
Distribution and Current State of Molecular Genetic Characterization in Pathogenic Free-Living Amoebae

 ,
,  ,
,  and
and
Abstract
:1. Introduction
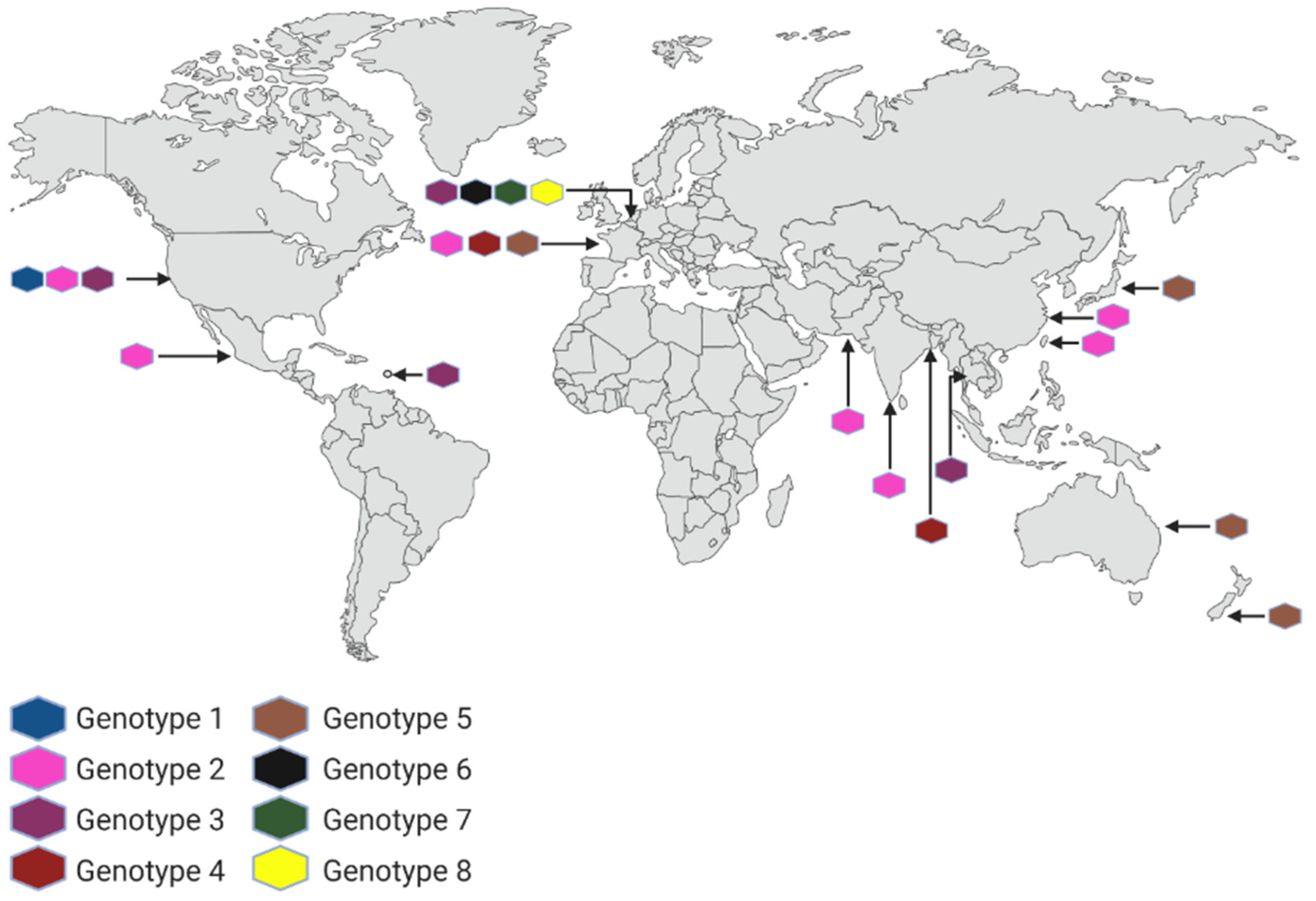
2. Naegleria fowleri
3. Acanthamoeba spp.
4. Balamuthia mandrillaris

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Symptoms | Time of Diagnosis | Reference |
|---|---|---|---|
| 13 | Bitemporal abdominal pain and headache | 8 days | [125] |
| 69 | Chronic nasal rash and ring lesions on the right frontal lobe | 19 days | [122] |
| 74 | Drowsiness and high temperature | 10 days | [126] |
| 69 | Confusion and expressive dysphasia | N/A | [102] |
| 71 | Partial spasm from the left thigh and fever | 10 days | [106] |
| 13 | Fever, vomiting, headaches, and right esotropia | 2 years | [115] |
| 51 | Fever, headache, altered mental status, disorientation, and seizures | 3 weeks | [127] |
| 9 | Fever, headache, altered state of consciousness, and blurred vision | 2 days | [16] |
| 60 | visual field loss, trouble walking, and fever | N/A | [128] |
| 50 | Headache, dizziness, dysarthria, and aphasia | N/A | [129] |
| 17 | Swelling in the left nostril, headache, weight loss, and fever | N/A | [116] |
| 54 | Numbness, weakness in left extremities | 44 days | [130] |
| 37 | Dizziness | 9 days | [131] |
| 15 | Fever and altered mental status | N/A | [132] |
| 4 | Epilepsy | N/A | [133] |
5. Sappinia pedata
6. Vermamoeba vermiformis
7. The FLA Molecular Epidemiology and the Genetic Changes
8. Advances in Genomic Epidemiology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Visvesvara, G.S. Infections with free-living amebae. Handb. Clin. Neurol. 2013, 114, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Król-Turmińska, K.; Olender, A. Human infections caused by free-living amoebae. Ann. Agric. Environ. Med. 2017, 24, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Armand, B.; Motazedian, M.H.; Asgari, Q. Isolation and identification of pathogenic free-living amoeba from surface and tap water of shiraz city using morphological and molecular methods. Parasitol. Res. 2016, 115, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Delafont, V.; Rodier, M.H.; Maisonneuve, E.; Cateau, E. Vermamoeba vermiformis: A free-living amoeba of interest. Microb. Ecol. 2018, 76, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Baquero, R.A.; Reyes-Batlle, M.; Nicola, G.G.; Martín-Navarro, C.M.; López-Arencibia, A.; Guillermo Esteban, J.; Valladares, B.; Martínez-Carretero, E.; Piñero, J.E.; Lorenzo-Morales, J. Presence of potentially pathogenic free-living amoebae strains from well water samples in Guinea-Bissau. Pathog. Glob. Health 2014, 108, 206–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelkha, N.; Jardot, P.; Moussaoui, I.; Levasseur, A.; La Scola, B.; Colson, P. Core gene-based molecular detection and identification of Acanthamoeba species. Sci. Rep. 2020, 10, 19–21. [Google Scholar] [CrossRef] [Green Version]
- Matos de Oliveira, Y.L.; Lima, E.T.S.; Rott, M.B.; Fernandes, R.P.M.; Jain, S.; de Aragão Batista, M.V.; Santana Dolabella, S. Occurrence, molecular diversity and pathogenicity of Acanthamoeba spp. isolated from Aquatic Environments of Northeastern Brazil. Int. J. Environ. Health Res. 2022, 1–11. [Google Scholar] [CrossRef]
- Coronado-Velázquez, D.; Silva-Olivares, A.; Castro-Muñozledo, F.; Lares-Jiménez, L.F.; Rodríguez-Anaya, L.Z.; Shibayama, M.; Serrano-Luna, J. Acanthamoeba mauritaniensis genotype T4D: An environmental isolate displays pathogenic behavior. Parasitol. Int. 2020, 74, 102002. [Google Scholar] [CrossRef] [PubMed]
- Kalra, S.K.; Sharma, P.; Shyam, K.; Tejan, N.; Ghoshal, U. Acanthamoeba and its pathogenic role in granulomatous amebic encephalitis. Exp. Parasitol. 2020, 208, 107788. [Google Scholar] [CrossRef]
- Hasni, I.; Andréani, J.; Colson, P.; La Scola, B. Description of virulent factors and horizontal gene transfers of keratitis-associated amoeba Acanthamoeba triangularis by genome analysis. Pathogens 2020, 9, 217. [Google Scholar] [CrossRef]
- Prithiviraj, S.R.; Rajapandian, S.G.K.; Gnanam, H.; Gunasekaran, R.; Mariappan, P.; Singh, S.S.; Prajna, L. Clinical presentations, genotypic diversity and phylogenetic analysis of Acanthamoeba species causing keratitis. J. Med. Microbiol. 2020, 69, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Juárez, M.M.; Tártara, L.I.; Cid, A.G.; Real, J.P.; Bermúdez, J.M.; Rajal, V.B.; Palma, S.D. Acanthamoeba in the eye, can the parasite hide even more? latest developments on the disease. Cont. Lens Anterior Eye 2018, 41, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castillo, M.; Cárdenas-Zúñiga, R.; Coronado-Velázquez, D.; Debnath, A.; Serrano-Luna, J.; Shibayama, M. Naegleria fowleri after 50 years: Is it a neglected pathogen? J. Med. Microbiol. 2016, 65, 885–896. [Google Scholar] [CrossRef]
- De Jonckheere, J.F. Origin and evolution of the worldwide distributed pathogenic amoeboflagellate Naegleria fowleri. Infect. Genet. Evol. 2011, 11, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Gharpure, R.; Bliton, J.; Goodman, A.; Ali, I.K.M.; Yoder, J.; Cope, J.R. Epidemiology and clinical characteristics of primary amebic meningoencephalitis caused by Naegleria fowleri: A global review. Clin. Infect. Dis. 2021, 73, E19–E27. [Google Scholar] [CrossRef]
- Yi, Z.; Zhong, J.; Wu, H.; Li, X.; Chen, Y.; Chen, H.; Yang, Y.; Yu, X. Balamuthia mandrillaris encephalitis in a child: Case report and literature review. Diagn. Microbiol. Infect. Dis. 2021, 100, 115180. [Google Scholar] [CrossRef]
- Lotonin, K.; Bondarenko, N.; Nassonova, E.; Rayko, M.; Smirnov, A. Balamuthia spinosa n. sp. (Amoebozoa, Discosea) from the brackish-water sediments of Nivå Bay (Baltic Sea, The Sound)—A novel potential vector of Legionella Pneumophila in the Environment. Parasitol. Res. 2022, 121, 713–724. [Google Scholar] [CrossRef]
- Scheid, P. Vermamoeba vermiformis—A free-living amoeba with public health and environmental health significance. Open Parasitol. J. 2019, 7, 40–47. [Google Scholar] [CrossRef]
- Cope, J.R.; Ali, L.K.; Visvesvara, G.S. Pathogenic and Opportunistic Free-Living Ameba Infections, 10th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9781416043904. [Google Scholar]
- Tang, P.; Croxen, M.A.; Hasan, M.R.; Hsiao, W.W.L.; Hoang, L.M. Infection control in the new age of genomic epidemiology. Am. J. Infect. Control 2017, 45, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Morales, A.; Balbin-Ramon, G.; Rabaan, A.; Sah, R.; Dhama, K.; Paniz-Mondolf, A.; Pagliano, P.; Esposito, S. Genomic epidemiology and its importance in the study of the COVID-19 pandemic. Infez. Med. 2020, 2, 139–142. [Google Scholar]
