
Prevalence of Enterococcus spp. and the Whole-Genome Characteristics of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Free-Living Birds in Poland
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
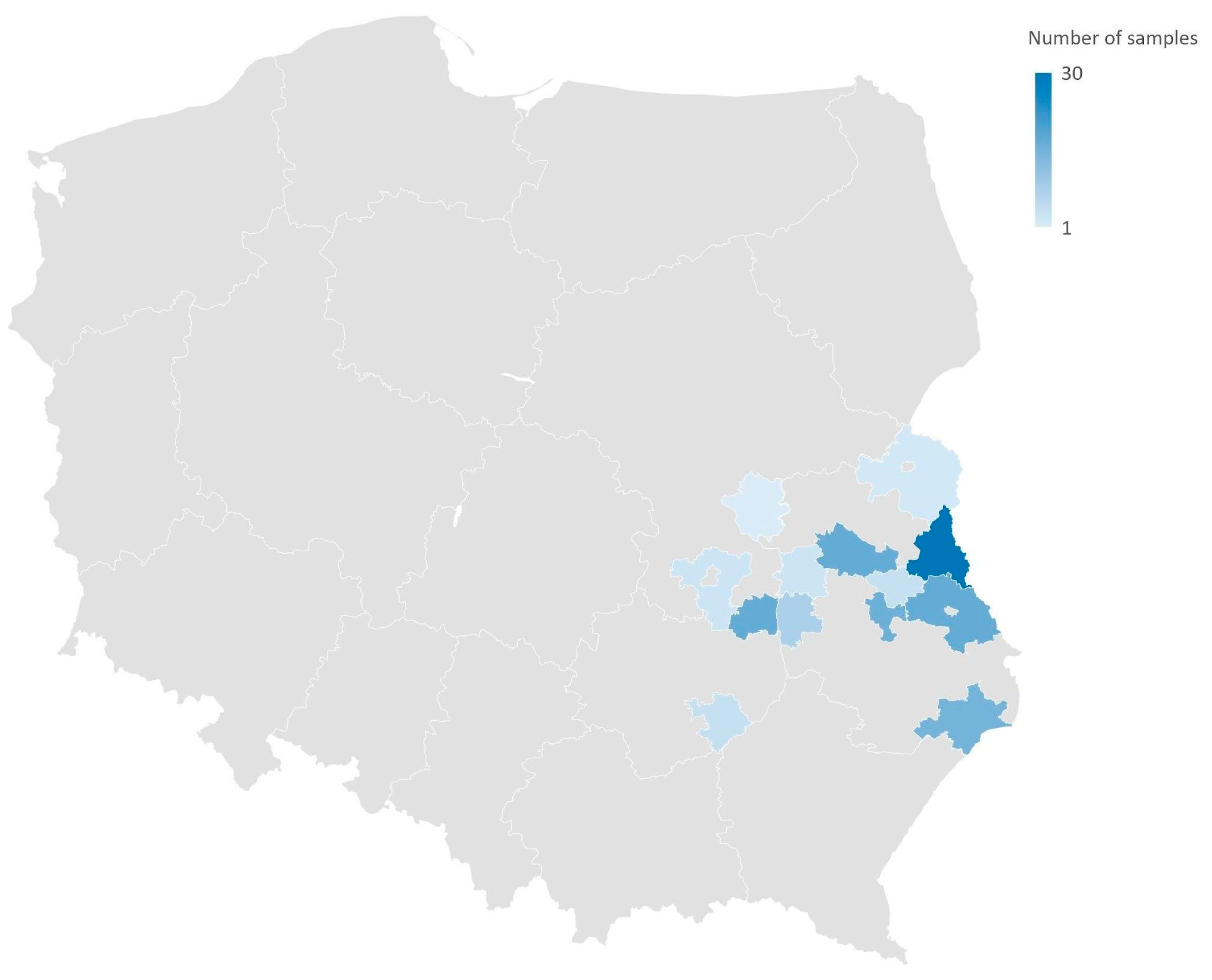
2.1. Sample Collection
2.2. Isolation and Identification
2.3. Antimicrobial Susceptibility Tests
2.4. Whole-Genome Sequencing
2.5. Bioinformatic Data Analysis
3. Results
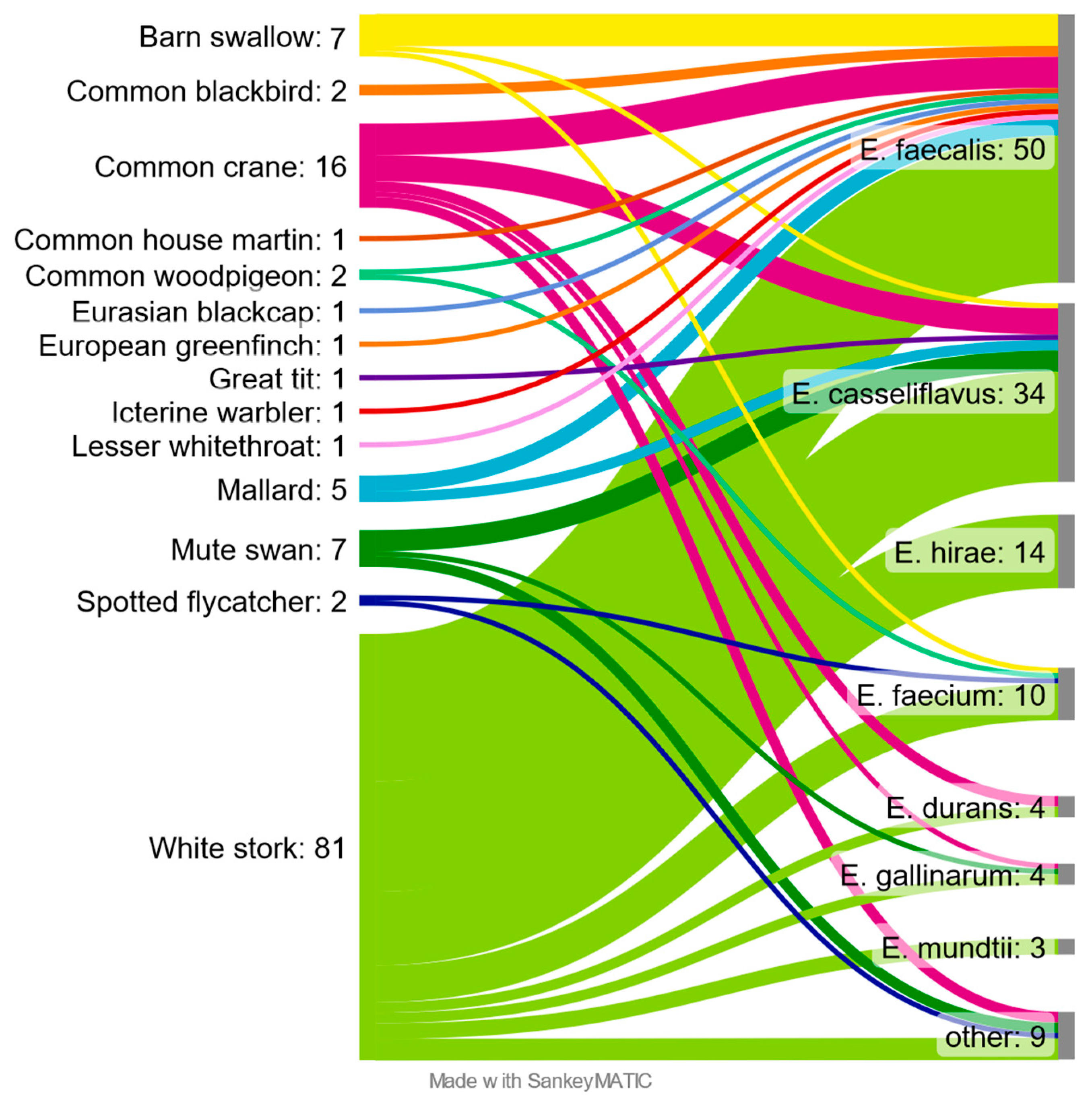
3.1. Enterococcus Prevalence
3.2. Antimicrobial Resistance Results
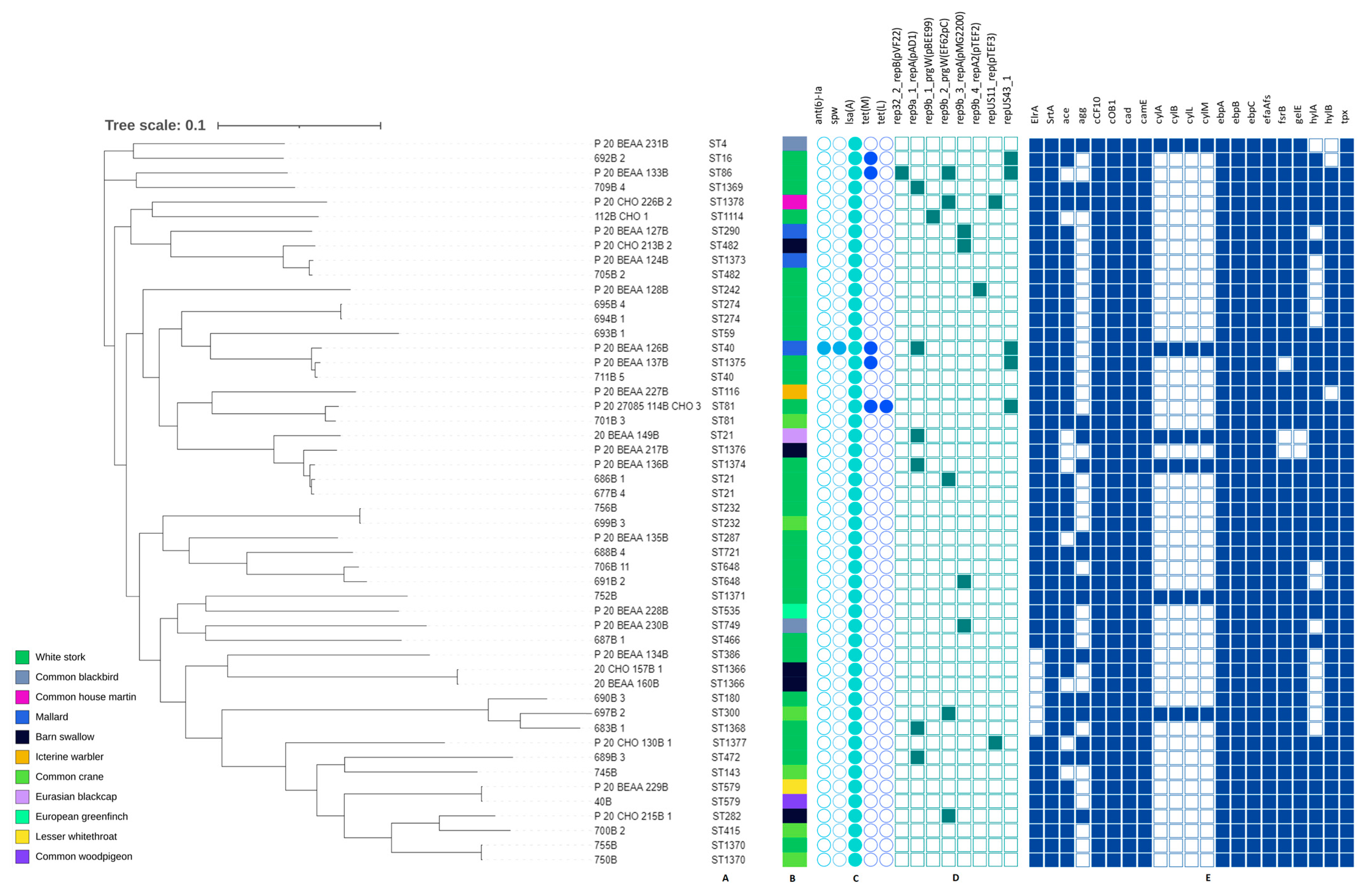
3.3. Genetic Characteristics of E. faecium
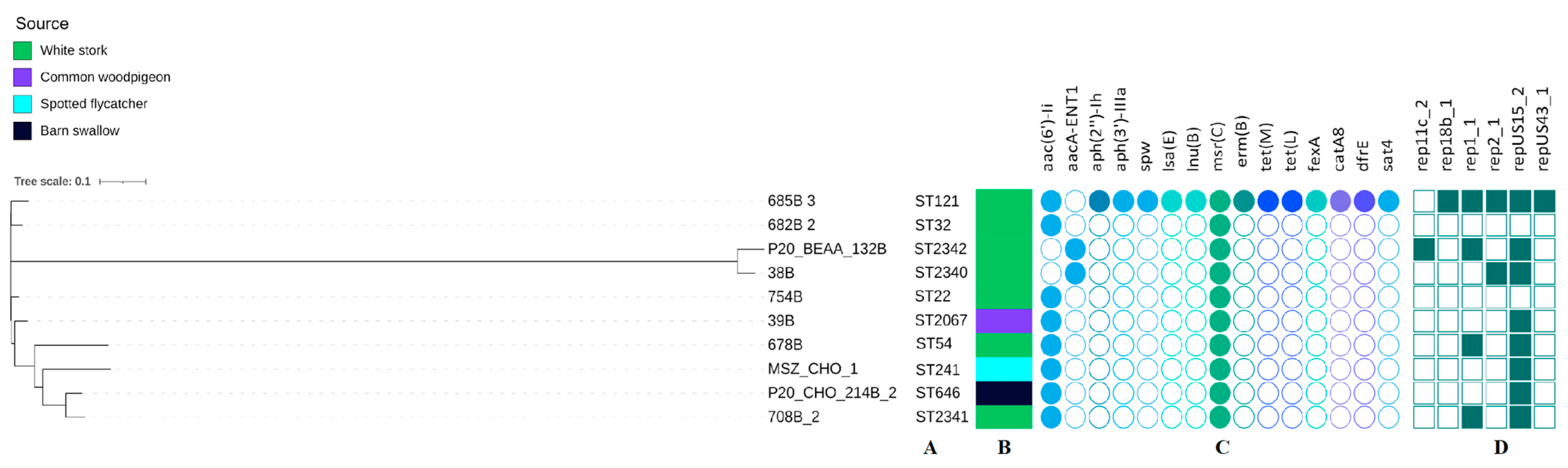
3.4. Genetic Characteristic of E. faecalis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ben Yahia, H.; Chairat, S.; Hamdi, N.; Gharsa, H.; Ben Sallem, R.; Ceballos, S.; Torres, C.; Ben Slama, K. Antimicrobial resistance and genetic lineages of faecal enterococci of wild birds: Emergence of vanA and vanB2 harbouring Enterococcus faecalis. Int. J. Antimicrob. Agents 2018, 52, 936–941. [Google Scholar] [CrossRef]
- Dec, M.; Stȩpień-Pyśniak, D.; Gnat, S.; Fratini, F.; Urban-Chmiel, R.; Cerri, D.; Winiarczyk, S.; Turchi, B. Antibiotic Susceptibility and Virulence Genes in Enterococcus Isolates from Wild Mammals Living in Tuscany, Italy. Microb. Drug Resist. 2020, 26, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Domig, K.J.; Mayer, H.K.; Kneifel, W. Methods used for the isolation, enumeration, characterisation and identification of Enterococcus spp.—2. Pheno- and genotypic criteria. Int. J. Food Microbiol. 2003, 88, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Różańska, H.; Lewtak-Piłat, A.; Osek, J. Enterokoki—Bakterie o wielu obliczach. Życie Weter. 2013, 88, 562–564. [Google Scholar]
