripa, L.; Simón, C.; Ceballos, S.; Ortega, C.; Zarazaga, M.; Torres, C.; Gómez-Sanz, E.S. pseudintermedius and S. aureus lineages with transmission ability circulate as causative agents of infections in pets for years. BMC Vet. Res. 2021, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.D.; McLaughlin, R.E.; Whiteaker, J.D.; Ambler, J.E.; Alm, R.A. Molecular characterization of MRSA isolates bracketing the current EUCAST ceftaroline-susceptible breakpoint for Staphylococcus aureus: The role of PBP2a in the activity of ceftaroline. J. Antimicrob. Chemother. 2015, 70, 2488–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcante, F.S.; Abad, E.D.; Lyra, Y.C.; Saintive, S.B.; Ribeiro, M.; Ferreira, D.C.; dos Santos, K.R.N. High prevalence of methicillin resistance and PVL genes among Staphylococcus aureus isolates from the nares and skin lesions of pediatric patients with atopic dermatitis. Braz. J. Med. Biol. Res. 2015, 48, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Hirose, M.; Aung, M.S.; Fukuda, A.; Yahata, S.; Fujita, Y.; Saitoh, M.; Hirose, Y.; Urushibara, N.; Kobayashi, N. Antimicrobial Resistance and Molecular Epidemiological Characteristics of Methicillin-Resistant and Susceptible Staphylococcal Isolates from Oral Cavity of Dental Patients and Staff in Northern Japan. Antibiotics 2021, 10, 1316. [Google Scholar] [CrossRef]
- Ruimy, R.; Angebault, C.; Djossou, F.; Dupont, C.; Epelboin, L.; Jarraud, S.; Armand Lefevre, L.; Bes, M.; Elena Lixandru, B.; Bertine, M.; et al. Are host genetics the predominant determinant of persistent nasal Staphylococcus aureus carriage in humans? J. Infect. Dis. 2010, 202, 924–934. [Google Scholar] [CrossRef] [Green Version]
- Van der Mee-Marquet, N.; Corvaglia, A.-R.; Valentin, A.-S.; Hernandez, D.; Bertrand, X.; Girard, M.; Kluytmans, J.; Donnio, P.-Y.; Quentin, R.; François, P. Analysis of prophages harbored by the human-adapted subpopulation of Staphylococcus aureus CC398. Infect. Genet. Evol. 2013, 18, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Cuny, C.; Abdelbary, M.; Layer, F.; Werner, G.; Witte, W. Prevalence of the immune evasion gene cluster in Staphylococcus aureus CC398. Vet. Microbiol. 2015, 177, 219–223. [Google Scholar] [CrossRef]
- Stegger, M.; Liu, C.M.; Larsen, J.; Soldanova, K.; Aziz, M.; Contente-Cuomo, T.; Petersen, A.; Vandendriessche, S.; Jiménez, J.N.; Mammina, C.; et al. Rapid Differentiation between Livestock-Associated and Livestock-Independent Staphylococcus aureus CC398 Clades. PLoS ONE 2013, 8, e79645. [Google Scholar] [CrossRef]
- Argudín, M.A.; Deplano, A.; Vandendriessche, S.; Dodémont, M.; Nonhoff, C.; Denis, O.; Roisin, S. CC398 Staphylococcus aureus subpopulations in Belgian patients. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 911–916. [Google Scholar] [CrossRef]