- Naegleria Fowleri, Illness & Symptoms. Available online: https://www.cdc.gov/parasites/naegleria/illness.html (accessed on 2 October 2022).
- Maciver, S.K.; Piñero, J.E.; Lorenzo-Morales, J. Is Naegleria fowleri an emerging parasite? Trends Parasitol. 2020, 36, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Tiewcharoen, S.; Junnu, V.; Sassa-Deepaeng, T.; Waiyawuth, W.; Ongrothchanakul, J.; Wankhom, S. Analysis of the 5.8S rRNA and internal transcribed spacers regions of the variant Naegleria fowleri Thai strain. Parasitol. Res. 2007, 101, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Wu, M.; Hu, B.C.; Chen, H.L.; Pan, J.R.; Ruan, W.; Yao, L.N. Identification and molecular typing of Naegleria fowleri from a patient with primary amebic meningoencephalitis in China. Int. J. Infect. Dis. 2018, 72, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, M.C.; Hsu, B.M.; Tao, C.W.; Lin, W.C.; Tsai, H.F.; Der Ji, D.; Shen, S.M.; Chen, J.S.; Shih, F.C.; Huang, Y.L. Identification and significance of Naegleria fowleri isolated from the hot spring which related to the first primary amebic meningoencephalitis (PAM) patient in Taiwan. Int. J. Parasitol. 2013, 43, 691–696. [Google Scholar] [CrossRef]
- Panda, A.; Khalil, S.; Mirdha, B.R.; Singh, Y.; Kaushik, S. Prevalence of Naegleria fowleri in environmental samples from northern part of India. PLoS ONE 2015, 10, e0137736. [Google Scholar] [CrossRef]
- Yoder, J.S.; Straif-bourgeois, S.; Roy, S.L.; Moore, T.A.; Visvesvara, G.S.; Ratard, R.C.; Hill, V.R.; Wilson, J.D.; Linscott, A.J.; Crager, R.; et al. Primary amebic meningoencephalitis deaths associated with sinus irrigation using contaminated tap water. Clin. Infect. Dis. 2012, 55, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Moussa, M.; de Jonckheere, J.F.; Guerlotté, J.; Richard, V.; Bastaraud, A.; Romana, M.; Talarmin, A. Survey of Naegleria fowleri in geothermal recreational waters of Guadeloupe (French west indies). PLoS ONE 2013, 8, e54414. [Google Scholar] [CrossRef] [Green Version]
- Cope, J.R.; Ratard, R.C.; Hill, V.R.; Sokol, T.; Causey, J.J.; Yoder, J.S.; Mirani, G.; Mull, B.; Mukerjee, K.A.; Narayanan, J.; et al. The first association of a primary amebic meningoencephalitis death with culturable Naegleria fowleri in tap water from a us treated public drinking water system. Clin. Infect. Dis. 2015, 60, e36–e42. [Google Scholar] [CrossRef]
- Sazzad, H.M.S.; Luby, S.P.; Sejvar, J.; Rahman, M.; Gurley, E.S.; Hill, V.; Murphy, J.L.; Roy, S.; Cope, J.R.; Ali, I.K.M. A case of primary amebic meningoencephalitis caused by Naegleria fowleri in Bangladesh. Parasitol. Res. 2019, 119, 339–344. [Google Scholar] [CrossRef]
- Vargas-Zepeda, J.; Gómez-Alcalá, A.V.; Vázquez-Morales, J.A.; Licea-Amaya, L.; De Jonckheere, J.F.; Lares-Villa, F. Successful treatment of Naegleria fowleri meningoencephalitis by using intravenous amphotericin b, fluconazole and rifampicin. Arch. Med. Res. 2005, 36, 83–86. [Google Scholar] [CrossRef]
- Aurongzeb, M.; Rashid, Y.; Habib, S.; Naqvi, A.; Khatoon, A.; Haq, S.A.; Azim, M.K.; Kaleem, I.; Bashir, S. Naegleria fowleri from Pakistan Has Type-2 Genotype. Iran. J. Parasitol. 2022, 17, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.K.; Mangrola, K.; Fitzpatrick, G.; Stockdale, K.; Matthias, L.; Ali, I.K.M.; Cope, J.R.; O’Laughlin, K.; Collins, S.; Beal, S.G.; et al. A case report of primary amebic meningoencephalitis in north Florida. IDCases 2021, 25, e01208. [Google Scholar] [CrossRef]
- Wang, Q.; Li, J.; Ji, J.; Yang, L.; Chen, L.; Zhou, R.; Yang, Y.; Zheng, H.; Yuan, J.; Li, L.; et al. A case of Naegleria fowleri related primary amoebic meningoencephalitis in China diagnosed by next-generation sequencing. BMC Infect. Dis. 2018, 18, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Ruan, W.; Zhang, L.; Hu, B.; Yang, X. Primary amebic meningoencephalitis: A case report. Korean J. Parasitol. 2019, 57, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vareechon, C.; Tarro, T.; Polanco, C.; Anand, V.; Pannaraj, P.S.; Dien Bard, J. Eight-year-old male with primary amebic meningoencephalitis. Open Forum Infect. Dis. 2019, 6, ofz349. [Google Scholar] [CrossRef]
- Moreira, L.R.; Rojas, L.Z.; Murillo, M.G.; Castro, S.E.M.; Sandí, E.A. Primary amebic meningoencephalitis related to groundwater in Costa Rica: Diagnostic confirmation of three cases and environmental investigation. Pathogens 2020, 9, 629. [Google Scholar] [CrossRef]
- Mubashir, H.; Zubair, S.M. Hemorrhage, cerebellar herniation, and cerebral venous thrombosis in a patient with Naegleria fowleri: A rare presentation. J. Med. Case Rep. Case Ser. 2021, 2, 1–4. [Google Scholar] [CrossRef]
- Huang, S.; Liang, X.; Han, Y.; Zhang, Y.; Li, X.; Yang, Z. A pediatric case of primary amoebic meningoencephalitis due to Naegleria fowleri diagnosed by next-generation sequencing of cerebrospinal fluid and blood samples. BMC Infect. Dis. 2021, 21, 1251. [Google Scholar] [CrossRef]
- Oncel, K.; Karaagac, L.; Dagcı, H.; Aykur, M. Real-time pcr confirmation of a fatal case of primary amoebic meningoencephalitis in Turkey caused by Naegleria fowleri or brain-eating amoeba. Acta Parasitol. 2022, 67, 697–704. [Google Scholar] [CrossRef]
- Pussard, M.; Pons, R. Morphologie de la paroi kystique et taxonomie du genre Acanthamoeba (protozoa, amoebida). Protistologica 1977, 13, 557–598. [Google Scholar]
- Stothard, D.R.; Schroeder-Diedrich, J.M.; Awwad, M.H.; Gast, R.J.; Ledee, D.R.; Rodriguez-Zaragoza, S.; Dean, C.L.; Fuerst, P.A.; Byers, T.J. The evolutionary history of the genus Acanthamoeba and the identification of eight new 18S rRNA gene sequence types. J. Eukaryot. Microbiol. 1998, 45, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Booton, G.C.; Kelly, D.J.; Chu, Y.; Seal, D.V.; Houang, E.; Lam, D.S.C.; Byers, T.J.; Fuerst, P.A. 18S ribosomal DNA typing and tracking of Acanthamoeba species isolates from corneal scrape specimens, contact lenses, lens cases, and home water supplies of Acanthamoeba keratitis patients in Hong Kong. J. Clin. Microbiol. 2002, 40, 1621–1625. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, P.A. Acanthamoeba and Free-Living Amoebae. Available online: https://u.osu.edu/acanthamoeba/acanthamoeba-sequence-types/ (accessed on 15 September 2022).