- Marrow, J.; Whittington, J.K.; Mitchell, M.; Hoyer, L.L.; Maddox, C. Prevalence and antibiotic-resistance characteristics of Enterococcus spp. isolated from free-living and captive raptors in central illinois. J. Wildl. Dis. 2009, 45, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Solaiman, S.; Patterson, R.; Davey, K.; Katz, Y.; Payne-Sturges, D.; Sapkota, A.R.; Micallef, S.A. Effects of season and water type on the distribution and antimicrobial resistance of Enterococcus faecalis and Ent. faecium from surface and reclaimed water. J. Appl. Microbiol. 2022, 133, 477–487. [Google Scholar] [CrossRef]
- Micallef, S.A.; Rosenberg Goldstein, R.E.; George, A.; Ewing, L.; Tall, B.D.; Boyer, M.S.; Joseph, S.W.; Sapkota, A.R. Diversity, distribution and antibiotic resistance of Enterococcus spp. recovered from tomatoes, leaves, water and soil on U.S. Mid-Atlantic farms. Food Microbiol. 2013, 36, 465–474. [Google Scholar] [CrossRef]
- Abriouel, H.; Omar, N.B.; Molinos, A.C.; López, R.L.; Grande, M.J.; Martínez-Viedma, P.; Ortega, E.; Cañamero, M.M.; Galvez, A. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol. 2008, 123, 38–49. [Google Scholar] [CrossRef]
- Fisher, K.; Phillips, C. The ecology, epidemiology and virulence of Enterococcus. Microbiology 2009, 155, 1749–1757. [Google Scholar] [CrossRef] [Green Version]
- Cagnoli, G.; Bertelloni, F.; Interrante, P.; Ceccherelli, R.; Marzoni, M.; Ebani, V.V. Antimicrobial-Resistant Enterococcus spp. in Wild Avifauna from Central Italy. Antibiotics 2022, 11, 852. [Google Scholar] [CrossRef]
- Lester, C.H.; Frimodt-Møller, N.; Sørensen, T.L.; Monnet, D.L.; Hammerum, A.M. In Vivo Transfer of the vanA Resistance Gene from an Enterococcus faecium Isolate of Animal Origin to an E. faecium Isolate of Human Origin in the Intestines of Human Volunteers. Antimicrob. Agents Chemother. 2006, 50, 596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clin. Microbiol. Infect. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Radhouani, H.; Poeta, P.; Gonçalves, A.; Pacheco, R.; Sargo, R.; Igrejas, G. Wild birds as biological indicators of environmental pollution: Antimicrobial resistance patterns of Escherichia coli and Enterococci isolated from common buzzards (Buteo buteo). J. Med. Microbiol. 2012, 61, 837–843. [Google Scholar] [CrossRef] [Green Version]