- Lekkerkerk, W.S.N.; Van Wamel, W.J.B.; Snijders, S.V.; Willems, R.J.; Van Duijkeren, E.; Broens, E.M.; Wagenaar, J.A.; Lindsay, J.A.; Vos, M.C. What Is the Origin of Livestock-Associated Methicillin-Resistant Staphylococcus aureus Clonal Complex 398 Isolates from Humans without Livestock Contact? An Epidemiological and Genetic Analysis. J. Clin. Microbiol. 2015, 53, 1836–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, I.M.; de Oliveira Ferreira, E.; de Araújo Penna, B. Dogs as reservoir of methicillin resistant coagulase negative staphylococci strains—A possible neglected risk. Microb. Pathog. 2019, 135, 103616. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, V.M.; Williams, N.J.; Pinchbeck, G.; Corless, C.E.; Shaw, S.; McEwan, N.; Dawson, S.; Nuttall, T. Antimicrobial resistance and characterisation of staphylococci isolated from healthy Labrador retrievers in the United Kingdom. BMC Vet. Res. 2014, 10, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phumthanakorn, N.; Prapasarakul, N.; Yindee, J.; Gronsang, D. Frequency, Distribution, and Antimicrobial Resistance of Coagulase-Negative Staphylococci Isolated from Clinical Samples in Dogs and Cats. Microb. Drug Resist. 2021, 28, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, X.; Liang, J.; Li, Q.; Lin, H.; Lin, C.; Liu, H.; Zhou, D.; Lu, W.; Sun, Z.; et al. Characterization of florfenicol resistance genes in the coagulase-negative Staphylococcus (CoNS) isolates and genomic features of a multidrug-resistant Staphylococcus lentus strain H29. Antimicrob. Resist. Infect. Control 2021, 10, 9. [Google Scholar] [CrossRef]
- Monecke, S.; Müller, E.; Schwarz, S.; Hotzel, H.; Ehricht, R. Rapid microarray-based identification of different mecA alleles in staphylococci. Antimicrob. Agents Chemother. 2012, 56, 5547–5554. [Google Scholar] [CrossRef] [Green Version]
- Nemeghaire, S.; Argudín, M.A.; Feßler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef]


{kind=link}
| Isolate | Host | Species | Molecular Typing | Antimicrobial Resistance | Virulence Factors | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ST (CC) | spa | SCCmec | agr | Phenotype | Genotype | IEC system | Other genes | ||||
| Case 1 | VS3182 | Hunter 1 | S. aureus | 7353 | t10042 | - | NT | PEN | blaZ | - | hla, hlb, hld |
| VS3183 | Dog 1 | S. aureus | 7353 | t10042 | - | NT | PEN | blaZ | E | hla, hld | |
| VS3184 | Dog 2 | S. aureus | 7353 | t10042 | - | NT | PEN | blaZ | E | hla, hld | |
| VS3185 | Dog 2 | S. pseudintermedius | - | - | - | - | PEN, FOX | mecA | lukS/F-I, siet | ||
| VS3186 | Dog 3 | S. cohnii | - | - | - | - | PEN | mecA | |||
| Case 2 | VS3187 | Hunter 2 | S. aureus | 30 (30) | t012 | - | III | Susceptible | - | E | hla, hld, tst |
| VS3188 | Dog 1 | S. aureus | 30 (30) | t012 | - | III | Susceptible | - | - | hla, hlb, hld, tst | |
| Case 3 | VS3189 | Hunter 3 | S. aureus | 9 (9) | t2922 | N.T. | II | PEN, FOX, CN, TOB, KAN, CD, TET, FD | mecA, blaZ, aac(6’)-Ie-aph(2’’)-Ia, aph(3′)-IIIa, lnuB, vgaA, tetM, tetK | - | hla, hlb, hld |
| VS3190 | Dog 1 | S. aureus | 9 (9) | t2922 | N.T. | II | PEN, FOX, CN, TOB, KAN, CD, TET, FD, C | mecA, blaZ, aac(6’)-Ie-aph(2’’)-Ia, aph(3′)-IIIa, vgaA, lnuB, tetM, tetK, catp221 | - | hla, hlb, hld | |
| VS3191 | Dog 2 | S. aureus | 9 (9) | t2922 | N.T. | II | PEN, FOX, CN, TOB, KAN, CD, TET, FD, C | mecA, blaZ, aac(6’)-Ie-aph(2’’)-Ia, aph(3′)-IIIa, vgaA, lnuB, tetM, tetK, catp221 | - | hla, hlb, hld | |
| VS3192 | Dog 5 | S. lentus | - | - | - | - | PEN | mecA | - | ||
| Case 4 | VS3193 | Hunter 4 | S. aureus | 8 (8) | t121 | IV | I | PEN, FOX | mecA, blaZ, | - | hla, hlb, hld |