- Fuerst, P.A.; Booton, G.C. Species, Sequence types and alleles: Dissecting genetic variation in Acanthamoeba. Pathogens 2020, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Fritsche, T.R.; Romesh, K.; Schleifer, K.; Wagner, M.; Mu, D. Novel bacterial endosymbionts of Acanthamoeba spp. Related to the Paramecium caudatum Symbiont Caedibacter caryophilus. Environ. Microbiol. 1999, 1, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Hewett, M.K.; Robinson, B.S.; Monis, P.T.; Saint, C.P. Identification of a new Acanthamoeba 18S rRNA gene sequence type, corresponding to the species Acanthamoeba jacobsi Sawyer, Nerad and Visvesvara, 1992 (Lobosea: Acanthamoebidae). Acta Protozool. 2003, 42, 325–329. [Google Scholar]
- Corsaro, D.; Venditti, D. Phylogenetic evidence for a new genotype of Acanthamoeba (amoebozoa, Acanthamoebida). Parasitol. Res. 2010, 107, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Nuprasert, W.; Putaporntip, C.; Pariyakanok, L.; Jongwutiwes, S. Identification of a novel T17 genotype of Acanthamoeba from environmental isolates and T10 genotype causing keratitis in Thailand. J. Clin. Microbiol. 2010, 48, 4636–4640. [Google Scholar] [CrossRef] [Green Version]
- Qvarnstrom, Y.; Nerad, T.A.; Visvesvara, G.S. Characterization of a new pathogenic Acanthamoeba species, A. byersi n. Sp., isolated from a human with fatal amoebic encephalitis. J. Eukaryot. Microbiol. 2013, 60, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Magnet, A.; Henriques-Gil, N.; Galván-Diaz, A.L.; Izquiedo, F.; Fenoy, S.; Del Aguila, C. Novel Acanthamoeba 18S rRNA gene sequence type from an environmental isolate. Parasitol. Res. 2014, 113, 2845–2850. [Google Scholar] [CrossRef]
- Fuerst, P.A.; Booton, G.C.; Crary, M. Phylogenetic analysis and the evolution of the 18S rRNA Gene Typing System of Acanthamoeba. J. Eukaryot. Microbiol. 2015, 62, 69–84. [Google Scholar] [CrossRef]
- Tice, A.K.; Shadwick, L.L.; Fiore-donno, A.M.; Geisen, S.; Kang, S.; Schuler, G.A.; Spiegel, F.W.; Wilkinson, K.A.; Bonkowski, M.; Dumack, K.; et al. Expansion of the molecular and morphological diversity of Acanthamoebidae (centramoebida, amoebozoa) and identification of a novel life cycle type within the group. Biol. Direct 2016, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putaporntip, C.; Kuamsab, N.; Nuprasert, W.; Rojrung, R.; Pattanawong, U.; Tia, T.; Yanmanee, S.; Jongwutiwes, S. Analysis of Acanthamoeba genotypes from public freshwater sources in thailand reveals a new genotype, T23 Acanthamoeba bangkokensis sp. nov. Sci. Rep. 2021, 11, 17290. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.W.; Hsu, B.M. Isolation and identification of Acanthamoeba from Taiwan spring recreation areas using culture enrichment combined with PCR. Acta Trop. 2010, 115, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Muchesa, P.; Leifels, M.; Jurzik, L.; Hoorzook, K.B.; Barnard, T.G.; Bartie, C. Coexistence of free-living amoebae and bacteria in selected south african hospital water distribution systems. Parasitol. Res. 2017, 116, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Niyyati, M.; Saberi, R.; Latifi, A.; Lasjerdi, Z. Distribution of Acanthamoeba genotypes isolated from recreational and therapeutic geothermal water sources in southwestern Iran. Environ. Health Insights 2016, 10, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo-Morales, J.; Ortega-Rivas, A.; Martínez, E.; Khoubbane, M.; Artigas, P.; Periago, M.V.; Foronda, P.; Abreu-Acosta, N.; Valladares, B.; Mas-Coma, S. Acanthamoeba isolates belonging to T1, T2, T3, T4 and T7 genotypes from environmental freshwater samples in the Nile delta region, Egypt. Acta Trop. 2006, 100, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Rivera, W.L.; Adao, D.E.V. Identification of the 18S-ribosomal-DNA genotypes of Acanthamoeba isolates from the Philippines. Ann. Trop. Med. Parasitol. 2008, 102, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Possamai, C.O.; Loss, A.C.; Costa, A.O.; Falqueto, A.; Furst, C. Correction to: Acanthamoeba of three morphological groups and distinct genotypes exhibit variable and weakly inter-related physiological properties. Parasitol. Res. 2018, 117, 1995. [Google Scholar] [CrossRef] [Green Version]
- Orosz, E.; Kriskó, D.; Shi, L.; Sándor, G.L.; Kiss, H.J.; Seitz, B.; Nagy, Z.Z.; Szentmáry, N. Clinical course of Acanthamoeba keratitis by genotypes T4 and T8 in Hungary. Acta Microbiol. Immunol. Hung. 2019, 66, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Dendana, F.; Trabelsi, H.; Neiji, S.; Sellami, H.; Kammoun, S.; Makni, F.; Feki, J.; Cheikhrouhou, F.; Ayadi, A. Isolation and molecular identification of Acanthamoeba spp. from oasis water in Tunisia. Exp. Parasitol. 2018, 187, 37–41. [Google Scholar] [CrossRef]
- Karakavuk, M.; Aykur, M.; Şahar, E.A.; Karakuş, M.; Aldemir, D.; Döndüren, Ö.; Özdemir, H.G.; Can, H.; Gürüz, A.Y.; Dağcı, H.; et al. First time identification of Acanthamoeba genotypes in the cornea samples of wild birds; Is Acanthamoeba keratitis making the predatory birds a target? Exp. Parasitol. 2017, 183, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, T.; Hameed, A.; Muazzam, A.G.; Jung, S.Y.; Gul, A.; Matin, A. Isolation and molecular characterization of potentially pathogenic Acanthamoeba genotypes from diverse water resources including household drinking water from Khyber Pakhtunkhwa, Pakistan. Parasitol. Res. 2013, 112, 2925–2932. [Google Scholar] [CrossRef] [PubMed]
- Nagyová, V.; Nagy, A.; Timko, J. Morphological, Physiological and molecular biological characterisation of isolates from first cases of Acanthamoeba keratitis in Slovakia. Parasitol. Res. 2010, 106, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Province, M.; Meighani, M.; Eslamirad, Z.; Hajihossein, R.; Ahmadi, A.; Saki, S. Isolation and genotyping of Acanthamoeba from soil samples in Markazi Province, Iran. Open Access Maced. J. Med. Sci. 2018, 6, 2290–2294. [Google Scholar]
- Spanakos, G.; Tzanetou, K.; Miltsakakis, D.; Patsoula, E.; Malamou-Lada, E.; Vakalis, N.C. Genotyping of pathogenic Acanthamoebae isolated from clinical samples in Greece-Report of a clinical isolate presenting T5 genotype. Parasitol. Int. 2006, 55, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.R.S.; Rivera, W.L. Genotype analysis of Acanthamoeba isolated from human nasal swabs in the Philippines. Asian Pac. J. Trop. Med. 2014, 7, S74–S78. [Google Scholar] [CrossRef] [Green Version]
- Alsam, S.; Kim, K.S.; Stins, M.; Rivas, A.O.; Sissons, J.; Khan, N.A. Acanthamoeba interactions with human brain microvascular endothelial cells. Microb. Pathog. 2003, 35, 235–241. [Google Scholar] [CrossRef]