- Moubareck, C.; Bourgeois, N.; Courvalin, P.; Doucet-Populaire, F. Multiple Antibiotic Resistance Gene Transfer from Animal to Human Enterococci in the Digestive Tract of Gnotobiotic Mice. Antimicrob. Agents Chemother. 2003, 47, 2993. [Google Scholar] [CrossRef] [Green Version]
- Doucet-Populaire, F.; Trieu-Cuot, P.; Dosbaa, I.; Andremont, A.; Courvalin, P. Inducible Transfer of Conjugative Transposon Tn1545 from Enterococcus faecalis to Listeria monocytogenes in the Digestive Tracts of Gnotobiotic Mice. Antimicrob. Agents Chemother. 1991, 35, 185–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, T.; Silva, N.; Igrejas, G.; Rodrigues, P.; Micael, J.; Rodrigues, T.; Resendes, R.; Gonçalves, A.; Marinho, C.; Gonçalves, D.; et al. Dissemination of antibiotic resistant Enterococcus spp. and Escherichia coli from wild birds of Azores Archipelago. Anaerobe 2013, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Stępień-Pyśniak, D.; Hauschild, T.; Nowaczek, A.; Marek, A.; Dec, M. Wild birds as a potential source of known and novel multilocus sequence types of antibiotic-resistant Enterococcus faecalis. J. Wildl. Dis. 2018, 54, 219–228. [Google Scholar] [CrossRef]
- Klibi, N.; Ben Amor, I.; Rahmouni, M.; Dziri, R.; Douja, G.; Ben Said, L.; Lozano, C.; Boudabous, A.; Ben Slama, K.; Mansouri, R.; et al. Diversity of species and antibiotic resistance among fecal enterococci from wild birds in Tunisia. Detection of vanA-containing Enterococcus faecium isolates. Eur. J. Wildl. Res. 2015, 61, 319–323. [Google Scholar] [CrossRef]
- Stȩpień-Pyśniak, D.; Hauschild, T.; Dec, M.; Marek, A.; Urban-Chmiel, R. Clonal Structure and Antibiotic Resistance of Enterococcus spp. from Wild Birds in Poland. Microb. Drug Resist. 2019, 25, 1227–1237. [Google Scholar] [CrossRef]
- Oravcova, V.; Zurek, L.; Townsend, A.; Clark, A.B.; Ellis, J.C.; Cizek, A.; Literak, I. American crows as carriers of vancomycin-resistant enterococci with vanA gene. Environ. Microbiol. 2014, 16, 939–949. [Google Scholar] [CrossRef]