| VS3194 | S. lentus | - | - | PEN | mecA | ||||||
| VS3195 | Dog 1 | S. aureus | 5 (5) | t179 | IV | II | PEN, FOX, ERY | mecA, blaZ ermC, | E | hla, hld | |
| VS3196 | S. pseudintermedius | - | - | - | - | PEN, FOX | mecA, blaZ | - | lukS/F-I | ||
| VS3197 | Dog 2 | S. aureus | 5 (5) | t179 | IV | II | PEN, FOX, ERY | mecA, blaZ, ermC, | E | hla, hld | |
| VS3198 | S. pseudintermedius | - | - | - | - | PEN, FOX, CN, TOB, KAN | mecA, blaZ, aac(6’)-Ie-aph(2’’)-Ia, aph(3′)-IIIa, str | - | lukS/F-I, siet | ||
| VS3199 | Dog 3 | S. aureus | 5 (5) | t179 | IV | II | PEN, FOX, ERY | mecA, blaZ | E | hla, hld | |
| VS3200 | S. pseudintermedius | - | - | - | - | PEN, FOX | mecA, blaZ | - | lukS/F-I, siet | ||
| Case 5 | VS3201 | Dog 2 | S. aureus | 718 | t11333 | IV | II | PEN, FOX, ERY, CD | mecA, blaZ, ermC, | E | hld |
| VS3202 | Dog 3 | S. aureus | 718 | t11333 | IV | II | PEN, FOX, ERY | mecA, blaZ, ermC, | E | hld | |
| Case 6 | VS3203 | Dog1 | S. aureus | 398 (398) | t5635 | - | I | Susceptible | - | - | hla, hld |
| VS3204 | Dog 3 | S. sciuri | - | - | - | - | PEN | mecA, blaZ | - | ||
| Case 7 | VS3205 | Hunter 5 | S. aureus | 34 (30) | t166 | - | III | PEN, ERY | blaZ, ermC | E | hld |
| VS3206 | Dog1 | S. pseudintermedius | - | - | - | - | PEN, FOX | mecA, blaZ | - | lukS/F-I, siet | |
| VS3207 | Dog2 | S. lentus | - | - | - | - | PEN | mecA | - | ||
| Case 8 | VS3208 | Hunter 6 | S. aureus | 718 | t11333 | II | Susceptible | - | E | hla, hld | |
| VS3209 | Dog 1 | S. vitulinus | - | - | PEN | mecA | |||||
| Case 9 | VS3210 | Hunter 7 | S. aureus | 398 (398) | t5635 | - | I | PEN, ERY | blaZ, ermT | scn | hla, hld |
| VS3211 | Dog1 | S. lentus | - | - | - | PEN, CN, KAN, CD, C | mecA, aac(6’)-Ie-aph(2’’)-Ia, aph(3′)-IIIa, mphC, catp221 | - | hla | ||
| Case 10 | VS3212 | Hunter 8 | S. aureus | 7343 | t012 | N.T. | III | PEN, FOX | mecA, blaZ | - | hla, hlb, hld |
| VS3213 | Dog | S. sciuri | - | - | PEN | mecA | |||||
| Case 11 | VS3214 | Hunter 9 | S. aureus | 5 (5) | t179 | IV | II | PEN, FOX, ERY, | mecA, blaZ, ermA | E | hla, hld |
| VS3215 | Dog 1 | S. lentus | - | - | - | - | PEN | mecA, blaZ | - | ||
| VS3216 | Dog 2 | S. lentus | - | - | - | - | PEN | mecA, blaZ, | - | ||
| Case 12 | VS3217 | Hunter 10 | S. aureus | 7 | t091 | - | I | PEN | blaZ | G | hla, hld |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.; Caniça, M.; Manageiro, V.; Vieira-Pinto, M.; Pereira, J.E.; Maltez, L.; Poeta, P.; Igrejas, G. Antimicrobial Resistance and Molecular Epidemiology of Staphylococcus aureus from Hunters and Hunting Dogs. Pathogens 2022, 11, 548. https://doi.org/10.3390/pathogens11050548
Silva V, Caniça M, Manageiro V, Vieira-Pinto M, Pereira JE, Maltez L, Poeta P, Igrejas G. Antimicrobial Resistance and Molecular Epidemiology of Staphylococcus aureus from Hunters and Hunting Dogs. Pathogens. 2022; 11(5):548. https://doi.org/10.3390/pathogens11050548
Chicago/Turabian StyleSilva, Vanessa, Manuela Caniça, Vera Manageiro, Madalena Vieira-Pinto, José Eduardo Pereira, Luís Maltez, Patrícia Poeta, and Gilberto Igrejas. 2022. "Antimicrobial Resistance and Molecular Epidemiology of Staphylococcus aureus from Hunters and Hunting Dogs" Pathogens 11, no. 5: 548. https://doi.org/10.3390/pathogens11050548