- Grün, A.; Stemplewitz, B.; Scheid, P. First report of an Acanthamoeba genotype T13 isolate as etiological agent of a keratitis in humans. Parasitol. Res. 2014, 113, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Kialashaki, E.; Daryani, A.; Sharif, M.; Gholami, S.; Dodangeh, S.; Moghddam, Y.D.; Sarvi, S.; Montazeri, M. Acanthamoeba spp. from water and soil sources in Iran: A systematic review and meta-analysis. Ann. Parasitol. 2018, 64, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Adamska, M. Molecular characterization of Acanthamoeba spp. occurring in water bodies and patients in Poland and redefinition of polish T16 genotype. J. Eukaryot. Microbiol. 2016, 63, 262–270. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, A.; Lin, J.; Geiseler, P.J.; Qvarnstrom, Y.; Bandea, R.; Roy, S.; Sriram, R.; Paddock, C.; Zaki, S.; Kim, G.; et al. Cutaneous Acanthamoebiasis with cns involvement post-transplantation: Implication for differential diagnosis of skin lesions in immunocompromised patients. J. Neuroparasitol. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Matsui, T.; Maeda, T.; Kusakabe, S.; Arita, H.; Yagita, K.; Morii, E. A case report of granulomatous amoebic encephalitis by group 1 Acanthamoeba genotype T18 diagnosed by the combination of morphological examination and genetic analysis. Diagn. Pathol. 2018, 13, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lares-Jiménez, L.F.; Borquez-Román, M.A.; Lares-García, C.; Otero-Ruiz, A.; Gonzalez-Galaviz, J.R.; Ibarra-Gámez, J.C.; Lares-Villa, F. Potentially pathogenic genera of free-living amoebae coexisting in a thermal spring. Exp. Parasitol. 2018, 195, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; Vanzzini-Zago, V.; Hernandez-Martinez, D.; Gonzalez-Robles, A.; Salazar-Villatoro, L.; Ramirez-Flores, E.; Oregon-Miranda, E.; Lorenzo-Morales, J.; Martinez-Palomo, A. Acanthamoeba genotypes T3 and T4 as causative agents of amoebic keratitis in Mexico. Parasitol. Res. 2016, 115, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Martín-Pérez, T.; Criado-Fornelio, A.; Martínez, J.; Blanco, M.A.; Fuentes, I.; Pérez-Serrano, J. Isolation and molecular characterization of Acanthamoeba from patients with keratitis in Spain. Eur. J. Protistol. 2017, 61, 244–252. [Google Scholar] [CrossRef]
- Tyndall, R.; Ali, I.; Newsome, A. Ramifications of the diverse mRNA patterns in Acanthamoeba royreba. bioRxiv 2020. [Google Scholar] [CrossRef]
- Reyes-Batlle, M.; Zamora-Herrera, J.; Vargas-Mesa, A.; Valerón-Tejera, M.A.; Wagner, C.; Martín-Navarro, C.M.; López-Arencibia, A.; Sifaoui, I.; Martínez-Carretero, E.; Valladares, B.; et al. Acanthamoeba genotypes T2, T4, and T11 in soil sources from El Hierro island, Canary Islands, Spain. Parasitol. Res. 2016, 115, 2953–2956. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Lindo, J.F.; Martinez, E.; Calder, D.; Figueruelo, E.; Valladares, B.; Ortega-Rivas, A. Pathogenic Acanthamoeba strains from water sources in Jamaica, west indies. Ann. Trop. Med. Parasitol. 2005, 99, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Maghsood, A.H.; Sissons, J.; Rezaian, M.; Nolder, D.; Warhurst, D.; Khan, N.A. Acanthamoeba genotype T4 from the UK and Iran and isolation of the T2 genotype from clinical isolates. J. Med. Microbiol. 2005, 54, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, M.d.C.; Rodríguez Fermepín, M.; Gracia Martínez, F.; Costamagna, S.R. Presence of Acanthamoeba spp. in watering trough in La Pampa province, Argentina. Rev. Argent. Microbiol. 2017, 49, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Favuzza, E.; Galano, A.; Di Filippo, M.M.; Ciccone, N.; Berrilli, F.; Mencucci, R.; Di Cave, D.; Rossolini, G.M. Regional spread of contact lens-related Acanthamoeba keratitis in Italy. New Microbiol. 2018, 41, 83–85. [Google Scholar] [PubMed]
- Di Cave, D.; Monno, R.; Bottalico, P.; Guerriero, S.; D’Amelio, S.; D’Orazi, C.; Berrilli, F. Acanthamoeba T4 and T15 genotypes associated with keratitis infections in Italy. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 607–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Serrano, F.D.B.; Caro-Magdaleno, M.; Perea-Pérez, R.; Rodríguez-De-La-Rúa, E.; Montero-Iruzubieta, J. Case Report: Acanthamoeba Keratitis management in a first-trimester pregnant patient. Optom. Vis. Sci. 2018, 95, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Fabres, L.F.; Maschio, V.J.; Dos Santos, D.L.; Kwitko, S.; Marinho, D.R.; De Araújo, B.S.; Locatelli, C.I.; Rott, M.B. Virulent T4 Acanthamoeba causing keratitis in a patient after swimming while wearing contact lenses in southern Brazil. Acta Parasitol. 2018, 63, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Cope, J.R.; Konne, N.M.; Jacobs, D.S.; Dhaliwal, D.K.; Rhee, M.K.; Yin, J.; Steinemann, T.L. Corneal infections associated with sleeping in contact lenses—Six cases, United States, 2016–2018. MMWR Morb. Mortal Wkly. Rep. 2018, 67, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.S.M.M.; Gonçalves, G.S.; Moraes, A.S.; Alves, L.M.; Rodrigues, J.; Hecht, M.M.; Nitz, N.; Gurgel-gonçalves, R.; Bernardes, G.; de Castro, A.M.; et al. Case report the first Acanthamoeba keratitis case in the midwest region of Brazil: Diagnosis, genotyping of the parasite and disease outcome. Rev. Soc. Bras. Med. Trop. 2018, 51, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Geith, S.; Walochnik, J.; Prantl, F.; Sack, S.; Eyer, F. Lethal outcome of granulomatous acanthamoebic encephalitis in a man who was human immunodeficiency virus-positive: A case report. J. Med. Case Rep. 2018, 12, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; Vanzzini-Zago, V.; Hernández-Martínez, D.; Reyes-Batlle, M.; Castelan-Ramírez, I.; Hernández-Olmos, P.; Salazar-Villatoro, L.; González-Robles, A.; Ramírez-Flores, E.; Servín-Flores, C. Acanthamoeba keratitis in Mexico: Report of a clinical case and importance of sensitivity assays for a better outcome. Exp. Parasitol. 2019, 196, 22–27. [Google Scholar] [CrossRef]
- Lau, H.L.; Corvino, D.F.D.L.; Jr, F.M.G.; Amer, M.; Lichtenberger, P.N.; Gultekin, S.H.; Ritter, J.M.; Ali, I.K.M.; Cope, J.R.; Post, M.J.D.; et al. Granulomatous amoebic encephalitis caused by Acanthamoeba in a patient with AIDS: A challenging diagnosis. Acta Clin. Belg. 2019, 76, 127–131. [Google Scholar] [CrossRef]
- Singh, A.; Acharya, M.; Jose, N.; Gandhi, A.; Sharma, S. 18S rDNA sequencing aided diagnosis of Acanthamoeba jacobsi keratitis -a case report. Indian J. Ophthalmol. 2019, 67, 1886–1888. [Google Scholar] [CrossRef] [PubMed]