- Timmermans, M.; Bogaerts, B.; Vanneste, K.; De Keersmaecker, S.C.J.; Roosens, N.H.C.; Kowalewicz, C.; Simon, G.; Argudín, M.A.; Deplano, A.; Hallin, M.; et al. Large diversity of linezolid-resistant isolates discovered in food-producing animals through linezolid selective monitoring in Belgium in 2019. J. Antimicrob. Chemother. 2021, 77, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Higgs, C.; Turner, A.M.; Nong, Y.; Gorrie, C.L.; Sherry, N.L.; Dyet, K.H.; Seemann, T.; Williamson, D.A.; Stinear, T.P.; et al. Daptomycin Resistance Occurs Predominantly in vanA-Type Vancomycin-Resistant Enterococcus faecium in Australasia and Is Associated With Heterogeneous and Novel Mutations. Front. Microbiol. 2021, 12, 749935. [Google Scholar] [CrossRef] [PubMed]
- Rathnayake, I.; Hargreaves, M.; Huygens, F. SNP diversity of Enterococcus faecalis and Enterococcus faecium in a South East Queensland waterway, Australia, and associated antibiotic resistance gene profiles. BMC Microbiol. 2011, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leinweber, H.; Alotaibi, S.M.I.; Overballe-Petersen, S.; Hansen, F.; Hasman, H.; Bortolaia, V.; Hammerum, A.M.; Ingmer, H. Vancomycin resistance in Enterococcus faecium isolated from Danish chicken meat is located on a pVEF4-like plasmid persisting in poultry for 18 years. Int. J. Antimicrob. Agents 2018, 52, 283–286. [Google Scholar] [CrossRef] [Green Version]
- PubMLST. Public Databases for Molecular Typing and Microbial Genome Diversity. Available online: https://pubmlst.org/organisms (accessed on 10 April 2023).
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- León-Sampedro, R.; del Campo, R.; Rodriguez-Baños, M.; Lanza, V.F.; Pozuelo, M.J.; Francés-Cuesta, C.; Tedim, A.P.; Freitas, A.R.; Novais, C.; Peixe, L.; et al. Phylogenomics of Enterococcus faecalis from wild birds: New insights into host-associated differences in core and accessory genomes of the species. Environ. Microbiol. 2019, 21, 3046–3062. [Google Scholar] [CrossRef]
- Kawalec, M.; Pietras, Z.; Daniłowicz, E.; Jakubczak, A.; Gniadkowski, M.; Hryniewicz, W.; Willems, R.J.L. Clonal structure of Enterococcus faecalis isolated from Polish hospitals: Characterization of epidemic clones. J. Clin. Microbiol. 2007, 45, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Stępień-Pyśniak, D.; Hauschild, T.; Dec, M.; Marek, A.; Brzeski, M.; Kosikowska, U. Antimicrobial resistance and genetic diversity of Enterococcus faecalis from yolk sac infections in broiler chicks. Poult. Sci. 2021, 100, 101491. [Google Scholar] [CrossRef]
- Talaga-Ćwiertnia, K.; Bulanda, M. Lekooporność rodzaju enterococcus—Aktualny problem wśród ludzi i zwierząt. Postep. Mikrobiol. 2018, 57, 244–250. [Google Scholar]
- Eputiene, V.S.; Bogdaite, A.; Ruž Auskas, M.; Suž, E. Antibiotic resistance genes and virulence factors in Enterococcus faecium and Enterococcus faecalis from diseased farm animals: Pigs, Cattle and Poultry. Pol. J. Vet. Sci. 2012, 15, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.-Z.; Zaheer, R.; Poulin-Laprade, D.; Scott, A.; Rehman, M.A.; Diarra, M.; Topp, E.; Van Domselaar, G.; Zovoilis, A.; McAllister, T.A. Comparative Genomic Analysis of Enterococci across Sectors of the One Health Continuum. Microorganisms 2023, 11, 727. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Silva, V.; Igrejas, G.; Carvalho, I.; Peixoto, F.; Cardoso, L.; Pereira, J.E.; Del Campo, R.; Poeta, P. Genetic Characterization of vanA-Enterococcus faecium Isolates from Wild Red-Legged Partridges in Portugal. Microb. Drug Resist. 2018, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.V.; Malathum, K.; Murray, B.E. Disruption of an Enterococcus faecium Species-Specific Gene, a Homologue of Acquired Macrolide Resistance Genes of Staphylococci, Is Associated with an Increase in Macrolide Susceptibility. Antimicrob. Agents Chemother. 2001, 45, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Ramos, E.; Cordero, J.; Molina-González, D.; Poeta, P.; Igrejas, G.; Alonso-Calleja, C.; Capita, R. Antimicrobial resistance and virulence genes in enterococci from wild game meat in Spain. Food Microbiol. 2016, 53, 156–164. [Google Scholar] [CrossRef]