- Chkheidze, R.; Evers, B.M.; Cavuoti, D.; Merritt, J.; Hogan, R.N. Bilateral Acanthamoeba panophthalmitis: A rare and unique case. Am. J. Ophthalmol. Case Rep. 2020, 20, 100970. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Srikumaran, D.; Zafar, S.; Salehi, M.; Liu, T.S.; Woreta, F.A. Case series: Delayed diagnoses of Acanthamoeba keratitis. Am. J. Ophthalmol. Case Rep. 2020, 19, 100778. [Google Scholar] [CrossRef]
- Korkmaz, I.; Barut Selver, O.; Simsek, C.; Palamar, M. Negative corneal fluorescein staining as an exceptionally early sign of Acanthamoeba Keratitis: A Case Report. Eye Contact Lens 2021, 47, 622–624. [Google Scholar] [CrossRef] [PubMed]
- Wankhade, A.; Patro, P.; Mishra, N.; Das, P. Successful culture of Acanthamoeba remains a key towards diagnosis of unusual clinical presentation of keratitis: A first case report from Chhattisgarh. J. Fam. Med. Prim. Care. 2021, 10, 3904. [Google Scholar] [CrossRef]
- Joshi, L.S.; Gurung, R. Acanthamoeba Keratitis—A case report. Nepal. J. Ophthalmol. 2021, 13, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Damhorst, G.L.; Watts, A.; Hernandez-Romieu, A.; Mel, N.; Palmore, M.; Ali, I.K.M.; Neill, S.G.; Kalapila, A.; Cope, J.R. Acanthamoeba castellanii encephalitis in a patient with AIDS: A case report and literature review. Lancet Infect. Dis. 2022, 22, e59–e65. [Google Scholar] [CrossRef]
- Dos Santos, D.L.; Virginio, V.G.; Berté, F.K.; Lorenzatto, K.R.; Marinho, D.R.; Kwitko, S.; Locatelli, C.I.; Freitas, E.C.; Rott, M.B. Clinical and molecular diagnosis of Acanthamoeba keratitis in contact lens wearers in southern Brazil reveals the presence of an endosymbiont. Parasitol. Res. 2022, 121, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara, G.S.; Shuster, F.L.; Martinez, A.J. Balamuthia mandrillaris, N.G., N. Sp., agent of amebic meningoencephalitis in humans and other animals. J. Eukaryot. Microbiol. 1993, 40, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Yohannan, B.; Feldman, M. Fatal Balamuthia mandrillaris Encephalitis. Case Rep. Infect. Dis. 2019, 2019, 9315756. [Google Scholar] [CrossRef] [Green Version]
- Matin, A.; Siddiqui, R.; Jayasekera, S.; Khan, N.A. Increasing importance of Balamuthia mandrillaris. Clin. Microbiol. Rev. 2008, 21, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Okamoto, K.; Genkai, N.; Kakita, A.; Abe, H. A homogeneously enhancing mass evolving into multiple hemorrhagic and necrotic lesions in amoebic encephalitis with necrotizing vasculitis. Clin. Imaging 2020, 60, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.C.; Fiester, S.E.; Madeline, L.A.; Fulcher, J.W.; Ward, M.E.; Schammel, C.M.G.; Hakimi, R.K. Acanthamoeba spp. and Balamuthia mandrillaris leading to fatal granulomatous amebic encephalitis. Forensic. Sci. Med. Pathol. 2020, 16, 171–176. [Google Scholar] [CrossRef]
- Kum, S.J.; Lee, H.W.; Jung, H.R.; Choe, M.; Kim, S.P. Amoebic encephalitis caused by Balamuthia mandrillaris. J. Pathol. Transl. Med. 2019, 53, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Hallur, V.; Goyal, M.K.; Sehgal, R.; Radotra, B.D. Emergence of Balamuthia mandrillaris meningoencephalitis in India. Indian J. Med. Microbiol. 2015, 33, 298–300. [Google Scholar] [CrossRef]
- Bravo, F.G.; Seas, C. Balamuthia mandrillaris amoebic encephalitis: An emerging parasitic infection. Curr. Infect. Dis. Rep. 2012, 14, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Todd, C.D.; Reyes-Batlle, M.; Piñero, J.E.; Martínez-Carretero, E.; Valladares, B.; Lindo, J.F.; Lorenzo-Morales, J. Balamuthia mandrillaris therapeutic mud bath in Jamaica. Epidemiol. Infect. 2015, 143, 2245–2248. [Google Scholar] [CrossRef] [PubMed]
- Tavares, M.; Da Costa, J.M.C.; Carpenter, S.S.; Santos, L.A.; Afonso, C.; Aguiar, Á.; Pereira, J.; Cardoso, A.I.; Schuster, F.L.; Yagi, S.; et al. Diagnosis of first case of Balamuthia amoebic encephalitis in Portugal by immunofluorescence and PCR. J. Clin. Microbiol. 2006, 44, 2660–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasaelap, A.; Prechawit, S.; Chansaenroj, J.; Punyahotra, P.; Puthanakit, T.; Chomtho, K.; Shuangshoti, S. Fatal Balamuthia amebic encephalitis in a healthy child: A case report with review of survival cases. Korean J. Parasitol. 2013, 51, 335–341. [Google Scholar] [CrossRef]
- Lares-Jiménez, L.F.; Booton, G.C.; Lares-Villa, F.; Velázquez-Contreras, C.A.; Fuerst, P.A. Genetic analysis among environmental strains of Balamuthia mandrillaris recovered from an artificial lagoon and from soil in Sonora, Mexico. Exp. Parasitol. 2014, 145, S57–S61. [Google Scholar] [CrossRef]
- Latifi, A.R.; Niyyati, M.; Lorenzo-Morales, J.; Haghighi, A.; Tabaei, S.J.S.; Lasjerdi, Z. Presence of Balamuthia mandrillaris in hot springs from Mazandaran Province, northern Iran. Epidemiol. Infect. 2016, 144, 2456–2461. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Vílchez, A.M.; Reyes-Batlle, M.; Montalbán-Sandoval, E.; Martín-Navarro, C.M.; López-Arencibia, A.; Elias-Letts, R.; Guerra, H.; Gotuzzo, E.; Martínez-Carretero, E.; Piñero, J.E.; et al. The isolation of Balamuthia mandrillaris from environmental sources from Peru. Parasitol. Res. 2014, 113, 2509–2513. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yan, G.; Han, S.; Ye, Y.; Cheng, X.; Gong, H.; Yu, H. Diagnosing Balamuthia mandrillaris encephalitis via next-generation sequencing in a 13-year-old girl. Emerg. Microbes. Infect. 2020, 9, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Saffioti, C.; Mesini, A.; Caorsi, R.; Severino, M.; Gattorno, M.; Castagnola, E. Balamuthia mandrillaris infection: Report of 1st autochthonous, fatal case in Italy. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 685–687. [Google Scholar] [CrossRef] [PubMed]
- White, J.M.L.; Barker, R.D.; Salisbury, J.R.; Fife, A.J.; Lucas, S.B.; Warhurst, D.C.; Higgins, E.M. Granulomatous amoebic encephalitis. Lancet 2004, 364, 220. [Google Scholar] [CrossRef]
- Laurie, M.; White, C.; Retallack, H.; Wu, W.; Moser, M.; Sakanari, J.; Ang, K.; Wilson, C.; Arkin, M.; DeRisi, J. Repurposing the quinoline antibiotic nitroxoline to treat infections caused by the brain-eating amoeba Balamuthia mandrillaris. bioRxiv 2018, 331785. [Google Scholar] [CrossRef] [Green Version]
- Cope, J.R.; Landa, J.; Nethercut, H.; Collier, S.A.; Glaser, C.; Moser, M.; Puttagunta, R.; Yoder, J.S.; Ali, I.K.; Roy, S.L. The epidemiology and clinical features of Balamuthia mandrillaris disease in the United States, 1974-2016. Clin. Infect. Dis. 2019, 68, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Vílchez, A.M.; Chura-Araujo, M.A.; Anicama Lima, W.E.; Vela, C.; Asencio, A.Y.; García, H.; del Carmen Garaycochea, M.; Náquira, C.; Rojas, E.; Martínez, D.Y. Fatal granulomatous amoebic encephalitis due to free-living amoebae in two boys in two different hospitals in Lima, Perú. Neuropathology 2020, 40, 180–184. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, X.; Min, L.; Dong, X.; Guan, Y. Balamuthia mandrillaris-related primary amoebic encephalitis in china diagnosed by Next Generation Sequencing and a review of the literature. Lab. Med. 2020, 51, 20–26. [Google Scholar] [CrossRef]
- Piper, K.J.; Foster, H.; Susanto, D.; Maree, C.L.; Thornton, S.D.; Cobbs, C.S. Fatal Balamuthia mandrillaris brain infection associated with improper nasal lavage. Int. J. Infect. Dis. 2018, 77, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Booton, G.C.; Carmichael, J.R.; Visvesvara, G.S.; Byers, T.J.; Fuerst, P.A. Genotyping of Balamuthia mandrillaris based on nuclear 18S and mitochondrial 16s rRNA genes. Am. J. Trop. Med. Hyg. 2003, 68, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Greninger, A.L.; Messacar, K.; Dunnebacke, T.; Naccache, S.N.; Federman, S.; Bouquet, J.; Mirsky, D.; Nomura, Y.; Yagi, S.; Glaser, C.; et al. Clinical metagenomic identification of Balamuthia mandrillaris encephalitis and assembly of the draft genome: The continuing case for reference genome sequencing. Genome Med. 2015, 7, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehab, K.W.; Aboul-nasr, K.; Elliott, S.P. Balamuthia mandrillaris granulomatous amebic encephalitis with renal dissemination in a previously healthy child: Case report and review of the pediatric literature. J. Pediatr. Infect. Dis. Soc. 2018, 7, 163–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Toyoshima, M.; Nakamura, M.; Sato, M.; Shimizu, H.; Inoue, C.; Shimizu, Y.; Yagita, K. An acute case of granulomatous amoebic encephalitis-Balamuthia mandrillaris infection. Intern. Med. 2018, 57, 1313–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyatanda, G.; Rand, K.; Lindner, M.S.; Hong, D.K.; Sait Albayram, M.; Gregory, J.; Kresak, J.; Ibne, K.M.A.; Cope, J.R.; Roy, S.; et al. Rapid, Noninvasive diagnosis of Balamuthia mandrillaris encephalitis by a plasma-based next-generation sequencing test. Open Forum Infect. Dis. 2020, 7, ofaa189. [Google Scholar] [CrossRef]
- Hirakata, S.; Sakiyama, Y.; Yoshimura, A.; Ikeda, M.; Takahata, K.; Tashiro, Y.; Yoshimura, M.; Arata, H.; Yonezawa, H.; Kirishima, M.; et al. The application of shotgun metagenomics to the diagnosis of granulomatous amoebic encephalitis due to Balamuthia mandrillaris: A case report. BMC Neurol. 2021, 21, 392. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yu, I.K.; Kim, S.M.; Kim, J.H.; Kim, H.Y. Fulminant disseminating fatal granulomatous amebic encephalitis: The first case report in an immunocompetent patient in south Korea. Yonsei Med. J. 2021, 62, 563–567. [Google Scholar] [CrossRef]
- Peng, L.; Zhou, Q.; Wu, Y.; Cao, X.; Lv, Z.; Su, M.; Yu, Y.; Huang, W. A patient with granulomatous amoebic encephalitis caused by Balamuthia mandrillaris survived with two excisions and medication. BMC Infect Dis 2022, 22, 54. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Y.; Yu, Y.; Yu, H.; Guo, S.; Shi, D.; He, J.; Hu, C.; Yang, J.; Fang, X.; et al. Encephalomyelitis caused by Balamuthia mandrillaris in a woman with breast cancer: A case report and review of the literature. Front. Immunol. 2022, 12, 768065. [Google Scholar] [CrossRef]
- Ai, J.; Zhang, H.; Yu, S.; Li, J.; Chen, S.; Zhang, W.; Mao, R. A case of fatal amoebic encephalitis caused by Balamuthia mandrillaris, China. Infect Genet. Evol. 2022, 97, 105190. [Google Scholar] [CrossRef]
- Cuoco, J.A.; Klein, B.J.; LeBel, D.P.; Faulhaber, J.; Apfel, L.S.; Witcher, M.R. Successful treatment of a Balamuthia mandrillaris cerebral abscess in a pediatric patient with complete surgical resection and antimicrobial therapy. Pediatr. Infect. Dis. J. 2022, 41, E54–E57. [Google Scholar] [CrossRef]
- Walochnik, J.; Wylezich, C.; Michel, R. The genus Sappinia: History, phylogeny and medical relevance. Exp. Parasitol. 2010, 126, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Wylezich, C.; Walochnik, J.; Corsaro, D.; Michel, R.; Kudryavtsev, A. Electron microscopical investigations of a new species of the genus Sappinia (Thecamoebidae, Amoebozoa), Sappinia Platani sp. nov., reveal a dictyosome in this genus. Acta Protozool. 2015, 54, 45–51. [Google Scholar] [CrossRef]
- Corsaro, D.; Wylezich, C.; Walochnik, J.; Venditti, D.; Michel, R. Molecular identification of bacterial endosymbionts of Sappinia strains. Parasitol. Res. 2017, 116, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Gelman, B.B.; Rauf, S.J.; Nader, R.; Popov, V.; Borkowski, J.; Chaljub, G.; Nauta, H.W.; Visvesvara, G.S. Amoebic encephalitis due to Sappinia diploidea. J. Am. Med. Assoc. 2001, 285, 2450–2451. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; da Silva, A.J.; Schuster, F.L.; Gelman, B.B.; Visvesvara, G.S. Molecular confirmation of Sappinia pedata as a causative agent of amoebic encephalitis. J. Infect. Dis. 2009, 199, 1139–1142. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.W.; Spiegel, F.W.; Silberman, J.D. Amoeba at attention: Phylogenetic affinity of Sappinia Pedata. J. Eukaryot. Microbiol. 2007, 54, 511–519. [Google Scholar] [CrossRef]
- Siddiqui, R.; Makhlouf, Z.; Khan, N.A. The increasing importance of Vermamoeba Vermiformis. J. Eukaryot. Microbiol. 2021, 68, e12857. [Google Scholar] [CrossRef]
- Chelkha, N.; Hasni, I.; Louazani, A.C.; Levasseur, A.; la Scola, B.; Colson, P. Vermamoeba vermiformis CDC-19 draft genome sequence reveals considerable gene trafficking including with candidate phyla radiation and giant viruses. Sci. Rep. 2020, 10, 19–21. [Google Scholar] [CrossRef] [Green Version]
- Haddad, M.H.F.; Habibpour, H.; Mahmoudi, M.R. Isolation and molecular identification of free-living amoebae (Naegleria spp., Acanthamoeba spp. and Vermamoeba spp.) from mineral springs in Guilan Province, northern Iran. J. Water Health 2020, 18, 60–66. [Google Scholar] [CrossRef]
- Baker, R.E.; Mahmud, A.S.; Miller, I.F.; Rajeev, M.; Rasambainarivo, F.; Rice, B.L.; Takahashi, S.; Tatem, A.J.; Wagner, C.E.; Wang, L.F.; et al. Infectious disease in an era of global change. Nat. Rev. Microbiol. 2022, 20, 193–205. [Google Scholar] [CrossRef]