- Padmasini, E.; Padmaraj, R.; Ramesh, S.S. High Level Aminoglycoside Resistance and Distribution of Aminoglycoside Resistant Genes among Clinical Isolates of Enterococcus Species in Chennai, India. Sci. World J. 2014, 2014, 329157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denys, A.; Denys, A.; Zajdel, R. Występowanie Genów Zjadliwości Wśród Szczepów Enterococcus Faecalis Izolowanych Od Pacjentów I Ze Środowiska Szpitalnego. Med. Dośw. 2009, 015, 125–132. [Google Scholar]
- Dong, J.; Zhang, L.; He, H.; Jin, Y.; Wang, Y.; Chen, L. Prevalence and conservation of ebp genes in Enterococcus faecalis originated from animals. J. Appl. Microbiol. 2022, 132, 3293–3301. [Google Scholar] [CrossRef]
- Singh, K.V.; Nallapareddy, S.R.; Sillanpää, J.; Murray, B.E. Importance of the Collagen Adhesin Ace in Pathogenesis and Protection against Enterococcus faecalis Experimental Endocarditis. PLoS Pathog. 2010, 6, e1000716. [Google Scholar] [CrossRef]
- Cui, P.; Feng, L.; Zhang, L.; He, J.; An, T.; Fu, X.; Li, C.; Zhao, X.; Zhai, Y.; Li, H.; et al. Antimicrobial Resistance, Virulence Genes, and Biofilm Formation Capacity Among Enterococcus species From Yaks in Aba Tibetan Autonomous Prefecture, China. Front. Microbiol. 2020, 11, 1250. [Google Scholar] [CrossRef] [PubMed]
- Prażmo, E.; Godlewska, R.; Kwaśny, M.; Mielczarek, A. Udział czynników wirulencji Enterococcus faecalis w rozwoju chorób miazgi i tkanek okołowierzchołkowych. Postępy Mikrobiol. 2016, 55, 247–254. [Google Scholar]
- Nallapareddy, S.R.; Weinstock, G.M.; Murray, B.E. Clinical isolates of Enterococcus faecium exhibit strain-specific collagen binding mediated by Acm, a new member of the MSCRAMM family. Mol. Microbiol. 2003, 47, 1733–1747. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.A.; Willems, R.J.L.; Bonten, M.J.M.; van Schaik, W. LPxTG surface proteins of enterococci. Trends Microbiol. 2009, 17, 423–430. [Google Scholar] [CrossRef]
- Brinster, S.; Posteraro, B.; Bierne, H.; Alberti, A.; Makhzami, S.; Sanguinetti, M.; Serror, P. Enterococcal Leucine-Rich Repeat-Containing Protein Involved in Virulence and Host Inflammatory Response. Infect. Immun. 2007, 75, 4463. [Google Scholar] [CrossRef] [Green Version]
- Nunez, N.; Derré-Bobillot, A.; Gaubert, S.; Herry, J.M.; Deschamps, J.; Wei, Y.; Baranek, T.; Si-Tahar, M.; Briandet, R.; Serror, P.; et al. Exploration of the role of the virulence factor ElrA during Enterococcus faecalis cell infection. Sci. Rep. 2018, 8, 1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woods, S.E.; Lieberman, M.T.; Lebreton, F.; Trowel, E.; De La Fuente-Nuñez, C.; Dzink-Fox, J.; Gilmore, M.S.; Fox, J.G. Characterization of Multi-Drug Resistant Enterococcus faecalis Isolated from Cephalic Recording Chambers in Research Macaques (Macaca spp.). PLoS ONE 2017, 12, 169293. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Name | NWT | Minimal Inhibitory Concentration Value (mg/L) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | 0.032 | 0.064 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | >1024 | |
| Ampicillin | 0.0% | 2 | 5 | 3 | ||||||||||||||
| Chloramphenicol | 10.0% | 2 | 7 | 1 | ||||||||||||||
| Ciprofloxacin | 10.0% | 2 | 1 | 4 | 2 | 1 | ||||||||||||
| Daptomycin | 0.0% | 5 | 5 | |||||||||||||||
| Erythromycin | 10.0% | 1 | 5 | 3 | 1 | |||||||||||||
| Gentamicin | 10.0% | 9 | 1 | |||||||||||||||
| Linezolid | 0.0% | 1 | 9 | |||||||||||||||
| Quinupristin/dalfopristin | 60.0% | 1 | 3 | 5 | 1 | |||||||||||||
| Teicoplanin | 0.0% | 10 | ||||||||||||||||
| Tetracycline | 10.0% | 9 | 1 | |||||||||||||||
| Tigecycline | 0.0% | 6 | 2 | 2 | ||||||||||||||
| Vancomycin | 0.0% | 8 | 2 | |||||||||||||||