- Siddiqui, R.; Boghossian, A.; Khatoon, B.; Kawish, M.; Alharbi, A.M.; Shah, M.R.; Alfahemi, H.; Khan, N.A. Antiamoebic properties of metabolites against Naegleria fowleri and Balamuthia mandrillaris. Antibiotics 2022, 11, 539. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P. Free-living amoebae as human parasites and hosts for pathogenic microorganisms. Proc. West Mark. Ed. Assoc. Conf. 2018, 2, 692. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Cheng, X. Various brain-eating amoebae: The protozoa, the pathogenesis, and the disease. Front. Med. 2021, 15, 842–866. [Google Scholar] [CrossRef] [PubMed]
- Angelici, M.C.; Walochnik, J.; Calderaro, A.; Saxinger, L.; Dacks, J.B. Free-living amoebae and other neglected protistan pathogens: Health emergency signals? Eur. J. Protistol. 2021, 77, 125760. [Google Scholar] [CrossRef] [PubMed]
- Hill, V.; Ruis, C.; Bajaj, S.; Pybus, O.G.; Kraemer, M.U.G. Progress and challenges in virus genomic epidemiology. Trends Parasitol. 2021, 37, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Mashe, T.; Takawira, F.T.; de Oliveira Martins, L.; Gudza-Mugabe, M.; Chirenda, J.; Munyanyi, M.; Chaibva, B.V.; Tarupiwa, A.; Gumbo, H.; Juru, A.; et al. Genomic epidemiology and the role of international and regional travel in the SARS-CoV-2 epidemic in Zimbabwe: A retrospective study of routinely collected surveillance data. Lancet Glob. Health 2021, 9, e1658–e1666. [Google Scholar] [CrossRef]
- Alteri, C.; Cento, V.; Piralla, A.; Costabile, V.; Tallarita, M.; Colagrossi, L.; Renica, S.; Giardina, F.; Novazzi, F.; Gaiarsa, S.; et al. Genomic epidemiology of SARS-CoV-2 reveals multiple lineages and early spread of SARS-CoV-2 infections in Lombardy, Italy. Nat. Commun. 2021, 12, 434. [Google Scholar] [CrossRef]
- Islam, A.; Sayeed, M.A.; Kalam, M.A.; Fedous, J.; Shano, S.; Abedin, J.; Islam, S.; Choudhury, S.D.; Saha, O.; Hassan, M.M. Transmission pathways and genomic epidemiology of emerging variants of SARS-CoV-2 in the environment. COVID 2022, 2, 916–939. [Google Scholar] [CrossRef]
- Mclaughlin, A.; Montoya, V.; Miller, R.L.; Mordecai, G.J.; Worobey, M.; Poon, A.F.; Joy, J.B. Genomic epidemiology of the first two waves of SARS-CoV-2 in Canada. eLife 2022, 11, 73896. [Google Scholar] [CrossRef]
- Fibke, C.D.; Croxen, M.A.; Geum, H.M.; Glass, M.; Wong, E.; Avery, B.P.; Daignault, D.; Mulvey, M.R.; Reid-Smith, R.J.; Parmley, E.J.; et al. Genomic epidemiology of major extraintestinal pathogenic Escherichia Coli lineages causing urinary tract infections in young women across Canada. Open Forum Infect. Dis. 2019, 6, ofz431. [Google Scholar] [CrossRef]
- Mäklin, T.; Kallonen, T.; Alanko, J.; Samuelsen, Ø.; Hegstad, K.; Mäkinen, V.; Corander, J.; Heinz, E.; Honkela, A. Bacterial genomic epidemiology with mixed samples. Microb. Genom. 2021, 7, 000691. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Maex, M.; Mattheus, W.; van den Bossche, A.; van Cauteren, D.; Laisnez, V.; Hammami, N.; Ceyssens, P.J. Genomic epidemiology of persistently circulating MDR Shigella Sonnei strains associated with men who have sex with men (MSM) in Belgium (2013-19). J. Antimicrob. Chemother. 2022, 77, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiang, X.; Yang, T.; Ju, Y.; Yin, Z.; Yue, L.; Ma, G.; Wang, X.; Jing, Y.; Luo, X.; et al. Genomic epidemiology of carbapenemase-producing Klebsiella Pneumoniae in China. Genom. Proteom. Bioinform. 2022. [Google Scholar] [CrossRef]
- Guglielmini, J.; Hennart, M.; Badell, E.; Toubiana, J.; Criscuolo, A.; Brisse, S. Genomic epidemiology and strain taxonomy of Corynebacterium diphtheriae. J. Clin. Microbiol. 2021, 59, e01581-21. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhou, W.; Zhan, Q.; Chen, Y.; Luo, Q.; Shen, P.; Xiao, Y. Genomic epidemiology and characterisation of penicillin-sensitive Staphylococcus Aureus isolates from invasive bloodstream infections in China: An increasing prevalence and higher diversity in genetic typing be revealed. Emerg. Microbes Infect. 2022, 11, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Verza, M.; Scheffer, M.C.; Salvato, R.S.; Schorner, M.A.; Barazzetti, F.H.; Machado, H.d.M.; Medeiros, T.F.; Rovaris, D.B.; Portugal, I.; Viveiros, M.; et al. Genomic epidemiology of Mycobacterium Tuberculosis in Santa Catarina, southern Brazil. Sci. Rep. 2020, 10, 12891. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.; Abdolrasouli, A.; Farrer, R.A.; Cuomo, C.A.; Aanensen, D.M.; Armstrong-James, D.; Fisher, M.C.; Schelenz, S. Genomic epidemiology of the UK outbreak of the emerging human fungal pathogen Candida auris. Emerg. Microbes Infect. 2018, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Brackin, A.P.; Hemmings, S.J.; Fisher, M.C.; Rhodes, J. Fungal genomics in respiratory medicine: What, how and when? Mycopathologia 2021, 186, 589–608. [Google Scholar] [CrossRef] [PubMed]
- Barker, B.M.; Cuomo, C.A.; Govender, N.P. Genomic characterization of emerging human fungal pathogens. Front. Genet. 2021, 12, 674765. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.; Abdolrasouli, A.; Dunne, K.; Sewell, T.R.; Zhang, Y.; Ballard, E.; Brackin, A.P.; van Rhijn, N.; Chown, H.; Tsitsopoulou, A.; et al. Population genomics confirms acquisition of drug-resistant Aspergillus Fumigatus infection by humans from the environment. Nat. Microbiol. 2022, 7, 663–674. [Google Scholar] [CrossRef]
- Voorhies, M.; Cohen, S.; Shea, T.P.; Petrus, S.; Muñoz, J.F.; Poplawski, S.; Goldman, W.E.; Michael, T.P.; Cuomo, C.A.; Sil, A.; et al. Chromosome-level genome assembly of a human fungal pathogen reveals synteny among geographically distinct species. Mbio 2022, 13, e02574-21. [Google Scholar] [CrossRef] [PubMed]
- Liechti, N.; Schürch, N.; Bruggmann, R.; Wittwer, M. The genome of Naegleria lovaniensis, the basis for a comparative approach to unravel pathogenicity factors of the human pathogenic amoeba N. fowleri. BMC Genom. 2018, 19, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, E.K.; Greninger, A.; van der Giezen, M.; Ginger, M.L.; Ramirez-Macias, I.; Miller, H.C.; Morgan, M.J.; Tsaousis, A.D.; Velle, K.; Vargová, R.; et al. Genomics and transcriptomics yields a system-level view of the biology of the pathogen Naegleria fowleri. BMC Biol. 2021, 19, 142. [Google Scholar] [CrossRef]
- Gu, X.; Lu, X.; Lin, S.; Shi, X.; Shen, Y.; Lu, Q.; Yang, Y.; Yang, J.; Cai, J.; Fu, C.; et al. A comparative genomic approach to determine the virulence factors and horizontal gene transfer events of clinical Acanthamoeba isolates. Microbiol. Spectr. 2022, 10, e00025-22. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wu, X.; Tan, M.; Wang, D.; Wang, S.; Wu, Y. Subacute Balamuthia mandrillaris encephalitis in an immunocompetent patient diagnosed by next-generation sequencing. J. Int. Med. Res. 2022, 50, 03000605221093217. [Google Scholar] [CrossRef] [PubMed]
- Phan, I.Q.; Rice, C.A.; Craig, J.; Noorai, R.E.; McDonald, J.R.; Subramanian, S.; Tillery, L.; Barrett, L.K.; Shankar, V.; Morris, J.C.; et al. The transcriptome of Balamuthia mandrillaris trophozoites for structure-guided drug design. Sci. Rep. 2021, 11, 21664. [Google Scholar] [CrossRef]
- Pedra-Rezende, Y.; Macedo, I.S.; Midlej, V.; Mariante, R.M.; Menna-Barreto, R.F.S. Different drugs, same end: Ultrastructural hallmarks of autophagy in pathogenic protozoa. Front. Microbiol. 2022, 13, 856686. [Google Scholar] [CrossRef]




| Location | Age (Years) | Symptoms | Genotypes | Time of Diagnosis | Reference |
|---|---|---|---|---|---|
| China | 42 | Headache and dyspnea | N/A | 2 days | [35] |
| China | 43 | Headache, fever, fatigue, pain, and chills | 2 | N/A | [25] |
| Bangladesh | 15 | Fever, severe headache, vomiting, weakness, and stiff neck | 4 | 3 years * | [31] |
| Zhejiang | 43 | Headache, fever, myalgia, and fatigue | N/A | 4 days | [36] |
| USA | 8 | Headache, stiff neck, photophobia, and vomiting | N/A | 2 days | [37] |
| Costa Rica | 15 | General discomfort, intense headaches, nausea, and vomiting | N/A | 7 days | [38] |
| Costa Rica | 5 | Headaches, vomiting, fever, lower limb pain, spasticity, hyperreflexia, and walking difficulty | N/A | N/A | [38] |
| Costa Rica | 1 | Fever, drowsiness, and an altered state of consciousness | N/A | 4 days | [38] |
| Pakistan | 26 | High fever, vomiting, and altered consciousness | N/A | 4 days | [39] |
| China | 8 | Headache, fever, vomiting, seizures, and coma | N/A | 24 days * | [40] |
| USA | 13 | Headache, vomiting, and fever | 1 | N/A | [34] |
| Turkey | 18 | Fever, nausea, vomiting, and changes in consciousness | N/A | 7–10 days * | [41] |
| Genotype | Strain | GenBank Accession Number |
|---|---|---|
| T1 | A. castellanii V006 | U07400 |
| T2 | A. palestinensis Reich | U07411 |
| T2/6A | A. polyphaga CCAP 1501/3b | AY026244 |
| T2/6B | Isolate OB3b_3A | AB425945 |
| T2/6C | A. palestinensis OX-1 | AF019051 |
| T3 | A. griffini S-7 | U07412 |
| T4A | A. castellanii | U07413 |
| T4B | A. castellanii Ma | U07414 |
| T4C | A. sp. Fernandez | U07409 |
| T4D | A. rhysodes Singh | AY351644 |
| T4E | A. polyphaga page-23 | AF019061 |
| T4F | A. triangularis SH621 | AF346662 |
| T4 Neff | A. castellanii Neff | U07416 |
| T5 | A. lenticulata Jc-1 | U94739 |
| T6 | A. palestinensis 2802 | AF019063 |
| T7 | A. astronyxis | AF019064 |
| T8 | A. tubiashi OC-15C | AF019065 |
| T9 | A. comandoni | AF019066 |
| T10 | A. culbertsoni Lilly A-1 | AF019067 |
| T11 | A. hatchetti BH-2 | AF019068 |
| T12 | A. healyi V013 OC-3A/AC-020 | AF019070 |
| T13 | UWC9 | AF132134 |
| T14 | PN13 | AF333609 |
| T15 | A. jacobsi 31-B | AY262360 |
| T16 | U/HC1 | AY026245 |
| T17 | Ac E1a | GU808277 |
| T18 | CDC:V621 | KC822461 |
| T19 | USP-AWW-A68 | KJ413084 |
| T20 | OSU 04-020 | DQ451161 |
| T21 | A. royreba | CDEZ01000000 |
| T22 | A. pyriformis | KX840327 |
| T23 | A. bangkokensis | MZ272148 and MZ272149 |
| Sampling Date | Age (Years) | Symptoms | Genotype | Time of Diagnosis | Reference |
|---|---|---|---|---|---|
| 2018 | 29 | Sensitivity to light, redness, and pain in the right eye | N/A | 1 day | [86] |
| 2018 | 27 | Severe eye pain, blurred vision, photophobia, and a foreign body sensation in the left eye | T4 | 1 day | [87] |
| 2018 | 34 | Redness of the left eye and blurred vision | N/A | 6 months | [88] |
| 2018 | 40 | Corneal ulcer and infected left eye | T3 | 6 months | [84] |
| 2018 | 69 | Central corneal ulcer deepening with hypopyon | T4 | >8 months | [84] |
| 2018 | 41 | Central corneal abscess with hypopyon | T4 | 5 months | [84] |
| 2018 | 22 | Corneal ulcer in the right eye | T4 | 7 months | [84] |
| 2018 | 28 | Red eye and foreign body sensation for a week | T4 | 1 week | [89] |
| 2018 | 54 | Progressive neurological deficits, including sensorimotor paralysis of the right leg and impaired alertness | T4 | 12 days | [90] |
| 2019 | 38 | Photophobia and inflammation | T4 | 8 months | [91] |
| 2019 | 24 | Multifocal stromal infiltrates | T4 | 3 weeks | [62] |
| 2019 | 28 | Multifocal stromal infiltrates | T8 | 2 weeks | [62] |
| 2019 | 53 | Altered mental status and fever | N/A | 2.5 months | [92] |
| 2019 | 9 | Decreased vision, pain, photophobia, and redness | T15 | ~5 weeks | [93] |
| 2020 | 65 | Bilateral Acanthamoeba panophthalmitis (first case) | N/A | N/A | [94] |
| 2020 | 54 | Blurred vision, photophobia, and excessive lacrimation | N/A | 2 weeks | [95] |
| 2020 | 18 | Eye pain and blurred vision associated with redness, swelling, and eye discharge | N/A | 7 weeks | [95] |
| 2021 | 30 | Irritation, itching, pain, and redness in the right eye | N/A | 5 days | [96] |
| 2021 | 59 | Redness, tearing, pain, and white discoloration of the cornea in the right eye | N/A | ~2.5 months | [97] |
| 2021 | 45 | Redness, pain, and tearing in the right eye | N/A | N/A | [98] |
| 2022 | 27 | Fever, chills, and lethargy | T1 | N/A | [99] |
| 2022 | 24 | Decreased vision, severe pain, eyelid swelling, redness, and irritation in the right eye | T4 | N/A | [100] |
| 2022 | 23 | Pain, hypopyon, and irritation in the left eye | T4 | N/A | [100] |
| 2022 | 42 | Vision loss, photophobia, and irritation in both eyes | T4 | N/A | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otero-Ruiz, A.; Gonzalez-Zuñiga, L.D.; Rodriguez-Anaya, L.Z.; Lares-Jiménez, L.F.; Gonzalez-Galaviz, J.R.; Lares-Villa, F. Distribution and Current State of Molecular Genetic Characterization in Pathogenic Free-Living Amoebae. Pathogens 2022, 11, 1199. https://doi.org/10.3390/pathogens11101199
Otero-Ruiz A, Gonzalez-Zuñiga LD, Rodriguez-Anaya LZ, Lares-Jiménez LF, Gonzalez-Galaviz JR, Lares-Villa F. Distribution and Current State of Molecular Genetic Characterization in Pathogenic Free-Living Amoebae. Pathogens. 2022; 11(10):1199. https://doi.org/10.3390/pathogens11101199
Chicago/Turabian StyleOtero-Ruiz, Alejandro, Leobardo Daniel Gonzalez-Zuñiga, Libia Zulema Rodriguez-Anaya, Luis Fernando Lares-Jiménez, Jose Reyes Gonzalez-Galaviz, and Fernando Lares-Villa. 2022. "Distribution and Current State of Molecular Genetic Characterization in Pathogenic Free-Living Amoebae" Pathogens 11, no. 10: 1199. https://doi.org/10.3390/pathogens11101199