| Antimicrobial Name | NWT | Minimal Ipnhibitory Concentration Value (mg/L) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | 0.032 | 0.064 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | >1024 | |
| Ampicillin | 0.0% | 6 | 26 | 18 | ||||||||||||||
| Chloramphenicol | 0.0% | 4 | 46 | |||||||||||||||
| Ciprofloxacin | 0.0% | 1 | 4 | 40 | 5 | |||||||||||||
| Daptomycin | 0.0% | 5 | 28 | 16 | 1 | |||||||||||||
| Erythromycin | 0.0% | 25 | 22 | 3 | ||||||||||||||
| Gentamicin | 0.0% | 10 | 37 | 3 | ||||||||||||||
| Linezolid | 0.0% | 4 | 46 | |||||||||||||||
| Quinupristin/dalfopristin | 0.0% | 2 | 37 | 11 | ||||||||||||||
| Teicoplanin | 0.0% | 48 | 2 | |||||||||||||||
| Tetracycline | 0.0% | 44 | 1 | 3 | 2 | |||||||||||||
| Tigecycline | 4.0% | 9 | 23 | 18 | ||||||||||||||
| Vancomycin | 0.0% | 29 | 18 | 3 | ||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwit, R.; Zając, M.; Śmiałowska-Węglińska, A.; Skarżyńska, M.; Bomba, A.; Lalak, A.; Skrzypiec, E.; Wojdat, D.; Koza, W.; Mikos-Wojewoda, E.; et al. Prevalence of Enterococcus spp. and the Whole-Genome Characteristics of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Free-Living Birds in Poland. Pathogens 2023, 12, 836. https://doi.org/10.3390/pathogens12060836
Kwit R, Zając M, Śmiałowska-Węglińska A, Skarżyńska M, Bomba A, Lalak A, Skrzypiec E, Wojdat D, Koza W, Mikos-Wojewoda E, et al. Prevalence of Enterococcus spp. and the Whole-Genome Characteristics of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Free-Living Birds in Poland. Pathogens. 2023; 12(6):836. https://doi.org/10.3390/pathogens12060836
Chicago/Turabian StyleKwit, Renata, Magdalena Zając, Aleksandra Śmiałowska-Węglińska, Magdalena Skarżyńska, Arkadiusz Bomba, Anna Lalak, Ewelina Skrzypiec, Dominika Wojdat, Weronika Koza, Emilia Mikos-Wojewoda, and et al. 2023. "Prevalence of Enterococcus spp. and the Whole-Genome Characteristics of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Free-Living Birds in Poland" Pathogens 12, no. 6: 836. https://doi.org/10.3390/pathogens12060836